Introduction
Breast cancer is one of the most frequent cancers in
females worldwide. Patients negative for the nuclear oestrogen
receptor ER-α (oestrogen receptor α) have a particularly poor
prognosis (1). Therefore, we
focused on neoplastic processes induced in mice by ER-α-negative
tumours. 4T1 cell line was chosen to induce malignancy. This cell
line is considered as a triple-negative model. Apart from ER-α,
this cell line lacks the protein expression of p53 (2), Wnt-5a and PgR (progesterone receptor)
(3).
The 4T1 mammary carcinoma cell line was isolated by
Miller et al (4) growing as
an adherent epithelial type in vitro, and are characterized
as murine mammary carcinoma cells. When introduced orthotopically
into BALB/c mice, 4T1 cells rapidly divide and spontaneously
metastasize from the mammary fat pad to several organs including
liver, lungs, lymph node and brain, as well as into bones (5). Since these tumours closely imitate
advanced human breast cancer, the 4T1 cell line serves as a model
for stage IV of breast cancer (6).
Many epidemiological studies have dealt with a close
link between higher breast tissue zinc levels and the development
of breast cancer (7,8). Compared with the adjacent tissues,
the zinc accumulation in the tumour tissue correlated with the
higher expression of cellular zinc importing proteins, prompting
that tumour cells are profiting from the increase of zinc uptake
(9). Furthermore, a low level of
zinc efflux transporter ZnT1 was found in breast tumour cells,
which caused higher levels of zinc, confirming a deregulation of
zinc transport in proliferating tumour cells (10,11).
The expression of ZIP6, 7 and 10 (zinc-influx transporters) has
been also associated with the breast cancer growth and metastasis
(8,12–14).
Breast cancer seems to be rare in its acquisition of zinc,
indicating a potential zinc demand for the development of breast
malignancy (8).
Proper supervision of cytosolic zinc is highly
needed for maintaining the redox status of the cells, because both
excessive and reduced zinc levels can induce oxidative stress
(15). As the excessive zinc is
toxic, detoxification of this ion has a crucial role in sustaining
cell homeostasis.
Zinc and other heavy metals are known to induce the
expression of metallothioneins (MT) (16–19).
MT expression was shown to be increased during oxidative stress
(20,21) to protect cells against cytotoxicity
(22,23) and DNA damage (24,25).
Metallothionein (MT) was also reported as a possible negative
regulator of apoptosis (26). The
main transcription factor involved in the metal regulation of MT
expression is MRE-binding transcription factor-1 (MTF-1) (27). The free zinc is capable to bind
MTF-1 and translocate it to the nucleus (28). In the nucleus, MTF-1 binds to metal
responsive elements (MRE) in the MT promoter and triggers the MT
expression to recoup for metal toxicity. Nevertheless, some studies
have demonstrated that only breast cancer epithelial cells with the
intact p53 can induce metallothionein (MT) synthesis after metal
exposure (29,30). Accordingly, it was also shown, that
a disruption of the p53 function sensitizes breast cancer MCF-7
cells to cisplatin (31).
Consequently, the aim of the present study was to
evaluate the effect of excessive zinc supplementation on
metallothionein 1 and 2, metal-regulatory transcription factor 1
and TP53 expression in a mouse model with the 4T1-induced advanced
breast cancer. The studied mammary tumour arose from 4T1 cells with
non-functional p53 and zinc transporters set to zinc accumulation
(13,32); thus, the hypothesis of the toxicity
of excessive zinc for the tumour tissue was tested. Furthermore,
the effect of tumour on the antioxidant capacity of tissues was
examined.
Materials and methods
Chemical and biochemical reagents
RPMI-1640 medium, fetal bovine serum (FBS)
(mycoplasma-free), penicillin/streptomycin and trypsin were
purchased from PAA Laboratories GmbH (Pasching, Austria).
Phosphate-buffered saline (PBS) was purchased from Invitrogen Corp.
(Carlsbad, CA, USA). Ethylenediaminetetraacetic acid (EDTA),
Zinc(II) sulphate (BioReagent grade, suitable for cell cultures),
RIPA buffer and all other chemicals of ACS purity were purchased
from Sigma-Aldrich Co. (St. Louis, MO, USA), unless noted
otherwise.
Cell culture and culture conditions
For in vivo and in vitro studies, the
4T1 cell line was used (cell line was kindly provided by Dr Lucia
Knopfova, Masaryk University, Brno, Czech republic). The 4T1 cells
grow as adherent epithelial type in vitro, and are
characterized as murine mammary carcinoma cells [American Type
Culture Collection (ATCC) catalogue no. CRL-2539, 2004]. The cells
were cultured in RPMI-1640 medium (PAA Laboratories) supplemented
with 10% fetal bovine serum (PAA Laboratories), 100 U/ml
penicillin, 100 μg/ml streptomycin, 1 mM sodium pyruvate, and 4.5
g/l glucose and kept in a 37°C humidified incubator with 5%
CO2 in air.
Cell number quantification
Total cell content was analysed using the Casy model
TT system (Roche Applied Science, Penzberg, Germany) and the
following protocol: first, calibration was performed from viable
and necrotic cells. For necrotic cells, 100 μl cell suspension and
800 μl Casy Blue solution was mixed and left for 5 min at room
temperature. Subsequently, 9 ml Casy Tone was added. To prepare a
viable cell standard, 100 μl of cell suspension was mixed with 10
ml of Casy Tone. All subsequent measurements were made in 100×
diluted 100 μl cell suspension. Prior to each measurement,
background was subtracted. All samples were measured in
duplicates.
Zinc(II) treatment of 4T1 cells
Once the cells grew up to 50–60% confluence in the
culture, the growth media were replaced by fresh medium for 24 h to
synchronize the cell growth. The 4T1 cells were then treated with
zinc sulphate (0–100 μM) in fresh medium for 48 h.
MTT viability assay
The MTT assay was used to determine cell viability.
The suspension of cells in the growth medium was diluted to a
density of 5,000 cells/1 ml and 200 ml were transferred to wells
2–11 of standard microtiter plates. The medium (200 ml) was added
to the first and to the last column (1 and 12). The plates were
incubated for 2 days at 37°C to ensure the cell growth. The medium
was removed from columns 2 through to 11. Columns 3–10 were filled
with 200 μl of the medium containing different concentrations of
zinc (0, 25, 50, 75 and 100 μM). As a control, columns 2 and 11
were fed with the medium only. The plates were incubated for 24 h;
then, the medium was removed and exchanged with a fresh medium,
daily three times. After that, columns 1–11 were fed with 200 ml of
the medium with 50 ml of MTT (5 mg/ml in PBS) and incubated for 4 h
in a humidified atmosphere at 37°C, wrapped in the aluminium foil.
After that, the medium-MTT was exchanged with 200 ml of 99.9% DMSO
to dissolve MTT-formazan crystals. Then, 25 ml of glycine buffer
was added to all wells with DMSO and the absorbance was recorded at
570 nm (VersaMax microplate reader; Molecular Devices, Sunnyvale,
CA, USA) (33).
Real-time impedance based cell growth and
proliferation assay
The impedance-based real-time cell analysis (RTCA)
xCELLigence system was used according to the instructions of the
supplier (Roche Applied Science and ACEA Biosciences, San Diego,
CA, USA). The xCELLigence system consists of four main components:
RTCA DP station, RTCA computer with integrated software and
disposable E-Plate 16. Firstly, the optimal seeding concentration
for proliferation and cytotoxic assay was determined. Optimal
response was found in the well containing 10,000 cells. After
seeding a total number of cells in 200 ml of medium to each well in
E-Plate 16, the attachment and proliferation of the cells were
monitored every 15 min. Duration of all experiments was 200 h.
Results are expressed as relative impedance using manufacturer’s
software (Roche Applied Science and ACEA Biosciences) (33).
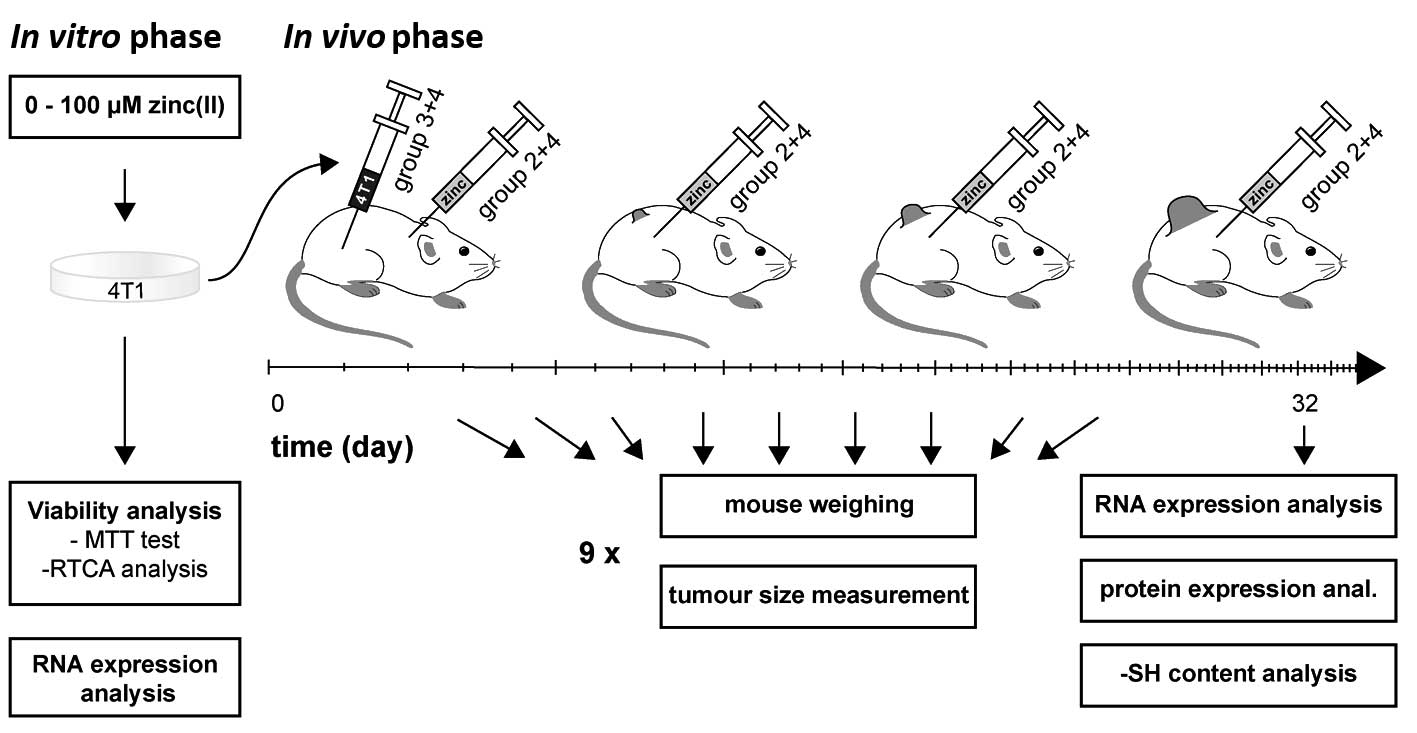
Tumour development and zinc
supplementation
Eight-week-old 40 Balb/c female mice (weight 21–24
g) were equally distributed to 2×2 groups according to the tumour
presence (tumour/control) and zinc supplementation
(supplemented/not supplemented). The 4T1 metastatic breast cancer
cell line was used to induce the neoplastic process. The 4T1 cells
used in the tumour development study were freshly harvested at 70%
confluence. The 4T1 cells (1×105) in 20 μl of PBS and
Matrigel (1:1 ratio) were administered into the fourth mammary fat
pad under general anesthesia (1% Narkamon + 2% Rometar 0.5 ml/100 g
of weight).
The intraperitoneally administered dose of zinc was
a half-LD50 dose as determined by Domingo et al
(34) (total dose 0.15 mg/g of
zinc sulphate). A total amount of 0.15 mg/g of zinc sulphate
divided into 4 doses (0.025 + 0.05 + 0.05 + 0.025 mg/g) was applied
to the mice intraperitoneally. The mice were fed ad libitum,
and observed daily. The cages were kept in a climate-controlled
room and cleaned weekly. Tumour volume (length × width2
× 0.5) was measured twice per week following the equation utilized
in similar tumour development studies (35,36).
After 1 month, the animals were sacrificed, the tumour weight and
volume were measured and the following organs were isolated:
primary tumour, brain, liver, lung, spleen and kidney. RNA was
isolated by using TriPure reagent (Roche, Boston, MA USA) and
qRT-PCR was performed with β-actin as housekeeping gene.
The use of the animals followed the European
Community Guidelines as accepted principles for the use of
experimental animals. The experiments were performed with the
approval of the Ethics Commission at the Faculty of Medicine,
Masaryk University, Brno, Czech Republic.
Tissue sample preparation
Approximately 0.01 g of the tissue in 500 ml of PBS
was mechanically disintegrated using Ultra-Turrax T8 homogenizer
(IKA-Werke, Staufen, Germany) placed in an ice bath for 3 min at
25,000 rpm.
The homogenates were kept at 99°C in a thermomixer
(Eppendorf 5430; Eppendorf, Hamburg, Germany) for 15 min with
shaking. The denatured homogenates were centrifuged at 4°C, 15,000
× g for 30 min (Eppendorf 5402; Eppendorf). Heat treatment
effectively denatures and removes thermolabile and high molecular
mass proteins from the samples. The prepared samples were used for
metallothionein and sulfhydryl group analyses.
Electrochemical detection of Zinc(II)
ions
Electrochemical analyser Model Metrohm AG (Herisau,
Switzerland) was used for the determination of Zn(II). The analyser
(757 VA Computrace) employs a conventional three-electrode
configuration with the hanging mercury drop electrode (HMDE) as a
working electrode: 0.4 mm2, Ag/AgCl/3M KCl as a
reference electrode and a platinum auxiliary electrode.
Differential pulse voltammetric measurements were carried out under
the following conditions: deoxygenating with argon for 60 sec;
deposition potential: −1.3 V; time of deposition: 240 sec; start
potential: −1.3 V; end potential: 0.15 V; pulse amplitude: 0.025 V;
pulse time: 0.04 sec; step potential: 5.035 mV; time of step
potential: 0.3 sec. For more details see Masarik et al
(33).
RNA isolation and reverse
transcription
High pure total-RNA isolation kit (Roche) was used
for isolation. The medium was removed and the samples were twice
washed with 5 ml of ice-cold PBS. The cells were scraped off,
transferred to clean tubes and centrifuged at 20,800 × g for 5 min
at 4°C. After this step, a lysis buffer was added and RNA isolation
was carried out according to manufacturer’s instructions. The
isolated RNA was used for cDNA synthesis. RNA (600 ng) was
transcribed using Transcriptor first strand cDNA synthesis kit
(Roche), which was applied according to manufacturer’s
instructions. The cDNA (20 μl) prepared from the total-RNA was
diluted with RNase-free water to 100 μl and the amount of 5 μl was
directly analysed by using the 7500 RT-PCR system (Applied
Biosystems).
Quantitative real-time polymerase chain
reaction
q-PCR was performed in triplicate using the TaqMan
gene expression assay system with the 7500 RT-PCR system (Applied
Biosystems) and the amplified DNA was analysed by the comparative
Ct method using β-actin as an endogenous control for
metallothionein MT2, MT1, MTF-1 and TP53 gene
expression quantification. The primer and probe sets for β-actin
(assay ID: Mm00607939_s1), MT2 (Mm04207591_g1), MT1
(Mm00496660_g1), TP53 (Mm01731290_g1) and MTF-1
(Mm00485274_m1) were selected from TaqMan gene expression assays
(Life Technologies, Waltham, MA, USA). q-PCR was performed under
the following amplification conditions: total volume of 20 μl,
initial incubation at 50°C/2 min followed by denaturation at
95°C/10 min, then 45 cycles at 95°C/15 sec and at 60°C/1 min.
Electrochemical detection of
metallothionein
Electrochemical detection was used to quantify
metallothionein in the thermolysate samples. The detection was
carried out using Autolab Analyser (Eco Chemie, Utrecht, The
Netherlands) with the classic three-electrode arrangement using
differential pulse voltammetry Brdicka reaction. The analysed
sample was accumulated on the surface of a working electrode, which
was represented by the hanging mercury drop electrode. After the
accumulation, the detection proceeded in a supporting electrolyte
containing cobaltic (cobalt3+) salt in ammonia buffer of
pH 9.6 (37).
Analysis of the total content of
sulfhydryl groups
The concentration of sulfhydryl groups (SH groups)
in the tissues was determined using 0.2 mmol/l 5,5′-dithiobis
(2-nitrobenzoic acid) (DTNB) reported by Ellmann (38) (the intra-assay CV was 3.5% and the
inter-assay CV was 5.5%). Total sulfhydryl group content values
were expressed in μmol/g of proteins.
Statistical analysis
The data were tested for normality and the
log-normal distributed data were recalculated. Factorial ANOVA and
the Bonferroni post-hoc test were used for the statistical
analysis. Pearson correlations were performed to reveal dependence
between the variables. Software Statistica 12 (StatSoft, Inc.,
Tulsa, OK, USA) was employed to perform the statistical analysis.
P-level of <0.05 was considered statistically significant.
Results
In vitro effects on the 4T1 cell
line
Effect of zinc(II) on 4T1
viability
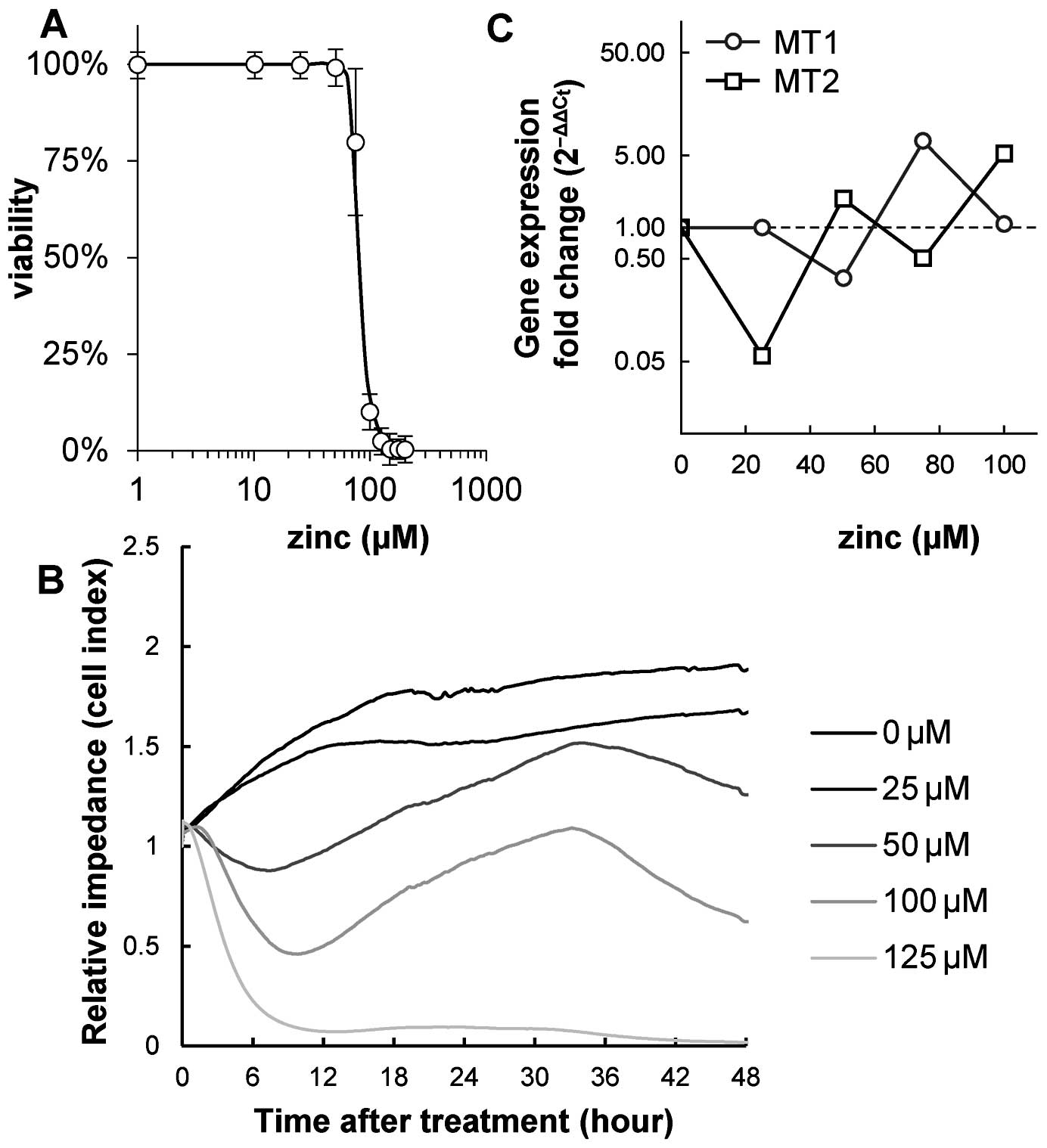
In the first step, an analysis of 4T1 cell line
viability after Zinc(II) treatment was performed to adjust the
optimal dose administered to animals in the next step. For the
proliferation and viability assay, zinc sulphate treatment
concentrations were optimized as follows: 0 (control), 25, 50, 75
and 100 μM. Consequently, IC50 of zinc sulphate for the
4T1 cell line was determined. The IC50 value obtained
from the MTT cytotoxicity assay was 83.7 μM (Fig. 2A). The IC50 value
obtained by using the impedance-based real-time cell analysis
(RTCA) system was also determined for the same period of time as
the MTT assay (48 h after the beginning of the experiment). The
IC50 value was 52.8 μM (Fig. 2B).
Effect of zinc(II) on 4T1 gene
expression
In the next step, the expression of MT1 and
MT2 in the 4T1 cell line after the zinc sulphate treatment
was detected. The following Zinc(II) concentrations were used to
analyse the effect on expression: 0, 25, 50, 75 and 100 μM. Using
one-way ANOVA, no significant effect of the ‘zinc concentration’
factor was determined for MT1 and MT2 expression. No
correlation between MT1 or MT2 expression and zinc
sulphate concentration was found (Fig.
2C).
Mouse models of breast tumours
Effect of zinc(II) on animal
weight
Animals were divided into the following groups: i)
control, ii) control administered with Zinc(II), iii) 4T1-induced
tumour mice, iv) 4T1-induced tumour mice administered with zinc.
The experimental scheme is illustrated in Fig. 1. To verify whether the administered
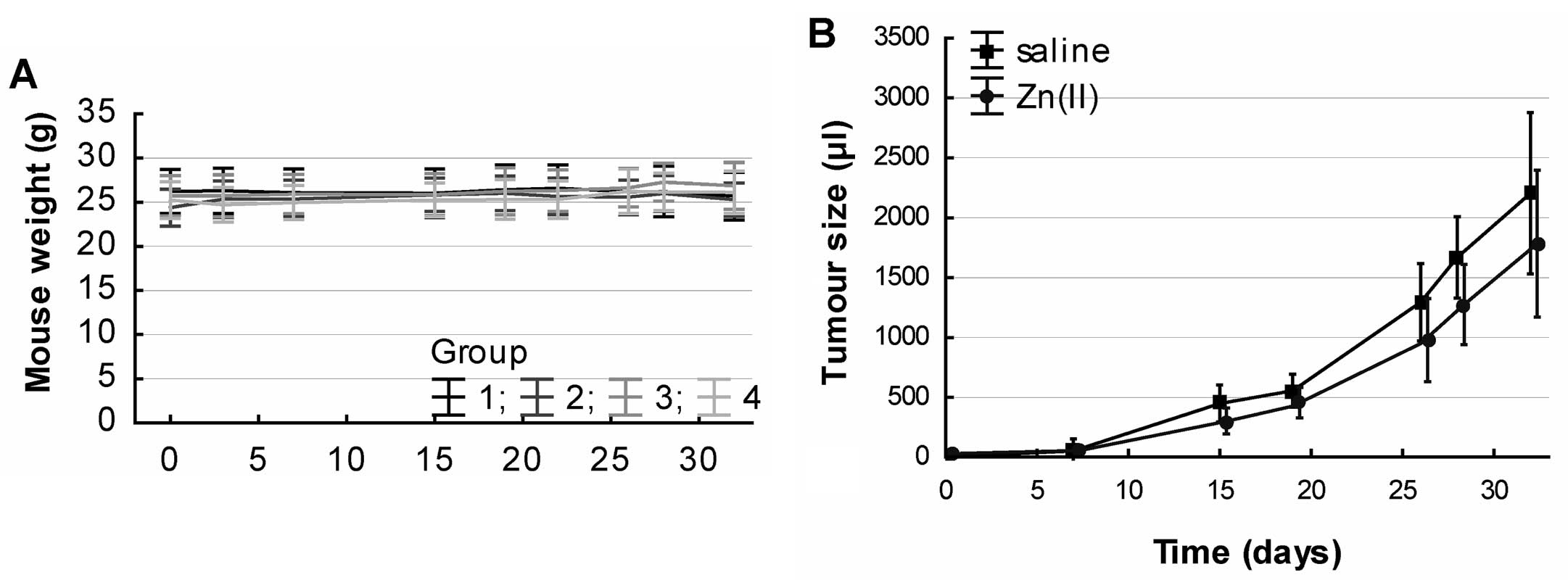
Zinc(II) doses cause symptoms of intoxication, the mice were
weighed during the experiment. There was no significant trend
during the treatment. Additionally, one-way ANOVA did not reveal
any significant weight difference between the groups (Fig. 3A). Based on this finding it can be
concluded, that Zinc(II) does not have any serious whole-body
effects.
Effect of Zinc(II) on tumour size
Tumours arose in all animals with the 4T1 breast
cancer cell application, and no tumour was found in animals without
the 4T1 tumour induction. The autopsy of organs revealed a clear
progression of metastases to liver, lungs, bones and spleen with
occasional metastases to brain and intestines in both groups (zinc
sulphate untreated or treated).
Concurrently, the tumour size was measured during
the experiment to verify whether the Zinc(II) administration
affects the tumour tissue. Using one-way ANOVA after the adjustment
to the treatment time, there was a significant effect of the
treatment group, F(1, 137)=7.2804, P=0.008). Tumour reduction up to
28% was observed in the zinc-treated subjects (Fig. 3B).
Gene expression
The effect of Zinc(II) supplementation was analysed
for association with the gene expression profile of metallothionein
1 and 2 (MT-1 and MT-2), metallothionein
transcription factor MTF-1 and TP53. The effect of
tumour induction, Zinc(II) treatment, and tissue type on gene
expression was analysed.
Using multivariate ANOVA, we observed a significant
effect of the tissue type on the gene expression level, F(20,
481.86)=12.96, P<0.001 after the adjustment of all other
variables. As expected, the highest expression levels of all the
above-mentioned genes were observed in hepatic and kidney tissues.
On the other hand, the expression of all genes was significantly
lower in the lung tissue, spleen and in the primary tumour as
assessed by Bonferroni post-hoc testing.
Using multivariate ANOVA, there was a significant
effect of tumour induction on the gene expression level, F(4,
145)=11.36, P<0.001 after the adjustment of all other variables;
the expression of all of the above-mentioned genes was lower in the
tumourous tissues. Consequently, the effect of treatment was
analysed using the same test after the adjustment of all other
variables. However, the expression of the selected genes did not
differ significantly in relation to this factor, F(4, 145)=1.40,
P=0.24. Therefore, a combined effect of the Zinc(II) treatment,
tumour induction and tissue type was analysed in relation to gene
expression to reveal whether the effect of treatment is recorded
only in the individual types of tissues or only in the
tumour-induced cases.
The ANOVA test revealed, that the expression of
genes is significantly affected by the combined effect of Zinc(II)
treatment*tumour induction*tissue, F(20, 481,86)=1.64, P=0.04
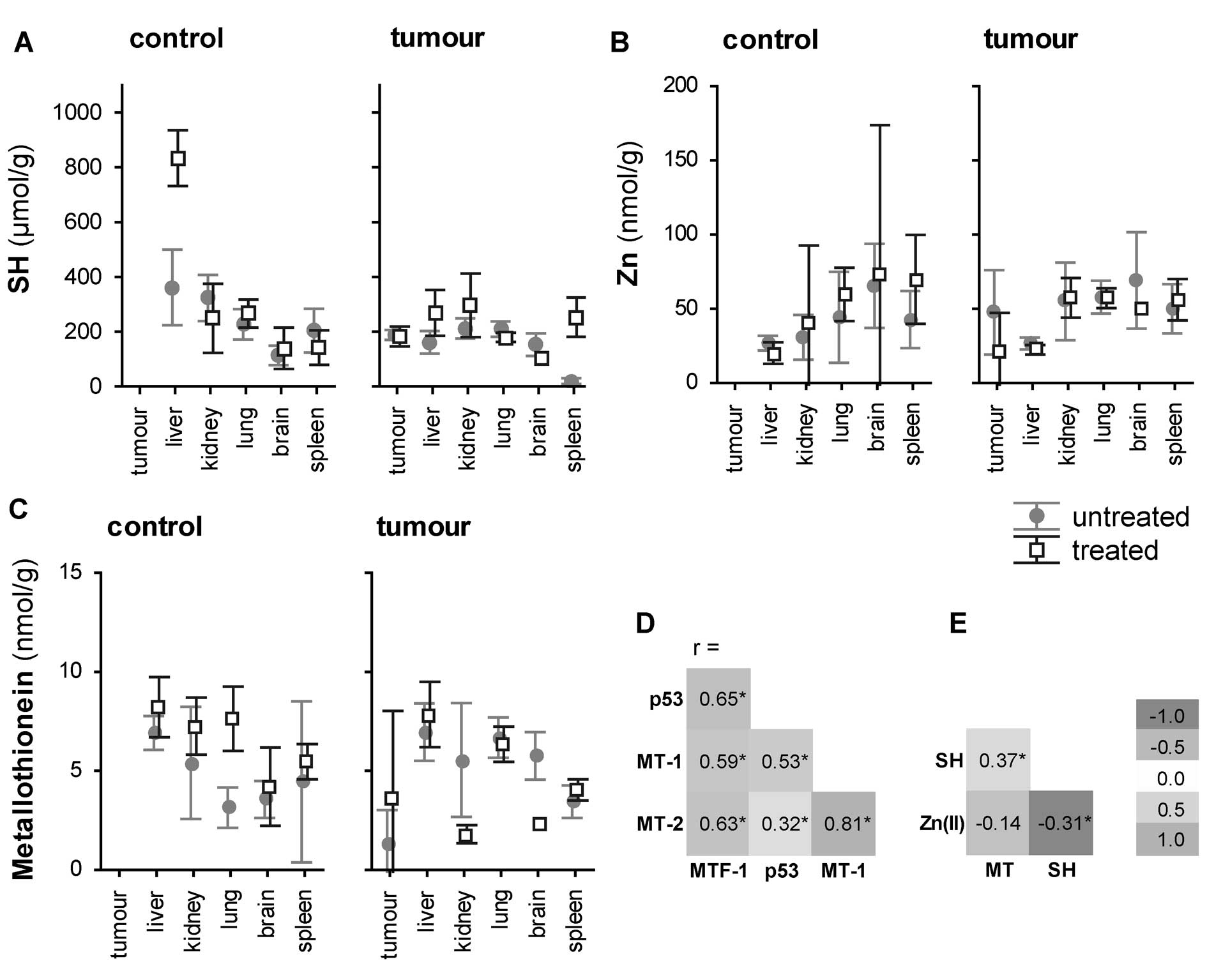
(Fig. 4). To reveal the trends of
individual genes, the Bonferroni post-hoc test was performed using
the ‘tissue-by-tissue’ manner. There was a strong co-expression
pattern of the genes with significant positive correlations; all
correlations were significant at P<0.01 with the highest
correlation observed between MT1 and MT2 (r=0.82) and
the lowest (but still significant) correlation observed between
TP53 and MT2 (r=0.32) (Fig. 5D). With regard to the
metallothionein expression, significant differences were recorded
in the lung tissue; animals with induced tumours had significantly
lower MT1 and MT2 levels than the controls. While no
other ‘intra-tissue’ trends were observed in the MT1
isoform, we recorded significant trends of MT2 levels in the
spleen and liver tissue. Similarly as in the lungs, the MT2
expression was significantly lower in the tumourous tissue than in
the control animals. By contrast, an inverse trend was apparent in
the liver tissue; the expression of MT2 in untreated
controls was significantly lower than in treated mice with the
induced tumours.
In summary, the expression levels of MT, TP53
and MTF-1 are affected mostly by the tissue type. Tumour
induction and Zinc(II) treatment affect the expression of
individual genes only, namely MT1 and MT2.
Analysis of sulfhydryl group compounds,
MT protein expression and zinc(II) ions
Experiments analysing the RNA levels pointed to a
significant variation of MT in relation to tumour induction. MTs
have a high content of cysteine residues that bind various heavy
metals, therefore an analysis of sulfhydryl group content was
carried out. The total-SH content does not reflect only MT, but
also other redox-active compounds, reduced glutathione (GSH) in
particular. Thus, MT was determined electrochemically. In addition,
Zinc(II) levels were detected.
A correlation between these variables was
determined. While only a weak positive correlation was found
between the content of -SH groups and MT (r=0.37 at P<0.001),
Zinc(II) correlated with the -SH group content negatively, r=−0.31
at P<0.001 (Fig. 5E). Thus, -SH
and metallothionein were interpreted separately in the following
statistical tests.
Similarly to the analysis of gene expression, the
effect of tissue on the concentration of sulfhydryl groups (-SH),
metallothionein protein (MT) and Zinc(II) was assessed using ANOVA.
Similar to the mRNA level, the concentrations of these substances
differed between the tissues. While there were significantly lower
concentrations of MT in the primary tumour as compared to brain,
lung and liver, an inverse trend was observed in the concentration
of Zinc(II): the concentration of Zinc(II) was significantly higher
in the tumorous tissue compared to that recorded in the liver.
Consequently, the effect of tumour induction of the
level of MT, -SH and zinc was assessed. A significantly 1–5-fold
lower -SH content was identified in animals with the induced
tumours using post-hoc testing. MT and zinc did not differ
significantly.
There was a significant effect of the treatment on
the metallothionein protein level, -SH groups and zinc level, F(3,
113)=4.90, P=0.003 (Fig. 5A–C).
Post-hoc testing revealed a significantly 1.4-fold higher content
of -SH groups in treated animals at P<0.05. This trend was
highlighted namely when the combined effect of treatment*tissue
type*tumour induction on the -SH concentration was analysed
(Fig. 5A). On the other hand, the
metallothionein protein and zinc levels were also elevated in the
treated animals, but below the level of statistical
significance.
Discussion
The aim of the present study was to assess if zinc
ions provided in excess may significantly modify the process of
mammary oncogenesis in mice. Zn(II) ions were chosen because of
their documented antitumour effect and inhibition of copper ions,
which are known as pro-oxidant and angiogenic factors (32,39–41).
Previously, it was also demonstrated in breast cancer cells that
zinc ions can inhibit NF-κB, which is exceedingly activated in
breast cancer with a hormone independent phenotype (42,43).
Moreover, zinc may have a direct impact on cell death in prostate
cancer cells (44). Even
physiological concentrations of zinc cause quick discharge of
cytochrome c from the inner membrane of mitochondria.
Thereafter, cytochrome c triggers the caspase pathway and
activates apoptosis (45). Zinc is
also involved in epigenetic mechanisms (46,47).
From the epigenetic perspective, zinc deficiency leads to decreased
DNA and histone methylation (48),
which may be due to the fact that histone deacetylases and histone
lysine methyltransferases are zinc-dependent enzymes (49–52).
Inhibitors of histone deacetylase were also shown to reactivate the
oestrogen receptor in oestrogen receptor-negative breast cancer
cells (53). Other effects of zinc
ions were expected through the oxidative stress and
metallothionein-mediated pathways.
In the present study, the intraperitoneally
administered dose of zinc was a half-LD50 dose as
determined by Domingo et al (34) (total dose 0.15 mg/g of zinc
sulphate). Using different dietary zinc doses at different time
periods, Hou et al (40)
found that doses <1 mg/g do not result in anorexia and body
weight loss in mice. Accordingly, zinc doses administrated in our
study caused no symptoms of intoxication, and there was no body
weight loss in the treated mice as compared with the control group.
The studied mammary tumour arose from 4T1 cells with non-functional
p53 and zinc transporters set to zinc accumulation (13), thus, the hypothesis about the
toxicity of excessive zinc for the tumour tissue was tested. Fan
and Cherian (29) and
Ostrakhovitch et al (30)
demonstrated that only breast cancer epithelial cells with the
intact p53 can induce metallothionein (MT) synthesis after the
metal exposure. In agreement with these findings, no significant
changes in MT1 or MT2 mRNA expression after the zinc
sulphate treatment of the 4T1 cell culture were found. No
correlation between MT1 or MT2 expression and zinc
sulphate concentration was established. Neither was a significant
change in MT1, MT2, TP53 and MTF1 gene
expression found in the tumour tissue after Zinc(II) exposure. The
tumour-free mice had significantly higher expression levels of the
studied genes (P<0.003).
Significant differences were also revealed among the
tissues (P<0.001). The highest expression levels were observed
in liver, which indicates the important role of this organ in the
oxidative stress processes and in the homeostasis of metal ions. In
the tumour-free mice, the amount of hepatic sulfhydryl groups was
significantly increased by the exposure to zinc sulphate, but
animals with tumour induction did not show a similar trend. In
contrast, no such trend was observed in metallothionein (compare
Fig. 5A and C). The difference as
well as the relatively weak correlation between MT and -SH can be
elucidated as follows: GSH is an important non-protein -SH
compound, together with MT, which includes an unusually high
cysteine content, these two -SH substances represent important
defence against stress (54). This
is why the total -SH content reflects mostly those compounds. Thus,
the zinc-induced -SH elevation reflects an increase of GSH content
in liver and emphasizes the importance of liver tissue in the
regulation of zinc(II).
The above confirms the significant effect of breast
tumour on the antioxidant capacity of the whole organism and shows
the inability of the exhausted body to cope with oxidative stress
conditions. According to Feng et al (55) in patients with malignant breast
cancer the overall antioxidant/oxidant balance is significantly
disrupted as compared with healthy controls. Kasapovic et al
(56) also showed that breast
carcinoma is associated with elevated plasma lipid peroxidation and
with a concomitant decrease in the antioxidant capacity of blood
cells.
In conformity with our hypothesis, the primary
tumour size of zinc-treated animals was 23% smaller F(1, 114)=9.71,
P=0.02 using ANOVA after adjustment for time; however, no
significant effect on metastasis progression due to the zinc
treatment was discovered. As a further confirmation of excessive
zinc toxicity for breast cancer cells, Al-Qubaisi et al
(57) found that the
IC50 value of NiZn ferrite nanoparticles in normal
breast MCF-10 cells was almost 15 times higher than that in breast
cancer MCF-7 cells lines. Surprisingly, no significant changes in
the final zinc accumulation between tissues or due to the zinc
treatment or tumour bearing were found. Nevertheless, Woo and Wu
(32) reported that MNU-induced
mammary tumourigenesis resulted in a zinc concentration in the
mammary tumour higher than in the mammary gland, whereas the body
zinc-distribution profile was not affected. Skrajnowska et
al (58) demonstrated that rat
diet supplemented with zinc or copper increased the Cu content in
the cancerous tissue as compared to normal tissue but did not lead
to changes in the final Zn levels. We hypothesize that the tumour
cells with the highest zinc accumulation succumbed to cell death,
which is why the final zinc levels remained unchanged.
In contrast to our study, some other studies have
shown that the growth of mammary carcinomas is suppressed rather by
zinc depletion (59–61). McQuity et al (62) discovered that mammary tumour growth
was markedly inhibited in the zinc-deficient group and only
slightly inhibited in the zinc-adequate and zinc-excess pair-fed
groups of rats. This difference might be due to the different cell
lines used in the studies. Pories et al (60), Dewys et al (61) and McQuity et al (62) used Walker 256 cells. Mills et
al (59) used R323OAC mammary
adenocarcinoma cells. Walker 256 cells arose spontaneously in the
mammary gland of a pregnant albino rat and were shown to be of
hematopoietic (i.e., monocytoid) origin (63). The R3230AC rat mammary
adenocarcinoma is a transplantable tumour model, which can be grown
in vitro, but unlike the 4T1 cells, it does not metastasize
spontaneously (64). It seems
there are no significant differences between the normal rat mammary
tissue and the oestrogen receptor-deficient R323OAC tumour with
regard to the number of prolactin binding sites, specificity of
binding, or the affinity of the receptor for prolactin (65). In contrast, 4T1 cells represent a
typical triple-negative breast cancer cell line (ER−,
PR−, HER2−) (66), they are highly metastatic, display
predominantly epithelial characteristics, though they also express
the mesenchymal marker VIM (67).
Similar to human breast cancer, 4T1 metastatic disease spreads from
the primary tumour. Also, the progressive dissemination of 4T1
metastases to lymph nodes and other organs is analogous to human
breast cancer (68). This evidence
indicates that 4T1 should be a more favourable breast cancer model
for the assessment of zinc treatment. According to the present
study and to the results of other authors (8,69,70),
zinc itself may mute the growth of primary breast tumours
especially at the early stages.
Acknowledgements
The financial support from doc CEITEC 02/2012 (JG),
MUNI/A/1003/2013, European Regional Development Fund - project
FNUSA-ICRC (CZ.1.05/1.1.00/02.0123) and Project for Conceptual
Development of Research Organization (Faculty of Medicine, Masaryk
University - ROZV/24/LF5/2014) is greatly acknowledged.
References
|
1
|
Giacinti L, Claudio PP, Lopez M and
Giordano A: Epigenetic information and estrogen receptor alpha
expression in breast cancer. Oncologist. 11:1–8. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yerlikaya A, Okur E and Ulukaya E: The
p53-independent induction of apoptosis in breast cancer cells in
response to proteasome inhibitor bortezomib. Tumor Biol.
33:1385–1392. 2012. View Article : Google Scholar
|
|
3
|
Ford CE, Ekstrom EJ and Andersson T:
Wnt-5a signaling restores tamoxifen sensitivity in estrogen
receptor-negative breast cancer cells (Retracted Article. See vol
107, pg 22360, 2010). Proc Natl Acad Sci USA. 106:3919–3924. 2009.
View Article : Google Scholar
|
|
4
|
Miller FR, Miller BE and Heppner GH:
Characterization of metastatic heterogeneity among subpopulations
of a single-mouse mammary-tumor: heterogeneity in phenotypic
stability. Invasion Metastasis. 3:22–31. 1983.
|
|
5
|
Tao K, Fang M, Alroy J and Sahagian GG:
Imagable 4T1 model for the study of late stage breast cancer. BMC
Cancer. 8:2282008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aslakson CJ and Miller FR: Selective
events in the metastatic process defined by analysis of the
sequential dissemination of subpopulations of a mouse
mammary-tumor. Cancer Res. 52:1399–1405. 1992.PubMed/NCBI
|
|
7
|
Cui Y, Vogt S, Olson N, Glass AG and Rohan
TE: Levels of zinc, selenium, calcium, and iron in benign breast
tissue and risk of subsequent breast cancer. Cancer Epidemiol
Biomark Prev. 16:1682–1685. 2007. View Article : Google Scholar
|
|
8
|
Grattan BJ and Freake HC: Zinc and cancer:
implications for LIV-1 in breast cancer. Nutrients. 4:648–675.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chasapis CT, Loutsidou AC, Spiliopoulou CA
and Stefanidou ME: Zinc and human health: an update. Arch Toxicol.
86:521–534. 2012. View Article : Google Scholar
|
|
10
|
Liuzzi JP and Cousins RJ: Mammalian zinc
transporters. Annu Rev Nutr. 24:151–172. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee R, Woo W, Wu B, Kummer A, Duminy H and
Xu Z: Zinc accumulation in N-methyl-N-nitrosourea-induced rat
mammary tumors is accompanied by an altered expression of ZnT-1 and
metallothionein. Exp Biol Med. 228:689–696. 2003.
|
|
12
|
Taylor KM, Morgan HE, Johnson A, Hadley LJ
and Nicholson RI: Structure-function analysis of LIV-1, the breast
cancer-associated protein that belongs to a new subfamily of zinc
transporters. Biochem J. 375:51–59. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kagara N, Tanaka N, Noguchi S and Hirano
T: Zinc and its transporter ZIP10 are involved in invasive behavior
of breast cancer cells. Cancer Sci. 98:692–697. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taylor KM, Vichova P, Jordan N, Hiscox S,
Hendley R and Nicholson RI: ZIP7-mediated intracellular zinc
transport contributes to aberrant growth factor signaling in
antihormone-resistant breast cancer cells. Endocrinology.
149:4912–4920. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maret W: Zinc coordination environments in
proteins as redox sensors and signal transducers. Antioxid Redox
Signal. 8:1419–1441. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gumulec J, Masarik M, Krizkova S, et al:
Insight to physiology and pathology of zinc(II) ions and their
actions in breast and prostate carcinoma. Curr Med Chem.
18:5041–5051. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gumulec J, Masarik M, Adam V, Eckschlager
T, Provaznik I and Kizek R: Serum and tissue zinc in epithelial
malignancies: a meta-analysis. PLoS One. 9:e997902014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gumulec J, Raudenska M, Adam V, Kizek R
and Masarik M: Metallothionein - immunohistochemical cancer
biomarker: a meta-analysis. PLoS One. 9:e853462014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holubova M, Axmanova M, Gumulec J, et al:
KRAS NF-kappa B is involved in the development of zinc resistance
and reduced curability in prostate cancer. Metallomics.
6:1240–1253. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ruttkay-Nedecky B, Nejdl L, Gumulec J, et
al: The role of metallothionein in oxidative stress. Int J Mol Sci.
14:6044–6066. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sato M and Bremner I: Oxygen free-radicals
and metallothionein. Free Radic Biol Med. 14:325–337. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Masarik M, Gumulec J, Hlavna M, et al:
Analysis of metallothionein and glutathione in prostate cells as
markers of oxidative stress. Int J Mol Med. 30:S46. 2012.
|
|
23
|
Aschner M, Conklin DR, Yao CP, Allen JW
and Tan KH: Induction of astrocyte metallothioneins (MTs) by zinc
confers resistance against the acute cytotoxic effects of
methylmercury on cell swelling, Na+ uptake, and
K+ release. Brain Res. 813:254–261. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cai L and Cherian MG: Zinc-metallothionein
protects from DNA damage induced by radiation better than
glutathione and copper- or cadmium-metallothioneins. Toxicol Lett.
136:193–198. 2003. View Article : Google Scholar
|
|
25
|
Min KS, Nishida K and Onosaka S:
Protective effect of metallothionein to ras DNA damage induced by
hydrogen peroxide and ferric ion-nitrilotriacetic acid. Chem Biol
Interact. 122:137–152. 1999. View Article : Google Scholar
|
|
26
|
Takahashi S: Molecular functions of
metallothionein and its role in hematological malignancies. J
Hematol Oncol. 5:412012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Otsuka F, Iwamatsu A, Suzuki K, Ohsawa M,
Hamer DH and Koizumi S: Purification and characterization of a
protein that binds to metal-responsive elements of the human
metallothionein II(a) gene. J Biol Chem. 269:23700–23707.
1994.PubMed/NCBI
|
|
28
|
Smirnova IV, Bittel DC, Ravindra R, Jiang
HM and Andrews GK: Zinc and cadmium can promote rapid nuclear
translocation of metal response element-binding transcription
factor-1. J Biol Chem. 275:9377–9384. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan LZ and Cherian MG: Potential role of
p53 on metallothionein induction in human epithelial breast cancer
cells. Br J Cancer. 87:1019–1026. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ostrakhovitch EA, Olsson PE, von Hofsten J
and Cherian MG: P53 mediated regulation of metallothionen
transcription in breast cancer cells. J Cell Biochem.
102:1571–1583. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan S, Smith ML, Rivet DJ, et al:
Disruption of p53 function sensitizes breast-cancer MCF-7 cells to
cisplatin and pentoxifylline. Cancer Res. 55:1649–1654.
1995.PubMed/NCBI
|
|
32
|
Woo W and Xu ZM: Body zinc distribution
profile during N-methyl-N-nitrosourea-induced mammary tumorigenesis
in rats at various levels of dietary zinc intake. Biol Trace Elem
Res. 87:157–169. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Masarik M, Gumulec J, Hlavna M, et al:
Monitoring of the prostate tumour cells redox state and real-time
proliferation by novel biophysical techniques and fluorescent
staining. Integr Biol. 4:672–684. 2012. View Article : Google Scholar
|
|
34
|
Domingo JL, Llobet JM, Paternain JL and
Corbella J: Acute zinc intoxication - comparison of the antidotal
efficacy of several chelating-agents. Vet Hum Toxicol. 30:224–228.
1988.PubMed/NCBI
|
|
35
|
Kotoh T, Dhar DK, Masunaga R, et al:
Antiangiogenic therapy of human esophageal cancers with thalidomide
in nude mice. Surgery. 125:536–544. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xanthopoulos JM, Romano AE and Majumdar
SK: Response of mouse breast cancer cells to anastrozole,
tamoxifen, and the combination. J Biomed Biotechnol. 2005:10–19.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kizek R, Trnkova L and Palecek E:
Determination of metallothionein at the femtomole level by constant
current stripping chronopotentiometry. Anal Chem. 73:4801–4807.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ellman GL: Tissue sulfhydryl groups. Arch
Biochem Biophys. 82:70–77. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Franklin RB and Costello LC: Zinc as an
anti-tumor agent in prostate cancer and in other cancers. Arch
Biochem Biophys. 463:211–217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hou G, Dick R, Zeng C and Brewer GJ:
Comparison of lowering copper levels with tetrathiomolybdate and
zinc on mouse tumor and doxorubicin models. Transl Res.
148:309–314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Silva MP, Silva DM, Conceicao ALC,
Ribeiro-Silva A and Poletti ME: Role of Ca, Fe, Cu and Zn in breast
cancer: study by X-ray fluorescence techniques and
immunohistochemical analysis. X-Ray Spectrom. 42:303–311. 2013.
View Article : Google Scholar
|
|
42
|
Nakshatri H, BhatNakshatri P, Martin DA,
Goulet RJ and Sledge GW: Constitutive activation of NF-kappa B
during progression of breast cancer to hormone-independent growth.
Mol Cell Biol. 17:3629–3639. 1997.PubMed/NCBI
|
|
43
|
Uzzo RG, Leavis P, Hatch W, et al: Zinc
inhibits nuclear factor-kappa B activation and sensitizes prostate
cancer cells to cytotoxic agents. Clin Cancer Res. 8:3579–3583.
2002.PubMed/NCBI
|
|
44
|
Franklin RB and Costello LC: The important
role of the apoptotic effects of zinc in the development of
cancers. J Cell Biochem. 106:750–757. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feng P, Li TL, Guan ZX, Franklin RB and
Costello LC: Direct effect of zinc on mitochondrial apoptogenesis
in prostate cells. Prostate. 52:311–318. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jones PA and Laird PW: Cancer epigenetics
comes of age. Nat Genet. 21:163–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ushijima T and Okochi-Takada E: Aberrant
methylations in cancer cells: where do they come from? Cancer Sci.
96:206–211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wallwork JC and Duerre JA: Effect of
zinc-deficiency on methionine metabolism, methylation reactions and
protein-synthesis in isolated perfused rat-liver. J Nutr.
115:252–262. 1985.PubMed/NCBI
|
|
49
|
Marks PA and Xu WS: Histone deacetylase
inhibitors: potential in cancer therapy. J Cell Biochem.
107:600–608. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Finnin MS, Donigian JR and Pavletich NP:
Structure of the histone deacetylase SIRT2. Nat Struct Biol.
8:621–625. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
51
|
Somoza JR, Skene RJ, Katz BA, et al:
Structural snapshots of human HDAC8 provide insights into the class
I histone deacetylases. Structure. 12:1325–1334. 2004. View Article : Google Scholar
|
|
52
|
Fatemi M, Hermann A, Pradhan S and Jeltsch
A: The activity of the murine DNA methyltransferase Dnmt1 is
controlled by interaction of the catalytic domain with the
N-terminal part of the enzyme leading to an allosteric activation
of the enzyme after binding to methylated DNA. J Mol Biol.
309:1189–1199. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhou Q, Atadja P and Davidson NE: Histone
deacetylase inhibitor LBH589 reactivates silenced estrogen receptor
alpha (ER) gene expression without loss of DNA hypermethylation.
Cancer Biol Ther. 6:64–69. 2007. View Article : Google Scholar
|
|
54
|
Shimizu M, Hochadel JF, Fulmer BA and
Waalkes MP: Effect of glutathione depletion and metallothionein
gene expression on arsenic-induced cytotoxicity and c-myc
expression in vitro. Toxicol Sci. 45:204–211. 1998.PubMed/NCBI
|
|
55
|
Feng JF, Lu L, Zeng P, et al: Serum total
oxidant/antioxidant status and trace element levels in breast
cancer patients. Int J Clin Oncol. 17:575–583. 2012. View Article : Google Scholar
|
|
56
|
Kasapovic J, Pejic S, Todorovic A,
Stojiljkovic V and Pajovic SB: Antioxidant status and lipid
peroxidation in the blood of breast cancer patients of different
ages. Cell Biochem Funct. 26:723–730. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Al-Qubaisi MS, Rasedee A, Flaifel MH, et
al: Cytotoxicity of nickel zinc ferrite nanoparticles on cancer
cells of epithelial origin. Int J Nanomed. 8:2497–2508. 2013.
View Article : Google Scholar
|
|
58
|
Skrajnowska D, Bobrowska B, Tokarz A,
Kuras M, Rybicki P and Wachowicz M: The Effect of Zinc- and copper
sulphate supplementation on tumor and hair concentrations of trace
elements (Zn, Cu, Fe, Ca, Mg, P) in rats with DMBA-induced breast
bancer. Pol J Environ Stud. 20:1585–1592. 2011.
|
|
59
|
Mills BJ, Broghamer WL, Higgins PJ and
Lindeman RD: Inhibition of tumor-growth by zinc depletion of rats.
J Nutr. 114:746–752. 1984.PubMed/NCBI
|
|
60
|
Pories WJ, DeWys WD, Flynn A, Mansour EG
and Strain WH: Implications of the inhibition of animal tumors by
dietary zinc deficiency. Adv Exp Med Biol. 91:243–257. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Dewys W, Pories WJ, Richter MC and Strain
WH: Inhibition of Walker 256 carcinosarcoma growth by dietary zinc
deficiency. Proc Soc Exp Biol Med. 135:17–22. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
McQuitty JT, Dewys WD, Monaco L, et al:
Inhibition of tumor growth by dietary zinc deficiency. Cancer Res.
30:1387–1390. 1970.PubMed/NCBI
|
|
63
|
Tang DG and Honn KV: Apoptosis of W256
carcinosarcoma cells of the monocytoid origin induced by NDGA
involves lipid peroxidation and depletion of GSH: role of
12-lipoxygenase in regulating tumor cell survival. J Cell Physiol.
172:155–170. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Gardner HA, Kellen JA, Wong AHC, Szalai JP
and Katic M: Characterization of different cell subpopulations
derived from an experimental tumor-model. Cancer Invest. 6:161–165.
1988. View Article : Google Scholar
|
|
65
|
Costlow ME, Buschow RA and McGuire WL:
Prolactin receptors in an estrogen receptor-deficient
mammary-carcinoma. Science. 184:85–86. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kau P, Nagaraja GM, Zheng H, et al: A
mouse model for triple-negative breast cancer tumor-initiating
cells (TNBC-TICs) exhibits similar aggressive phenotype to the
human disease. BMC Cancer. 12:1202012. View Article : Google Scholar
|
|
67
|
Drasin DJ, Robin TP and Ford HL: Breast
cancer epithelial-to-mesenchymal transition: examining the
functional consequences of plasticity. Breast Cancer Res.
13:2262011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Pulaski BA and Ostrand-Rosenberg S: Mouse
4T1 breast tumor model. Curr Protoc Immunol. Chapter 20(Unit 20):
22001.
|
|
69
|
Ames BN: Micronutrients prevent cancer and
delay aging. Toxicol Lett. 103:5–18. 1998. View Article : Google Scholar
|
|
70
|
Ho E: Zinc deficiency, DNA damage and
cancer risk. J Nutr Biochem. 15:572–578. 2004. View Article : Google Scholar : PubMed/NCBI
|