Introduction
Ovarian cancer is a malignancy that is associated
with a high mortality. Worldwide, this form of cancer represents
~3% of all cancers in women and is the fifth leading cause of death
in the female population (1).
Despite developments in cancer therapy in the past two decades, the
mortality from ovarian cancer has remained constant (1). Early diagnosis and improved
prognostic markers are required to improve patient survival rates
for ovarian cancer (2). The
identification of new molecular markers may also lead to more
effective and targeted therapies for ovarian cancer.
MicroRNAs (miRNAs) are a class of small, non-coding,
endogenous RNAs that are ~22 nucleotides in length and play
important roles in post-transcriptional regulation (3). miRNAs can inhibit target gene
expression by binding to the 3′-untranslated region (3′-UTR) of
target mRNA, which results in either mRNA degradation or prevention
of translation to protein expression (4,5).
miRNAs have several roles in normal cellular processes involving
cell proliferation, apoptosis, cell differentiation and stress
response (6–8). In recent years, miRNAs have received
attention in cancer research, as 98 miRNAs are located at genomic
regions involved in malignancy, indicating that miRNAs could be
used as diagnostic and prognostic biomarkers (9). However, miRNAs have been reported to
have both tumor suppressor and oncogenic activities (10).
In particular, miR-221 has been reported to have
both tumor suppressor and oncogenic roles in different types of
human cancer, including breast cancer (11,12)
and hepatocellular carcinoma (13). Recently, miR-221 has been shown to
have as tumor suppressor role in non-small cell lung cancer
(14). miRNA-221 is encoded by a
gene located on chromosome Xp11.3 and acts as an oncogene in tumors
of epithelial origin and as an oncosuppressor in hemopoietic
malignancies (12–14). The increasing awareness of the role
of miRNAs in human malignancy has raised the possibility of their
future role as diagnostic and prognostic biomarkers (15,16).
Recent studies have also indicated that miRNAs may be potential
therapeutic targets in human malignancy (17,18).
In 2007, a study in human prostate cancer cell lines
demonstrated that miR-221 mediated its effects on the control of
cell proliferation by targeting p27Kip1 (20). It is now known that miR-221 exerts
its oncogenic effects through the inhibition of the
cyclin-dependent kinase (CDK) inhibitors p27Kip1 and
p57, upregulating ZEB2, an epithelial-to-mesenchymal transition
(EMT)-inducing gene through TRPS1 (19,20).
Overexpression of miR-221 in malignant glioblastoma cells has been
shown to promote the cell cycle through G1 into S phase and to
promote cell apoptosis (21). In
addition, miR-221 has been shown to be downregulated in Kaposi's
sarcoma-associated herpes virus-associated cancers, including
Kaposi sarcoma and primary effusion lymphoma (PELs) (22). A recent study has shown that the
overexpression of miR-221 enhances chemosensitivity to gemcitabine
in cholangiocarcinoma cells (23).
These data demonstrate that miR-221 has a bimodal function in the
tumorigenesis of human cancers.
Apoptotic protease activating factor 1 (APAF1) is a
crucial component of the apoptosome that is present in humans and
mice (24,25) and is assembled in response to
cellular stress, including DNA damage, hypoxia and oncogene
activation (26–28). APAF1 is closely related to several
oncogenes and tumor suppressor genes, including B-cell lymphoma-2
(Bcl-2) and p53 (29,30). A series of studies have shown that
APAF1 protein is an important apoptosis factor that is abnormally
expressed in cancer tissues (27,31).
APAF1 is downregulated in human colorectal cancer, and its
expression is associated with adverse patient prognosis (32,33).
Since the role of miR-221 in the development and
progression of ovarian cancer remains unknown, the present study
was done using both established tumor cell lines and tumor tissue
samples from patients with histopathologically confirmed primary
ovarian carcinoma.
Materials and methods
Patients studied and tissue
collection
Sixty-three patients with histopathologically
diagnosed ovarian cancer had tumor tissue sampled as part of their
routine diagnosis with adjacent normal tissue samples. All patients
underwent surgery at the First Affiliated Hospital of Sun Yat-sen
University between 2008 and 2010. The diagnosis of primary ovarian
cancer was based on histopathological evaluation according to the
International Federation of Obstetrics and Gynecology (FIGO)
criteria. Clinical pathology information was available for all
patient samples, as shown in Table
I. No other treatments were conducted in these patients before
surgery and tissue sampling. Informed patient consents were
obtained from every study participant. The present study was
approved by the Research Ethics Committee of the First Affiliated
Hospital of Sun Yat-sen University. All tissues were collected and
immediately frozen in liquid nitrogen and stored at −80°C for
further study.
 | Table IClinicopathological characteristics
and GAS5 expression in 63 patient samples of ovarian cancer. |
Table I
Clinicopathological characteristics
and GAS5 expression in 63 patient samples of ovarian cancer.
| Clinical
parameters | No. of cases
(%) |
|---|
| Age (years) | |
| <55 | 29 (46.03) |
| >55 | 34 (53.96) |
| Size (cm) | |
| >5 | 33 (52.38) |
| <5 | 30 (47.62) |
| Histologic
differentiation | |
| Well | 5 (7.94) |
| Moderate | 23 (36.51) |
| Poor | 28 (44.44) |
|
Undifferentiated | 7
(11.11) |
| Invasion depth | |
| T1 | 15 (23.81) |
| T2 | 16 (25.40) |
| T3 | 18 (28.57) |
| T4 | 14 (22.22) |
| FIGO stages | |
| I | 8
(12.70) |
| II | 22 (34.92) |
| III | 27 (42.86) |
| IV | 6 (9.52) |
| Lymphatic
metastasis | |
| Yes | 30 (47.62) |
| No | 33 (52.38) |
| Distant
metastasis | |
| Yes | 8
(12.70) |
| No | 55 (87.30) |
| Expression of
miR-221 | |
| Low
expression | 31 (50.79) |
| High
expression | 32 (49.21) |
Cell culture
Human ovarian surface epithelial cells (IOSE25) were
immortalized and cultured as previously described (34). Four human ovarian cancer cell
lines, A2780, OVCAR3, SKOV3 and 3AO were purchased from the Cell
Bank of the Chinese Academy of Sciences (Shanghai, China) and the
American Type Culture Collection (ATCC; Manassas, VA, USA),
respectively.
The ovarian cancer cell lines were maintained
according to the vendor's instructions. Briefly, A2780 and 3AO cell
lines were routinely cultured in Dulbecco's modified Eagle's medium
(DMEM) with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA, USA).
SKOV3 cells were cultured in McCoy's 5A Modified Medium (ATCC) with
10% FBS (Gibco). OVCAR3 cells were cultured in RPMI-1640 medium
(ATCC) with 20% FBS (Gibco). All the media contained 1%
penicillin-streptomycin (100 U/ml penicillin and 100 µg/ml
streptomycin). The ovarian cancer cells were cultured and
maintained in a humidified incubator at 37°C and supplemented with
5% CO2.
Plasmid construction
The apoptotic protease activating factor 1 (APAF1)
3′-untranslated region (3′-UTR) containing putative binding sites
for microRNA-221 (miR-221) was cloned downstream of the psi-CHECK2
vector (Promega, Madison, WI, USA) and named as APAF1-3′UTR-WT.
APAF1 mutant 3′-UTR recombinant plasmid was generated by QuikChange
Site-Directed Mutagenesis kit (Stratagene, La Jolla, CA, USA),
which generated a mutation of 7 bp from AUGUAGC to UACGCGU in the
predicted miR-221 target binding site, named as APAF1-3′UTR-MUT.
The full-length cDNA encoding human APAF1 was amplified and the
recombinant plasmid, pcDNA3.1-APAF1 was constructed. All plasmids
were confirmed by DNA sequencing.
Cell transfection
The miR-221 mimics, miR-221 inhibitor and scrambled
sequence pre-miR negative control (NC) were purchased from a
commercial manufacturer (Guangzhou RiboBio, Co., Ltd., Guangzhou,
China). Ovarian cancer cells (100 µl) were seeded in 24-well
plates (1×106 cells/ml) and incubated for 24 h, then
cells were transfected with miRNA mimics/inhibitor (50 mM) or
vectors (2 µg) using Lipofectamine 2000 (Invitrogen,
Carlsbad, CA, USA) in serum-free medium in accordance with the
manufacturer's instructions.
RNA extraction and quantitative real-time
PCR (qRT-PCR) analysis
TRIzol reagent (Invitrogen) was used to extract
total RNA from tissue samples or cultured cells. For quantitative
real-time polymerase chain reaction (qRT-PCR), 2 µg of total
RNA was used for the reverse transcription reaction and cDNA
synthesis by using a Reverse Transcription kit (Takara, Dalian,
China). Quantitative real-time PCR analysis was performed with
SYBR-Green Real-Time Master Mix (Toyobo, Co., Ltd, Osaka, Japan).
Results were normalized to a constitutive expression gene, GAPHD.
The relative expression of miRNA-221 was detected using a SYBR
PrimeScript miRNA RT PCR kit (Takara) in accordance with the
manufacturer's instructions and U6 was used as an internal control.
The gene-specific primers were synthesized by Sangon Biotech, Co.,
Ltd. (Shanghai, China) (Table
III). qRT-PCR and data collection were performed on an Applied
Biosystems 7500 Sequence Detection system (Applied Biosystems,
Foster City, CA, USA). The relative expression of miR-221 and APAF1
were calculated and normalized using the 2−ΔΔCt
method.
| Table IIIPrimer sequences used for miRNA and
mRNA expression analysis. |
Table III
Primer sequences used for miRNA and
mRNA expression analysis.
| Name | Primer sequence
(5′-3′) |
|---|
| miR-221-RT |
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTGGGGTATT |
| U6-RT |
CGCTTCACGAATTTGCGTGTCAT |
| U6-F |
CTCGCTTCGGCAGCACA |
| U6-R |
AACGCTTCACGAATTTGCGT |
| miR-221-F |
ACACTCCAGCTGGGTGTCAGTTTGTCAA |
| Universal-R |
CTCAACTGGTGTCGTGGA |
| APAF1-F |
TTGCTGCCCTTCTCCATGAT |
| APAF1-R |
TCCCAACTGAAACCCAATGC |
| GAPDH-F |
TGTTCGTCATGGGTGTGAA |
| GAPDH-R |
ATGGCATGGACTGTGGTCAT |
Protein extraction and western blot
analysis
Total proteins were extracted from tissue samples or
cultured cells with SDS lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) on ice for 20 min and the protein
concentrations were determined using BCA protein assay kit (Pierce,
Rockford, IL, USA). Equal amounts of total proteins were separated
by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) at 120 V
for 2 h, transferred to 0.22 µm polyvinylidene difluoride
membranes (PVDF) (Millipore, Billerica, MA, USA) and incubated with
APAF1 antibodies (1:1,000, ab32372; Abcam). Proteins were detected
by enhanced chemiluminescence (ECL) as described by the
manufacturer (Beyotime Institute of Biotechnology) and the
intensity of the bands was quantified by densitometry (Quantity One
software; Bio-Rad Laboratories, Hercules, CA, USA). Results were
normalized to a constitutive expression gene, GAPHD (1:2,000,
ab9485; Abcam).
Luciferase assays
For the reporter assay, 1×105 cells were
cultured in 24-well plates one day before transfection. A total of
100 ng APAF1-3′UTR-WT or -MUT vectors were co-transfected with 50
nM miR-221 mimics or negative control into cells using
Lipofectamine 2000 reagent. After 48 h of transfection, luciferase
activities were measured using Dual-luciferase reporter assay
system (Promega) according to the manufacturer's instructions.
Firefly luciferase activity was normalized to Renilla
luciferase activity.
Cell proliferation assays
Cell Counting kit-8 (CCK-8; Dojindo Laboratories,
Kumamoto, Japan) was used to examine the cell proliferation,
according to the manufacturer's instruction. Briefly,
2×104 cells were seeded into a 96-well plate. Cell
viability was evaluated with CCK-8 at daily intervals from the next
24, 48 and 72 h after seeding. Following the CCK-8 assay at 37°C
for 1 h, ovarian cancer cells were used to measure the absorbency
at 450 nm using a microplate reader Thermo Plate (Rayto Life and
Analytical Science, Co., Ltd., Hamburg, Germany).
Cell migration and invasion assays
A Transwell migration assay and a Matrigel invasion
assay were performed separately using 24-well Transwell inserts
with 8-µm pore size (Corning Costar Corp., Corning, NY,
USA). For the Transwell migration assay, 2×104 ovarian
cancer cells were plated in 100 µl corresponding culture
medium without FBS into the upper chamber of a Transwell insert
with a non-coated membrane. For the Matrigel invasion assay,
4×104 ovarian cancer cells were suspended in 100
µl serum-free corresponding culture medium were loaded in
the upper Matrigel-coated chamber. In both assays, 500 µl of
culture medium containing 20% FBS was added to the lower chamber.
Cells were then allowed to migrate or invade for 24 h at 37°C. The
cells that migrated or invaded into the bottom chamber were fixed
with 100% methanol for 30 min and stained using 0.5% crystal violet
(Sigma-Aldrich, St. Louis, MO, USA) for 20 min, and the permeating
cells were counted under a phase-contrast microscope (Olympus
Corp., Tokyo, Japan).
Cell apoptosis
Cell apoptosis was studied using an Annexin
V-fluorescein isothiocyanate (FITC) and propidium iodide (PI)
apoptosis detection kit (BestBio, Shanghai China) with flow
cytometry and in accordance with the manufacturer's instruction.
Briefly, 1×106 cells were harvested and resuspended in
cold PBS, and then stained using the Annexin V FITC/PI apoptosis
detection kit according to the manufacturer's instructions. Samples
were analyzed using Becton-Dickinson flow cytometer. Annexin
V(+)/PI(−) and Annexin V(+)/PI(+) represented the ovarian cancer
cells in early and late apoptosis or necrosis, respectively.
Hoechst 33342 nuclear staining method was performed
to detect cell apoptosis. Briefly, cells were cultured with Hoechst
33342 blue fluorescent nuclear dye (Sigma-Aldrich) in 6-well cell
culture plates for 30 min. The nuclear morphology was examined
using fluorescence microscopy with a filter for Hoechst 33342 at
365 nm.
Statistical analysis
All results were expressed as the mean ± standard
deviation (SD). Statistical analysis was performed using the SPSS
18.0 (SPSS, Inc., Chicago, IL, USA). Statistical significance
between the groups was determined using the Student's t-test or the
Chi-square test. Survival analysis was performed using the
Kaplan-Meier method. The log-rank (Mantel-Cox) test was used to
compare the differences between the patient groups.
Results
miR-221 upregulation in human ovarian
cancer tissues
Fig. 1A shows that
miR-221 was upregulated in ovarian cancer tissues compared with
matched adjacent normal tissues in 63 cases, using quantitative
reverse transcription polymerase chain reaction (qRT-PCR). In tumor
specimens, miR-221 expression levels were greater than in adjacent
normal tissues (median ratio of tumor to normal of 3.68).
miR-221 expression and
clinicopathological features in ovarian cancer patients
The clinical pathology findings in 63 ovarian cancer
patients studied are shown in Table
I. The 63 ovarian cancer patients were classified into two
groups based on the median relative miR-221 expression (3.68) in
ovarian cancer tissues. Group 1: a low miR-221 expression group
(n=31, miR-221 expression ≤ median). Group 2: a high miR-221
expression group (n=32, miR-221 expression ≥ median) (Fig. 1B).
A comparison of the clinicopathological features of
the two groups is shown in Table
II. The high-miR-221 group (group 2) had a larger tumor size
(P=0.0010), deeper depth of tumor invasion (P=0.0003) and higher
FIGO stage (P=0.0076) when compared with the low-miR-221 group
(group 1). However, the miR-221 expression level was not associated
with other parameters such as age (P=0.8052), histologic
differentiation (tumor grade) (P=0.9463), lymphatic spread
(P=0.4985), or distant metastasis (P=0.6723) (Table II).
| Table IIThe relationship between the GAS5
expression and the clinicopathological factors in ovarian cancer
patients. |
Table II
The relationship between the GAS5
expression and the clinicopathological factors in ovarian cancer
patients.
| Clinical
parameter | miR-221
| Chi-squared test
P-value |
|---|
| Group 1 | Group 2 |
|---|
| Age (years) | | | 0.8052 |
| <55 | 14 | 15 | |
| >55 | 18 | 16 | |
| Size (cm) | | | 0.0010 |
| >5 | 9 | 24 | |
| <5 | 21 | 9 | |
| Histologic
differentiation | | | 0.9463 |
| Well | 3 | 2 | |
| Moderate | 11 | 12 | |
| Poor | 14 | 14 | |
|
Undifferentiated | 4 | 3 | |
| Invasion depth | | | 0.0003 |
| T1 | 10 | 5 | |
| T2 | 13 | 3 | |
| T3 | 4 | 14 | |
| T4 | 3 | 12 | |
| FIGO stages | | | 0.0076 |
| I | 6 | 2 | |
| II | 15 | 7 | |
| III | 8 | 19 | |
| IV | 1 | 5 | |
| Lymphatic
metastasis | | | 0.4985 |
| Yes | 18 | 12 | |
| No | 17 | 16 | |
| Distant
metastasis | | | 0.6723 |
| Yes | 3 | 5 | |
| No | 25 | 30 | |
miR-221 expression and patient
survival
Disease-free survival (DFS) and overall survival
(OS) curves were plotted according to miR-221 expression levels,
using the Kaplan-Meier analysis and the log-rank test,
respectively. The results are presented in Fig. 1C and 1D. Patients with high miR-221 expression
levels had a reduced DFS (P=0.0014) and OS (P=0.0058). For the OS,
3 years of overall accumulative survival rates of ovarian cancer
patients with low and high miR-221 expression level were 38.57 and
20.68%, respectively. High expression of miR-221 was associated
with a greater OS time for ovarian cancer patients (median OS, 14
months) compared with patients with low miR-221 expression (median
OS, 27 months). The 3-year disease-free survival rates for high and
low expression of miR-221 patients were 35.21 and 18.54%,
respectively. The median survival time for patients with low
miR-221 expression was 22 months, and 9 months for patients with
high miR-221 expression.
Decreased APAF1 protein expression in
ovarian cancer tissues; negative correlation with miR-221
expression
The results of western blotting in ovarian cancer
tissues and ovarian cancer lines SKOV3, OVCAR-3, A2780, 3AO and
human ovarian surface epithelium cells showed that APAF1 protein
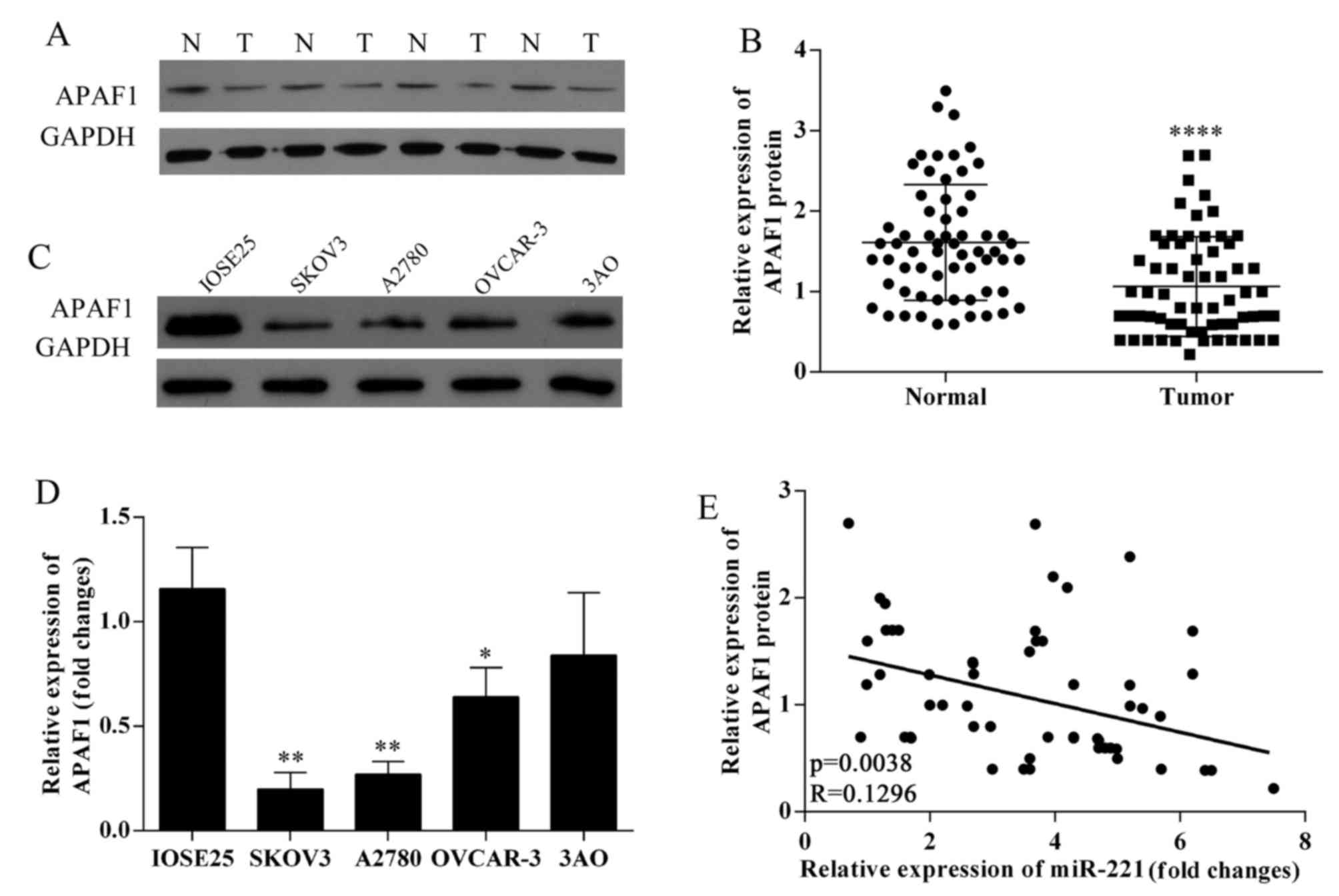
expression was downregulated in ovarian cancer tissues (Fig. 2A and B). APAF1 protein expression
was down-regulated in ovarian cancer cell lines (Fig. 2C). The expression of APAF1 in
SKOV3, OVCAR-3, A2780, 3AO and human ovarian surface epithelium
cells were detected using qRT-PCR (Fig. 2D); SKOV3 and A2780 cells were
chosen for further study. APAF1 protein expression levels were
negatively associated with miR-221 expression levels in 63 ovarian
cancer tissues (P=0.0038; R=0.1296; Fig. 2E).
APAF1 as a potential target of
miR-221
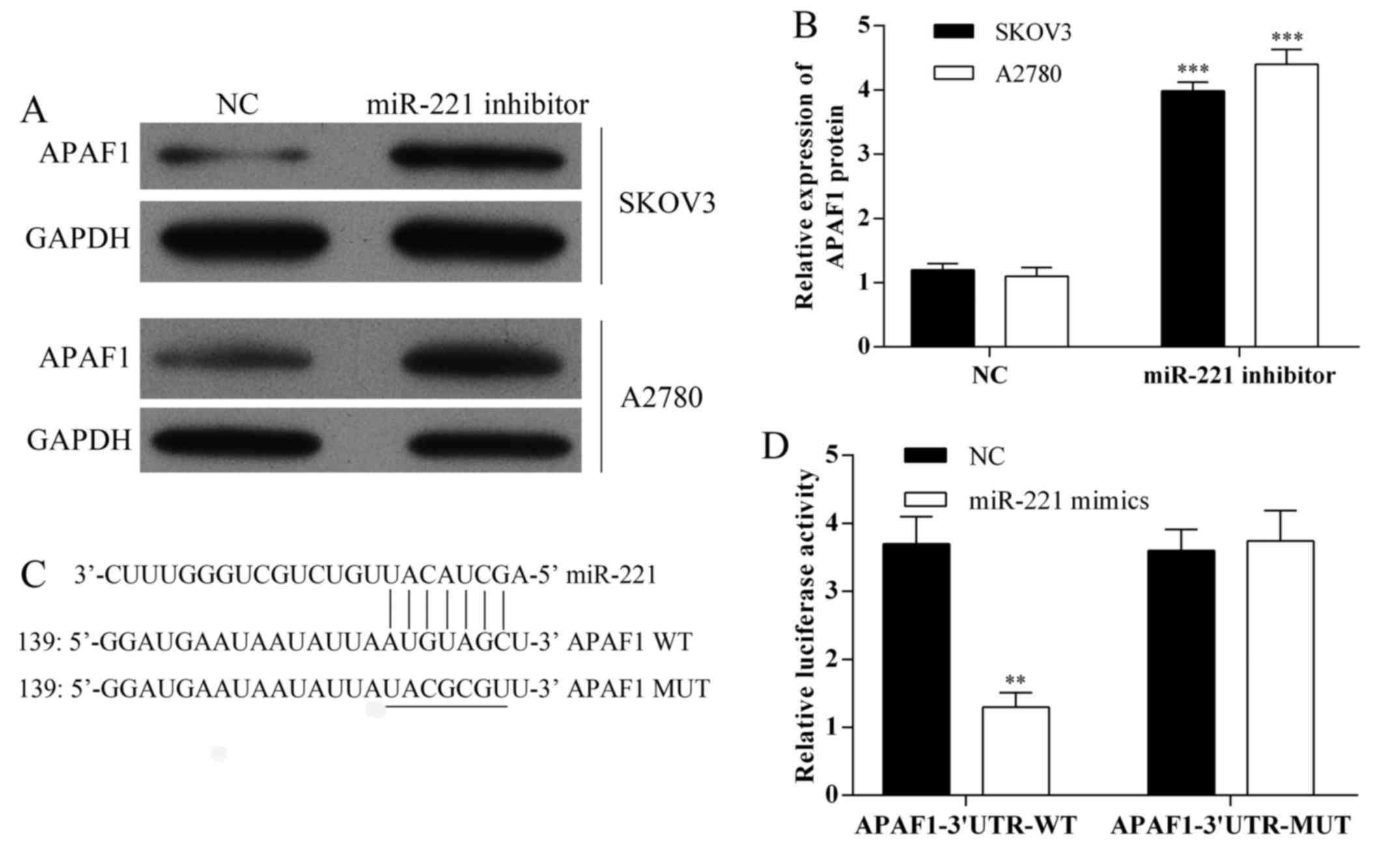
The expression of APAF1 in SKOV3 and A2780 cells
following transfection with miR-221 inhibitor were detected using
western blot analysis (Fig. 3A and
B). Also, miRNA target predication databases (TargetScan:
www.targetscan.org and MICRORNA.ORG: www.microrna.org) were used for computational
analysis. miR-221 had one predictive target site in the human
APAF1-3′-UTR (Fig. 3C). A
Dual-luciferase reporter system was used to determine whether APAF1
was a direct target of miR-221. The results showed that miR-221
mimics downregulated the luciferase activity of the reporter and
the luciferase expression of mutant APAF1-3′-UTR was not regulated
by miR-221 (Fig. 3D). These
results indicated that this site of APAF1-3′-UTR was the regulation
site for miR-221.
Transfection with miR-221 inhibitor and
suppression of ovarian cancer cell proliferation, migration and
invasion in vitro
The proliferation of SKOV3 and A2780 cells were
suppressed following miR-221 inhibitor transfection (Fig. 4A). Using the Transwell assay the
cell migration and invasion of SKOV3 and A2780 were inhibited
following miR-221 inhibitor transfection (Fig. 4B and C).
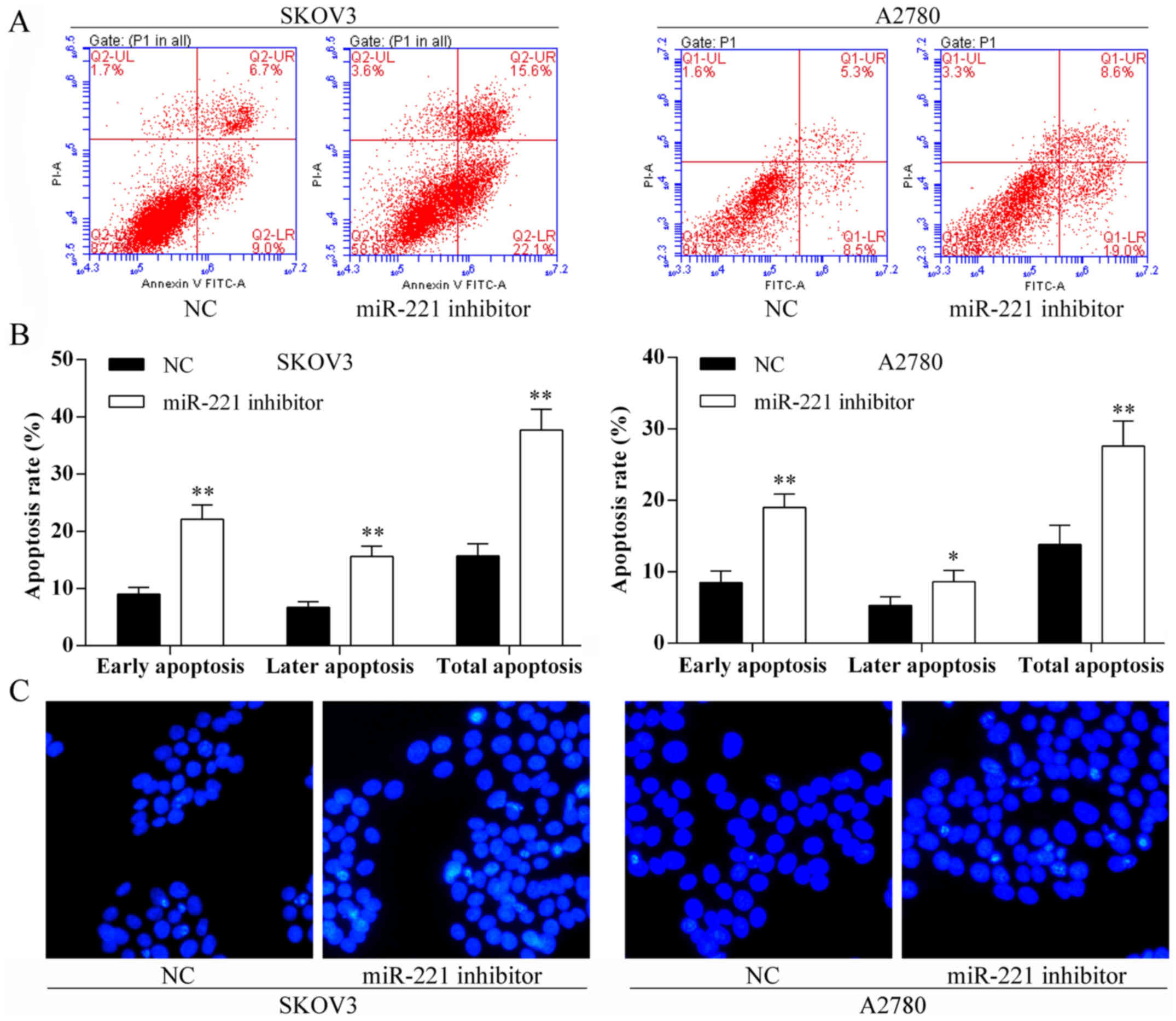
Transfection with miR-221 inhibitor and
ovarian cancer cell apoptosis in vitro
Using the Annexin V-FITC/PI staining and Hoechst
staining methods, the SKOV3 and A2780 cell apoptosis rates were
induced after miR-221 inhibitor transfection (Fig. 5).
Overexpression of APAF1 and suppression
of ovarian cancer cell proliferation, migration and invasion in
vitro
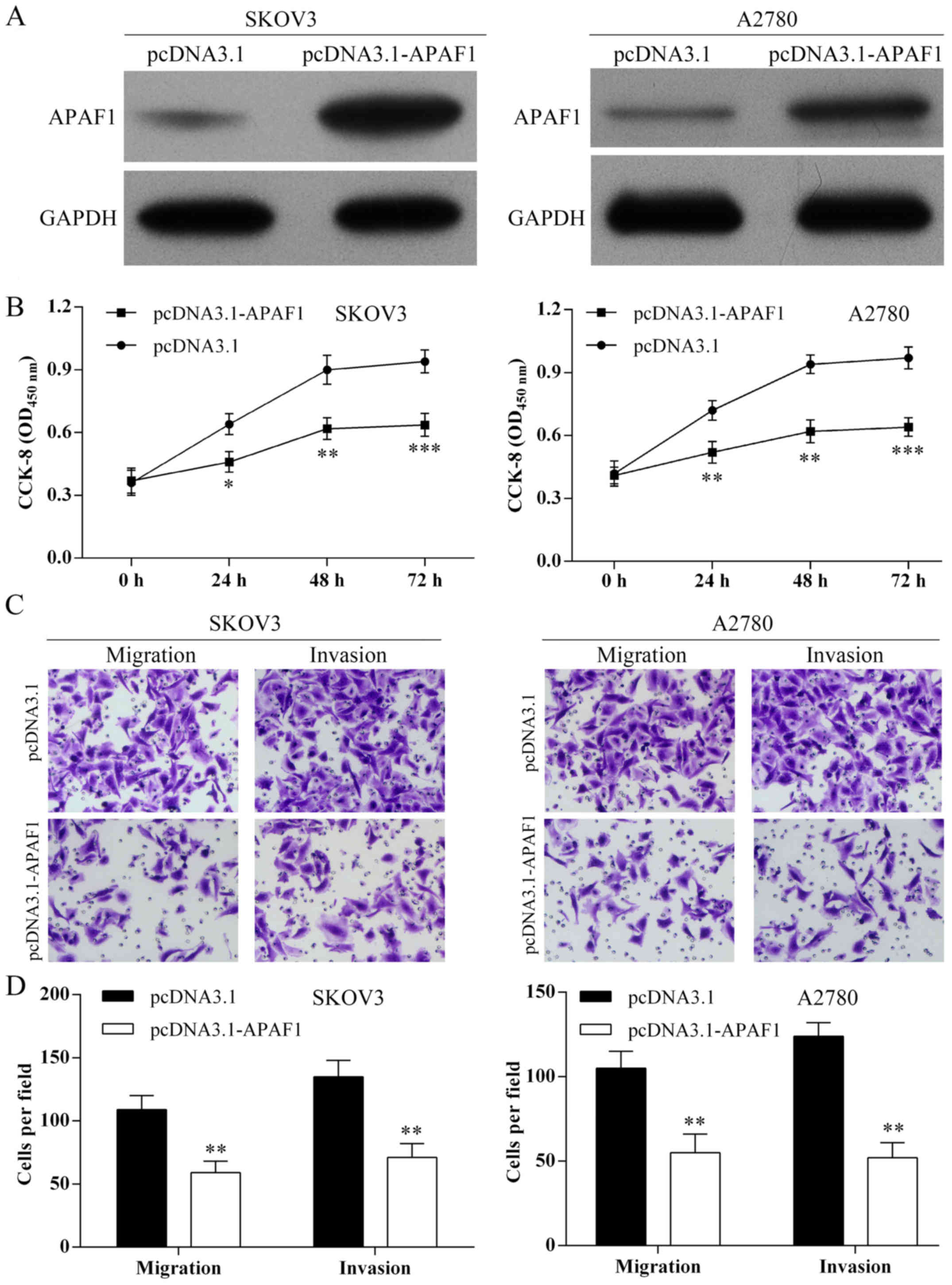
As shown in Fig.
6A, APAF1 was overexpressed in SKOV3 and A2780 cells following
pcDNA3.1-APAF1 transfection. Using the CCK-8 assay, overexpression
of APAF1 inhibited ovarian cancer cell proliferation (Fig. 6B). Using Transwell assay,
overexpression of APAF1 suppressed the SKOV3 and A2780 cell
migration and invasion (Fig. 6C and
D).
Overexpression of APAF1 and ovarian
cancer cell apoptosis in vitro
The Annexin V-FITC/PI staining and Hoechst staining
methods showed that the ovarian cancer cell apoptosis rates were
induced following overexpression of APAF1 (Fig. 7).
Discussion
The present study was designed to investigate the
expression of microRNA-221 (miR-221) in ovarian cancer tissues and
ovarian carcinoma cell lines and to undertake a preliminary study
of the prognostic significance of miR-221 in patients following
surgery for ovarian carcinoma. The results showed that the
expression of miR-221 was increased in ovarian cancer tissues
compared with matched adjacent normal tissue. Increased expression
levels of miR-221 were associated with increased ovarian tumor
size, increased depth of tumor invasion, a higher tumor stage,
reduced patient prognosis and survival. Tumor cell transfection
with the miR-221 inhibitor induced apoptosis protease activating
factor 1 (APAF1) protein expression and confirmed that APAF1 is a
target gene for miR-221. Following miR-221 inhibitor transfection,
ovarian cancer cell proliferation, migration and invasion were
inhibited in vitro and cell apoptosis was induced in
vitro. The findings of the present study indicate that miR-221
may act as an oncogene by regulating tumor cell growth, invasion,
migration and apoptosis in ovarian cancer, and that miR-221-APAF1
may represent a new potential diagnosis and therapeutic biomarker
in ovarian cancer.
The findings of this study on ovarian cancer cell
lines and tissue support the finding of other studies on the role
of miR-221 in other types of human cancer (11,12).
Also, this study confirmed the findings of other studies on human
malignancy that APAF1 expression is associated with inhibition of
tumor cell apoptosis (27,31). This study also confirmed that the
expression of APAF1 is associated with adverse patient prognosis
(32,33). However, this study showed, for the
first time that the APAF1 gene is a direct target of miR-221
in human ovarian cancer cells and that the overexpression of APAF1
suppressed ovarian cancer cell proliferation and induced cell
apoptosis in vitro.
The present study was preliminary in nature, and so
further, larger studies are required to confirm and expand the
findings. Although multiple tumor cell lines were studied, the
behavior of immortalized tumor cells in vitro cannot
replicate that of human cancer cells in vitro. For this
reason, human ovarian cancer tissues were studied in parallel.
However, the number of human tumor samples was small and confined
to those from a single center, we suggest that, for future studies,
a larger number of tumor samples should be studied and from
multiple centers.
In conclusion, the APAF1 gene was confirmed
as a direct target of miR-221 and overexpression of APAF1
suppressed ovarian cancer cell proliferation and induced cell
apoptosis in vitro. These findings indicate that
miR-221-APAF1 should be studied further as a potential new
diagnostic or prognostic biomarker for ovarian cancer.
Acknowledgments
We thank Guangzhou Vipotion Biotechnology Co., Ltd.
for the assistance in the vector construction of luciferase
reporter assay.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Breuer EK and Murph MM: The role of
proteomics in the diagnosis and treatment of women's cancers:
current trends in technology and future opportunities. Int J
Proteomics. 2011:3735842011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Doench JG and Sharp PA: Specificity of
microRNA target selection in translational repression. Genes Dev.
18:504–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meyer SU, Thirion C, Polesskaya A,
Bauersachs S, Kaiser S, Krause S and Pfaffl MW: TNF-α and IGF1
modify the microRNA signature in skeletal muscle cell
differentiation. Cell Commun Signal. 13:42015. View Article : Google Scholar
|
|
7
|
Tian L, Fang YX, Xue JL and Chen JZ: Four
microRNAs promote prostate cell proliferation with regulation of
PTEN and its downstream signals in vitro. PLoS One. 8:e758852013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ambros V: MicroRNA pathways in flies and
worms: Growth, death, fat, stress, and timing. Cell. 113:673–676.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M,
et al: Human microRNA genes are frequently located at fragile sites
and genomic regions involved in cancers. Proc Natl Acad Sci USA.
101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hummel R, Hussey DJ and Haier J:
MicroRNAs: Predictors and modifiers of chemo- and radiotherapy in
different tumour types. Eur J Cancer. 46:298–311. 2010. View Article : Google Scholar
|
|
11
|
Nassirpour R, Mehta PP, Baxi SM and Yin
MJ: miR-221 promotes tumorigenesis in human triple negative breast
cancer cells. PLoS One. 8:e621702013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stinson S, Lackner MR, Adai AT, Yu N, Kim
HJ, O'Brien C, Spoerke J, Jhunjhunwala S, Boyd Z, Januario T, et
al: TRPS1 targeting by miR-221/222 promotes the
epithelial-to-mesenchymal transition in breast cancer. Sci Signal.
4:ra412011.PubMed/NCBI
|
|
13
|
Rong M, Chen G and Dang Y: Increased
miR-221 expression in hepatocellular carcinoma tissues and its role
in enhancing cell growth and inhibiting apoptosis in vitro. BMC
Cancer. 13:212013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamashita R, Sato M, Kakumu T, Hase T,
Yogo N, Maruyama E, Sekido Y, Kondo M and Hasegawa Y: Growth
inhibitory effects of miR-221 and miR-222 in non-small cell lung
cancer cells. Cancer Med. 4:551–564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Adams BD, Kasinski AL and Slack FJ:
Aberrant regulation and function of microRNAs in cancer. Curr Biol.
24:R762–R776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Berindan-Neagoe I, Monroig PC, Pasculli B
and Calin GA: MicroRNAome genome: A treasure for cancer diagnosis
and therapy. CA Cancer J Clin. 64:311–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hauser B, Zhao Y, Pang X, Ling Z, Myers E,
Wang P, Califano J and Gu X: Functions of MiRNA-128 on the
regulation of head and neck squamous cell carcinoma growth and
apoptosis. PLoS One. 10:e01163212015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Visone R, Russo L, Pallante P, De Martino
I, Ferraro A, Leone V, Borbone E, Petrocca F, Alder H, Croce CM, et
al: MicroRNAs (miR)-221 and miR-222, both overexpressed in human
thyroid papillary carcinomas, regulate p27Kip1 protein
levels and cell cycle. Endocr Relat Cancer. 14:791–798. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fornari F, Gramantieri L, Ferracin M,
Veronese A, Sabbioni S, Calin GA, Grazi GL, Giovannini C, Croce CM,
Bolondi L, et al: MiR-221 controls CDKN1C/p57 and CDKN1B/p27
expression in human hepatocellular carcinoma. Oncogene.
27:5651–5661. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Medina R, Zaidi SK, Liu CG, Stein JL, van
Wijnen AJ, Croce CM and Stein GS: MicroRNAs 221 and 222 bypass
quiescence and compromise cell survival. Cancer Res. 68:2773–2780.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
O'Hara AJ, Wang L, Dezube BJ, Harrington
WJ Jr, Damania B and Dittmer DPP: Tumor suppressor microRNAs are
underrepresented in primary effusion lymphoma and Kaposi sarcoma.
Blood. 113:5938–5941. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Okamoto K, Miyoshi K and Murawaki Y:
miR-29b, miR-205 and miR-221 enhance chemosensitivity to
gemcitabine in HuH28 human cholangiocarcinoma cells. PLoS One.
8:e776232013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cecconi F, Alvarez-Bolado G, Meyer BI,
Roth KA and Gruss P: Apaf1 (CED-4 homolog) regulates programmed
cell death in mammalian development. Cell. 94:727–737. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou H, Henzel WJ, Liu X, Lutschg A and
Wang X: Apaf-1, a human protein homologous to C. elegans CED-4,
participates in cytochrome c-dependent activation of caspase-3.
Cell. 90:405–413. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao Y, Liang W, Hu X, Zhang W, Stetler RA,
Vosler P, Cao G and Chen J: Neuroprotection against
hypoxic-ischemic brain injury by inhibiting the apoptotic protease
activating factor-1 pathway. Stroke. 41:166–172. 2010. View Article : Google Scholar
|
|
27
|
Yong FL, Wang CW, Roslani AC and Law CW:
The involvement of miR-23a/APAF1 regulation axis in colorectal
cancer. Int J Mol Sci. 15:11713–11729. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De Zio D, Bordi M, Tino E, Lanzuolo C,
Ferraro E, Mora E, Ciccosanti F, Fimia GM, Orlando V and Cecconi F:
The DNA repair complex Ku70/86 modulates Apaf1 expression upon DNA
damage. Cell Death Differ. 18:516–527. 2011. View Article : Google Scholar :
|
|
29
|
Ho CK, Bush JA and Li G: Tissue-specific
regulation of Apaf-1 expression by p53. Oncol Rep. 10:1139–1143.
2003.PubMed/NCBI
|
|
30
|
Marsden VS, O'Connor L, O'Reilly LA, Silke
J, Metcalf D, Ekert PG, Huang DC, Cecconi F, Kuida K, Tomaselli KJ,
et al: Apoptosis initiated by Bcl-2-regulated caspase activation
independently of the cytochrome c/Apaf-1/caspase-9 apoptosome.
Nature. 419:634–637. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mustika R, Budiyanto A, Nishigori C,
Ichihashi M and Ueda M: Decreased expression of Apaf-1 with
progression of melanoma. Pigment Cell Res. 18:59–62. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Paik SS, Jang KS, Song YS, Jang SH, Min
KW, Han HX, Na W, Lee KH, Choi D and Jang SJ: Reduced expression of
Apaf-1 in colorectal adenocarcinoma correlates with tumor
progression and aggressive phenotype. Ann Surg Oncol. 14:3453–3459.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zlobec I, Minoo P, Baker K, Haegert D,
Khetani K, Tornillo L, Terracciano L, Jass JR and Lugli A: Loss of
APAF-1 expression is associated with tumour progression and adverse
prognosis in colorectal cancer. Eur J Cancer. 43:1101–1107. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li NF, Broad S, Lu YJ, Yang JS, Watson R,
Hagemann T, Wilbanks G, Jacobs I, Balkwill F, Dafou D, et al: Human
ovarian surface epithelial cells immortalized with hTERT maintain
functional pRb and p53 expression. Cell Prolif. 40:780–794. 2007.
View Article : Google Scholar : PubMed/NCBI
|