Introduction
Gastric cancer is one of the most common types of
cancer, currently, and it remains a worldwide burden as the second
leading cause of cancer-associated death, whereas the incidence
rate is gradually rising every year in the world (1,2).
Though a variety of advances, such as surgery, have been achieved
to inhibit gastric cancer progression in recent years, the
therapeutic effect is still far from satisfactory (3). Furthermore, various chemical drugs
are suggested as the first-line therapy. However, side effects are
observed due to cytotoxicity (4).
In addition, the drug resistance to chemotherapeutics develops in
gastric cancer cells, resulting in tumor recurrence and even
further progression (5,6). Therefore, the drug resistance of
chemotherapy is considered as a major clinical obstacle to
successful treatment, and new chemotherapeutics, including drugs in
combination, used against human gastric cancer are still needed
(7). Cisplatin (DDP)-based
chemotherapy is reported as the main strategy to prevent human
gastric cancer. However, drug resistance is also a major
obstruction for DDP chemotherapy (8,9).
Licorice, known as medicinal plant in China, Korea
and Japan, has a variety of pharmacological activities, including
antibacterial, anti-inflammatory and antioxidant activities, and
even anticancer ability (10,11).
Liquiritin (LIQ) is a main component among the licorice flavonoids,
and has also been suggested to possess anti-inflammatory and
anticancer abilities (12,13). However, its effects, as well as its
combination with DDP, have not been well investigated on the
regulation of human gastric cancer with drug resistance.
In the present study, for the first the synergistic
effect of LIQ on DDP-induced apoptosis and autophagy was evaluated
in human gastric cancer SGC-7901/DDP cells. Our results indicated
that LIQ enhanced the sensitivity of SGC-7901/DDP cells to DDP
treatment through inducing G0/G1 cell cycle arrest, increasing the
pro-apoptosis via upregulating the cleaved caspase-8/-9/-3 and
PARP, and inducing autophagy by improving LC3B and Beclin 1
expression. In vivo, LIQ enhanced the suppressive effects of
DDP on tumor growth in SGC-7901/DDP xenograft model. Our results
suggested that LIQ might serve as a synergistic drug with
chemotherapeutic drugs DDP in the prevention of gastric cancer.
Materials and methods
Cells and culture
The drug (DDP)-resistant SGC7901/DDP cells were
purchased from the KeyGen Biotech Development Co., Ltd. (Nanjing,
China). The cells were grown in RPMI-1640 medium containing 10%
(v/v) fetal calf serum (Gibco, USA), 100 U/ml penicillin, and 100
µg/ml streptomycin. All cells were then incubated in a 37°C
incubator with 5% CO2. SGC7901/DDP cells were routinely
maintained in the above RPMI-1640 medium containing 10 µg/ml
cisplatin (Sigma-Aldrich Chemical Co., USA). Lquiritin (purity,
HPLC ≥98%) was obtained from Xi'an Jing Cheng Biological Technology
Co., Ltd. (Xi'an, China).
Cell death analysis
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide
(MTT) (Beyotime, Nanjing, China) was used to calculate cell
viability. Cells (2×103)/well were seeded on 96-well
plates and treated under different conditions as indicated and
incubated at 37°C (14,15). MTT solution (300 µl/well)
was added after incubation. Following incubation at 37°C for an
additional 4 h, the supernatants were removed and 200 µl
dimethyl sulfoxide (DMSO, Sigma-Aldrich) was added into each well
to dissolve the formazan crystals. The 96-well plates were then
placed in a microplate reader to determine the absorbance at 490
nm. Each test was carried out in triplicate.
Migration assays
SGC7901/DDP cells (1×104) after
incubation with DPP, LIQ or the co-treatment of the two drugs for
24 h were seeded into the upper chamber of a Transwell®
system (Corning, USA) following the manufacturer's instructions.
The medium in the lower chamber contained 5% FBS as a source of
chemoattractants. Cancer cells were suspended in serum-free medium.
The cultures were rinsed with PBS and the medium was replaced with
fresh medium alone or supplemented with 10% FBS. The cells were
then incubated for 24 h at 37°C. The number of cells in migration
was counted under a microscope.
Colony formation analysis
SGC7901/DDP cells (1×104) were first
incubated with DPP, LIQ or the two in combination for 24 h. Also,
then the medium was removed and the cells were seeded into the
upper chamber of Transwell systems (Corning) in accordance with the
manufacturer's instructions. The medium in the lower chamber was
supplemented with 5% fetal bovine serum (FBS, Gibco) as a source of
chemoattractants. SGC7901/DDP cells were then suspended in
serum-free medium. The cultures were washed with PBS and the medium
was replaced with fresh medium alone or supplemented with 5% FBS.
Next, SGC7901/DDP cells were incubated for another 24 h at 37°C.
The number of migrated cells was counted with a microscope.
The wound healing analysis
SGC7901/DDP cells were seeded and grown on a 6-well
plate treated with DPP, LIQ or the two in combination for 24 h.
Then, the monolayers of cancer cells were wounded using a pipette
tip. Gastric cancer cells were then rinsed with PBS to remove
cellular debris and subjected to migration for 24 h. The
representative images were captured on 0 and 24 h, respectively,
after the wound using an inverted microscope.
Cell cycle arrest analysis
The effect of LIQ, DDP or LIQ/DDP co-treatment on
cell cycle distribution was examined using flow cytometry analysis.
After treatment under various conditions, all cells were harvested
and fixed in 70% ethanol. The cells were then washed with PBS twice
and stained with PI solution containing 50 µg/ml PI and 25
µg/ml RNAse for 30 min. Finally, all cells were analyzed by
a FACSCalibur flow cytometer (BD Pharmingen, USA).
Assessment of apoptosis
SGC7901/DDP cells treated under different conditions
as indicated were harvested in PBS. Then, the cells were incubated
with Annexin V-FITC and PI (Sigma-Aldrich) for 15 min in a darkroom
at room temperature. Then, the stained cells were analyzed through
flow cytometry (BD Pharmingen).
Hoechst 33258 staining of SGC7901/DDP cells was used
to determine the apoptotic status. Cells (1×106)/ml
SGC7901/DDP cells were seeded in 6-well plates and then treated as
indicated. The cells were collected after various treatments,
washed with PBS twice, and fixed with 4% paraformaldehyde (Taize
Ruida Technology Co. Ltd., Beijing, China) for 10 min and then
washed with PBS three times. Next, the cells were stained with
Hoechst 33258 solution (Beyotime) following the manufacturer's
instructions. The representative images were immediately captured
through a fluorescence microscope (Olympus Corp., Japan) to
calculate the apoptotic cells.
SGC7901/DDP cells were exposed to LIQ, DDP or the
two in combination for 24 h on 12-well plates. Next, the One Step
TUNEL Apoptosis assay kit (KeyGen Biotech) was used to determine
the apoptosis following the manufacturer's instructions. The
staining intensity was assessed through a fluorescence
microscopy.
Mitochondrial potential evaluation
JC-1 and JC-10 dye loading solutions (Abcam, USA)
were added to SGC7901/DDP cells after various treatments as
described and incubated for 30 min. The fluorescent intensities for
both J-aggregates and monomeric forms of JC-1 and JC-10 were
calculated at Ex/Em = 490/525 nm and 490/590 nm using a microplate
reader.
Measurement of caspase activity
Caspase activity kits (Beyotime Biotech) were used
to assess caspase activity according to the manufacturer's
instructions. SGC7901/DDP cells underwent various treatments, then
the lysates were prepared. Activities of caspase-8/-9 and -3 was
measured by the use of substrate peptides Ac-IETD-pNA, Ac-LEHD-pNA
and Ac-DEVD-pNA, respectively. Also, the pNA release was determined
by calculating the absorbance at 405 nm using a microplate
reader.
Transmission electron microscopy (TEM)
assays
After different treatments as indicated, SGC7901/DDP
cells were collected and washed with PBS and then fixed in
glutaraldehyde (2.5%) overnight. The cells were washed by PBS (0.1
M) and fixed with OsO4 (1%). Next, the SGC7901/DDP cells
were dehydrated with a range of alcohol concentrations for 15 min.
The cells were then embedded into paraffin and sliced using an
LKB-V ultramicrotome (LKB, Bromma, Sweden). For acquiring TEM
images, the prepared sections were observed on a JEM-2100
microscope operating at an accelerating voltage of 200 kV (Jeol
Ltd., Tokyo, Japan).
Western blot analysis
SGC7901/DDP cells after different treatments were
harvested and washed with chilled PBS. The cells and gastric tumor
tissues were lysed in 50 mM Tris-Cl (pH 7.5) containing 150 mM
NaCl, 0.5% deoxycholic acid, 1% NP-40, 0.1% SDS, and a protease
inhibitor (Sigma-Aldrich) on ice for 10 min to yield a homogenate.
The final supernatants were obtained by centrifugation at 15,000 ×
g for 15 min at 4°C. The concentration of protein was measured
using BCA protein assay kit (Thermo Fisher Scientific, USA) with
bovine serum albumin as a standard. Sample-loading buffer was
added, the mixture was boiled for 5 min. The total protein extract
are used for western blot analysis. Total protein (40 µg)
was loaded and the proteins were then separated using 10% SDS-PAGE
and electrophoretically transferred to the polyvinylidene
difluoride membranes (Millipore, USA). Then, the membranes were
blocked with skim milk (5%) Tris-buffered saline with 0.1% Tween-20
(TBST), washed, and then incubated with primary antibody overnight
at 4°C. The membrane was then washed with TBST three times,
followed by incubation with a horseradish peroxidase
(HRP)-conjugated secondary antibody (Santa Cruz Biotechnology Inc.,
USA) at room temperature for 2 h. Following another round of
washing with TBST, the membrane was then developed using ECL, and
exposed to Kodak (Eastman Kodak Co., USA) X-ray film. The protein
expression levels were defined as grey value by the use of ImageJ
1.38 software (National Institutes of Health, USA) and standardized
to housekeeping gene of GAPDH and expressed as a fold of control.
All experiments were performed in triplicate independently. The
primary antibodies for GAPDH (1:1,000, ab8245, Abcam), p53
(1:1,000, ab1431, Abcam), p62 (1:1,000, ab56416, Abcam), Beclin 1
(1:1,000, ab62557, Abcam), LC3B (1:1,000, ab48394, Abcam),
caspase-8 (1:1,000, ab25901, Abcam), caspase-9 (1:1,000, ab25758,
Abcam), cyclin A (1:1,000, 4656, Cell Signaling Technology, USA),
cyclin D1 (1:1,000, 2978, Cell Signaling Technology), CDK4 (1:1000,
ab137675, Abcam), p21 (1:500, KD1406, Santa Cruz Biotechnology),
caspase-3 (1:1,000, ab52293, Abcam), and PARP (1:1,000, ab4830,
Abcam) were included.
Animals and treatments
Forty, 5-week-old male BALB/c-nu mice, weighed 15–18
g, were subcutaneously injected with 2×105 SGC7901/DDP
cells in the left flank. All mice were purchased from Shanghai
Laboratory Animal Center (Shanghai, China). Before the experiments,
the mice were acclimatized to the new environment for a week. All
protocols used followed the Regulations of Experimental Animal
Administration issued by the Ministry of Science and Technology of
the People's Republic of China. Mouse care and usage were performed
in accordance with the ethical guidelines of Huai'an First People's
Hospital, Nanjing Medical University (Nanjing, China). The mice
were raised in air-conditioned pathogen-free rooms at 25±2°C with
50±10% humidity under controlled lighting (12 h light/day) and fed
with standard laboratory food and water. When the tumors were
visible (the tumor size reached to 50 mm3), the mice
were randomly divided into four groups (10 mice per group). The
control group received the vehicle (PBS) injection i.p., and the
treatment group was administered with LIQ (15 mg/kg), DDP (3 mg/kg)
and the two in combination i.p. every day (16,17).
Liquiritin was dissolved in PBS, and it was dispersed in PBS to
form an aqueous suspension in ultrasonic water bath for i.p.
injection. After treatments for 21 days, all animals were
sacrificed. The tumors were excised and measured. Also, the tumor
tissues were fixed in 10% formalin for further research.
The immunohistochemical analysis
Formalin-fixed tumor tissue samples were first
embedded in paraffin (4%) and then the paraffin-embedded samples
were cut into serial sections (3 µm thickness). Primary
tumor sections were stained with hematoxylin and eosin (H&E).
Mouse gastric tumors were sectioned (3 µM thickness), and
stained with Ki-67 (Abcam), and analyzed through a microscope. The
representative images were arranged using TissueFAXs
(Tissue-Gnostics) software. The percentage of Ki-67-positive cells
in each tumor section was quantified. The apoptosis of tumor
tissues was also measured via TUNEL assay using a In situ Apoptosis
Detection kit (KeyGen Biotech) in line with the manufacturer's
protocol. After deparaffinization and hydration, tumor tissue
sections were rinsed with PBS for twice and then incubated with
proteinase K (20 µg/ml, Abcam) at 37°C for 25 min, which was
followed by washing with PBS. Next, all sections were incubated
with TUNEL mixture. The gastric tumor sections were counter-stained
with DAPI. In the end, the tissue sections were observed with a
microscope. In addition, for immunohistochemical analysis of
cleaved caspase-3 and LC3B, tumor tissue sections were
deparaffinized in xylene, followed by hydration through graded
alcohol, and treated with 3% H2O2 for 15 min.
The boiling sodium citrate buffer was used for antigen retrieval
for 10 min. Next, tumor tissue sections were incubated with 10%
normal goat serum for 15 min, and then incubated with primary
antibodies of active caspase-3 and LC3B at 4°C overnight. Next, the
slides were rinsed with PBS three times, the sections were
incubated using secondary antibody for 30 min at 37°C.
3,3′-diaminobenzidine (DAB, KeyGen Biotech) was used for the
staining of the nucleus. Also, hematoxylin (Beyotime) was added for
the counterstaining of the background.
Statistical assays
The quantitative data are presented as the mean ±
SEM of triplicate experiments. Statistically significant values
were compared using the ANOVA and the Dunnett's post hoc test, and
p<0.05 was considered to indicate a statistically significant
result.
Results
Liquiritin potentiates the suppressive
effects of DDP on the proliferation of DDP-resistant gastric cancer
cells
DDP-resistant gastric cancer cells (SGC7901/DDP)
were treated with LIQ to explore if it could potentiate the effects
of DDP on the suppression of gastric cancer progression. SGC7901
cells were cultured with DDP (2 µg/ml) and LIQ (0, 20, 40,
60 and 80 µM) or the two in combination as indicated for 24
h. Then, the cell death condition was investigated using MTT
analysis. As shown in Fig. 1A, we
found that treatment of DDP showed no influence on the cell death
compared to the control group in the absence of DDP and LIQ
(DDP−/LIQ−). LIQ single treatment was found
to significantly increase the percentage of cell death. Of note,
co-treatment of DDP and LIQ further elevated the cell death levels,
which was comparable to the DDP−/LIQ+ group.
Also, LIQ-induced cell death was dose-dependent. Next, the
potential cytotoxicity of LIQ was measured. The rat gastric mucosal
cells (RGM-1, left) and human gastric epithelium cells (GES-1,
right) were cultured with different concentrations (0, 5, 10, 20,
40, 80 and 160 µM) of LIQ for 24 h, followed by MTT
analysis. As shown in Fig. 1B, the
data indicated that there was no significant difference between the
control group and the LIQ-treated groups. Thus, LIQ showed no
cytotoxicity to normal gastric cells treated within our conditions,
demonstrating its safety for application. Following, the morphology
of SGC7901/DDP cells treated with DDP, LIQ or the two in
combination was observed under a microscope. DDP and LIQ
co-treatment showed stronger suppressive role in the proliferation
of SGC7901/DDP cells. Also, clonogenic analysis further confirmed
the effective role of DDP/LIQ in suppressing the gastric cancer
cells. In addition, the migration and wound healing analysis
indicated that DDP/LIQ combination could markedly inhibit the
migration of SGC7901/DDP cells in comparison to the
DDP−/LIQ+ group (Fig. 1C–F). In conclusion, LIQ was able to
remarkably increase the sensitivity of SGC7901/DDP cells to DDP,
contributing to the cell death.
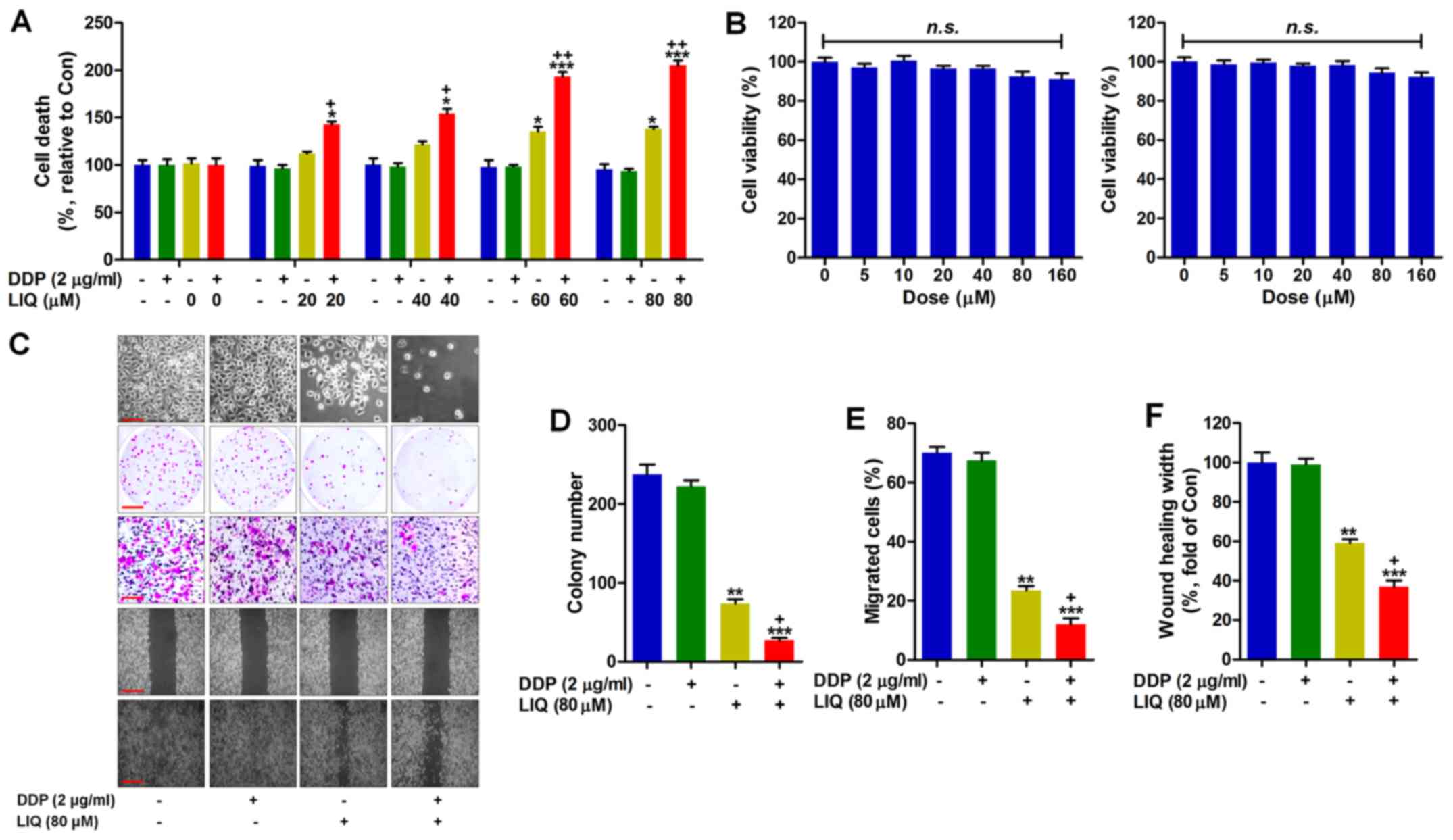 | Figure 1Liquiritin potentiates the
suppressive effects of DDP on the proliferation of DDP-resistant
gastric cancer cells. (A) SGC7901 cells were treated with DDP (2
µg/ml) and LIQ (0, 20, 40, 60 and 80 µM) or the two
in combination as indicated for 24 h. Then, the cell viability was
calculated using MTT analysis. (B) The rat gastric mucosal cells
(RGM-1, left) and human gastric epithelium cells (GES-1, right)
were cultured with different concentrations (0, 5, 10, 20, 40, 80
and 160 µM) of LIQ for 24 h, followed by MTT analysis to
calculate the cell viability. (C) SGC7901/DDP cells were treated as
indicated for 24 h. Then, the morphology of cells was observed
under a microscope. The colony formation was analyzed. The
migration of cancer cells was determined; and the wound healing
assay was carried out to further evaluate the migration of
DDP-resistant gastric cancer cells treated by LIQ. (D) The number
of clonogenetic cells was quantified. (E) The percentage of
migrated cells is shown. (F) The relative wound healing width was
quantified. Data are presented as the mean ± SEM, *p<0.05,
**p<0.01 and ***p<0.001 versus the
DDP−/LIQ− group. +p<0.05,
++p<0.01 and +++p<0.001 versus the
DDP−/LIQ+ group. |
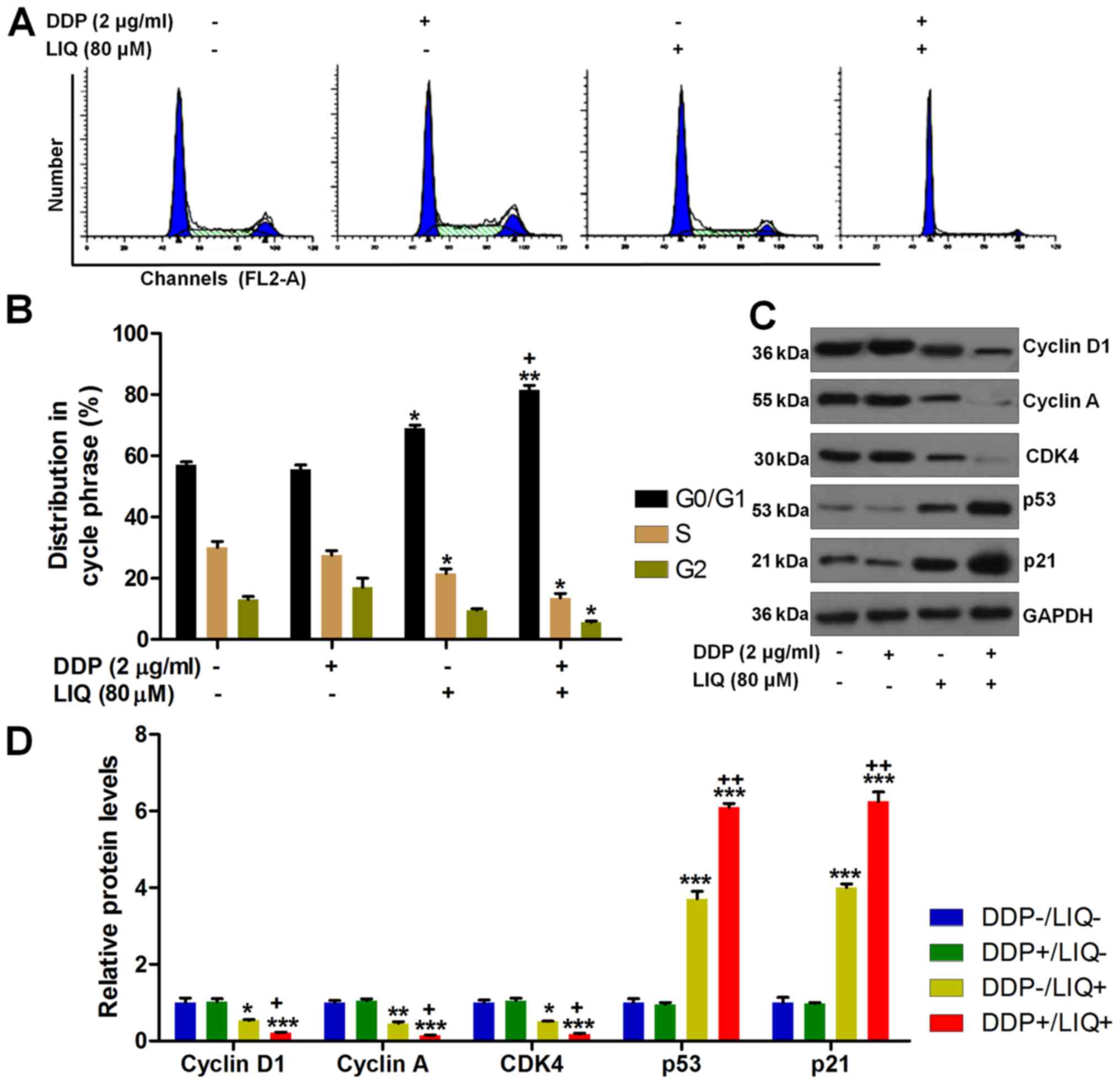
Liquiritin induces G0/G1 cell cycle
arrest in DPP-resistant gastric cancer cells
Cell proliferation has a close relationship with the
distribution of cell cycle phase. Thus, in order to verify if the
growth suppression caused by DDP/LIQ was related to cell cycle
arrest, the role of DDP/LIQ in the cell cycle distribution was
measured using flow cytometry. As shown in Fig. 2A and B, the proportion of
SGC7901/DDP cells at G1/S was significantly increased after DDP/LIQ
co-treatment compared to the LIQ-single treatment. Next, the cell
cycle-associated molecules were calculated through western blot
analysis. The results indicated that DDP and LIQ in combination
markedly reduced cyclin D1, cyclin A and CDK4 expressions, while
p53 and p21 were found to be enhanced, which were comparable to the
single treatment of LIQ (Fig. 2C and
D). Taken together, the findings above indicated that DDP/LIQ
induced G0/G1 cell cycle arrest through modulating the important
signals regarding to DNA damage.
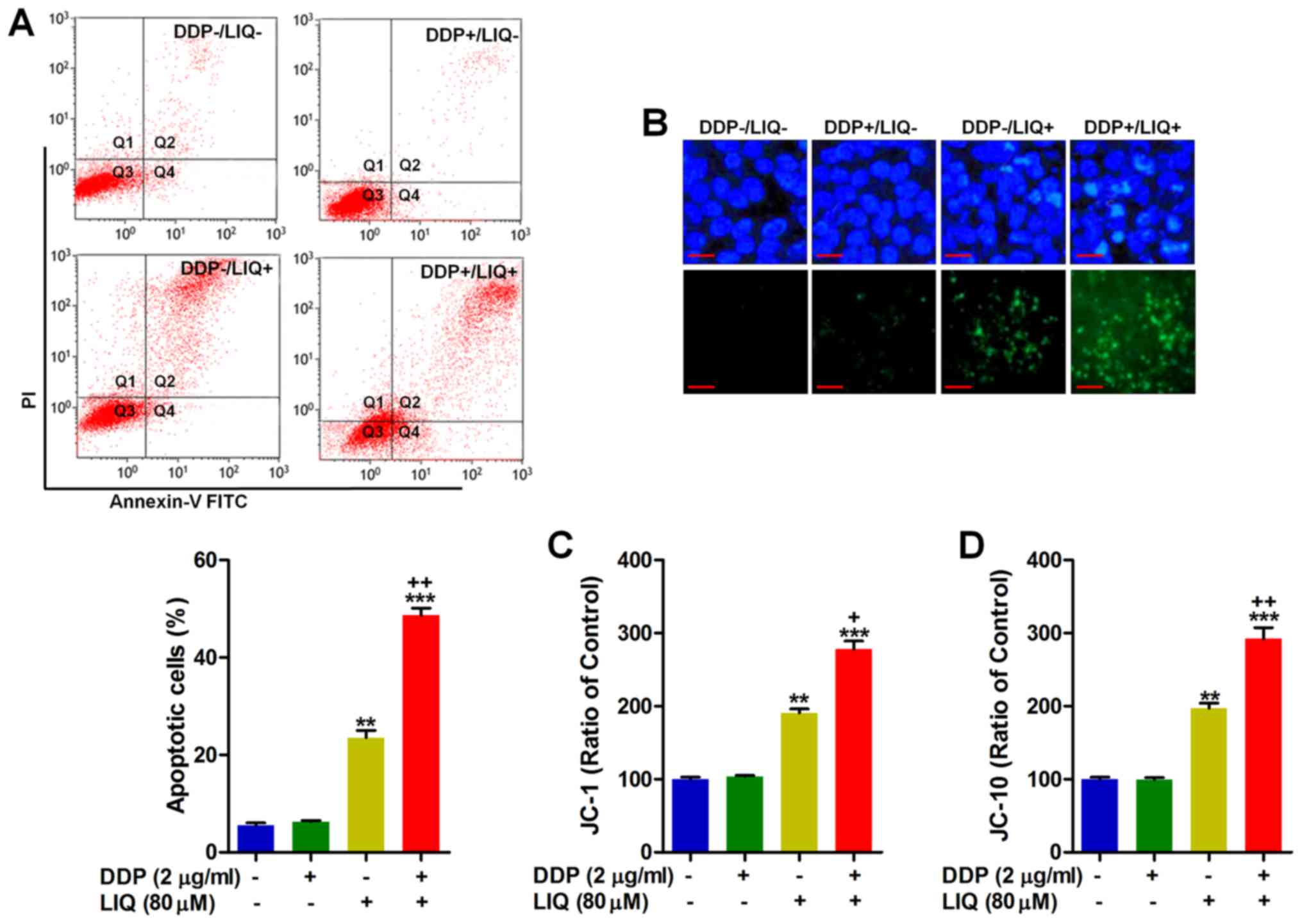
Liquiritin and DDP co-treatment enhances
apoptosis in DDP-resistant gastric cancer cells through promoting
caspase-3 expression
Apoptosis plays an essential role in inducing cell
death, which is widely used as a vital molecular mechanism to
explore new and effective therapies against the progression of
various tumors (18,19). Fig.
3A illustrates that LIQ could effectively induce apoptosis in
SGC7901/DDP cells, and considerably, LIQ combined with DDP further
improved the apoptotic proportion. Similarly, Hoechst 33258
staining and TUNEL staining suggested that co-treatment of DDP and
LIQ could dramatically induce apoptosis in SGC7901/DDP cells
compared to the LIQ group (Fig.
3B). Further, the fluorescent intensities for both J-aggregates
and monomeric forms of JC-1 and JC-10 were markedly enhanced by
DDP/LIQ co-treatment in SGC7901/DDP cells, which further proved
that apoptosis was induced in DDP-resistant gastric cancer cells
(Fig. 3C and D). Together, the
results above indicated that DDP/LIQ could induce apoptosis in
SGC7901/DDP cells, contributing to cell death.
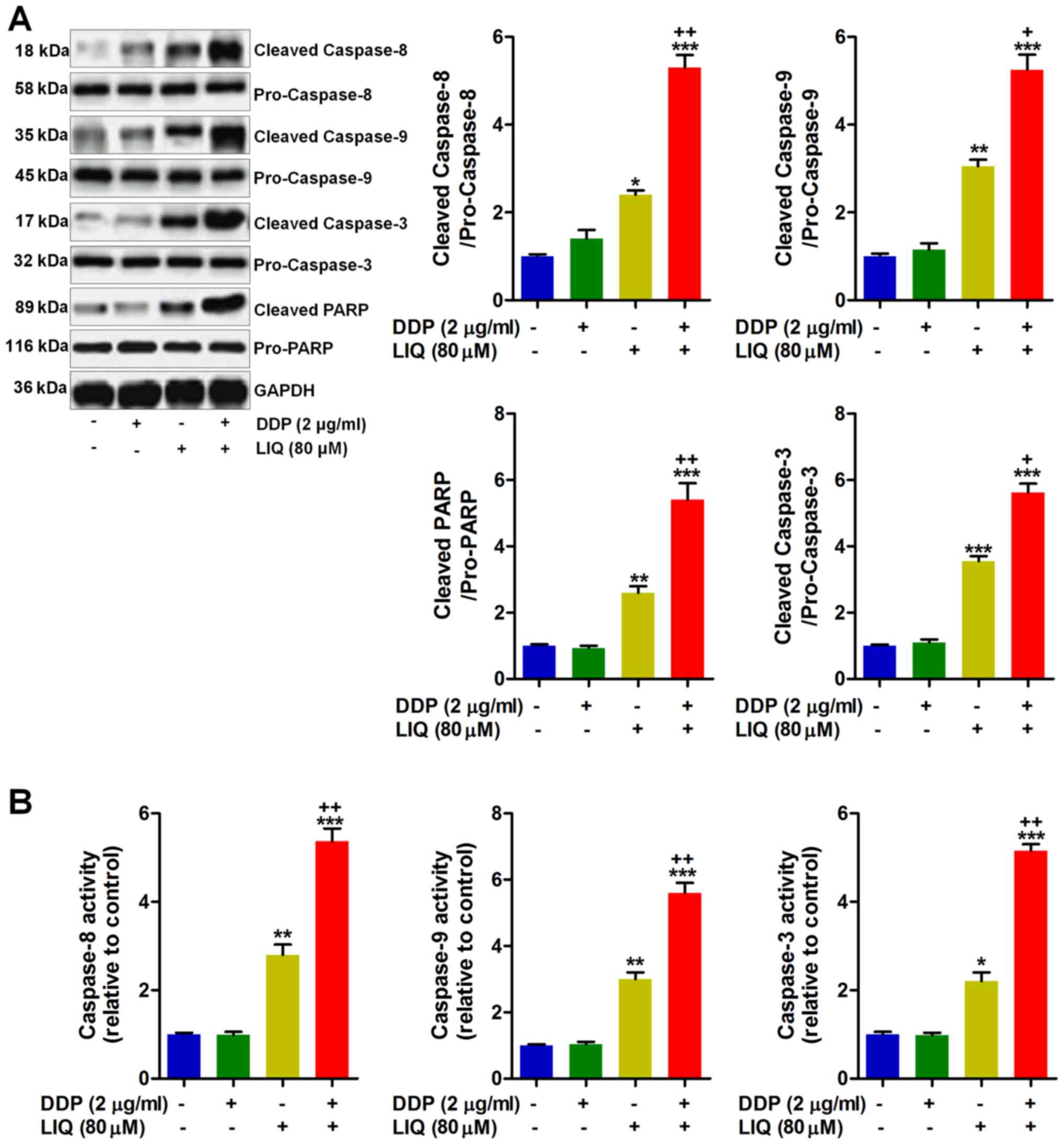
Caspases are important signals involved in apoptosis
induction (20). Thus, here we
attempted to explore how these signals altered in SGC7901/DDP cells
after various treatments. From Fig.
4A, we found that cleaved caspase-8/-9/-3 and PARP were highly
induced by LIQ, and of note, DDP and LIQ in combination showed
stronger effects to induce the expression of these proteins.
Moreover, the activity of caspase-8/-9 and -3 in DDP-resistant
gastric cancer cells was measured. As shown in Fig. 4B, DDP/LIQ co-treatment considerably
enhanced caspase-8/-9 and -3 activiy, respectively, indicating that
LIQ/DDP-induced apoptosis was tightly associated with caspase-3
pathway in SGC7901/DDP cells.
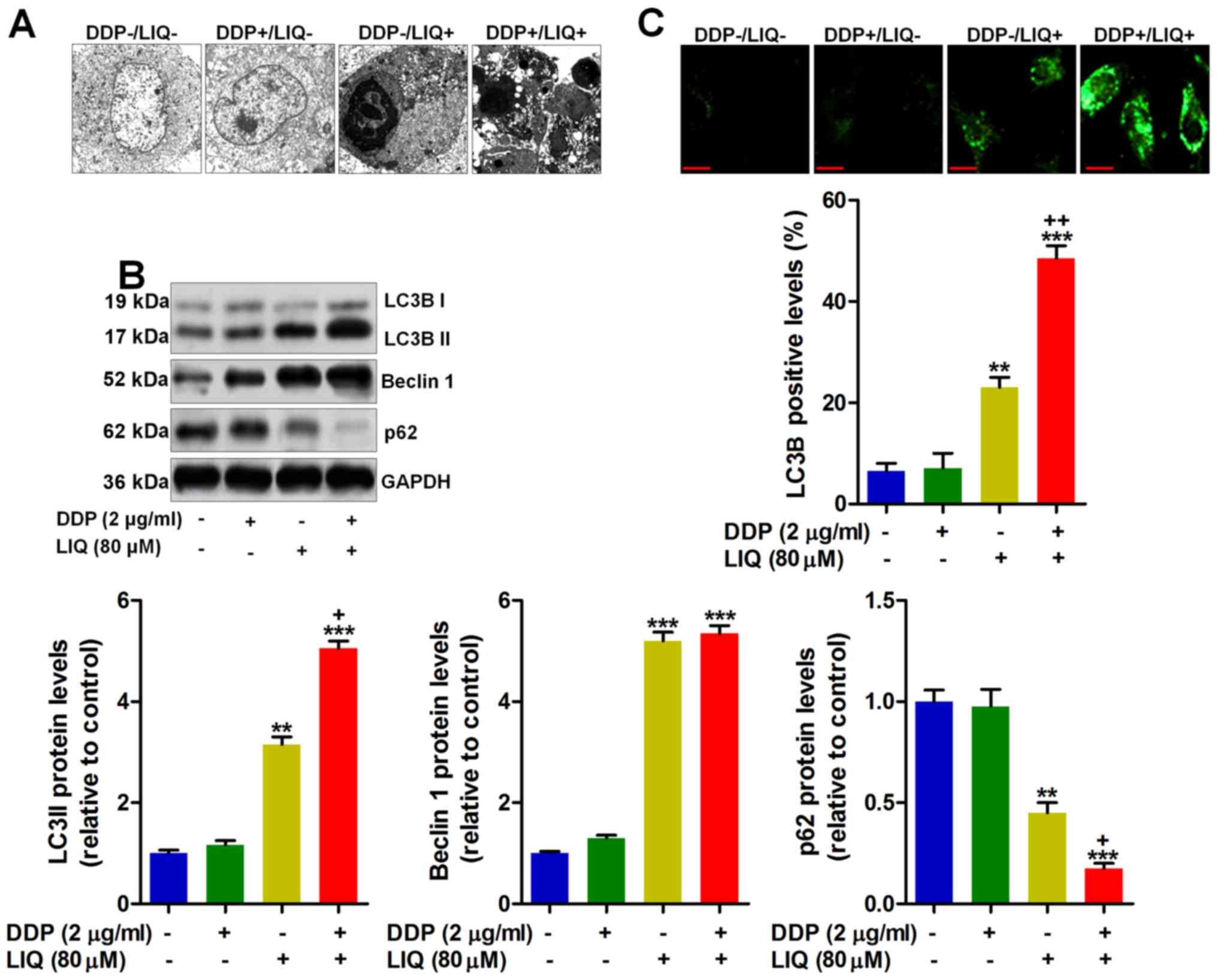
Liquiritin and DDP double therapy
triggers autophagy in DDP-resistant gastric cancer cells
Autophagy is another molecular mechanism by which
the cells are induced to death. Thus, autophagy plays an essential
role in drug exploration for preventing tumor growth (21,22).
As shown in Fig. 5A, the TEM
analysis indicated that the cell structure was severely damaged by
LIQ treatment, which was accelerated due to its combination with
DDP. Following, western blot analysis suggested that
autophagy-related signals of LC3BI/II, and Beclin 1 were highly
induced by LIQ single treatment, while p62 was found to be
suppressed. Notably, DDP/LIQ co-treatment further augmented
LC3BI/II and Beclin 1 expression and reduced p62 expression,
resulting in autophagy in SGC7901/DDP cells (Fig. 5B). Finally, fluorescent analysis
further evidenced that LC3B were highly induced by LIQ/DDP
co-treatment compared to the DDP−/LIQ+ group,
indicating that autophagy could be induced by DDP/LIQ, resulting in
gastric cancer cell death (Fig.
5C).
Liquiritin and DDP co-treatment
suppresses tumor growth of xenograft mice in vivo
In vivo, the SGC7901/DDP cells
(2×105) were subcutaneously inoculated into nude mice.
When tumor size reached to ~50 mm3, mice were randomly
grouped to receive DDP (3 mg/kg) and LIQ (15 mg/kg) or the two in
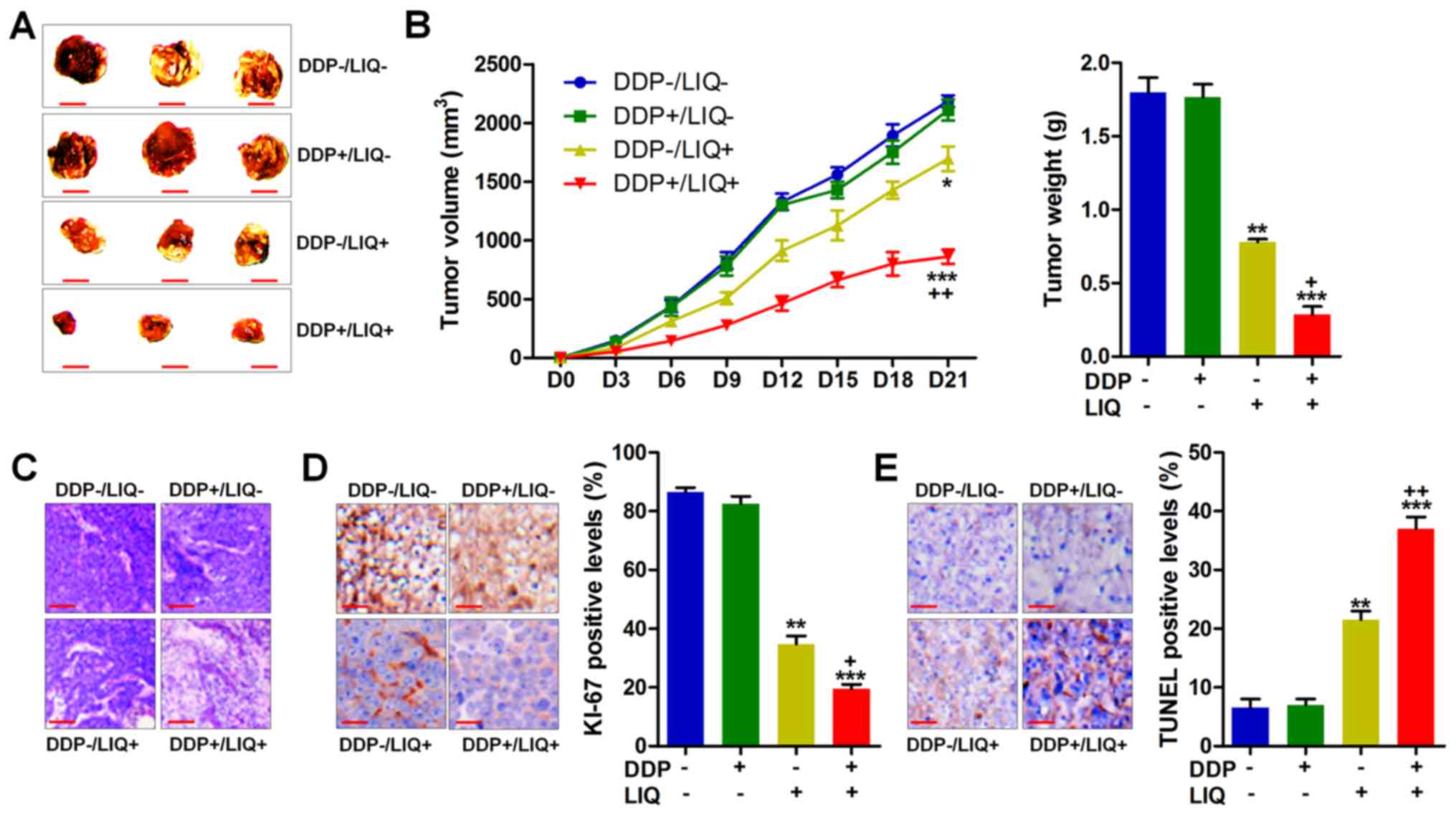
combination for 21 days. As shown in Fig. 6A and B, we found that DDP and LIQ
in combination significantly reduced the size, volume and weight of
tumor compared to LIQ single treatment, suggesting that
co-treatment of LIQ and DDP could reduce the growth of tumor in
vivo. Fig. 6C also indicated
that the number of tumor cells was obviously reduced by DDP and LIQ
double therapy. Finally, the immunohistochemical analysis showed
dramatically downregulated Ki-67 levels and upregulated
TUNEL-positive cells in tumor tissue sections with DDP/LIQ
treatment together, further indicating that DDP/LIQ impeded the
tumor growth and induced apoptosis in vivo (Fig. 6D and E).
Liquiritin and DDP co-treatment induces
apoptosis and autophagy in tumor tissues
The results above elucidated that DDP/LIQ
co-treatment could trigger apoptosis and autophagy in vitro.
Thus, here we attempted to explore if the possible molecular
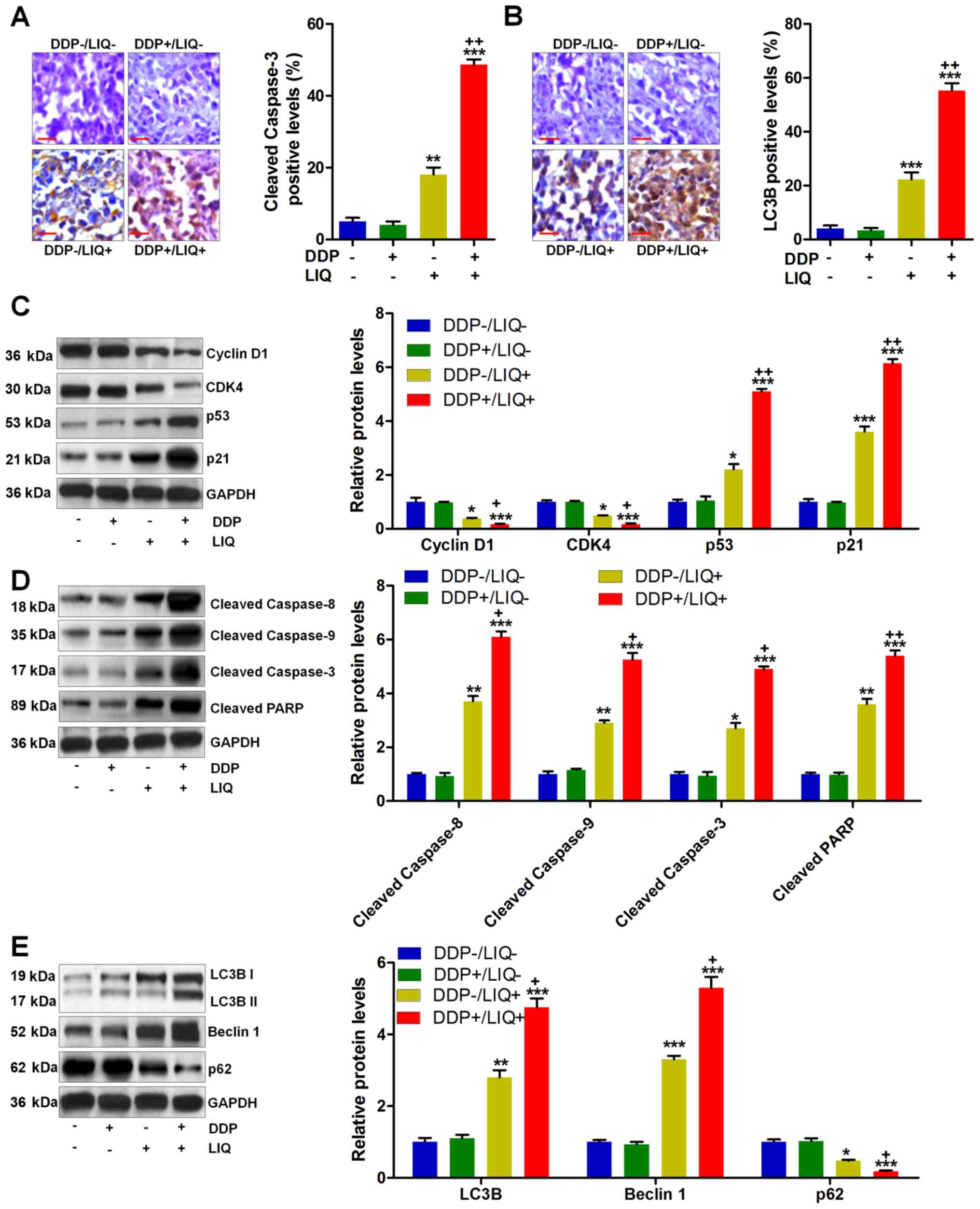
mechanism was involved in vivo. As shown in Fig. 7A and B, cleaved caspase-3- and
LC3B-positive cells were significantly elevated in DPP/LIQ-treated
group of tumors. Furthermore, western blot analysis showed that
high expression of cyclin D1 and CDK4 was reduced by LIQ, which
were further downregulated by DDP/LIQ co-treatment. In contrast,
p53 and p21 expressions were found to be markedly elevated in
DDP/LIQ group, suggesting the suppressive effects of DDP/LIQ on
gastric cancer growth (Fig. 7C).
Also, the expression levels of cleaved caspase-8/-9/-3 and PARP,
contributing to the progression of apoptosis, were dramatically
improved by co-treatment of DDP and LIQ (Fig. 7D). Also, autophagy-related
molecules of LC3B and Beclin 1 were considerably induced by
DDP/LIQ, while p62 was reduced, which were in line with the results
in vitro (Fig. 7E). In
summary, the data above indicated that co-treatment of DDP/LIQ
could induce apoptosis and autophagy in gastric tumor samples in
vivo, performing its role in suppressing gastric tumor
growth.
Discussion
During the process of tumor chemotherapy, one of the
most intractable problems is the occurrence of drug resistance of
cancer cells to chemotherapeutic drugs (8,23,24).
Resistance to chemotherapy is a major obstacle for the effective
treatment of cancers. The mechanism of chemoresistance is still
poorly understood. The development of multidrug resistance is a
crucial problem of therapy failure in gastric cancer, which results
in disease recurrence and metastasis (25,26).
In the clinical practice, a large number of Chinese medicine drugs
have exhibited effective synergism in chemotherapy. The procedure
has been evidenced in numerous studies (27,28).
Recently, liquiritin (LIQ) displayed comprehensive ability to
prevent the progression of tumors, such as the non-small cell lung
cancer (NSCLC) by inducing apoptosis (29). Though LIQ has been reported to have
anticancer ability, how it suppressed cancer development and the
underlying molecular mechanisms are not well known. Thus, further
study is still required to fully explain its bioactivities against
different types of cancer, including gastric carcinoma. Modern
pharmacological studies have indicated that application of two
drugs in combination could suppress the growth, proliferation,
migration and invasion of various tumor cells, induce apoptosis and
autophagy of tumor cells and impede the role of tumor-promoting
substances on the potential tumor cells (30–32).
In order to further explore the role of LIQ in
preventing gastric cancer, gastric cancer cells of SGC7901 with DDP
resistance were used in our study. SGC7901/DDP cells show
resistance to a large number of chemotherapeutic drugs (33,34).
We combined LIQ and DDP to prevent SGC7901/DDP cells. The results
indicated that LIQ could enhance the killing ability of DDP on
SGC7901/DDP cells and promote the effects of DDP on the induction
of apoptosis and autophagy in SGC7901/DDP cells. Further, the
cytotoxicity of LIQ was measured. MTT analysis indicated that there
was no significant difference between the Con and LIQ-treated
groups, indicating its safety for application within our conditions
(14,15). In vivo, LIQ and DDP in
combination showed strongly suppressive effects on the growth of
SGC7901/DDP xenograft tumor in nude mice. The results above
suggested that LIQ could enhance the sensitivity of SGC7901/DDP
cells to DDP treatment, reducing the drug resistance.
Cancer is characterized by abnormal cell growth,
which evolves, at least partly by over-riding the regulation of
cellular proliferation (35).
Cyclins and cyclin-dependent kinases (CDKs) are tightly included in
the process of cell cycle in tumor cells. CDKs are important
modulators of cell cycle machinery, influencing the progression of
cell cycle from one phase to the next (36,37).
Unusual cyclins and CDK activity leads to dysregulation of
programmed cell death or apoptotic development, which contributes
to selective growth advantage for tumor cells. Dys-regulated cell
cycle process is an essential factor during development and
progression of cancer (38,39).
Controlling the process of cell cycle in tumor cells is an
effective therapeutic strategy to inhibit tumor growth and
development, and cell cycle regulators are deregulated in most
common malignancies (40,41). Regulating the cell cycle at G1
checkpoint is complex, including multiple molecular processes. P53
is a frequent target for mutation in various human tumors (42). Additionally, p53 could respond to
different stresses, such as the cell cycle arrest, DNA repair, and
apoptosis (43). As reported
previously, p21 plays an essential role in diminishing the G1 cell
cycle arrest in different tumor cells. P53 inhibits the cell cycle
progression by triggering CDK suppressors, including p21 (44). In our study, we found that G1/G0
cell cycle arrest was induced by LIQ, which was further enhanced by
LIQ and DDP in combination. Also, consistently, cyclin D1, cyclin A
and CDK4 were highly induced by LIQ/DDP co-treatment, while p53 and
p21 were reduced, indicating that LIQ potentiated the effects of
DDP on the suppression of SGC7901/DDP cells.
Apoptosis is a morphologically obvious form of
programmed cell death, which plays an essential role in regulating
homeostasis and development of organisms. In addition, the
progression of numerous diseases is linked with apoptosis,
including different cancers (45,46).
Inducing apoptosis is an important molecular mechanism by which a
variety of anticancer drugs were explored (47). In our present study, we found that
apoptosis was highly induced by LIQ and DDP co-treatment through
flow cytometry, Hoechst 33258 and TUNEL staining. Caspase is a
family of cysteine proteases, modulating apoptosis (48). In response to cell death stimuli,
Bax could migrate to mitochondria to induce a cascade of
mitochondrial permeability transition pore-related events,
resulting in a downregulation in the mitochondrial membrane
potential and apoptosis (49). In
the end, caspase-9 is activated, promoting the cleavage of
caspase-3 and PARP, which contributes to apoptosis and ultimately
cell death (50). Caspase-8
participates in the extrinsic apoptosis signaling, which is another
mechanism to induce apoptosis (51). In our study, we found that cleaved
caspase-8/-9/-3 and PARP were expressed highly in the group of
LIQ/DDP, revealing that LIQ might enhance the role of DDP in
inducing apoptosis in SGC7901/DDP cells, dependent on both
intrinsic and extrinsic signaling pathways.
Autophagy is a homeostatic and catabolic process,
which is crucial for the packaging and degradation of cytoplasmic
proteins and organelles (52).
Therefore, autophagy is necessary to maintain genomic stability and
overall cell survival (21).
Microtubule-associated protein light chain 3 (LC3) is localized and
accumulated on the autophagosome, which is well known as an
important hallmark of autophagy (53). LC3B transforms from LC3B-I to
LC3B-II, resulting in autophagosome formation (54). Beclin 1 is another important marker
of autophagy. Beclin 1 forms a complex with type III
phosphatidylinositol that allows the nucleation of the autophagic
vesicles (55). p62 has a close
relationship with autophagy, possessing an LC3-interacting region
(56). In our study, LIQ improved
the effects of DPP to trigger autophagy in SGC7901/DDP cells,
evidenced by the considerable increasing of LC3B, Beclin 1 and
decreasing of p62. In the present study in vivo, we found
that LIQ/DDP co-treatment significantly downregulated the
SGC7901/DDP tumor growth, proved by limited tumor volume and tumor
weight. Furthermore, the western blot and histochemistry analysis
further ensured the suppression of cyclin D1 and CDK4, improvement
of p53, p21, cleaved caspase-8/-9/-3 and PARP, as well as elevation
of LC3B-II and Beclin 1 after treatment of LIQ/DDP in animals,
indicating that LIQ might be effective in blunting tumor growth and
inducing apoptosis and autophagy.
In conclusion, the results above indicated that LIQ
could enhance the pro-apoptotic effect of DDP on human gastric
cancer SGC7901/DDP cells through arresting G0/G1 phase cycle,
inducing apoptosis and autophagy. In the xenograft models, LIQ/DDP
administration showed obvious anticancer activity with reduced
tumor volume and weight. Thus, LIQ could be considered as
synergistic drug with DDP to prevent gastric cancer.
Acknowledgments
This study was supported by the Science and
Technology Planning Project: ‘The relationship between difference
of different stem cell subsets and prognosis in liver metastasis of
colorectal cancer’, Jilin, P.R. China (20170414034GH).
References
|
1
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: Defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sakuramoto S, Sasako M, Yamaguchi T,
Kinoshita T, Fujii M, Nashimoto A, Furukawa H, Nakajima T, Ohashi
Y, Imamura H, et al ACTS-GC Group: Adjuvant chemotherapy for
gastric cancer with S-1, an oral fluoropyrimidine. N Engl J Med.
357:1810–1820. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brambilla G and Martelli A: Genotoxic and
carcinogenic risk to humans of drug-nitrite interaction products.
Mutat Res. 635:17–52. 2007. View Article : Google Scholar
|
|
5
|
Van Cutsem E, Moiseyenko VM, Tjulandin S,
Majlis A, Constenla M, Boni C, Rodrigues A, Fodor M, Chao Y, Voznyi
E, et al V325 Study Group: Phase III study of docetaxel and
cisplatin plus fluorouracil compared with cisplatin and
fluoro-uracil as first-line therapy for advanced gastric cancer: A
report of the V325 Study Group. J Clin Oncol. 24:4991–4997. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsutani Y, Yoshida K, Sanada Y, Wada Y,
Konishi K, Fukushima M and Okada M: Decreased orotate
phosphoribo-syltransferase activity produces 5-fluorouracil
resistance in a human gastric cancer cell line. Oncol Rep.
20:1545–1551. 2008.PubMed/NCBI
|
|
7
|
Zheng LH, Bao YL, Wu Y, Yu CL, Meng X and
Li YX: Cantharidin reverses multidrug resistance of human hepatoma
HepG2/ADM cells via down-regulation of P-glycoprotein expression.
Cancer Lett. 272:102–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kang YK, Kang WK, Shin DB, Chen J, Xiong
J, Wang J, Lichinitser M, Guan Z, Khasanov R, Zheng L, et al:
Capecitabine/cisplatin versus 5-fluorouracil/cisplatin as
first-line therapy in patients with advanced gastric cancer: A
randomised phase III noninferiority trial. Ann Oncol. 20:666–673.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rennicke A, Voigt W, Mueller T, Fruehauf
A, Schmoll HJ, Beyer C and Dempke W: Resistance mechanisms
following cisplatin and oxaliplatin treatment of the human
teratocarcinoma cell line 2102EP. Anticancer Res. 25A:1147–1155.
2005.
|
|
10
|
Tian ML, Yan HY and Row KH: Simultaneous
extraction and separation of liquiritin, glycyrrhizic acid, and
glabridin from licorice root with analytical and preparative
chromatography. Biotechnol Bioprocess Eng; BBE. 13:671–676. 2008.
View Article : Google Scholar
|
|
11
|
Lee JY, Lee JH, Park JH, Kim SY, Choi JY,
Lee SH, Kim YS, Kang SS, Jang EC and Han Y: Liquiritigenin, a
licorice flavonoid, helps mice resist disseminated candidiasis due
to Candida albicans by Th1 immune response, whereas liquiritin, its
glycoside form, does not. Int Immunopharmacol. 9:632–638. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang W, Hu X, Zhao Z, Liu P, Hu Y, Zhou J,
Zhou D, Wang Z, Guo D and Guo H: Antidepressant-like effects of
liquiritin and isoliquiritin from Glycyrrhiza uralensis in the
forced swimming test and tail suspension test in mice. Prog
Neuropsychopharmacol Biol Psychiatry. 32:1179–1184. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong S, Inoue A, Zhu Y, Tanji M and Kiyama
R: Activation of rapid signaling pathways and the subsequent
transcriptional regulation for the proliferation of breast cancer
MCF-7 cells by the treatment with an extract of Glycyrrhiza glabra
root. Food Chem Toxicol. 45:2470–2478. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Aboue A and Al-Shawi AAA: Liquiritin (LG),
isolated and identified from Radix Glycyrrhizae (RG), inhibits the
proliferation and induces apoptosis of human gastric cancer cells
(MGC-803). Int J Curr Microbiol Appl Sci. 3:89–98. 2014.
|
|
15
|
Gao YX, Cheng BF, Lian JJ, Guo D-D, Qin
J-W, Zhang Y-B, Yang H-J, Wang M, Wang L and Feng Z-W: Liquiritin,
a flavone compound from licorice, inhibits IL-1β-induced
inflammatory responses in SW982 human synovial cells. J Funct
Foods. 33:142–148. 2017. View Article : Google Scholar
|
|
16
|
Selvakumaran M, Pisarcik DA, Bao R, Yeung
AT and Hamilton TC: Enhanced cisplatin cytotoxicity by disturbing
the nucleotide excision repair pathway in ovarian cancer cell
lines. Cancer Res. 63:1311–1316. 2003.PubMed/NCBI
|
|
17
|
Kim YW, Ki SH, Lee JR, Lee SJ, Kim CW, Kim
SC and Kim SG: Liquiritigenin, an aglycone of liquiritin in
Glycyrrhizae radix, prevents acute liver injuries in rats induced
by acetaminophen with or without buthionine sulfoximine. Chem Biol
Interact. 161:125–138. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vanella L, Di Giacomo C, Acquaviva R,
Barbagallo I, Cardile V, Kim DH, Abraham NG and Sorrenti V:
Apoptotic markers in a prostate cancer cell line: Effect of ellagic
acid. Oncol Rep. 30:2804–2810. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang XH, Zheng X, Cao JG, Xiang HL, Liu F
and Lv Y: 8-Bromo-7-methoxychrysin-induced apoptosis of
hepatocellular carcinoma cells involves ROS and JNK. World J
Gastroenterol. 16:3385–3393. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gyrd-Hansen M and Meier P: IAPs: From
caspase inhibitors to modulators of NF-kappaB, inflammation and
cancer. Nat Rev Cancer. 10:561–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang ZJ, Chee CE, Huang S and Sinicrope F:
Autophagy modulation for cancer therapy. Cancer Biol Ther.
11:169–176. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhai B, Hu F, Jiang X, Xu J, Zhao D, Liu
B, Pan S, Dong X, Tan G, Wei Z, et al: Inhibition of Akt reverses
the acquired resistance to sorafenib by switching protective
autophagy to autophagic cell death in hepatocellular carcinoma. Mol
Cancer Ther. 13:1589–1598. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao D, Jiang Y, Dong X, Liu Z, Qu B,
Zhang Y, Ma N and Han Q: Arsenic trioxide reduces drug resistance
to adriamycin in leukemic K562/A02 cells via multiple mechanisms.
Biomed Pharmacother. 65:354–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao Y, You H, Liu F, An H, Shi Y, Yu Q
and Fan D: Differentially expressed gene profiles between multidrug
resistant gastric adenocarcinoma cells and their parental cells.
Cancer Lett. 185:211–218. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang XW, Bu P, Liu L, Zhang XZ and Li J:
Overexpression of long non-coding RNA PVT1 in gastric cancer cells
promotes the development of multidrug resistance. Biochem Biophys
Res Commun. 462:227–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang S, Chen M, Shen Y, Shen W, Guo H,
Gao Q and Zou X: Inhibition of activated Stat3 reverses drug
resistance to chemotherapeutic agents in gastric cancer cells.
Cancer Lett. 315:198–205. 2012. View Article : Google Scholar
|
|
27
|
Ganta S and Amiji M: Coadministration of
Paclitaxel and curcumin in nanoemulsion formulations to overcome
multidrug resistance in tumor cells. Mol Pharm. 6:928–939. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim MK, Choi HS, Cho SG, Shin YC and Ko
SG: Rubus coreanus Miquel extract causes apoptosis of
doxorubicin-resistant NCI/ADR-RES ovarian cancer cells via JNK
phosphorylation. Mol Med Rep. 13:4065–4072. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou Y and Ho WS: Combination of
liquiritin, isoliquiritin and isoliquirigenin induce apoptotic cell
death through upregulating p53 and p21 in the A549 non-small cell
lung cancer cells. Oncol Rep. 31:298–304. 2014. View Article : Google Scholar
|
|
30
|
Matsumoto H, Inaba H, Kishi M, Tominaga S,
Hirayama M and Tsuda T: Orally administered delphinidin
3-rutinoside and cyanidin 3-rutinoside are directly absorbed in
rats and humans and appear in the blood as the intact forms. J
Agric Food Chem. 49:1546–1551. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hagiwara Y, Kasukabe T, Kaneko Y, Niitsu N
and Okabe-Kado J: Ellagic acid, a natural polyphenolic compound,
induces apoptosis and potentiates retinoic acid-induced
differentiation of human leukemia HL-60 cells. Int J Hematol.
92:136–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang ZC and Ma J: Actein enhances TRAIL
effects on suppressing gastric cancer progression by activating
p53/Caspase-3 signaling. Biochem Biophys Res Commun. Nov
30–2016.Epub ahead of print. View Article : Google Scholar
|
|
33
|
Kumar A, Wessels D, Daniels KJ, Alexander
H, Alexander S and Soll DR: Sphingosine-1-phosphate plays a role in
the suppression of lateral pseudopod formation during Dictyostelium
discoideum cell migration and chemotaxis. Cell Motil Cytoskeleton.
59:227–241. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li G, Yang F, Gu S, Li Z and Xue M:
MicroRNA-101 induces apoptosis in cisplatin-resistant gastric
cancer cells by targeting VEGF-C. Mol Med Rep. 13:572–578. 2016.
View Article : Google Scholar
|
|
35
|
Schwartz GK and Shah MA: Targeting the
cell cycle: A new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aggarwal P, Vaites LP, Kim JK, Mellert H,
Gurung B, Nakagawa H, Herlyn M, Hua X, Rustgi AK, McMahon SB, et
al: Nuclear cyclin D1/CDK4 kinase regulates CUL4 expression and
triggers neoplastic growth via activation of the PRMT5
methyl-transferase. Cancer Cell. 18:329–340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bockstaele L, Bisteau X, Paternot S and
Roger PP: Differential regulation of cyclin-dependent kinase 4
(CDK4) and CDK6, evidence that CDK4 might not be activated by CDK7,
and design of a CDK6 activating mutation. Mol Cell Biol.
29:4188–4200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Perisanidis C, Perisanidis B, Wrba F,
Brandstetter A, El Gazzar S, Papadogeorgakis N, Seemann R, Ewers R,
Kyzas PA and Filipits M: Evaluation of immunohistochemical
expression of p53, p21, p27 cyclin D1, and Ki67 in oral and
oropharyngeal squamous cell carcinoma. J Oral Pathol Med. 41:40–46.
2012. View Article : Google Scholar
|
|
39
|
Kim YA, Lee WH, Choi TH, Rhee SH, Park KY
and Choi YH: Involvement of p21WAF1/CIP1, pRB, Bax and NF-kappaB in
induction of growth arrest and apoptosis by resveratrol in human
lung carcinoma A549 cells. Int J Oncol. 23:1143–1149.
2003.PubMed/NCBI
|
|
40
|
Liu P, Wang X, Hu C and Hu T: Inhibition
of proliferation and induction of apoptosis by trimethoxyl stilbene
(TMS) in a lung cancer cell line. Asian Pac J Cancer Prev.
12:2263–2269. 2011.
|
|
41
|
Le NT and Richardson DR: The role of iron
in cell cycle progression and the proliferation of neoplastic
cells. Biochim Biophys Acta. 1603:31–46. 2002.PubMed/NCBI
|
|
42
|
Alao JP: The regulation of cyclin D1
degradation: Roles in cancer development and the potential for
therapeutic invention. Mol Cancer. 6:242007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Horn HF and Vousden KH: Coping with
stress: Multiple ways to activate p53. Oncogene. 26:1306–1316.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chatterjee SJ, Datar R, Youssefzadeh D,
George B, Goebell PJ, Stein JP, Young L, Shi SR, Gee C, Groshen S,
et al: Combined effects of p53, p21, and pRb expression in the
progression of bladder transitional cell carcinoma. J Clin Oncol.
22:1007–1013. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen MB, Wu XY, Gu JH, Guo QT, Shen WX and
Lu PH: Activation of AMP-activated protein kinase contributes to
doxorubicin-induced cell death and apoptosis in cultured myocardial
H9c2 cells. Cell Biochem Biophys. 60:311–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Circu ML and Aw TY: Glutathione and
modulation of cell apoptosis. Biochim Biophys Acta. 1823:1767–1777.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Andersen JL and Kornbluth S: The tangled
circuitry of metabolism and apoptosis. Mol Cell. 49:399–410. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Suzuki Y, Nakabayashi Y, Nakata K, Reed JC
and Takahashi R: X-linked inhibitor of apoptosis protein (XIAP)
inhibits caspase-3 and -7 in distinct modes. J Biol Chem.
276:27058–27063. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xiong Y, Lu QJ, Zhao J and Wu GY:
Metformin inhibits growth of hepatocellular carcinoma cells by
inducing apoptosis via mitochondrion-mediated pathway. Asian Pac J
Cancer Prev. 13:3275–3279. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hasan TN, B LG, Shafi G, Al-Hazzani AA and
Alshatwi AA: Anti-proliferative effects of organic extracts from
root bark of Juglans Regia L. (RBJR) on MDA-MB-231 human breast
cancer cells: Role of Bcl-2/Bax, caspases and Tp53. Asian Pac J
Cancer Prev. 12:525–530. 2011.PubMed/NCBI
|
|
51
|
Sträter J, Herter I, Merkel G, Hinz U,
Weitz J and Möller P: Expression and prognostic significance of
APAF-1, caspase-8 and caspase-9 in stage II/III colon carcinoma:
Caspase-8 and caspase-9 is associated with poor prognosis. Int J
Cancer. 127:873–880. 2010.
|
|
52
|
Schleicher SM, Moretti L, Varki V and Lu
B: Progress in the unraveling of the endoplasmic reticulum
stress/autophagy pathway and cancer: Implications for future
therapeutic approaches. Drug Resist Updat. 13:79–86. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen ZH, Lam HC, Jin Y, Kim HP, Cao J, Lee
SJ, Ifedigbo E, Parameswaran H, Ryter SW and Choi AM: Autophagy
protein microtubule-associated protein 1 light chain-3B (LC3B)
activates extrinsic apoptosis during cigarette smoke-induced
emphysema. Proc Natl Acad Sci USA. 107:18880–18885. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Klionsky DJ, Abeliovich H, Agostinis P,
Agrawal DK, Aliev G, Askew DS, Baba M, Baehrecke EH, Bahr BA,
Ballabio A, et al: Guidelines for the use and interpretation of
assays for monitoring autophagy in higher eukaryotes. Autophagy.
4:151–175. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yue Z, Jin S, Yang C, Levine AJ and Heintz
N: Beclin 1, an autophagy gene essential for early embryonic
development, is a haploinsufficient tumor suppressor. Proc Natl
Acad Sci USA. 100:15077–15082. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mizushima N and Yoshimori T: How to
interpret LC3 immuno-blotting. Autophagy. 3:542–545. 2007.
View Article : Google Scholar : PubMed/NCBI
|