Introduction
Glioblastoma (GBM) is one of the most common and
fatal tumors of the central nervous system (1–3). In spite
of the therapeutic advances in strategies for treatment, the poor
prognosis of patients diagnosed with GBM has not improved and the
median survival is <1 year (2–4). One of
the major reasons leading to the poor prognosis of patients is
recurrence as a result of the diffuse infiltration of tumor cells
into the surrounding brain tissue. Therefore, understanding the
molecular mechanisms of astrocytic tumor cell invasion in the brain
is essential for developing effective strategies for the control of
GBM.
MicroRNAs (miRNAs/miRs) belong to a class of small
non-coding single-stranded RNAs of 18–23 nucleotides, which take
part in the control of gene expression by destabilizing target mRNA
or by repressing protein translation. It is now clear that miRNAs
are involved in regulating important cellular functions, including
the stress response, death, differentiation, apoptosis, invasion,
proliferation and development (5,6). Moreover,
emerging evidence has shown that the dysfunction or deregulation of
miRNAs is involved in the processes of cancer initiation,
progression and metastasis (7,8). The
aberrant expression of miR-133b has been reported to be
downregulated in certain types of cancer, including cervical
carcinoma, lung cancer, renal cell carcinoma (9), colon cancer (10), osteosarcoma (11) and prostate cancer (1,12).
Furthermore, miR-133b has been suggested to function as a tumor
suppressor in specific types of cancer by the direct targeting of
certain oncogenes (13). However, the
role of miR-133b in GBM is not clear.
The aim of the present study was to investigate the
expression of miRNA-133b on the migration and invasion of GBM
cells.
Materials and methods
Tumor and normal tissue samples
Normal brain tissue and human GBM samples were
obtained from the Department of Neurosurgery at the Tumor Hospital
(Harbin Medical University Harbin, Heilongjiang, China). None of
the patients had received radiotherapy or chemotherapy prior to
surgery. Samples were snap-frozen in liquid nitrogen and stored at
−80°C until RNA extraction. Study approval was obtained from the
Ethics Committee of The Tumor Hospital, Harbin Medical University
and all patients involved provided written informed consent.
Cell culture and transfection
Two human glioma cell lines, U87 and U251, purchased
from Cell Bank of the Chinese Academy of Sciences (Shanghai,
China), were included in this study. The cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, UT, USA)
containing 100 mg/ml streptomycin (Beyotime Institute of
Biotechnology, Haimen, China), 100 U/ml penicillin ((Beyotime
Institute of Biotechnology) and 10% fetal bovine serum (FBS;
Hyclone) in a humidified atmosphere containing 5% CO2 at
37°C. On the day prior to transfection, the cells were seeded onto
6-well plates (5×105 cells/well) and inoculated in
complete medium without antibiotics (2 ml/well).
Transfections with mature miR-133b mimics, negative
control (NC) and luciferase reporter plasmid were carried out using
50 nm Lipofectamine 2000 (Shanghai GenePharma Co., Ltd., Shanghai,
China) according to the manufacturer's instructions. The sequences
were as follows: hsa-miR-133b mimic, 5′-UUUGGUCCCCUUCAACCAGCUA-3′;
and NC, 5′-UUCUCCGAACGUGUCACGUTT-3.
Wound healing assay
The U87 and U251 cells were plated onto 96-well
plates and treated with miR-133b mimics and NC for 24 h. A 100 μl
plastic pipette tip was then dragged across the plate to create a
wound line that is a cell-free area. Images (magnification, ×40) of
wound lines for each treatment were taken at 0 and 24 h using a
digital camera (Nikon, Tokyo, Japan). The distance of wound closure
(compared with the control at 0 h) was measured. These experiments
were repeated three times.
Transwell migration and invasion
assays
A Transwell invasion assay was performed using a
24-well Transwell chamber with polycarbonate membranes with 8-μm
pores (Corning-Costar Inc., Corning, NY, USA) coated with Matrigel
(BD Biosciences, Franklin Lakes, NJ, USA). Transfected cells
(miR-133 mimics or NC) were loaded onto the upper filter (0.2 ml)
at a density of 5×105 cells/ml of serum-free medium.
DMEM/F12 medium containing 15% FBS (0.5 ml) was added to the lower
chamber. The cells were incubated for 24 h at 37°C in a humidified
incubator containing 5% CO2. A cotton swab was used to
scrape off cells and Matrigel on the upper side of the Transwell
insert filter. Invading cells were fixed with 4% paraformaldehyde
(Beyotime Institute of Biotechnology, Haimen, China) for 10 min,
stained with hematoxylin for 2 min, rinsed in running tap water for
10 min, differentiated with 0.3% acid alcohol for 1 min, rinsed in
running tap water for 10 min, stained with eosin for 2 min, and
counted by capturing images in three independent fields for each
Transwell insert filter (magnification, ×200). The numbers of
invading cells were averaged. A migration assay was performed
following the same procedure, except that the cells were incubated
for 12 h and the polycarbonate filters were not coated with
Matrigel. These experiments were repeated three times.
Western blotting
The cells were washed twice with phosphate-buffered
saline and lysed directly in lysis buffer (1 mmol/l PMSF, 1 mmol/l
EDTA, 40 mmol/l Tris-HCl, 100 mmol/l NaVO3, 150 mmol/l KCl and 1%
Triton X-100) on ice for 15 min. Subsequent to centrifugation
(12,000 × g for 20 min at 4°C), the Bradford protein assay kit
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to measure
the protein concentrations of the lysates. Equal amounts of
proteins were separated on 10% SDS-PAGE gels and then
electrotransferred onto a polyvinylidene difluoride membrane. The
membranes were blocked in 5% skimmed milk-Tris-buffered saline plus
Tween-20 solution and incubated with rabbit MMP14 monoclonal
antibody (1:1,000; Abcam, Cambridge, UK) or rabbit polyclonal
β-actin antibody (1:1,000; Bioss, Beijing, China). Following
incubation with peroxidase-conjugated AffiniPure goat anti-rabbit
IgG (1:2,000; ZSGB-BIO, Beijing, China), protein bands were
detected with Fujifilm Las-4000 (Fujifilm, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA). RNA (~1 μg) was
reverse transcribed using the miR-133b qPCR Quantitation kit
(Shanghai GenePharma Co., Ltd.). The primers of miR-133b and U6
were synthesized by Shanghai Genepharma Co., Ltd., and RT-qPCR was
performed on a 7500 Fast Real Time PCR System (Applied Biosystems,
Foster City, CA, USA) using SYBR Green PCR Master Mix (Applied
Biosystems). The reaction conditions were 95°C for 10 min, followed
by 40 cycles of 95°C for 30 sec and 60°C for 1 min. The mRNA
expression levels of miR-133b were normalized to U6 using the
standard ΔΔCt method. Each assay was performed in triplicate and
all experiments were repeated at least three times.
miRNA target prediction and luciferase
reporter assay
The following target prediction software was used to
identify miRNA-133b that could potentially bind MMP14: Pictar
(http://www.pictar.mdc-berlin.de/),
TargetScan (http://www.targetscan.org/), miRanda (http://www.microrna.org/) and miRWalk (http://www.umm.uni-heidelberg.de/). The U87 and
U251 cells were seeded in 24-well plates. One day later, the cells
were co-transfected with luciferase vectors, either the wild-type
MMP14 3′-untranslated region (3′UTR) reporter plasmid or mutated
MMP14 3′UTR reporter plasmid together with miR-133b mimics or
miRNA-NC, and the Renilla control. After 48 h, luciferase activity
was measured using the Dual-Luciferase Reporter Assay System
(Promega Corporation, Madison, WI, USA) and normalized to Renilla
luciferase activity.
Statistical analysis
SPSS statistical software, version 19.0 (IBM,
Armonk, NY, USA) was used for statistical analysis. The expression
of miR-133b in the tissue samples was analyzed using the
Mann-Whitney U test for comparing the results of two groups. Other
data were expressed as the mean ± standard deviation, and a
two-sided Student's t-test was used for the statistical analysis.
P<0.05 was used to indicate a statistically significant
difference.
Results
Expression of miR-133b is lower in GBM
specimens than normal brain tissues
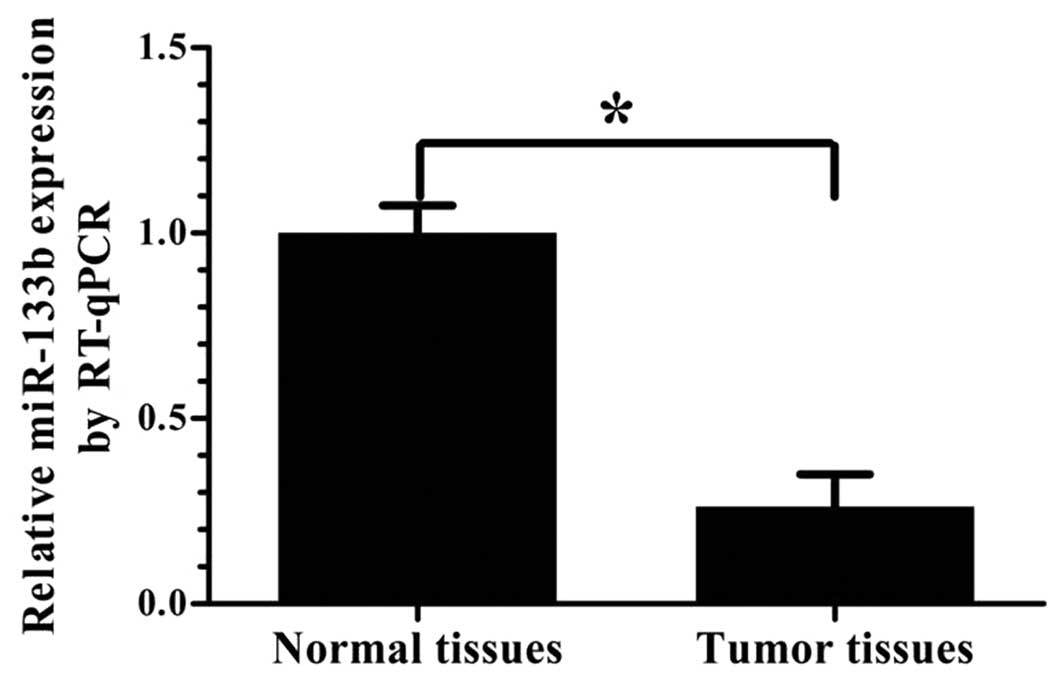
The expression levels of miR-133b were analyzed by
RT-qPCR in 21 GBM specimens and 9 normal brain tissue samples. As
shown in Fig. 1, the expression of
miR-133b was significantly lower in the GBM specimens than in the
normal brain tissues (P<0.05).
Expression of miR-133b following
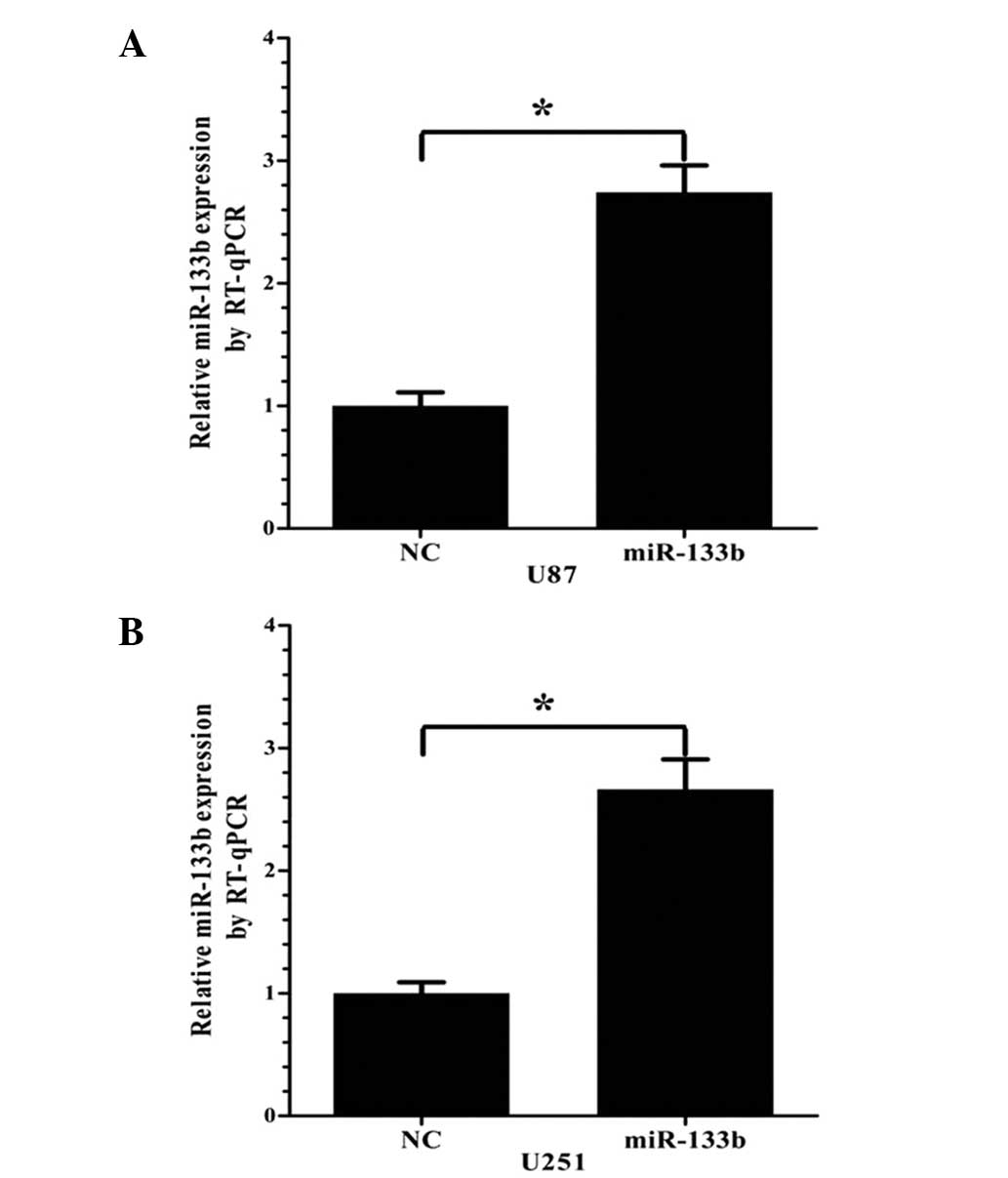
transfection of miR-133b mimics in U87 and U251 cells
miR-133b or NC mimics were transfected into the U87
and U251 cells. After 24 h, the ectopic expression of miR-133b in
the cells was analyzed by RT-qPCR. Compared with the NC group, the
miR-133b mimic group showed significantly upregulated miR-133b
expression in the two cell lines (P<0.05; Fig. 2).
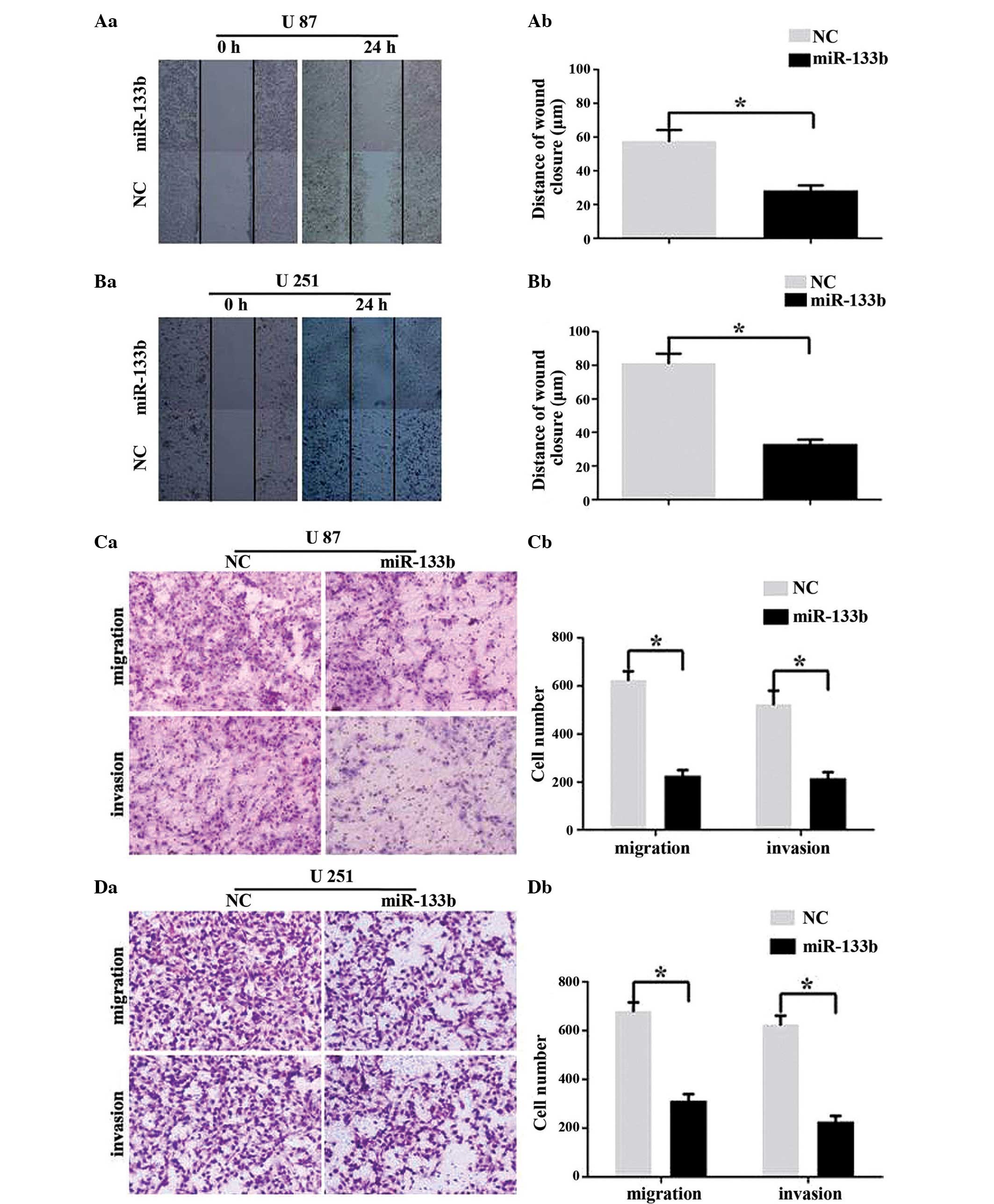
miR-133b inhibits GBM cell migration
and invasion
To determine whether the overexpression of miR-133b
has an effect on the migration and invasion of GBM cells, wound
healing and Transwell assays were performed. As demonstrated in
Fig. 3A and B (wound healing assays),
the overexpression of miR-133b using mimics significantly inhibited
the migratory ability of the U87 and U251 cells (P<0.05). As
shown in Fig. 3C and D, the ability
of the U87 and U251 cells to migrate through a Transwell insert
filter was clearly inhibited by miR-133b (P<0.05). The Transwell
invasion assay also showed that the number of invasive cells
transfected with miR-133b mimics was significantly lower compared
with the NC transfected cells (P<0.05). Therefore, these
findings indicated that miR-133b acts as an invasive suppressor in
GBM cells.
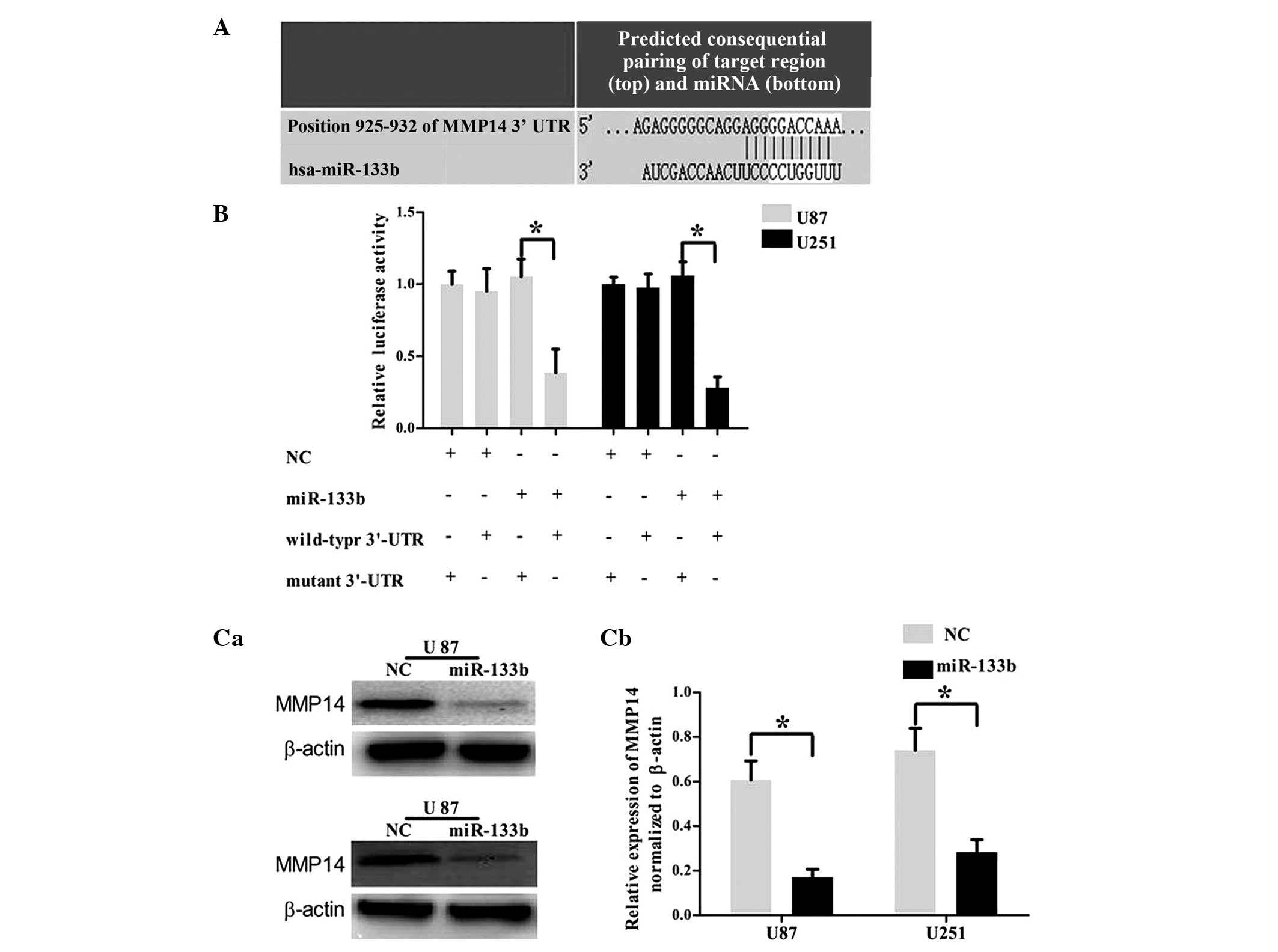
miR-133b targets the 3′UTR of
MMP14
To investigate the mechanism by which miR-133b
regulates cell invasion, an miRNA target search was performed with
four target prediction databases (miRanda, TargetScan, PicTar and
miRWalk), and all four databases showed that the MMP14 mRNA
contained miR-133b binding sites (Fig.
4A). To examine the hypothesis that miR-133b targeted MMP14, a
luciferase reporter assay was performed. The wild-type MMP-9 3′UTR
reporter plasmid or mutated MMP-9 3′UTR reporter plasmid was
cotransfected into the U87 and U251 cells with miR-377 mimic or NC.
As shown in Fig. 4B, the U87 and U251
cell lines cotransfected with miR-133b mimic and wild-type MMP14
3′UTR reporter plasmid showed a significant decrease in reporter
activity (P<0.05). To confirm that the expression of MMP14 is
regulated by miR-133b, western blot analysis was performed. As
shown in Fig. 4C, MMP14 was
significantly downregulated in the U87 and U251 GBM cell lines
following the overexpression of miR-131b (P<0.05). These
findings demonstrated that the 3′UTR of MMP14 was the directed
target of miR-133b.
Discussion
The present study showed for the first time a novel
molecular mechanism of miR-133b and MMP14 in glioma cell invasion.
It was found that miR-133b was frequently downregulated in the
tumor tissues of GBM patients. In addition, MMP14 was identified as
target of miR-133b, and miR-133b significantly induced the protein
expression of MMP14 in glioma cells. More significantly, it was
also concluded that miR-133b suppressed glioma cell invasion via
its target MMP14.
miRNAs are key regulators of gene expression at the
post-transcriptional level and mediate a large variety of
physiological and pathological processes (8,14).
Mounting evidence has indicated that alterations in the expression
of miRNAs correlate with the pathogenesis of different types of
human malignancy, where they serve as either oncogenes or tumor
suppressors. Accumulating studies have demonstrated that miR-133b
functions as a tumor suppressive miRNA in various cancers (15). For instance, it was reported that
miR-133b inhibited the cell viability and colony formation ability
of non-small cell lung cancer (16).
Wu et al showed that miR-133b suppressed the cell
proliferation, migration and invasion of renal cell carcinoma cells
(9). However, the role of miR-133b in
GBM remains unknown. The present study first analyzed the
expression of miR-133b in GBM and found that this expression was
lower in GBM compared with normal brain tissues. The inhibition of
migration and invasion caused by the overexpression of miR-133b in
the human GBM U87 and U251 cell lines was also observed. This
result demonstrates that miR-133b acts as a tumor suppressor and
inhibits GBM cell migration and invasion.
To date, certain targets of miR-133b have been
reported, including FGFR1 in gastric cancer (16), FSCN-1 in esophageal squamous cell
carcinoma (17) and CXCR4 in
colorectal cancer (18). In the
present study, it was shown that miR-133b targets MMP14, revealing
a potential mechanism associated with the development of GBM.
The MMPs, a family of proteolytic enzymes, have been
indicated to be important in the tissue remodeling associated with
various physiological or pathological processes including
arthritis, cirrhosis, tissue repair, angiogenesis, morphogenesis,
tumor invasion and neoplastic metastasis (19). There are 24 soluble and
membrane-anchored members of the MMP family in humans, and MMP14 is
one of them. MMP14 acts either as a pericellular collagenase or as
a proMMP-2 activator via formation of a TIMP-2/roMMP-2/MMP14
complex. Multiple studies have demonstrated that MMP14 is expressed
in various malignant tumors, and its overexpression in tumor cells
promotes tumorigenesis and metastasis. For example, in previous
studies, high MMP14 mRNA expression was found to be an independent
factor of lymph node metastasis and tumor invasion in carcinoma of
the cervix, lung and stomach (20,21). In
addition, it is proposed that a ternary protein complex composed of
αVβ3 integrin, ADAM12 and MMP14 at the tumor cell surface regulates
the function of MMP14 to affect tumor growth (22). More and more studies have confirmed
that MMP14 also plays significant role in glioma progression.
Increases in MMP14 were reported to correlate with poor overall
survival, and may represent a diagnostic and prognostic marker of
glioma (23). Recently, certain
studies have further investigated the mechanisms that regulate
MMP14 expression. A study by Xie et al also found that the
expression of MMP14 was high in human brain gliomas of different
pathological grades and that this may be regulated by activation of
the extracellular signal-regulated kinase 1/2 signaling pathway
(24). Another study found that MMP14
could contribute to the invasion of glioma via HOXD10, which is a
target of miR-10b (10). However, the
results of the present study indicated that MMP14 was regulated by
miR-133b and suggested that miR-133b may suppresses GBM cell
migration and invasion via the downregulation of MMP14.
In summary, the present study showed that miR-133b
was downregulated in GBM. The decreased expression of miR-133b
promoted GBM cell invasion by modulating MMP14, which was
identified as a novel target of miR-133b. Based on these
observations, the study indicates that miR-133b may represent a
potential target for an effective treatment strategy to suppress
GBM invasion and metastasis.
Acknowledgements
This study was supported by the Scientific and
Technological Project of Heilongjiang Province of China (grant no.
GC12C303-1) and the Foundation of Heilongjiang Educational
Committee (grant no. 12541467).
References
|
1
|
Nava F, Tramacere I, Fittipaldo A,
Bruzzone MG, Dimeco F, Fariselli L, Finocchiaro G, Pollo B,
Salmaggi A, Silvani A, et al: Survival effect of first- and
second-line treatments for patients with primary glioblastoma: A
cohort study from a prospective registry, 1997–2010. Neuro Oncol.
16:719–727. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang JB, Dong DF, Gao K and Wang MD:
Mechanisms underlying the biological changes induced by isocitrate
dehydrogenase-1 mutation in glioma cells. Oncol Lett. 7:651–657.
2014.PubMed/NCBI
|
|
3
|
Zhou Y, Xu Y, Wang H, Niu J, Hou H and
Jiang Y: Histone deacetylase inhibitor, valproic acid,
radiosensitizes the C6 glioma cell line in vitro. Oncol Lett.
7:203–208. 2014.PubMed/NCBI
|
|
4
|
Shen SH, Kwan AL, Chen YY and Wang ZX:
Effect of silencing HIF-1α on proliferation, invasion and migration
of glioblastoma U87 cells. Neurol Sci. 34:365–371. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu JH, Yao YL, Gu T, Wang ZY, Pu XY, Sun
WW, Zhang X, Jiang YB and Wang JJ: MiR-421 regulates apoptosis of
BGC-823 gastric cancer cells by targeting caspase-3. Asian Pac J
Cancer Prev. 15:5463–5468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lagos-Quintana M, Rauhut R, Lendeckel W
and Tuschl T: Identification of novel genes coding for small
expressed RNAs. Science. 294:853–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang TS, Yang XH, Wang XD, Wang YL, Zhou B
and Song ZS: MiR-214 regulate gastric cancer cell proliferation,
migration and invasion by targeting PTEN. Cancer Cell Int.
13:682013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu D, Pan H, Zhou Y, Zhou J, Fan Y and Qu
P: microRNA-133b downregulation and inhibition of cell
proliferation, migration and invasion by targeting matrix
metallopeptidase-9 in renal cell carcinoma. Mol Med Rep.
9:2491–2498. 2014.PubMed/NCBI
|
|
10
|
Sun L, Yan W, Wang Y, Sun G, Luo H, Zhang
J, Wang X, You Y, Yang Z and Liu N: MicroRNA-10b induces glioma
cell invasion by modulating MMP-14 and uPAR expression via HOXD10.
Brain Res. 1389:9–18. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao H, Li M, Li L, Yang X, Lan G and
Zhang Y: MiR-133b is down-regulated in human osteosarcoma and
inhibits osteosarcoma cells proliferation, migration and invasion
and promotes apoptosis. PLoS One. 8:e835712013 View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wen D, Li S, Ji F, Cao H, Jiang W, Zhu J
and Fang X: MiR-133b acts as a tumor suppressor and negatively
regulates FGFR1 in gastric cancer. Tumour Biol. 34:793–803. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nohata N, Hanazawa T, Enokida H and Seki
N: MicroRNA-1/133a and microRNA-206/133b clusters: Dysregulation
and functional roles in human cancers. Oncotarget. 3:9–21. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
et al: A mammalian microRNA expression atlas based on small RNA
library sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karatas OF, Guzel E, Suer I, Ekici ID,
Caskurlu T, Creighton CJ, Ittmann M and Ozen M: MiR-1 and miR-133b
are differentially expressed in patients with recurrent prostate
cancer. PLoS One. 9:e986752014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu L, Shao X, Gao W, Zhang Z, Liu P, Wang
R, Huang P, Yin Y and Shu Y: MicroRNA-133b inhibits the growth of
non-small-cell lung cancer by targeting the epidermal growth factor
receptor. FEBS J. 279:3800–3812. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kano M, Seki N, Kikkawa N, Fujimura L,
Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and
Matsubara H: MiR-145, miR-133a and miR-133b: Tumor-suppressive
miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J
Cancer. 127:2804–2814. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duan FT, Qian F, Fang K, Lin KY, Wang WT
and Chen YQ: MiR-133b, a muscle-specific microRNA, is a novel
prognostic marker that participates in the progression of human
colorectal cancer via regulation of CXCR4 expression. Mol Cancer.
12:1642013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Al-Raawi D, Abu-El-Zahab H, El-Shinawi M
and Mohamed MM: Membrane type-1 matrix metalloproteinase (MT1-MMP)
correlates with the expression and activation of matrix
metalloproteinase-2 (MMP-2) in inflammatory breast cancer. Int J
Clin Exp Med. 4:265–275. 2011.PubMed/NCBI
|
|
20
|
Arroyo AG, Genís L, Gonzalo P,
Matí-as-Román S, Pollán A and Gálvez BG: Matrix metalloproteinases:
New routes to the use of MT1-MMP as a therapeutic target in
angiogenesis-related disease. Curr Pharm Des. 13:1787–1802. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Basile JR, Holmbeck K, Bugge TH and
Gutkind JS: MT1-MMP controls tumor-induced angiogenesis through the
release of semaphorin 4D. J Biol Chem. 282:6899–6905. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Albrechtsen R, Kveiborg M, Stautz D,
Vikeså J, Noer JB, Kotzsh A, Nielsen FC, Wewer UM and Fröhlich C:
ADAM12 redistributes and activates MMP-14, resulting in gelatin
degradation, reduced apoptosis and increased tumor growth. J Cell
Sci. 126:4707–4720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang L, Yuan J, Tu Y, Mao X, He S, Fu G,
Zong J and Zhang Y: Co-expression of MMP-14 and MMP-19 predicts
poor survival in human glioma. Clin Transl Oncol. 15:139–145. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie H, Xue YX, Liu LB, Wang P, Liu YH and
Ying HQ: Expressions of matrix metalloproteinase-7 and matrix
metalloproteinase-14 associated with the activation of
extracellular signal-regulated kinase1/2 in human brain gliomas of
different pathological grades. Med Oncol. (28 Suppl 1): S433–S438.
2011. View Article : Google Scholar : PubMed/NCBI
|