Introduction
Oxidative stress is associated with the pathogenesis
of neurological disorders, including dysautonomia, Alzheimer's
disease and Parkinson's disease (1,2). Hydrogen
peroxide (H2O2), which is one of the major
reactive oxygen species, is considered as a major cause of neuronal
cell death (3). Therefore,
pharmacological approaches for intervening in oxidative stress may
be potential therapeutic strategies for neurodegenerative disorders
(4).
Natural antioxidants with neuroprotective potential
are being considered as a promising approach to prevent or slow the
effects of neurological illness, due to their low toxicity and
absence of clear side effects. It has been reported that numerous
natural antioxidants, including resveratrol, celastrol and
salidroside, may protect neurons from oxidative stress injury
(5–7).
Daphnetin (7,8-dihydroxycoumarin), an active
ingredient extracted from Changbai daphne (Daphne Korean
Nakai), exhibits various pharmacological effects, including
anti-inflammatory, anti-oxidative and anti-tumor effects (8,9). However,
whether daphnetin exerts neuroprotection against
H2O2-induced neuronal-like rat
pheochromocytoma PC12 cell apoptosis, and the mechanisms
responsible for this effect, remains unclear.
Inducible heat shock protein (HSP) 70, a member of
the HSP superfamily, is an important protective protein induced by
various stimuli that prevents cell apoptosis (10,11). A
previous study has suggested that HSP70 is protective in
neurodegenerative diseases, including Parkinson's disease, through
its chaperone and direct antiapoptotic role (11). It has also been reported that natural
antioxidants, including celastrol, protect nerve cell damage by
inducing the expression of HSP70 (12).
The present study investigated the activity of
daphnetin in neuronal apoptosis and the underlying mechanisms of
this effect. The present study demonstrated that daphnetin
dose-dependently attenuated H2O2-induced PC12
cell apoptosis via suppression of p38 and c-Jun N-terminal kinases
(JNK) phosphorylation. In addition, the present study revealed that
HSP70 expression was elevated in daphnetin-treated PC12 cells, and
HSP70 expression was regulated by extracellular signal-regulated
kinase (ERK) signaling. Overall, the present study concluded that
daphnetin attenuates p38 and JNK activation and upregulates HSP70
expression in H2O2-treated PC12 cells. These
two mechanisms reduce H2O2-induced PC12
apoptosis, and are protective in oxidative stress-induced neuronal
injury.
Materials and methods
Antibodies and reagents
Daphnetin (purity >98%) was obtained from
Sigma-Aldrich (St. Louis, MO, USA), and the ERK inhibitor U0126 was
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
H2O2 (30%) was purchased from Beyotime
Institute of Biotechnology (Shanghai, China). Rabbit monoclonal
antibodies against β-actin (catalog no., 4970S), Akt, phospho
(p)-Akt (Ser 473; catalog no., 9272S), p38 mitogen-activated
protein kinase (MAPK; catalog no., 8690S), p-p38 MAPK
(Thr180/Tyr182; catalog no., 4511S), ERK (catalog no., 4695S),
p-ERK (Thr202/Tyr204; catalog no., 4376S), JNK/stress-activated
protein kinase (SAPK; catalog no., 9258S), p-JNK/SAPK
(Thr183/Tyr185; catalog no., 4668S), poly ADP-ribose polymerase
(PARP; catalog no., 9532S), cleaved-caspase 3 (catalog no., 9664S;
1:500), pro-caspase 3 (catalog no., 9665S) and HSP70 (catalog no.,
4872S) were all purchased from Cell Signaling Technology, Inc and
used at 1:1,000 dilution, unless otherwise specified. Rabbit
polyclonal antibody against glyceraldehyde 3-phosphate
dehydrogenase (catalog no., AP0063; 1:1,000) was purchased from
Bioworld Technology, Inc. (St. Louis Park, MN, USA). Secondary
antibodies coupled to IRDye800 fluorophore (catalog no., 926-32211;
dilution, 1:5,000) for use with the Odyssey® Infrared
Imaging System (LI-COR Biosciences, Lincoln, NE, USA) were
purchased from LI-COR Biosciences.
Cell culture
Rat pheochromocytoma PC12 cells were obtained from
the American Type Culture Collection (Manassas, VA, USA) and
cultured in Dulbecco's modified Eagle's medium (Invitrogen™; Thermo
Fisher, Scientific, Inc., Waltham, MA, USA) supplemented with 10%
(v/v) heat-inactivated horse serum, 5% fetal bovine serum and 1%
antibiotics (100 U/ml penicillin and 100 µg/ml streptomycin)
(Hyclone™; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C in
5% CO2/95% air humidified atmosphere. Culture medium was
changed every 2–3 days.
Cell viability assay
Cell viability was detected using Cell Counting
Kit-8 (CCK-8; KeyGen Biotech Corp., Ltd., Nanjing, China). Briefly,
PC12 cells were plated onto 96-well plates at a density of
2×104 cells per well 24 h prior to treatment. Cells were
treated with various concentrations of daphnetin (0, 2.5, 5.0, 10.0
and 20.0 µM) for 2 h, then stimulated with
H2O2 (200 µΜ) for 24 h, followed by
incubation with 10 µl CCK-8 working solution at 37% for 4 h. The
absorbance of each well at 450 nm was measured using a Multiskan™
GO Microplate Spectrophotometer (Thermo Fisher Scientific, Inc.).
Three repeats were performed for each of the different
treatments.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) assay
The apoptotic ratio was analyzed using Annexin-V/PI
Double Staining Assay (KeyGen Biotech Corp., Ltd.), according to
the manufacturer's protocol. Briefly, the cells were washed,
trypsinized and resuspended with 500 µl binding buffer, then
stained with Annexin V-FITC (5 µl) and PI (5 µl). The stained cells
were visualized directly using a Guava EasyCyte™ System (EMD
Millipore, Billerica, MA, USA), and the data were analyzed using
Guava TUNEL ExpressPro version 8.0 software (EMD Millipore).
Western blotting
Cells were rinsed twice with ice-cold
phosphate-buffered saline (PBS) and lysed on ice in a lysis buffer
containing 20 mM Tris (pH 7.5), 2 mM EDTA, 135 mM NaCl, 2 mM DTT, 2
mM sodium pyrophosphate, 25 mM β-glycerophosphate, 10% glycerol, 1%
Triton X-100, 1 mM Na3VO4, 10 mM NaF, 10
µg/ml leupeptin, 10 µg/ml aprotinin and 1 mM PMSF, supplemented
with a complete protease inhibitor cocktail (Roche Diagnostics,
Indianapolis, IN, USA) for 30 min. Lysates were centrifuged at
12,500 × g for 15 min at 4°C. Equal amounts of proteins were
subjected to 12% SDS-PAGE, and then transferred onto nitrocellulose
membranes (GE Healthcare Life Sciences, Chalfont, UK). The membrane
was blocked with 5% skimmed milk for 1 h at room temperature,
washed with Tris-buffered saline with Tween 20 three times, then
incubated with the indicated primary antibodies at 4°C overnight.
Subsequently, the membranes were incubated with secondary
antibodies for 1 h at room temperature. The antibody-antigen
complexes were visualized by the Odyssey Infrared Imaging System
using IRDye800 fluorophore-conjugated antibodies
(LI-CORBiosciences).
Morphological observation
PC12 cells were pretreated with various
concentrations of daphnetin (5, 10 and 20 µM) for 2 h, then
stimulated with H2O2 (200 µΜ) for 24 h.
Subsequently, cell morphology was observed using an inverted
microscope (DP72; Olympus Corporation, Tokyo, Japan).
Immunofluorescence microscopy and
4′,6-diamidino-2-phenylindole (DAPI) staining
Cells were pre-incubated with daphnetin for 2 h,
then stimulated with H2O2 for 24 h. Cells
were fixed with 4% paraformaldehyde for 20 min at room temperature,
permeabilized in 0.2% Triton X-100 and then incubated with DAPI (1
µg/ml) for 5 min in the dark. Subsequent to washing with PBS,
nuclear morphology was observed and photographed using a
fluorescence microscope (DP72; Olympus Corporation).
Statistical analysis
Data are expressed as the mean ± standard deviation.
One-way analysis of variance was used to determine the significance
of the difference between two groups. P<0.05 was considered to
indicate a statistically significant difference. Statistical
analysis was performed using SPSS version 17.0 software (SPSS,
Inc., Chicago, IL, USA).
Results
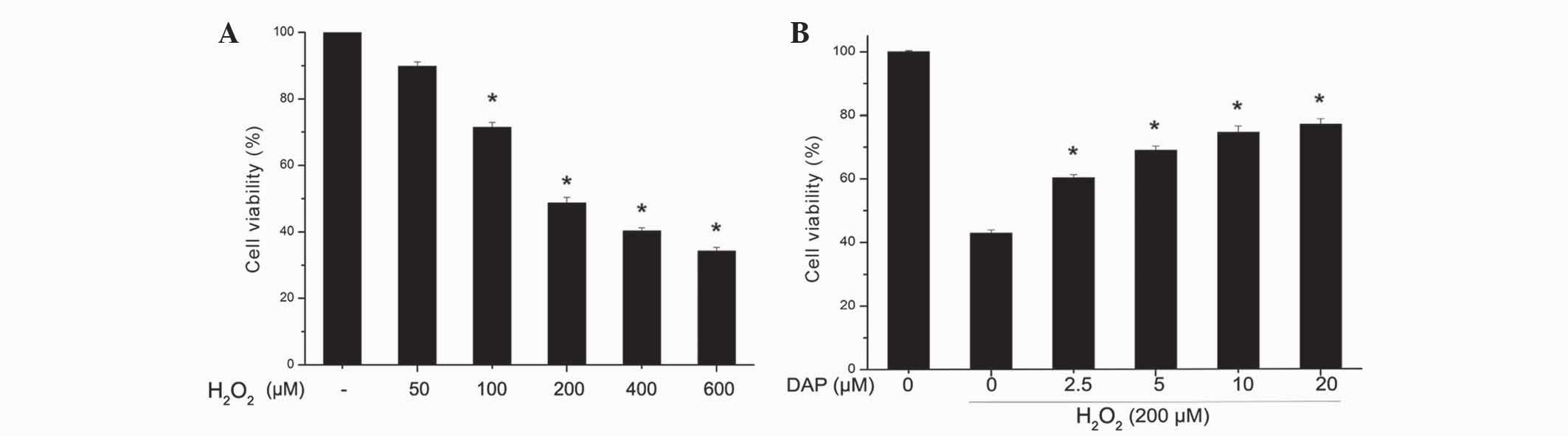
Effect of daphnetin on
H2O2-induced cell viability
PC12 cells were treated with various concentrations
of H2O2 (50, 100, 200, 400 and 600 µM) for 24
h, and subsequently cell viability was determined using CCK-8. As
shown in Fig. 1A,
H2O2 reduced cell viability in a
dose-dependent manner (P<0.05 vs. control). For the cells
treated with 200 µM H2O2, the cell viability
was 48.75±1.63%. Since, the cell viability was reduced by ~50% with
200 µM H2O2 at 24 h, in subsequent
experiments these variables were selected as the standard dose and
time point for the induction of PC12 cell apoptosis. In PC12 cells
treated with various doses of daphnetin and then stimulated with
200 µM H2O2, cell viability was increased in
a daphnetin dose-dependent manner (P<0.05 vs.
H2O2-treated group; Fig. 1B).
Daphnetin attenuated
H2O2-induced PC12 cell apoptosis
Observation of cell morphology revealed that
untreated PC12 cells exhibited typical long fusiform-like
morphology. By contrast, H2O2-treated (200
µM; 24 h) PC12 cells were shrunken, scattered and floating.
Pretreating cells with daphnetin significantly reversed the
apoptotic morphological alterations observed with
H2O2-treated cells (Fig. 2A). To further determine the role of
daphnetin in suppressing H2O2-induced PC12
cell apoptosis, the apoptotic ratio was determined by Annexin
V-FITC/PI assay. As shown in Fig. 2B,
cells exposed to H2O2 for 24 h had an
apoptotic ratio of 55.6%. However, the number of apoptotic cells,
when cells were pretreated with various concentrations of
daphnetin, was significantly reduced; the apoptotic ratio was 34.6,
31.3, 26.6%, for 5, 10 and 20 µM daphnetin, respectively.
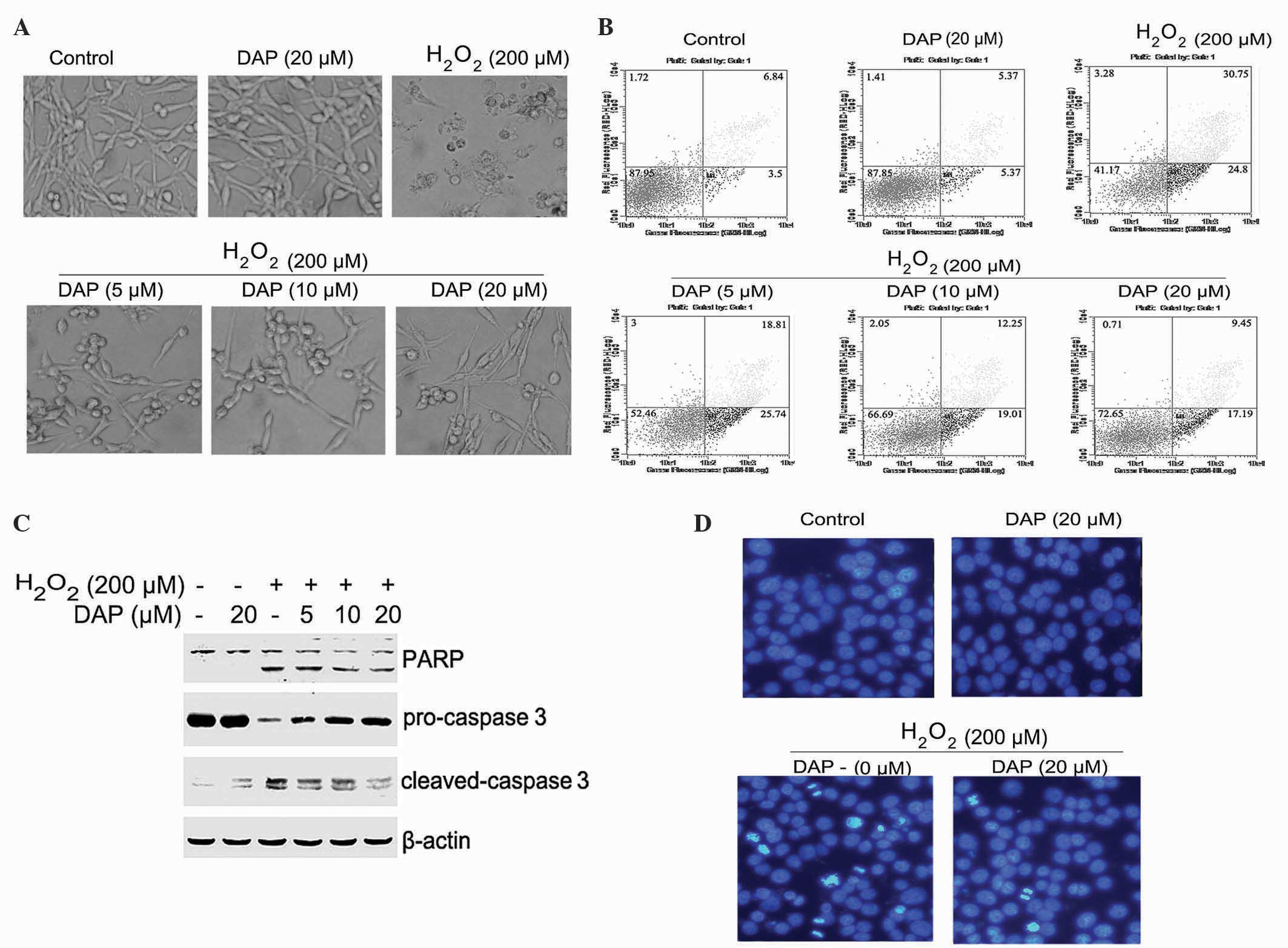 | Figure 2.DAP attenuated
H2O2-induced apoptosis in rat
pheochromocytoma PC12 cells. PC12 cells were pretreated with
various concentrations of daphnetin for 2 h, then stimulated with
H2O2 (200 µΜ) for 24 h. (A) Cell morphology
was detected using an inverted microscope. (B) The apoptotic ratio
was determined by Annexin V-FITC/PI assay. (C) Cleavage of PARP
(89/116 kDa) and caspase 3 (17/19 kDa), and the level of
pro-caspase 3 (35 kDa) were detected by western blotting (β-actin,
45 kDa). (D) Cells were stained with 4′,6-diamidino-2-phenylindole,
and nuclear morphology was observed and photographed using a
fluorescence microscope. H2O2, hydrogen
peroxide; DAP, daphnetin; FITC, fluorescein isothiocyanate; PI,
propidium iodide; PARP, poly ADP-ribose polymerase. |
The activation of apoptosis-associated proteins was
detected using western blotting to confirm the occurrence of
apoptosis. Daphnetin pretreated cells clearly attenuated the
H2O2-induced cleavage of PARP and caspase 3,
and enhanced the level of pro-caspase 3 (Fig. 2C) compared with cells not pretreated
with daphnetin. DAPI staining was used to determine the apoptotic
status of the PC12 cells. Staining with DAPI revealed that the
round nuclei of normal cells was homogeneous, and when exposed to
200 µM H2O2 for 24 h cells underwent nuclear
condensation and fragmentation. However, these alterations in
nuclear morphology were significantly attenuated by pretreatment
with daphnetin (Fig. 2D). Overall,
these results suggest that daphnetin has a protective effect in
H2O2-induced PC12 cell apoptosis.
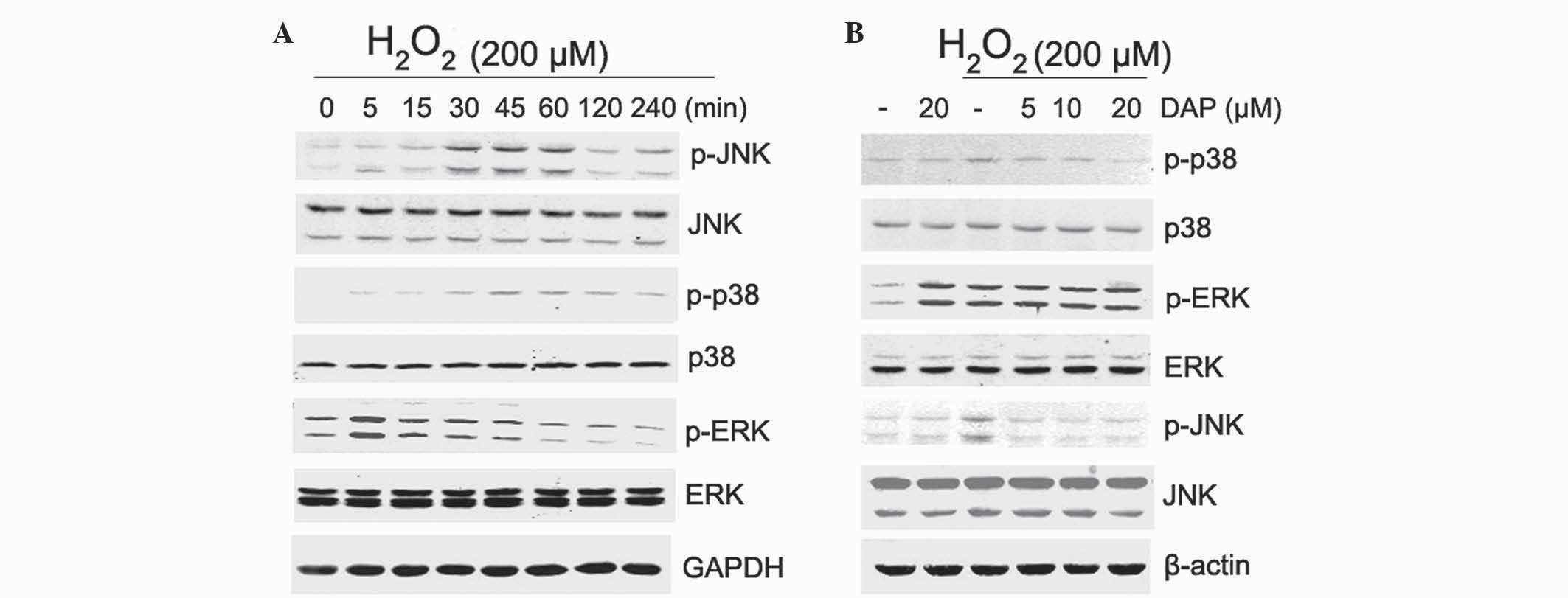
Daphnetin reduced the activation of
p38 MAPK and JNK induced by H2O2
To determine the mechanism by which daphnetin
inhibits H2O2-induced apoptosis, the
phosphorylation levels of MAPKs in PC12 cells were investigated.
PC12 cells were stimulated with 200 µM H2O2
for various times, and the phosphorylation levels of p38, ERK and
JNK were detected by western blotting. The results showed that
stimulation of PC12 cells with H2O2 resulted
in an increase in phosphorylation of JNK, p38 and ERK (Fig. 3A). The enhanced phosphorylation of JNK
and p38 was significantly attenuated by daphnetin pretreatment in a
dose-dependent manner (Fig. 3B).
However in the presence of daphnetin, increased ERK phosphorylation
observed with H2O2 stimulation was not
attenuated. In addition, with daphnetin stimulation,
phosphorylation of ERK was clearly elevated, but ERK activation had
no additive effect on daphnetin and H2O2
stimulation (Fig. 3B).
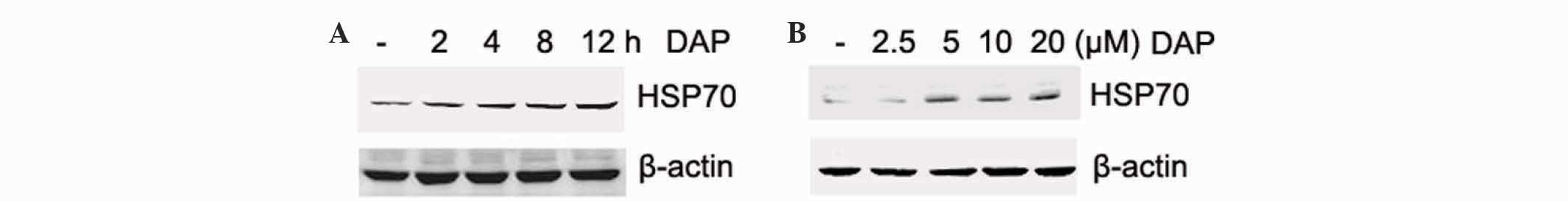
Daphnetin induced HSP70 expression in
PC12 cells
In order to determine other mediators in daphnetin
neuroprotection, HSP70, a well-known chaperone with cytoprotective
effects, was investigated. PC12 cells were stimulated with
daphnetin, and total cellular protein was extracted and subjected
to western blotting with an anti-HSP70 antibody. The present
results revealed that the level of HSP70 was increased upon
daphnetin stimulation in a dose- and time-dependent manner
(Fig. 4A and B). The level of HSP70
peaked at 12 h.
ERK signaling is involved in
daphnetin-induced HSP70 expression
The phosphorylation of upstream kinases was examined
to investigate the possible signal transduction pathways involved
in daphnetin-induced HSP70 expression. PC12 cells were treated with
daphnetin (20 µM) for various times and the phosphorylation level
of ERK and Akt was detected by western blotting. As shown in
Fig. 5A, the activation of ERK was
enhanced in a time-dependent manner following daphnetin
stimulation. ERK activation peaked at 12 h, which is consistent
with the expression of HSP70. By contrast, Akt phosphorylation was
not significantly different compared with unstimulated PC12 cells.
Overall, these observations suggested that ERK signaling activation
may be involved in daphnetin-induced HSP70 expression.
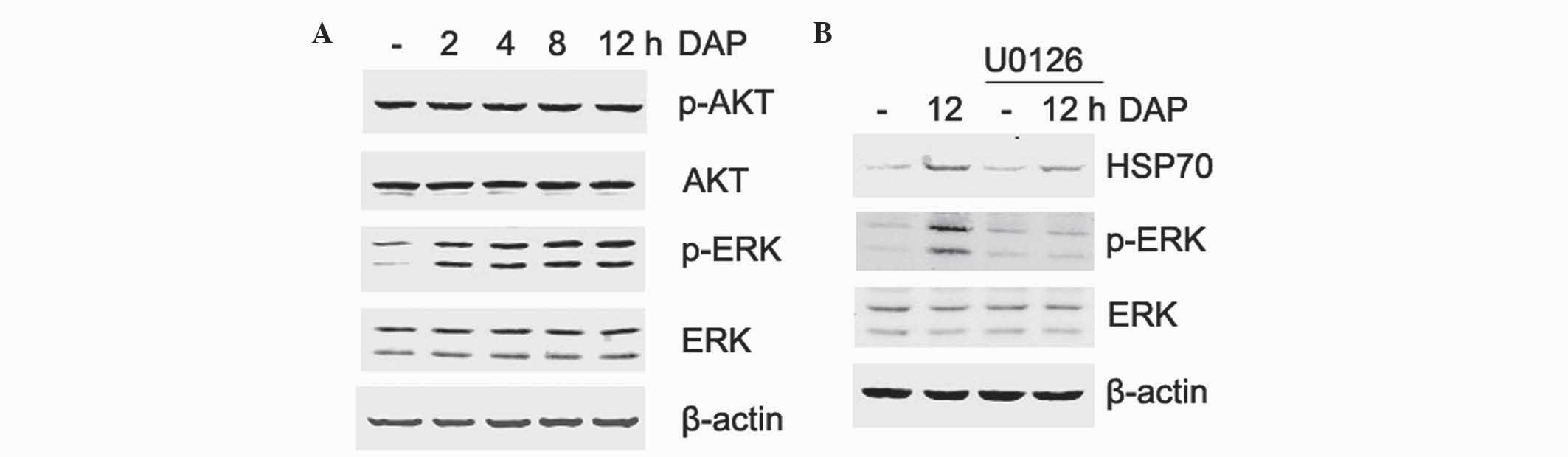 | Figure 5.ERK signaling is involved in
DAP-induced HSP70 expression. (A) Rat pheochromocytoma PC12 cells
were stimulated with 20 µM daphnetin for 2, 4, 8 and 12 h, and
total cellular protein was extracted and subjected to western
blotting with p-Akt (60 kDa), Akt (60 kDa), p-ERK (42/44 kDa) and
ERK (42/44 kDa) antibodies. (B) PC12 cells were pretreated with
U0126 for 2 h, then treated with daphnetin for 12 h. Cell lysates
were prepared and subjected to western blotting using HSP70 (70
kDa), p-ERK and ERK antibodies. Equal protein loading was confirmed
by β-actin (45 kDa). DAP, daphnetin; HSP, heat shock protein; ERK,
extracellular signal-regulated kinase; p, phospho. |
To test this hypothesis, U0126, a specific inhibitor
of ERK, was used to verify the activation of ERK in
daphnetin-induced HSP70 expression. PC12 cells were pretreated with
20 µM U0126 for 2 h, and then exposed to 20 µM daphnetin for a
further 12 h, and the expression of HSP70 and phosphorylated ERK
were detected using western blotting. As shown in Fig. 5B, attenuation of ERK activation by
U0126 clearly reduced the protein expression level of HSP70.
Therefore, the present data suggest that HSP70 is involved in
daphnetin-mediated cytoprotection via ERK signaling.
Discussion
Oxidative stress has been widely implicated in
neuronal cell death, and is associated with a variety of chronic
neurodegenerative diseases, including Parkinson's, Alzheimer's and
Huntington's disease. H2O2 is a strong
oxidant, which reduces cell viability and increases cell apoptosis
(13). Currently, great effort is
being made to identify potent natural antioxidants with
neuroprotective potential, due to their low toxicity and absence of
clear side effects. Daphnetin, a natural antioxidant, exhibits a
variety of biological effects, including anti-inflammatory and
antitumor effects (14). Recently,
studies have demonstrated that daphnetin exhibits a neurotrophic
effect on peripheral nerve regeneration by suppressing nuclear
factor-κB expression (15), provides
a neuroprotective effect against glutamate-induced toxicity in
immortalized mouse hippocampal HT22 cells and ischemic brain injury
(16), and prevents excitotoxicity by
inhibiting the N-methyl D-aspartate receptor subtype 2B -containing
receptors and the subsequent calcium overload in cultured cortical
neurons (17). These studies suggest
that daphnetin has a neuroprotective effect, which is consistent
with the present findings. The present study confirmed that
treating PC12 cells with H2O2 resulted in
suppression of cell viability in a dose-dependent manner; however,
pretreatment of these cells with various concentrations (5, 10, 20
µM) of daphnetin significantly increased PC12 cell viability
(Fig. 1). The present study further
investigated whether daphnetin has protective effects against
H2O2-induced nerve cell apoptosis. Cell
morphology analysis revealed that untreated PC12 cells exhibited
typical long fusiform-like morphology, while treatment with
H2O2 (200 µM) for 24 h resulted in shrunken,
scattered and floating cells. However, pre-incubation with
daphnetin clearly reversed the morphology induced by
H2O2 (Fig. 2A).
The cell apoptotic ratio was detected using flow cytometry, and the
present data revealed that the number of apoptotic cells was
significantly reduced in daphnetin-pretreated PC12 cells compared
with cells stimulated with H2O2 (Fig. 2B). The present study also examined the
activation of apoptotic proteins by immunoblotting, in order to
confirm the occurrence of apoptosis. Upon
H2O2 stimulation, cleavage of PARP and
caspase 3 were clearly enhanced in PC12 cells; however,
pretreatment with daphnetin significantly decreased the cleavage of
PARP and caspase 3 in a concentration-dependent manner (Fig. 2C). DAPI staining of
H2O2-treated cells revealed typical apoptotic
morphology. However, pre-incubation with daphnetin clearly
attenuated these morphological alterations (Fig. 2D). These results suggest that
daphnetin exerts its neuroprotective effect via inhibition of PC12
cell apoptosis.
Certain studies have revealed that oxidative
stress-induced toxicity is associated with intracellular signaling,
including the activation of JNK and p38 signaling pathways
(7,13). In addition, it has been reported that
the activation of ERK may protect neurons from oxidative
stress-induced cell death (18). The
present study hypothesized that daphnetin may protect PC12 cells
against H2O2-induced apoptosis via the MAPK
signaling pathway. Western blotting revealed that p38, ERK and JNK
were phosphorylated upon H2O2 stimulation,
and ERK was activated earlier than p38 MAPK and JNK activation.
Additionally, the enhanced phosphorylation of JNK and p38 observed
following stimulation with H2O2 was clearly
attenuated by daphnetin pre-treatment in a dose-dependent manner.
However, phosphorylation of ERK upon H2O2
stimulation was not attenuated by daphnetin treatment (Fig. 3). Furthermore, daphnetin was
demonstrated to induce ERK activation, and ERK activation had no
cumulative effect in PC12 cells stimulated with daphnetin and
H2O2. Therefore, the present study
hypothesized that ERK activation reached the maximum when PC12
cells were treated with daphnetin. These results suggest that
daphnetin protects PC12 cells by suppressing p38 MAPK and JNK, and
enhances ERK signaling.
HSP70 is induced in cells in response to a wide
variety of chemical and physiological stresses, and its expression
provides protection against cell death (10,19,20). A
previous study also suggested that HSP70 is protective in nervous
system diseases (21), and HSP70
protects against neuronal, apoptosis at least in part, by
inhibiting caspase-dependent and caspase-independent programmed
cell-death pathways (22). Therefore,
the present study investigated the effect of daphnetin on the
expression of HSP70 in PC12 cells. The present data revealed that
daphnetin clearly upregulates HSP70 expression in a dose- and
time-dependent manner, which reached its maximum at 12 h (Fig. 4A and B).
To further determine the possible signal
transduction pathway involved in daphnetin-induced HSP70
expression, the present study investigated pro-survival signaling
protein activation, including Akt and ERK, upon daphnetin
treatment. The activation of ERK was enhanced time-dependently upon
daphnetin stimulation (Fig. 5A). By
contrast, Akt phosphorylation did not exhibit a significant
difference compared with unstimulated PC12 cells. Furthermore, a
specific inhibitor of ERK revealed that ERK significantly
suppressed HSP70 protein expression (Fig.
5B). These data confirmed that ERK, but not Akt, signaling is
involved in daphnetin-mediated HSP70 expression.
In conclusion, the present study elucidated that
daphnetin protects PC12 cells against
H2O2-induced apoptosis by suppressing p38
MAPK and JNK signaling activation and increasing HSP70 expression
via ERK signaling. Overall, the present results suggest that
daphnetin may be a potential candidate for the treatment of
neurodegenerative diseases. However, daphnetin should be further
tested in animal models mimicking neurodegenerative diseases prior
to being considered as a candidate for a clinical trial to prevent
neurodegenerative disease progression in humans.
Acknowledgements
The present study was supported by the Natural
Science Research Project of Anhui Colleges and Universities (grant
no. KJ2016SD59), Outstanding Young Talent Support Program Key
Projects in Anhui Colleges and Universities (grant no.
gxyqZD2016173), National Nature Science Foundation of China (grant
no. 31301171), Natural Science Research Project of Anhui Provincial
Education Department (grant no. KJ2013B311) and Anhui Province Key
Laboratory of Active Biological Macro-molecules (grant no.
1306C083008).
Glossary
Abbreviations
Abbreviations:
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
H2O2
|
hydrogen peroxide
|
|
HSP
|
heat shock protein
|
References
|
1
|
Gong QH, Shi XR, Hong ZY, Pan LL, Liu XH
and Zhu YZ: A new hope for neurodegeneration: Possible role of
hydrogen sulfide. J Alzheimers Dis. 24(Suppl 2): S173–S182.
2011.
|
|
2
|
Pan LL, Liu XH, Jia YL, Wu D, Xiong QH,
Gong QH, Wang Y and Zhu YZ: A novel compound derived from danshensu
inhibits apoptosis via upregulation of heme oxygenase-1 expression
in SH-SY5Y cells. Biochim Biophys Acta. 1830:2861–2871. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chong ZZ, Li F and Maiese K: Oxidative
stress in the brain: Novel cellular targets that govern survival
during neurodegenerative disease. Prog Neurobiol. 75:207–246. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Asaithambi A, Kanthasamy A, Saminathan H,
Anantharam V and Kanthasamy AG: Protein kinase D1 (PKD1) activation
mediates a compensatory protective response during early stages of
oxidative stress-induced neuronal degeneration. Mol Neurodegener.
6:432011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karuppagounder SS, Pinto JT, Xu H, Chen
H-L, Beal MF and Gibson GE: Dietary supplementation with
resveratrol reduces plaque pathology in a transgenic model of
Alzheimer's disease. Neurochem Int. 54:111–118. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin W, Zhang X, Shi X and Li Y: Curcumin
protects SH-SY5Y cells from oxidative stress by up-regulating HO-1
via Phosphatidylinositol 3 Kinase/Akt/Nrf-2 and down-regulating
HO-2. Mol Neurodegener. 7(Suppl 1): S142012. View Article : Google Scholar
|
|
7
|
Zhang L, Ding W, Sun H, Zhou Q, Huang J,
Li X, Xie Y and Chen J: Salidroside protects PC12 cells from
MPP+-induced apoptosis via activation of the PI3K/Akt
pathway. Food Chem Toxicol. 50:2591–2597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ouyang X and Yu T: Effect of Daphnetin on
the expression of β-APP after the ischemia in the front brain of
the mouse. Zhonghua Naoxueguanbing Zazhi. 6:2012.(In Chinese).
|
|
9
|
Kuang NZ, Fu YY, Gong SQ, Zeng ZP, Zhang
ZQ and Chen ZY: Effects of Daphnetin extract on the expression of
Bcl-2 and Bax mRNA during the apoptosis of SMMC-7721 cells.
Guangdong Yiyao Zazhi. 33:1374–1377. 2012.(In Chinese).
|
|
10
|
Gao Y, Han C, Huang H, Xin Y, Xu Y, Luo L
and Yin Z: Heat shock protein 70 together with its co-chaperone
CHIP inhibits TNF-alpha induced apoptosis by promoting proteasomal
degradation of apoptosis signal-regulating kinase1. Apoptosis.
15:822–833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sabirzhanov B, Stoica BA, Hanscom M, Piao
CS and Faden AI: Over-expression of HSP70 attenuates
caspase-dependent and caspase-independent pathways and inhibits
neuronal apoptosis. J Neurochem. 123:542–554. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chow AM, Tang DW, Hanif A and Brown IR:
Induction of heat shock proteins in cerebral cortical cultures by
celastrol. Cell Stress Chaperones. 18:155–160. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kou X, Shen K, An Y, Qi S, Dai WX and Yin
Z: Ampelopsin inhibits H2O2-induced apoptosis
by ERK and Akt signaling pathways and up-regulation of heme
oxygenase-1. Phytother Res. 26:988–994. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Li CF, Pan LM and Gao ZL:
7,8-Dihydroxycoumarin inhibits A549 human lung adenocarcinoma cell
proliferation by inducing apoptosis via suppression of Akt/NF-kB
signaling. Exp Ther Med. 5:1770–1774. 2013.PubMed/NCBI
|
|
15
|
Du JS, Zhao Q, Zhang YL, Wang Y and Ma M:
7,8-dihydroxycoumarin may promote sciatic nerve regeneration by
suppressing NF-kB expression in mice. Mol Med Rep. 8:1525–1530.
2013.PubMed/NCBI
|
|
16
|
Du G, Tu H, Li X, Pei A, Chen J, Miao Z,
Li J, Wang C, Xie H, Xu X and Zhao H: Daphnetin, a natural coumarin
derivative, provides the neuroprotection against glutamate-induced
toxicity in HT22 cells and ischemic brain injury. Neurochem Res.
39:269–275. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang L, Yang Q, Zhang K, Li YJ, Wu YM, Liu
SB, Zheng LH and Zhao MG: Neuroprotective effects of daphnetin
against NMDA receptor-mediated excitotoxicity. Molecules.
19:14542–14555. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu S, Shen Y, Liu J and Ding F:
Involvement of ERK1/2 pathway in neuroprotection by salidroside
against hydrogen peroxide-induced apoptotic cell death. J Mol
Neurosci. 40:321–331. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takayama S, Reed JC and Homma S:
Heat-shock proteins as regulators of apoptosis. Oncogene.
22:9041–9047. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao Y, Han C, Huang H, Xin Y, Xu Y, Luo L
and Yin Z: Heat shock protein 70 together with its co-chaperone
CHIP inhibits TNF-alpha induced apoptosis by promoting proteasomal
degradation of apoptosis signal-regulating kinase1. Apoptosis.
15:822–833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Turturici G, Sconzo G and Geraci F: Hsp70
and its molecular role in nervous system diseases. Biochem Res Int.
2011:6181272011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sabirzhanov B, Stoica BA, Hanscom M, Piao
CS and Faden AI: Over-expression of HSP70 attenuates
caspase-dependent and caspase-independent pathways and inhibits
neuronal apoptosis. J Neurochem. 123:542–554. 2012. View Article : Google Scholar : PubMed/NCBI
|