Introduction
Docosahexaenoic acid (DHA) is an omega (ω)-3 fatty
acid (22:6Δ4,7,10,13,16,19) and a member of a family of compounds
known to possess multiple benefits for human health, including
anticancer properties (1).
Epidemiological evidence strongly links fish oil, which is rich in
DHA and eicosapentaenoic acid (EPA), with a reduced incidence of
several types of cancer, including breast cancer (2,3).
Epidemiological studies, dietary studies in mice and humans, and
tissue culture studies have substantiated the beneficial role of
DHA in breast cancer prevention (4–7).
Cancer treatment failure largely occurs due to
cancer cell proliferation, invasion and metastasis, which
ultimately lead to mortality. Invasion and metastasis are the major
causes of morbidity and mortality in breast cancer patients
(3). Metastasis involves the
penetration of cancer cells into the extracellular basement
membrane by proteolytic degradation of components of the
extracellular matrix (ECM) (8),
including type IV collagen, laminin, heparin sulfate proteoglycan,
nidogen and fibronectin (9), which
normally provide biochemical and mechanical barriers to cell
movement (10).
ECM degradation requires extracellular proteinases
such as matrix metalloproteinases (MMPs), a group of zinc and
calcium-dependent endopeptidases that can be divided into different
subclasses (including collagenases, gelatinases and stromelysins)
based on their substrate (8). MMP-9
is directly associated with invasion, metastasis and poor prognosis
in breast cancer (11,12). MMP-9 is stimulated by growth factors
(including fibroblast growth factor 2, epidermal growth factor and
hepatocyte growth factor), cytokines (such as tumor necrosis factor
α), oncogenes (such as Ras) and
12-O-tetradecanoylphorbol-13-acetate (TPA) (13–17). TPA
is a selective activator of protein kinase C (14). TPA can stimulate MMP-9 synthesis and
secretion during breast cancer cell invasion (18,19).
Cytokine and TPA treatment induce MMP-9 expression via activation
of transcription factors such as nuclear factor (NF)-κB and
activator protein (AP)-1 (20–22), since
the MMP-9 gene promoter contains binding sites for both factors
(23). Mitogen-activated protein
kinase (MAPK) signaling is important for AP-1 and NF-κB activation,
and requires nuclear factor of κ light polypeptide gene enhancer in
B cells inhibitor (IκB) kinase, extracellular signal-regulated
kinase (ERK), c-Jun N-terminal kinase (JNK) or p38 MAPK, depending
on the cell type (17,24,25). Thus,
inhibiting MMP-9 expression and/or its upstream regulatory pathways
may aid in the treatment of malignant tumors, including breast
carcinoma.
Peroxisome proliferator-activated receptors (PPARs)
belong to the nuclear receptor superfamily of ligand-activated
transcription factors. Three types of PPARs [-α, -β and -δ (β)]
have been identified thus far (26).
PPAR-γ is expressed in various cell types and tissues, and is
important in the regulation of inflammatory (27) and immune reactions (28), as well as in NF-κB activation
(29). In a previous study, agents
that inhibit MMP-9 expression in MCF-7 cells were described
(30–32). The present study reports for the first
time that increased PPAR-γ expression inhibits breast cancer
metastasis via regulation of NF-κB activation. These results
provide an insight into the anti-cancer actions of an ω-3 fatty
acid, which could aid in the development of new cancer therapeutic
strategies involving the use of fish oil as a dietary
supplement.
Materials and methods
Cells and reagents
MCF-7 cells were obtained from the American Type
Culture Collection (Manassas, VA, USA). Cells were cultured in
Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal
bovine serum (FBS) and 1% antibiotics at 37°C in a 5%
CO2 incubator. TPA, cis-4,7,10,13,16,19-DHA, MTT,
GW9662 (a PPAR-γ antagonist) and anti-β-actin antibody (catalog no.
A5441) were obtained from Sigma-Aldrich (Merck Millipore,
Darmstadt, Germany). Antibodies against p38 (catalog no. 9212),
phosphorylated (p)-p38 (catalog no. 9211), JNK (catalog no. 9252),
p-JNK (catalog no. 9251), ERK (catalog no. 9102) and p-ERK (catalog
no. 9101) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Antibodies against PPAR-γ (catalog no. 7169),
MMP-9 (catalog no. 12759), p50 (catalog no. 7178), p65 (catalog no.
372) and proliferating cell nuclear antigen (catalog no. 7907), and
horseradish peroxidase (HRP)-conjugated immunoglobulin (Ig)G
(catalog no. 2004, 2005) were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). [γ-32P] adenosine
triphosphate was obtained from GE Healthcare Life Sciences
(Chalfont, UK). High glucose DMEM, FBS and PBS were acquired from
Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Cell viability assay
The effect of DHA on MCF-7 cell viability was
determined by MTT assay. A stock solution of 200 mM DHA was
prepared in dimethylsulfoxide (DMSO) and diluted with DMEM prior to
the experiments. Briefly, 3xl04 cells/well were seeded
in 96-well plates and incubated at 37°C for 24 h to allow
attachment. Cells were then left untreated, or were treated with
50, 100 and 200 µM DHA for 24 h at 37°C. Cells were next washed
with PBS prior to the addition of MTT (0.5 mg/ml in PBS), and
incubated at 37°C for 30 min. Formazan crystals were dissolved with
DMSO (100 µl/well), and absorbance was measured at 570 nm using a
Model 3550 microplate reader (Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Western blot analysis
MCF-7 cells (5×105) were pre-treated with
50 and 100 µM DHA for 1 h, and then incubated with TPA for 24 h at
37°C. Cells were lysed with ice-cold M-PER® Mammalian
Protein Extraction Reagent (Pierce; Thermo Fisher Scientific,
Inc.). The protein concentration in the lysate was determined by
the Bradford method (33). Samples
(20 µg) were separated by 10% SDS-PAGE and transferred to Hybond
polyvinylidene difluoride membranes (GE Healthcare Life Sciences).
Membranes were blocked for 2 h with 2% bovine serum albumin
(Sigma-Aldrich; Merck Millipore) or 5% skimmed milk, and then
incubated overnight at 4°C with primary antibodies at 1:2,000
dilution, followed by incubation with HRP-conjugated IgG at 1:2,000
dilution for 2 h at 4°C. Protein expression levels were measured by
signal analysis using an image analyzer (Fujifilm, Tokyo, Japan)
and specific immunoreactive signals were visualized with an
enhanced chemiluminescence kit (GE Healthcare Life Sciences).
Gelatin zymography assay
Conditioned media were collected after 24 h of cell
stimulation, mixed with non-reducing sample buffer and resolved by
PAGE containing 0.1% (w/v) gelatin. The gel was washed at room
temperature for 30 min with 2.5% Triton X-100 solution, and
incubated at 37°C for 16 h in 5 mM CaCl2, 0.02% Brij
(Sigma-Aldrich; Merck Millipore) and 50 mM Tris-HCl (pH 7.5). The
gel was stained for 30 min with 0.25% (w/v) Coomassie Brilliant
Blue in 40% (v/v) methanol/7% (v/v) acetic acid, and photographed
on an image analyzer (Fujifilm). Proteolysis was imaged as a white
zone in a dark blue field. Densitometric analysis was performed
using MultiGauge image analysis software (version 3.0;
Fujifilm).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using a FastPure
RNA kit (Takara Bio, Inc., Otsu, Japan). The RNA concentration and
purity were assessed by measuring the absorbance value at 260/280
nm. Complementary DNA was synthesized from 1 µg total RNA using a
PrimeScript RT reagent kit (Takara Bio, Inc.) at 37°C for 15 min
and 85°C for 5 sec. MMP-9 and GAPDH messenger (m)RNA expression
were determined by qPCR using the ABI PRISM 7900HT Sequence
Detection System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and SYBR Green PCR Master Mix (Applied Biosystems; Thermo
Fisher Scientific, Inc.) with the following sense and antisense
primer sets: MMP-9 (NM_004994), 5′-CCTGGAGACCTGAGAACCAATCT-3′
(forward) and 5′-CCACCCGAGTGTAACCATAGC-3′ (reverse), and GAPDH
(NM_002046), 5′-ATGGAAATCCCATCACCATCTT-3′ (forward) and
5′-CGCCCCACTTGATTTTGG-3′ (reverse). To control for variation in
mRNA concentration, the results were normalized to the level of
GAPDH. Relative quantitation was performed using the comparative
2−∆∆Cq method (34),
according to the manufacturer's protocol.
Preparation of nuclear extract
MCF-7 cells (2×106) were treated with DHA
in the presence or absence of TPA for 4 h. Cells were immediately
washed twice, scraped into 1.5 ml ice-cold PBS (pH 7.5) and
pelleted at 1,500 × g for 3 min. Cytoplasmic and nuclear
extracts were prepared from cells using the NE-PER®
Nuclear and Cytoplasmic Extraction kit (Pierce; Thermo Fisher
Scientific, Inc.).
Electrophoretic mobility shift assay
(EMSA)
NF-κB activation was evaluated with a gel mobility
shift assay using nuclear extracts. Oligonucleotides containing a
binding site for the κ chain (κB, 5′-CCGGTTAACAGAGGGGGCTTTCCGAG-3′)
or AP-1 (5′-CGCTTGATGAGTCAGCCGGAA-3′) binding site was synthesized
and used as a probe (Promega, Madison, WI, USA). The two
complementary strands were annealed and labeled with
[α-32P] deoxycytidine triphosphate. Labeled
oligonucleotides (10,000 cpm) were combined with 10 µg nuclear
extracts and binding buffer [10 mM Tris-HCl (pH 7.6), 500 mM KCl,
10 mM EDTA, 50% glycerol, 100 ng poly (dI:dC) (Roche, Basel,
Switzerland) and 1 mM dithiothreitol], and incubated for 30 min at
room temperature in a final volume of 20 µl. The reaction products
were analyzed by 4% PAGE in 0.5X Tris-borate buffer. The gels were
then dried and examined by autoradiography. A 50-fold excess of
cold κB oligonucleotide was used as a control to confirm binding
specificity.
Invasion assay
The invasion assay was conducted in 24-well chambers
(8-µm pore size) coated with 20 µl Matrigel diluted in DMEM. The
Matrigel coating was re-hydrated in 0.5 ml DMEM for 30 min
immediately prior to the experiment. Cells (2×105) were
added to the upper chamber, with the chemoattractant in the bottom
well. Conditioned medium (0.5 ml) was added to the lower
compartment of the invasion chamber, followed by incubation for 24
h. Subsequently, cells on the upper side of the chamber were
removed using cotton swabs, while those that had migrated were
fixed and stained with Toluidine Blue solution. Invading cells were
counted in five random areas of the membrane under a light
microscope. Data from three individual experiments performed in
triplicate were analyzed and presented as the mean ± standard error
of the mean.
Statistical analysis
Data were evaluated by analysis of variance and
Duncan's test using the Microsoft 2010 Excel program (Microsoft
Corporation, Redmond, WA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
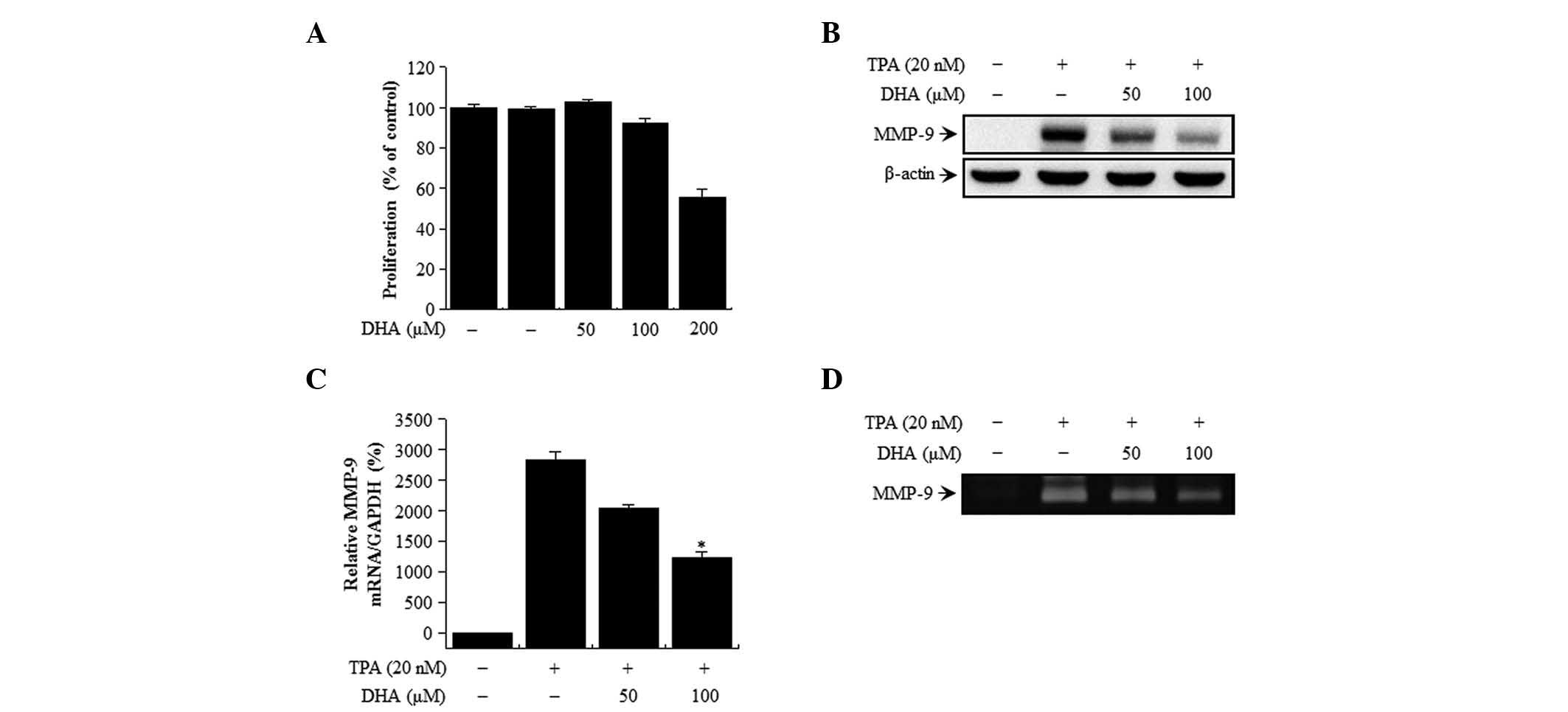
MCF-7 cell viability is unaffected by
DHA treatment
The cytotoxicity of DHA on MCF-7 cells was evaluated
by MTT assay. There were no changes in cell viability or morphology
upon treatment with the indicated concentrations of DHA for 24 h
(Fig. 1A). Therefore, the subsequent
experiments were performed at optimal, non-toxic DHA concentrations
of 50 and 100 µM.
DHA suppresses TPA-induced MMP-9
activation in MCF-7 cells
The effect of DHA on TPA-induced MMP-9 expression in
MCF-7 cells was examined by western blot analysis, RT-qPCR and
gelatin zymography. DHA treatment blocked the upregulation of MMP-9
protein expression induced by TPA, as determined by western
blotting (Fig. 1B). Accordingly,
RT-qPCR analysis revealed that the increase in MMP-9 expression
induced by TPA treatment was abrogated by DHA in a dose-dependent
manner (Fig. 1C). MMP-9 secretion was
stimulated by TPA, but this effect was abrogated by treatment with
DHA, as determined by zymography (Fig.
1D). These results indicate that DHA potently inhibits the
TPA-induced increase in MMP-9 levels in MCF-7 cells.
DHA inhibits TPA-induced NF-κB but not
AP-1 DNA binding activity, as well as MAPK signaling
To investigate the mechanism of DHA-mediated
inhibition of MMP-9 expression, the effect of DHA on TPA-induced
NF-κB activation was evaluated by EMSA. TPA increased the NF-κB
binding activity, whereas pre-treatment with DHA abolished this
effect for NF-κB (Fig. 2A) but not
for AP-1 (data not shown). These results suggest that DHA
specifically blocks NF-κB activation in MCF-7 cells. DHA inhibited
the phosphorylation of p38 and ERK, but not that of JNK, 30 min
after TPA treatment (Fig. 2B).
Additionally, TPA induced the phosphorylation of cytoplasmic IκBα
and the consequent nuclear translocation of the NF-κB subunits p50
and p65, as determined by western blot analysis. In addition, c-Jun
but not c-Fos expression was upregulated upon TPA treatment.
Additionally, TPA also increased p-IκBα expression, as well as p65
and p50 translocation, which was suppressed by treatment with DHA
(Fig. 2C and D). However, the TPA
induced phosphorylation of c-Jun (a major subunit of AP-1) was
unaffected by DHA (Fig. 2D). These
results suggest that the effect of DHA on the regulation of
TPA-induced MMP-9 expression is via modulation of MAPK
signaling.
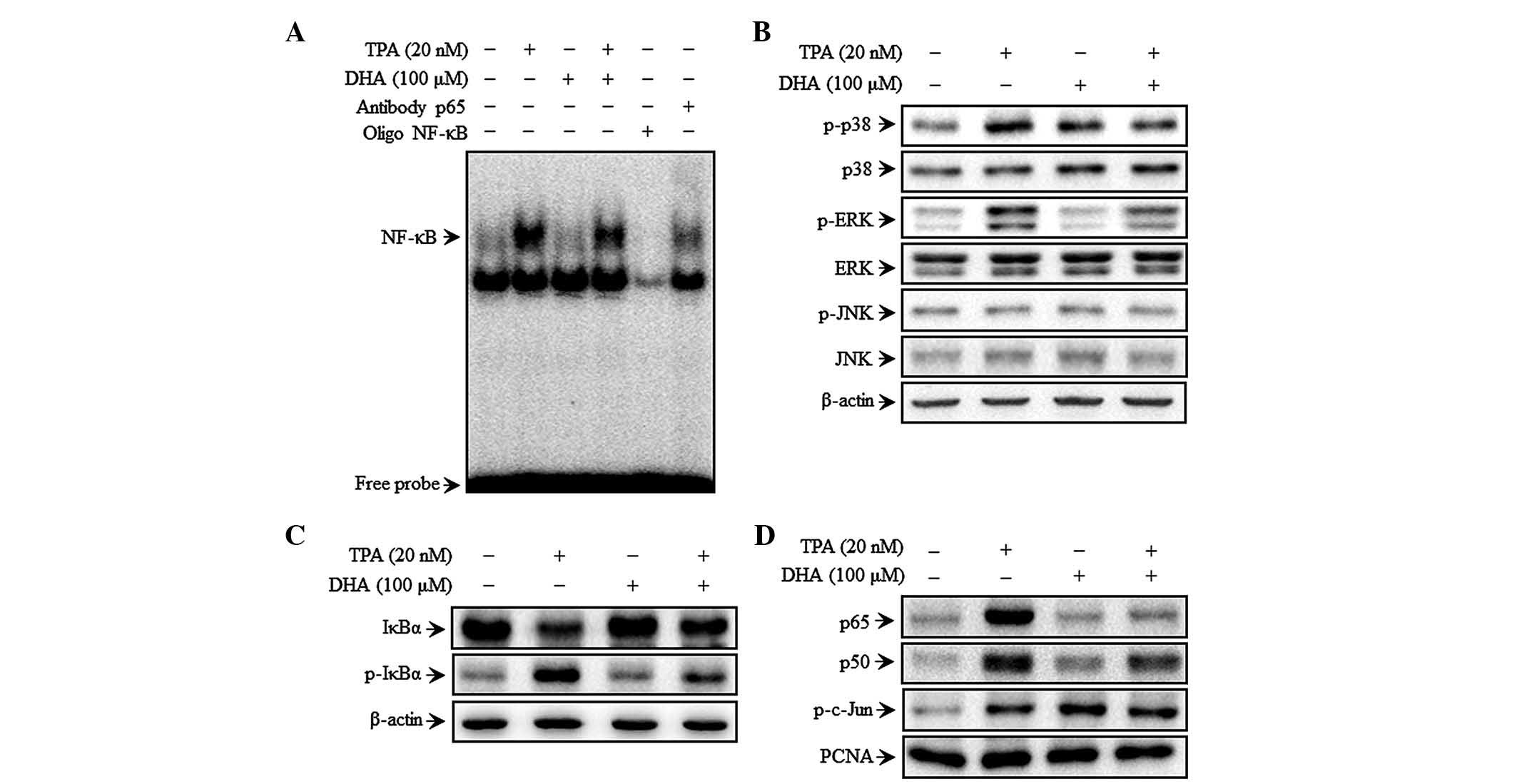 | Figure 2.DHA inhibits TPA-induced
transcriptional activation of matrix metalloproteinase-9 and p38
phosphorylation in MCF-7 cells. Cells were treated with DHA in the
presence of TPA for 4 h, followed by nuclear extraction. (A) NF-κB
DNA binding was analyzed by electrophoretic mobility shift assay.
(B) Cells were pretreated with TPA for 30 min in the presence or
absence of DHA, and cell lysates were prepared for western blotting
to detect p-p38, p38, p-JNK, JNK, p-ERK and ERK expression. (C)
Cytoplasmic levels of NF-κB subunits, IκBα and p-IκBα, were
determined by western blotting. (D) Nuclear levels of NFκB (p50 and
p65) and activator protein 1 (p-c-Jun) subunits were determined by
western blotting. TPA, 12-O-tetradecanoylphorbol-13-acetate;
DHA, docosahexaenoic acid; NF, nuclear factor; ERK, extracellular
signal-regulated kinase; JNK, c-Jun N-terminal kinase; IκB, nuclear
factor of κ light polypeptide gene enhancer in B cells inhibitor;
p, phosphorylated; PCNA, proliferating cell nuclear antigen. |
DHA increases PPAR-γ expression
DHA treatment increased PPAR-γ expression in a
dose-dependent manner in MCF-7 cells (Fig. 3A). To verify whether MMP-9 expression
is regulated by PPAR-γ, TPA-treated cells were treated concurrently
with DHA in the presence or absence of GW9662. MMP-9 expression was
higher upon treatment in the presence of GW9662 than following
treatment with DHA alone (Fig. 3B).
GW9662 also rescued the expression of p65 and p50 that was
abolished by DHA treatment (Fig. 3C).
These results indicate that DHA inhibits MAPK signaling and MMP-9
expression by increasing PPAR-γ expression.
DHA treatment inhibits TPA-induced
MCF-7 cell invasion
The upregulation of MMP-9 expression contributes to
cancer cell invasion (10,27). Therefore, the effect of DHA on the
invasive potential of MCF-7 cells was investigated by Matrigel
assay. TPA treatment stimulated MCF-7 cell invasion 10-fold more,
compared with untreated control cells. Treatment with DHA decreased
TPA-induced cell invasion by 60%. However, a decrease of only 30%
was observed in the presence of both GW9662 and DHA (Fig. 4), indicating that DHA suppresses the
invasive potential of breast cancer cells via PPAR-γ
activation.
Discussion
The protective effects of fish oil are attributable
to ω-3 polyunsaturated fatty acids (PUFAs), including EPA (20:5
ω-3) and DHA (22:6 n-3), which are converted by fish from
α-linolenic acid (LNA; 18:3 ω-3) present in ingested cold water
vegetation (1). A study of metastatic
mouse mammary carcinoma reported that a diet containing α-LNA-rich
linseed oil effectively arrested tumor progression (12). Furthermore, tumor growth and
metastasis were inhibited by diets containing fish oil, EPA or DHA
(27,35). ω-3 PUFAs may affect carcinogenesis by
altering transcription factor activity, gene expression and/or
signal transduction (7,36). DHA is a particularly potent enhancer
of tumor cell chemosensitivity (37).
However, there is little information on the anti-metastatic effects
of DHA in human breast cancer cells.
Metastasis is the primary cause of breast cancer
mortality, and involves cell proliferation, ECM degradation, cell
migration and tumor growth at secondary sites (19,20). Tumor
cell invasion is an early step in this process, representing the
transition from a benign state to malignancy (11). Tumor invasion is morphologically
distinguished by a distortion of the primary tumor edge where
individual or cohorts of tumor cells actively invade the
surrounding ECM (38). MMP-9 is a
critical molecule in tumor invasion and metastasis, and its
activation is associated with the progression of mammary tumors
(39). Inflammatory cytokines, growth
factors or phorbol esters stimulate MMP-9 by activating different
intracellular signaling pathways in breast cancer cells (40–42). Thus,
inhibiting the expression and/or activity of MMP-9 may be an
important strategy for slowing or preventing tumor metastasis.
AP-1 belongs to the basic region/leucine zipper
motif group of DNA-binding proteins, and homo or heterodimerizes in
response to signaling events to indirectly or directly activate
c-Jun and c-Fos expression (43–46). NF-κB
is a member of a family of inducible transcription factors that
regulate host inflammatory and immune responses (47) as a result of MAPK signaling, which
requires cell type-specific IκB kinase expression (17,24,25,48,49).
Both AP-1 and NF-κB have been implicated in TPA-mediated MMP-9 gene
induction (20,21). The present results demonstrate that
DHA inhibits TPA-induced MMP-9 expression via activation of NF-κB,
but not of AP-1, in MCF-7 breast cancer cells.
PPAR-γ is a member of the nuclear receptor and
ligand-activated steroid hormone receptor-regulated transcription
factor superfamily (26). ω-3 PUFAs
are natural ligands of nuclear receptors such as PPAR-α and -γ, and
can upregulate PPAR-γ expression (50). PPAR-γ has been reported to inhibit the
NF-κB signaling pathway by reducing NF-κB binding activity and
physically interacting with both p65 and p50 (51,26). The
present study confirmed that PPAR-γ expression was increased by
treatment with DHA, which consequently led to a decrease in p65 and
p50 levels, and a decrease in MMP-9 expression. NF-κB inhibition by
DHA was reversed by treatment with the PPAR-γ-specific antagonist
GW9662, demonstrating that TPA-induced tumor cell invasion was
suppressed by DHA through a PPAR-γ-dependent mechanism.
In conclusion, DHA is a potent inhibitor of
TPA-induced MMP-9 expression, and blocks breast carcinoma cell
invasion by modulating the NF-κB signaling pathway via PPAR-γ
upregulation. These findings suggest that DHA may be an effective
therapeutic agent for preventing breast tumor invasion and
metastasis.
Acknowledgements
The present study was supported by the Korea Atomic
Energy Research Institute (grant no. NRF-2015M2A2A6021673) and by a
National Research Foundation of Korea grant funded by the Korean
government (grant no. No-2011-0030130). The authors thank Editage
(Cactus Communications Co. Ltd., Shanghai, China) for English
language editing the present manuscript.
References
|
1
|
Stillwell W and Wassall SR:
Docosahexaenoic acid: Membrane properties of a unique fatty acid.
Chem Phys Lipids. 126:1–27. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirayama T: Epidemiology of breast cancer
with special reference to the role of diet. Prev Med. 7:173–195.
1978. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wynder EL, Rose DP and Cohen LA: Diet and
breast cancer in causation and therapy. Cancer. 58:(Suppl 8).
S1804–S1813. 1986. View Article : Google Scholar
|
|
4
|
Rose DP, Connolly JM and Coleman M: Effect
of omega-3 fatty acids on the progression of metastases after the
surgical excision of human breast cancer cell solid tumors growing
in nude mice. Clin Cancer Res. 2:1751–1756. 1996.PubMed/NCBI
|
|
5
|
Wu M, Harvey KA, Ruzmetov N, Welch ZR,
Sech L, Jackson K, Stillwell W, Zaloga GP and Siddiqui RA: Omega-3
polyunsaturated fatty acids attenuate breast cancer growth through
activation of a neutral sphingomyelinase-mediated pathway. Int J
Cancer. 117:340–348. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartsch H, Nair J and Owen RW: Dietary
polyunsaturated fatty acids and cancers of the breast and
colorectum: Emerging evidence for their role as risk modifiers.
Carcinogenesis. 20:2209–2218. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Larsson SC, Kumlin M, Ingelman-Sundberg M
and Wolk A: Dietary long-chain n-3 fatty acids for the prevention
of cancer: A review of potential mechanisms. Am J Clin Nutr.
79:935–945. 2004.PubMed/NCBI
|
|
8
|
Chambers AF and Matrisian LM: Changing
views of the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakajima M, Welch DR, Belloni PN and
Nicolson GL: Degradation of basement membrane type IV collagen and
lung subendothelial matrix by rat mammary adenocarcinoma cell
clones of differing metastatic potentials. Cancer Res.
47:4869–4876. 1987.PubMed/NCBI
|
|
10
|
Woessner JF Jr: Matrix metalloproteinases
and their inhibitors in connective tissue remodeling. FASEB J.
5:2145–2154. 1991.PubMed/NCBI
|
|
11
|
Velinov N, Poptodorov G, Gabrovski N and
Gabrovski S: The role of matrixmetalloproteinases in the tumor
growth and metastasis. Khirurgiia (Sofiia). 44–49. 2010.(In
Bulgarian). PubMed/NCBI
|
|
12
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: A tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gum R, Wang H, Lengyel E, Juarez J and
Boyd D: Regulation of 92 kDa type IV collagenase expression by the
jun aminoterminal kinase- and the extracellular signal-regulated
kinase-dependent signaling cascades. Oncogene. 14:1481–1493. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Newton AC: Regulation of protein kinase C.
Curr Opin Cell Biol. 9:161–167. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeigler ME, Chi Y, Schmidt T and Varani J:
Role of ERK and JNK pathways in regulating cell motility and matrix
metalloproteinase 9 production in growth factor-stimulated human
epidermal keratinocytes. J Cell Physiol. 180:271–284. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hozumi A, Nishimura Y, Nishiuma T, Kotani
Y and Yokoyama M: Induction of MMP-9 in normal human bronchial
epithelial cells by TNF-alpha via NF-kappa B-mediated pathway. Am J
Physiol Lung Cell Mol Physiol. 281:L1444–L1452. 2001.PubMed/NCBI
|
|
17
|
Weng CJ, Chau CF, Hsieh YS, Yang SF and
Yen GC: Lucidenic acid inhibits PMA-induced invasion of human
hepatoma cells through inactivating MAPK/ERK signal transduction
pathway and reducing binding activities of NF-kappaB and AP-1.
Carcinogenesis. 29:147–156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin CW, Hou WC, Shen SC, Juan SH, Ko CH,
Wang LM and Chen YC: Quercetin inhibition of tumor invasion via
suppressing PKC delta/ERK/AP-1-dependent matrix metalloproteinase-9
activation in breast carcinoma cells. Carcinogenesis. 29:1807–1815.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee SO, Jeong YJ, Kim M, Kim CH and Lee
IS: Suppression of PMA-induced tumor cell invasion by capillarisin
via the inhibition of NF-kappaB-dependent MMP-9 expression. Biochem
Biophys Res Commun. 366:1019–1024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oh JH, Chung AS, Steinbrenner H, Sies H
and Brenneisen P: Thioredoxin secreted upon ultraviolet A
irradiation modulates activities of matrix metalloproteinase-2 and
tissue inhibitor of metalloproteinase-2 in human dermal
fibroblasts. Arch Biochem Biophys. 423:218–226. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hong S, Park KK, Magae J, Ando K, Lee TS,
Kwon TK, Kwak JY, Kim CH and Chang YC: Ascochlorin inhibits matrix
metalloproteinase-9 expression by suppressing activator
protein-1-mediated gene expression through the ERK1/2 signaling
pathway: Inhibitory effects of ascochlorin on the invasion of renal
carcinoma cells. J Biol Chem. 280:25202–25209. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Woo MS, Jung SH, Kim SY, Hyun JW, Ko KH,
Kim WK and Kim HS: Curcumin suppresses phorbol ester-induced matrix
metalloproteinase-9 expression by inhibiting the PKC to MAPK
signaling pathways in human astroglioma cells. Biochem Biophys Res
Commun. 335:1017–1025. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eberhardt W, Huwiler A, Beck KF, Walpen S
and Pfeilschifter J: Amplification of IL-1 beta-induced matrix
metalloproteinase-9 expression by superoxide in rat glomerular
mesangial cells is mediated by increased activities of NF-kappa B
and activating protein-1 and involves activation of the
mitogen-activated protein kinase pathways. J Immunol.
165:5788–5797. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karin M: The regulation of AP-1 activity
by mitogen-activated protein kinases. J Biol Chem. 270:16483–16486.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Madrid LV, Mayo MW, Reuther JY and Baldwin
AS Jr: Akt stimulates the transactivation potential of the RelA/p65
Subunit of NF-kappa B through utilization of the Ikappa B kinase
and activation of the mitogen-activated protein kinase p38. J Biol
Chem. 276:18934–18940. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Delerive P, Fruchart JC and Staels B:
Peroxisome proliferator-activated receptors in inflammation
control. J Endocrinol. 169:453–459. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Daynes RA and Jones DC: Emerging roles of
PPARs in inflammation and immunity. Nat Rev Immunol. 2:748–759.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee JH, Woo JH, Woo SU, Kim KS, Park SM,
Joe EH and Jou I: The 15-deoxy-delta 12,14-prostaglandin J2
suppresses monocyte chemoattractant protein-1 expression in
IFN-gamma-stimulated astrocytes through induction of MAPK
phosphatase-1. J Immunol. 181:8642–8649. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li CC, Yang HT, Hou YC, Chiu YS and Chiu
WC: Dietary fish oil reduces systemic inflammation and ameliorates
sepsis-induced liver injury by up-regulating the peroxisome
proliferator-activated receptor gamma-mediated pathway in septic
mice. J Nutr Biochem. 25:19–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee YR, Noh EM, Oh HJ, Hur H, Kim JM, Han
JH, Hwang JK, Park BH, Park JW, Youn HJ, et al:
Dihydroavenanthramide D inhibits human breast cancer cell invasion
through suppression of MMP-9 expression. Biochem Biophys Res
Commun. 405:552–557. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim BS, Lee SH, Lee SJ, et al: Curcumin
suppresses the TPA-induced invasion through inhibition of
PKCalpha-dependent MMP-expression in MCF-7 human breast cancer
cells. Phytomedicine. 19:1085–1092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim MS, Lee SJ, Jung SH, et al: Suppression of
TPA-induced tumor cell invasion by sulfuretin via inhibition of
NF-κB-dependent MMP-9 expression. Oncol Rep. 29:1231–1237.
2013.PubMed/NCBI
|
|
33
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Altenburg JD, Bieberich AA, Terry C,
Harvey KA, Vanhorn JF, Xu Z, Jo Davisson V and Siddiqui RA: A
synergistic antiproliferation effect of curcumin and
docosahexaenoic acid in SK-BR-3 breast cancer cells: Unique
signaling not explained by the effects of either compound alone.
BMC Cancer. 11:1492011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim J, Lim SY, Shin A, Sung MK, Ro J, Kang
HS, Lee KS, Kim SW and Lee ES: Fatty fish and fish omega-3 fatty
acid intakes decrease the breast cancer risk: A case-control study.
BMC Cancer. 9:2162009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun H, Berquin IM, Owens RT, O'Flaherty JT
and Edwards IJ: Peroxisome proliferator-activated receptor
gamma-mediated up-regulation of syndecan-1 by n-3 fatty acids
promotes apoptosis of human breast cancer cells. Cancer Res.
68:2912–2919. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Scorilas A, Karameris A, Arnogiannaki N,
Ardavanis A, Bassilopoulos P, Trangas T and Talieri M:
Overexpression of matrix-metalloproteinase-9 in human breast
cancer: A potential favourable indicator in node-negative patients.
Br J Cancer. 84:1488–1496. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cho HJ, Kang JH, Kwak JY, Lee TS, Lee IS,
Park NG, Nakajima H, Magae J and Chang YC: Ascofuranone suppresses
PMA-mediated matrix metalloproteinase-9 gene activation through the
Ras/Raf/MEK/ERK- and Ap1-dependent mechanisms. Carcinogenesis.
28:1104–1110. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kajanne R, Miettinen P, Mehlem A, Leivonen
SK, Birrer M, Foschi M, Kahari VM and Leppa S: EGF-R regulates MMP
function in fibroblasts through MAPK and AP-1 pathways. J Cell
Physiol. 212:489–497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Srivastava AK, Qin X, Wedhas N, Arnush M,
Linkhart TA, Chadwick RB and Kumar A: Tumor necrosis factor-alpha
augments matrix metalloproteinase-9 production in skeletal muscle
cells through the activation of transforming growth
factor-beta-activated kinase 1 (TAK1)-dependent signaling pathway.
J Biol Chem. 282:35113–35124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Frigo DE, Tang Y, Beckman BS, Scandurro
AB, Alam J, Burow ME and McLachlan JA: Mechanism of AP-1-mediated
gene expression by select organochlorines through the p38 MAPK
pathway. Carcinogenesis. 25:249–261. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shaulian E and Karin M: AP-1 in cell
proliferation and survival. Oncogene. 20:2390–2400. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yokoo T and Kitamura M: Dual regulation of
IL-1 beta-mediated matrix metalloproteinase-9 expression in
mesangial cells by NF-kappa B and AP-1. Am J Physiol.
270:F123–F130. 1996.PubMed/NCBI
|
|
46
|
Lungu G, Covaleda L, Mendes O,
Martini-Stoica H and Stoica G: FGF-1-induced matrix
metalloproteinase-9 expression in breast cancer cells is mediated
by increased activities of NF-kappaB and activating protein-1. Mol
Carcinog. 47:424–435. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yamamoto Y and Gaynor RB: Therapeutic
potential of inhibition of the NF-kappaB pathway in the treatment
of inflammation and cancer. J Clin Invest. 107:135–142. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yao J, Xiong S, Klos K, Nguyen N, Grijalva
R, Li P and Yu D: Multiple signaling pathways involved in
activation of matrix metalloproteinase-9 (MMP-9) by heregulin-beta1
in human breast cancer cells. Oncogene. 20:8066–8074. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Amin AR Ruhul, Senga T, Oo ML, Thant AA
and Hamaguchi M: Secretion of matrix metalloproteinase-9 by the
proinflammatory cytokine, IL-1beta: A role for the dual signalling
pathways, Akt and Erk. Genes Cells. 8:515–523. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Marion-Letellier R, Déchelotte P, Iacucci
M and Ghosh S: Dietary modulation of peroxisome
proliferator-activated receptor gamma. Gut. 58:586–593. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Delerive P, Martin-Nizard F, Chinetti G,
Trottein F, Fruchart JC, Najib J, Duriez P and Staels B: Peroxisome
proliferator-activated receptor activators inhibit thrombin-induced
endothelin-1 production in human vascular endothelial cells by
inhibiting the activator protein-1 signaling pathway. Circ Res.
85:394–402. 1999. View Article : Google Scholar : PubMed/NCBI
|