Introduction
Ovarian cancer is a common malignant tumor of the
female reproductive system (1,2). As the
majority of patients with ovarian cancer have insidious onset and
methods of early diagnosis are lacking, 70% of patients are at an
advanced stage when the tumor is identified (FIGO stage III or IV)
and the 5-year survival rate is approximately 40% (3). The mortality rate ranks first in
gynecologic malignant tumors (4), and
it is extremely difficult to treat this disease.
Ovarian cancer has the characteristic that pelvic
cavity spreads extensively, and metastasis of cancer cells is
multifactorial and includes multiple steps. Interleukin-8 (IL-8) is
a CXC inflammatory chemokine that is highly expressed in tumor
microenvironment and has the functions of promoting migration and
tumor cell growth (5). IL-8 is
consistently highly expressed in patients with ovarian cancer and
the ascitic fluid level of IL-8 is obviously higher than serum
(6), suggesting that as an
inflammatory chemokine, IL-8 is involved in the occurrence and
development of ovarian cancer.
Experiments conducted in the present study aimed to
exclude interference of endogenous IL-8 through lentiviral
transfection technology, to observe its impact on invasion and
migration of ovarian carcinoma cells and explore preliminarily its
possible molecular mechanism to provide experimental basis for
molecular target therapy for ovarian carcinoma.
Materials and methods
Materials
Ovarian carcinoma SKOV3 cell lines were obtained
from the Shanghai Institute of Cell Library (Shanghai, China). The
small interfering RNA (siRNA) was designed and constructed as a
vector (plasmid without vector and bacterial strain containing this
plasmid were all produced and identified by Nanjing Moji Biological
Technology Development Co. Ltd., Nanjing, China). The main reagents
used were: CCK-8, IL-8, RPMI-1640 basal culture medium,
double-antibody and fetal bovine serum (FBS) (both from Gibco,
Grand Island, NY, USA), acrylamide, methylene acrylamide,
Tris-base, ammonium peroxydisulfate, TEMED, glycine and SDS (all
from Biosharp, Hefei, China). The PVDF membrane, and
chemiluminescent liquid were purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA).
Methods
siRNA and siNC were confirmed as effective target
sites by verifying the effective sequence from three sequences, and
it was designed as shRNA. pLV2-shSIRT1 and pLV2-scramble vectors
were constructed (Fig. 1). Cells
infected with nonsense sequences were the scramble group.
Non-infected cells served as the control group.
Plasmid was extracted by collecting 50 µl of fresh
bacteria fluid to inoculate with 30 ml LB culture medium
(containing moderate antibiotics), followed by shock culturing for
16 h at 37°C. The culture was centrifuged at 5,000 × g for 10 min,
at 25°C, to collect the thallus and absorb supernatant as much as
possible.
Buffer A1 (2.5 ml) was added and transferpettor
positive displacement pipette (Sigma-Aldrich, St. Louis, MO, USA)
was used to ensure that sedimentation of bacteria was resuspended.
Subsequently, 2.5 ml buffer B1 was used and reversed gently 10
times to mix evenly and allowed to stand for 5 min until the
solution was viscous and clear. Subsequently, 1 ml buffer N3 was
added, and mixed immediately 5 times, followed by vigorous
agitation 5 times, until white flocculent precipitate appeared. The
lysate was transferred to high-speed centrifuge at 12,000 × g for
10 min at room temperature. Then, 5 ml of 1 volume of buffer RET
(Biosharp) was added to 3 ml of 100% ethanol lysate and was
agitated vigorously 5 times to mix evenly. The DNA column was
centrifuged immediately and 6 ml lysate was transferred to a DNA
column with a collecting pipe immediately and centrifuged at 5,000
× g for 2 min at room temperature. The filtrate collected in the
pipe was discarded and column was placed back in collecting pipe,
this step was repeated until the appropriate solution passed
through the DNA column. Five milliliters of 70% ethanol was added
into the centrifugal column and centrifuged at 5,000 × g for 1 min
at room temperature. The filtrate collected in the pipe was
discarded and the column was placed back in the collecting pipe, a
step that was repeated once again. The centrifugal column was
placed back in the high-speed centrifuge at 5,000 × g for 10 min at
room temperature with open cover to remove residual ethanol.
The centrifugal column was placed in a new 15-ml
centrifuge tube. Endo-free elution buffer (Biosharp) was added in
middle of the DNA column membrane and kept for 10 min at room
temperature. The column was centrifuged again at 5,000 × g for 5
min to elute plasmid DNA. The eluent of the 15-ml centrifuge tube
was placed in the column, to elute for 1 min, and centrifuged at
5,000 × g for 5 min.
Virus transfection
Target cells (1–2×105) were inoculated in
a 6-well plate for one day prior to lentiviral transfection. The
medication treatment was given after 4 days. One milliliter of
virus supernatant (removed at −80°C in advance and melted on ice)
was added to the wells, and mixed gently. Then, 6–8 µl polybrene,
and cell state were added and observed after returning the cell
plate to the incubator for 6- to 8-h incubation, and 1 ml fresh
culture medium was added. After infecting for 24 h, fresh culture
medium was added. Puromycin (final concentration was 5 µg/ml) was
added into the wells and infected for 2 days to screen infected
SKOV3 cells, followed by replacement with new culture medium. After
1 week of culturing, positive cells were obtained.
Scratching test
A marker was used to draw horizontal lines under the
6-well plate uniformly, and a line approximately every 0.5 to 1 cm
and across wells. Each well was crossed by at least 5 lines.
Approximately 5×105 cells were added in the well, and a
specific number was different from cell to cell, by average
overnight. On the second day, a spearhead was used on the straight
edge, which was perpendicular to the scratched lines.
Phosphate-buffered saline (PBS) was used to wash the cells three
times, and the cells that had been drawn were removed and placed
back into serum-free medium. The cells were then incubated at 37°C,
in a 5% CO2 incubator, and a sample was taken at 0, 4,
24 and 48 h to capture images.
Transwell invasion test
The chambers were placed onto the cultured board.
Preheated serum-free medium (300 µl)was added in the upper chamber
and allowed to stand for 15–30 min at 25°C. Subsequently, the
inoculum was aspirated. After completion of digestion, the inoculum
was centrifuged and removed, the PBS was used twice to wash, and
serum-free medium containing BSA was used for resuspension. The
cell density was adjusted to 3×105. The cells were
inoculated by adding 200 µl of cell suspension to Transwell
chamber. Then, 500 µl of culture medium containing FBS was added to
the lower chamber of a 24-well plate and cultured for 14 h. After
0.1% crystal violet staining, the cells were observed using a
microscope (SZ61; Olympus, Tokyo, Japan), images were captured and
five horizons were randomly selected to count the cell numbers.
Western blot analysis and
antibodies
Western blot analysis was used to detect the
expression of IL-8 and p-NF-κB protein. When confluence of the
number of growing cells reached >80%, the culture medium was
removed. The prechilled PBS was used to wash the cells followed by
radioimmunoprecipitation assay (RIPA). The cells were homogenized
in RIPA buffer and subsequently centrifuged to extract total
protein. The extracted protein was preserved at −20°C.
Western blotting was performed as previously
reported (7). The primary rabbit
polyclonal IL-8 antibody (dilution, 1:500; cat. no. ab7747; Abcam,
Cambridge, MA, USA) was added to the membrane for incubation at 4°C
for 12 h. After incubation, the membrane was repeatedly washed in
0.2% TBST and incubated with secondary goat anti-rabbit (HRP) IgG
antibody (dilution, 1:2,000; cat. no. ab6721; Abcam) at 25°C.
Images of separated proteins were captured after chemiluminescence
reaction, and Gel-Pro gel Analyzer software (Silver Spring, MD,
USA) was used to detect brand intensity. GAPDH was used as the
internal control for normalization.
Statistical analysis
SPSS 19.0 software (IBM, Armonk, NY, USA) was used
for statistical analysis. All quantitative data were expressed as
means ± standard deviation. Comparison between groups was done
using one-way ANOVA test followed by post hoc test (Least
Significant Difference). P<0.05 indicated statistically
significant results.
Results
Impact of SKOV3 cells on the
expression of IL-8 after lentiviral transfection
It was found that in comparison to the control and
nonsense sequence groups (scramble group), the IL-8 expression
level of SKOV3 cells in the si-IL-8 group decreased following the
tranfection of si-IL-8 pLV2-shSIRT1, and its interference rate was
72.3% (p<0.01) (Fig. 2).
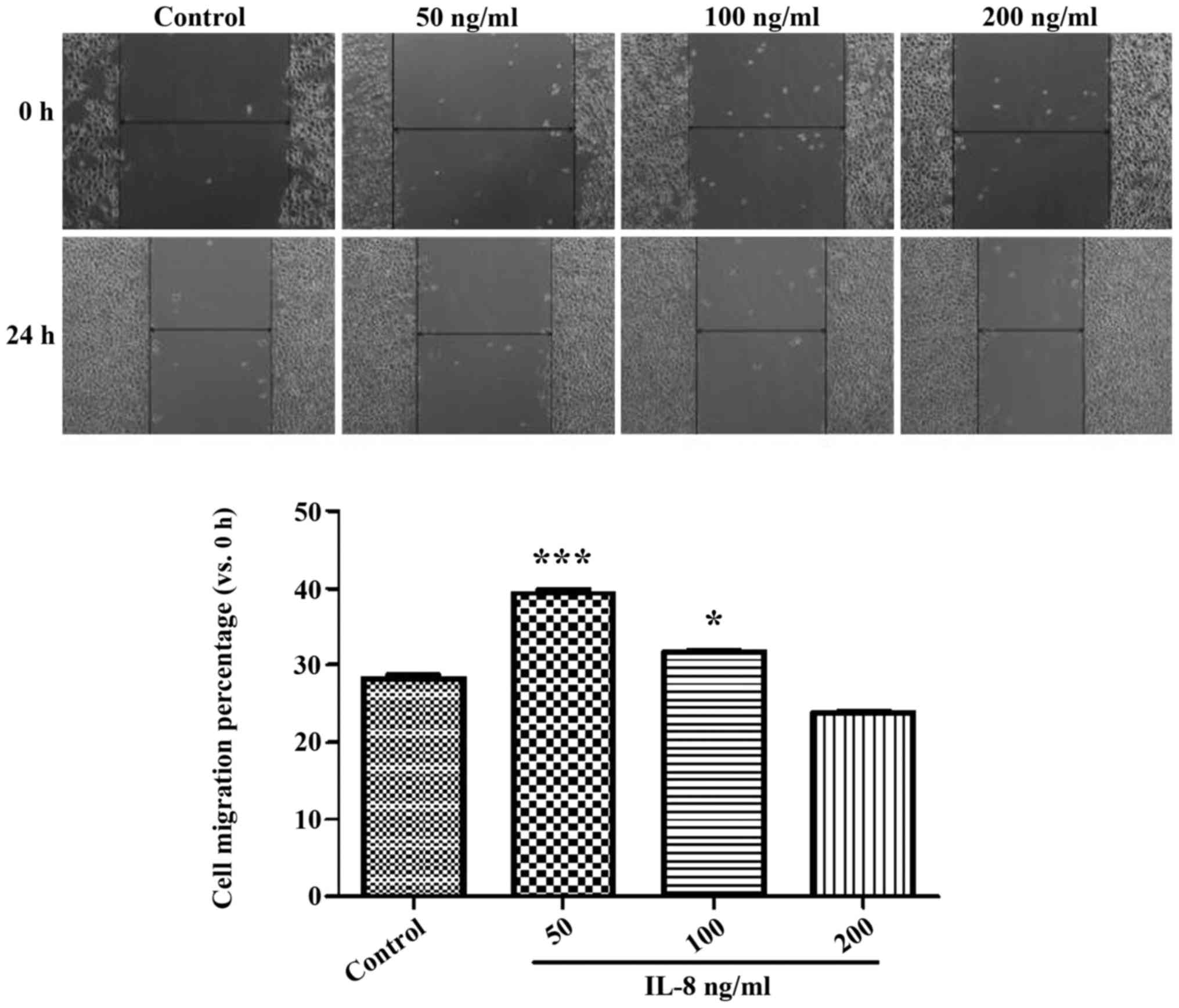
Impact of different concentrations of
IL-8 on the migration of SKOV3
The result of the experiment of CCK determined the
concentration of IL-8, which was set at 50 ng/ml as a low-dose
group, at 100 ng/ml as a medium-dose group and at 200 ng/ml as a
high-dose group. Fig. 3 shows the
width of the cell scratch in the IL-8 medium- and high-dose groups
was significantly higher than that of the control group
(p<0.05), and the 50 ng/ml group showed more obvious effects
(p<0.05). These results indicated that certain concentrations of
echogenic IL-8 can promote SKOV3 cell migration.
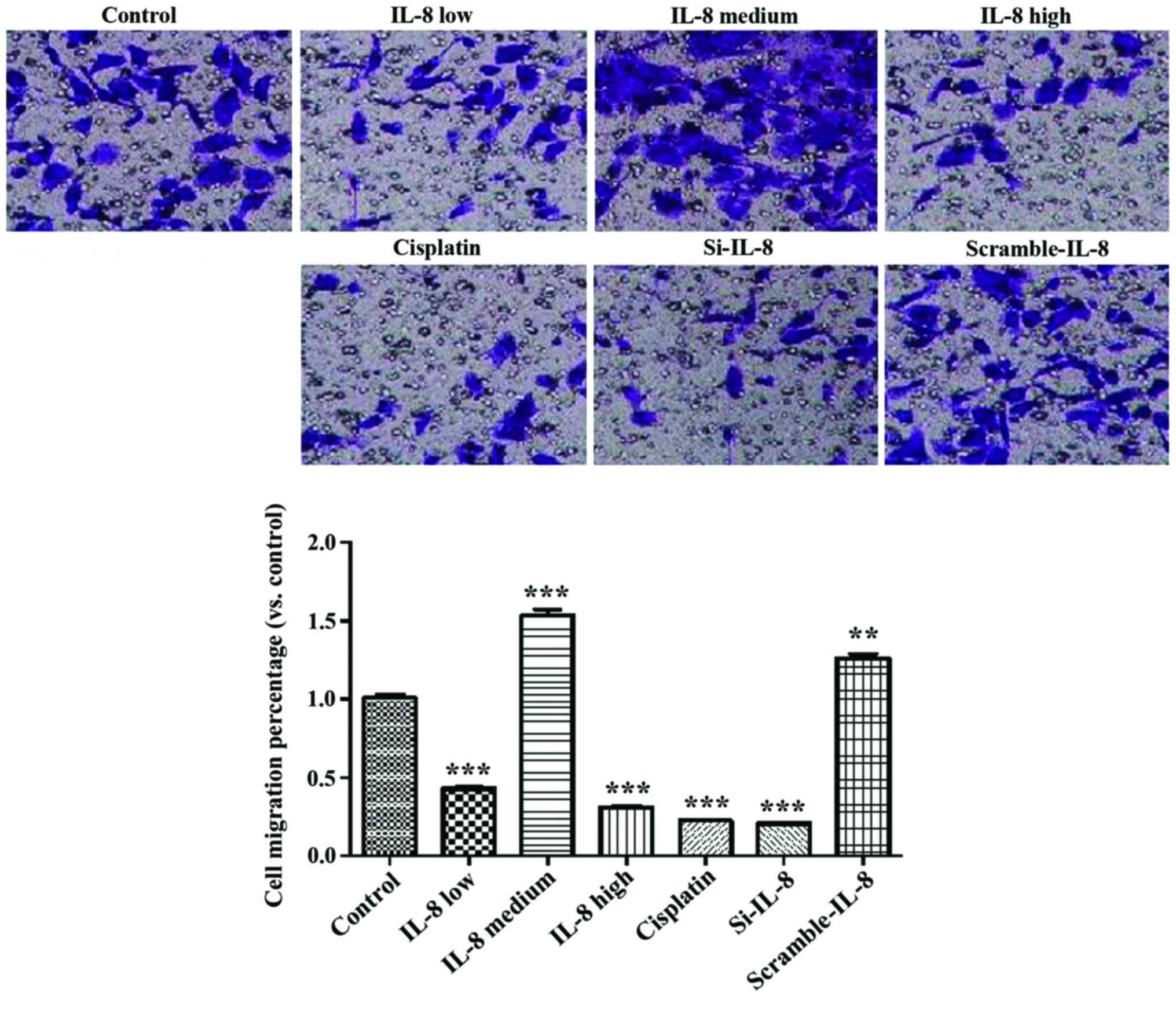
Impact of IL-8 on the invasion of
SKOV3 cells
Fig. 4 shows that
cisplatin and IL-8 silencing inhibited the invasion ability of
SKOV3 (p<0.01), and 100 ng/ml IL-8 enhanced the invasion ability
of SKOV3 (p<0.01).
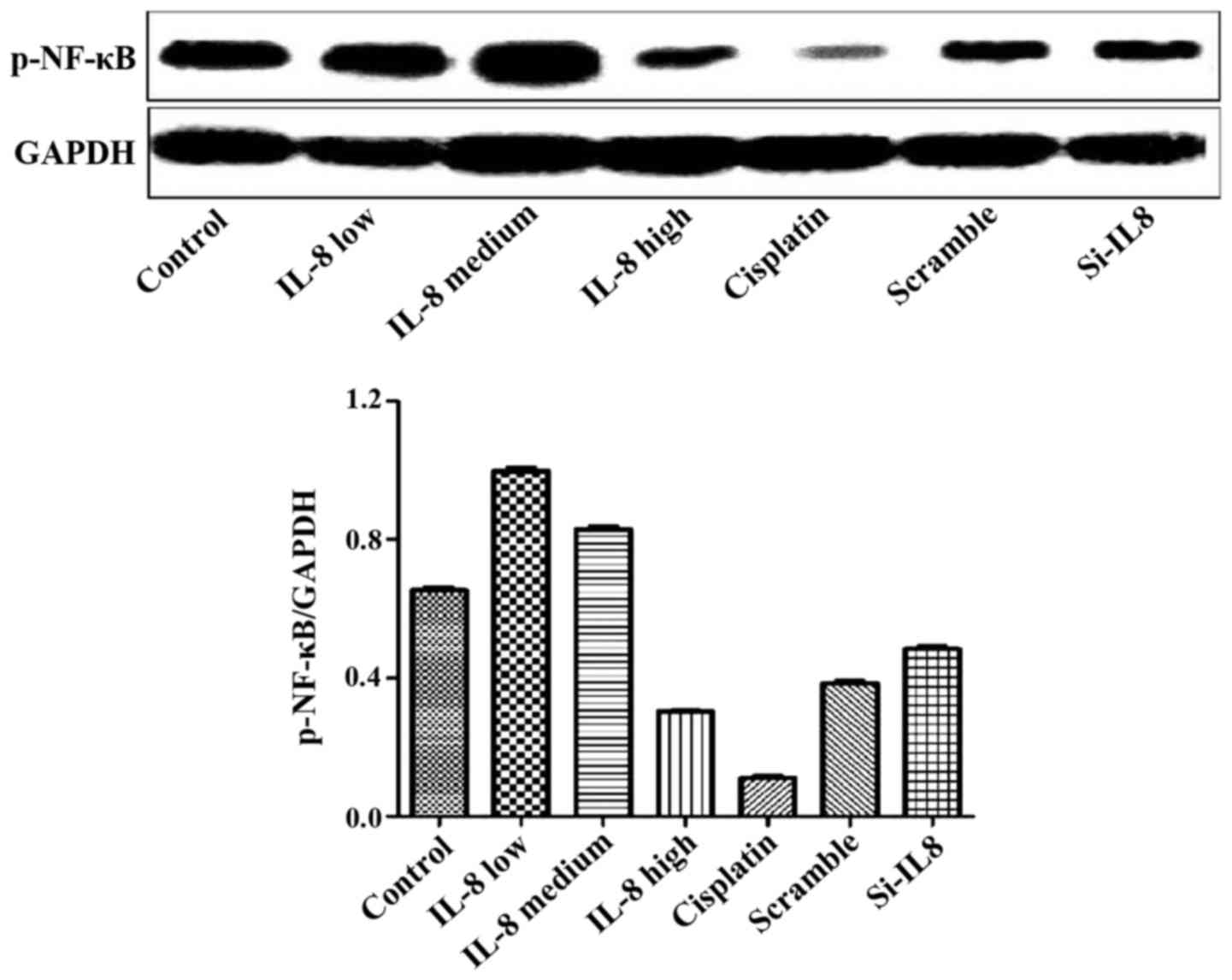
Detection of the expression of p-NF-κB
protein in each group
The experimental results are shown in Fig. 5. Cisplatin and IL-8 silencing
decreased the expression of p-NF-κB protein to some extent, and 100
ng/ml IL-8 strengthened the expression of p-NF-κB protein. However,
there was no statistical difference, suggesting that silencing of
IL-8 and the expression of p-NF-κB protein may not be linked.
Discussion
Ovarian cancer is a common female malignant tumor.
Since ovary is in the deep pelvic cavity and early lesions are not
easy to identify, most patients present at an advanced stage of
disease. Applications of ideal cytoreductive surgery and
chemotherapy regimens which are mainly platinum, have improved the
treatment effects of ovarian cancer obviously, albeit the 5-year
survival rate is only 30–40% (8) and
the mortality rate ranks first in gynecologic tumors. The main
reason is that 70% of patients are at an advanced stage at
diagnosis, and 70% of patients have recurrence after treatment
(8).
Therefore, research on better diagnosis and
therapeutic approaches is crucial for ovarian cancer. With the
rapid development of tumor immunology, biological treatment on
tumor may be an important supplement to surgery, chemotherapy,
radiotherapy and other traditional treatment methods (9). Use of biological treatment plays an
increasingly important role in controlling tiny remnant tumors,
delaying recurrence, increasing survival time, improving life
quality and other aspects. It is an antitumor therapy that has
developed rapidly during recent decade. Immunotherapy mainly
constitutes cytokines and its antitumor effect has been highly
valued. Thus, identifying target points of cytokines in the
proliferation and metastasis of ovarian cancer is crucial to
develop traditional treatments and improve the patient survival
rate.
Cisplatin is a metal fluor-complex of platinum and
can bind to DNA, leading to crosslinks of double strands of DNA,
thereby inhibiting cell mitosis and proliferation. It is a cell
cycle non-specific agent that can be applied to chemotherapy of
various types of tumors and acts as a first-line chemotherapeutic
agent to treat ovarian cancer (10).
Appropriate concentrations were screened through the cell scratch
test and mobility through screening at different time points.
Results were selected at 0 and 24 h to exclude the impact of
duration of cisplatin on this experiment.
It was found that TNF-α, IL-6, IL-8 and other
cytokines and chemotactic factors produced by inflammatory
corpuscle within the tumor microenvironment promote cell growth,
inhibit mutant cell apoptosis and play important roles in the
growth and metastasis of malignant tumors (11). According to lentiviral transfection,
targeting silent lentivirus infected and constructed silent SKVO3
cell lines of IL-8, avoiding the influence of endogenous IL-8. At
the same time, we included a blank control group, low-dose,
medium-dose, and high-dose IL-8 groups, cisplatin group, IL-8
silence group and nonsense sequence group. Results of the Transwell
invasion experiment showed that the cisplatin group and
IL-8-silenced group were able to inhibit tumor cell invasion, and
100 ng/ml IL-8 enhanced the invasion ability of cells, both of
which suggested that IL-8 produced by tumor cells plays an
important role in the cell invasion progress.
Inflammation is a pathophysiologic reaction that
concerns inflammatory cells and inflammatory cytokines. Previous
findings have shown that inflammation plays an important role in
the occurrence and development of tumors and the mechanism of
inflammation inducing tumors may contain two main aspects (12). First, inflammatory cells of tumor
microenvironment produce reactive oxygen species (ROS) and reactive
nitrogen cluster (NOS) which cause damage of cellular DNA.
Secondly, cytokines and chemotactic and other factors released by
inflammation cells enhance the signal of cell proliferation,
promote rapid proliferation and cell differentiation in order to
inhibit cell apoptosis (12).
Tumor-associated inflammation concerns the
transcription factor nuclear factor pathway (13). NF-κB engages in the differentiation,
proliferation and activation of immunological cells, and in the
growth control of some cells, has the function of anti-apoptosis
and plays a central role in the activation process of many
pro-inflammatory cytokines (14).
Expression of IL-8 is mainly regulated by activator protein-1
(AP-1) and NF-κB, and it needs to combine with the G
protein-coupled receptors, CXCR1 and CXCR2, to conduct biological
effects. Previous findings have shown that a high expression of the
content of IL-8 in a variety of malignant tumor tissues, such as
melanoma, gastroenteric tumor, cancerous goiter, ovarian cancer,
and lung carcinoma, was closely assoicated with the angiogenesis,
growth, metastasis, recurrence of tumors (15). Furthermore, the serum level of IL-8 of
patients with ovarian cancer prior to surgery was higher than that
in healthy individuals. As the autocrine growth factor of tumor
cells, IL-8 induces its own proliferation and promotes tumor growth
(16).
Signal channel of inducing cancer cells to secrete
IL-8 is complex. Inflammatory factors such as TNF-d and LPS can
pass the NF-κB signal channel and induce the expression of IL-8
(17). In addition, the MAPK
signaling channel is closely associated with IL-8, and after
changes of the p38MAPK, ERK1/2 phosphorylation level, the signaling
channels may have the ability of promoting the migration of tumor
cells by regulating the stability of IL-8 mRNA (18).
Cisplatin and silencing of IL-8 decreased the
expression of p-NF-κB protein while 100 ng/ml IL-8 enhanced the
expression of p-NF-κB protein (19).
Thus, IL-8 and cisplatin may inhibit the migration and
proliferation of SKOV3 ovarian cancer cells by activating the NF-κB
channel.
Multigenes and multiprocedures are involved in
development of malignant tumors, including ovarian cancer (20). Malignant tumor cells, especially
ovarian cancer which are difficult to detect, diffuse rapidly at an
advanced stage, while prognosis is delayed.
Thus, identification of appropriate therapeutic
targets is crucial in the latest evolution in the treatment of
ovarian cancer. Previous studies have reported the impact of IL-8
on the migration and invasion of tumor cells (21). However, those studies neglected that
SKOV3 cells themselves can produce IL-8 thus impacting cell
migration and invasion.
In the present study, the impact that the cells
themselves produced through lentiviral transfection technology was
not considered while it was verified that IL-8 has the function of
promoting the migration of ovarian cells. The study emphasizes the
role of IL-8 by suggesting that antibacterial therapy plays an
important role in complex treatments of ovarian cancer. From a
standpoint of controlling the metastasis of ovarian cancer, the
study provided new insight into treatments of ovarian cancer. At
the same time, IL-8 can be regarded as the key providing a new
approach for controlling the invasion and migration of ovarian
cancer.
References
|
1
|
Kim H, Wu R, Cho KR, Thomas DG, Gossner G,
Liu JR, Giordano TJ, Shedden KA, Misek DE and Lubman DM:
Comparative proteomic analysis of low stage and high stage
endometrioid ovarian adenocarcinomas. Proteomics Clin Appl.
2:571–584. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen WQ, Zhang SW, Zou XN and Zhao P:
Cancer incidence and mortality in China, 2006. Chin J Cancer Res.
23:3–9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Watanabe H, Iwase M, Ohashi M and Nagumo
M: Role of interleukin-8 secreted from human oral squamous cell
carcinoma cell lines. Oral Oncol. 38:670–679. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barshishat M, Ariel A, Cahalon L, Chowers
Y, Lider O and Schwartz B: TNFalpha and IL-8 regulate the
expression and function of CD44 variant proteins in human colon
carcinoma cells. Clin Exp Metastasis. 19:327–337. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee LF, Hellendall RP, Wang Y, Haskill JS,
Mukaida N, Matsushima K and Ting JP: IL-8 reduced tumorigenicity of
human ovarian cancer in vivo due to neutrophil infiltration. J
Immunol. 164:2769–2775. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ozols RF: Update on the management of
ovarian cancer. Cancer J. 8:(Suppl 1). S22–S30. 2002.PubMed/NCBI
|
|
10
|
Armstrong DK, Bundy B, Wenzel L, Huang HQ,
Baergen R, Lele S, Copeland LJ, Walker JL and Burger RA:
Gynecologic Oncology Group: Intraperitoneal cisplatin and
paclitaxel in ovarian cancer. N Engl J Med. 354:34–43. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu H, Ouyang W and Huang C: Inflammation,
a key event in cancer development. Mol Cancer Res. 4:221–233. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li QQ, Chen ZQ, Cao XX, Xu JD, Xu JW, Chen
YY, Wang WJ, Chen Q, Tang F, Liu XP, et al: Involvement of
NF-κB/miR-448 regulatory feedback loop in chemotherapy-induced
epithelial-mesenchymal transition of breast cancer cells. Cell
Death Differ. 18:16–25. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bromberg J and Wang TC: Inflammation and
cancer: IL-6 and STAT3 complete the link. Cancer Cell. 15:79–80.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartels M, Schweda AT, Dreikhausen U,
Frank R, Resch K, Beil W and Nourbakhsh M: Peptide-mediated
disruption of NFkappaB/NRF interaction inhibits IL-8 gene
activation by IL-1 or Helicobacter pylori. J Immunol.
179:7605–7613. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Subramaniam D, Ramalingam S, May R,
Dieckgraefe BK, Berg DE, Pothoulakis C, Houchen CW, Wang TC and
Anant S: Gastrin-mediated interleukin-8 and cyclooxygenase-2 gene
expression: differential transcriptional and posttranscriptional
mechanisms. Gastroenterology. 134:1070–1082. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee JW, Wang P, Kattah MG, Youssef S,
Steinman L, DeFea K and Straus DS: Differential regulation of
chemokines by IL-17 in colonic epithelial cells. J Immunol.
181:6536–6545. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Estrada Y, Dong J and Ossowski L: Positive
crosstalk between ERK and p38 in melanoma stimulates migration and
in vivo proliferation. Pigment Cell Melanoma Res. 22:66–76. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Padda RS, Gkouvatsos K, Guido M, Mui J,
Vali H and Pantopoulos K: A high-fat diet modulates iron metabolism
but does not promote liver fibrosis in hemochromatotic
Hjv−/− mice. Am J Physiol Gastrointest Liver Physiol.
308:G251–G261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mabuchi S, Ohmichi M, Nishio Y, Hayasaka
T, Kimura A, Ohta T, Saito M, Kawagoe J, Takahashi K,
Yada-Hashimoto N, et al: Inhibition of NFkappaB increases the
efficacy of cisplatin in in vitro and in vivo ovarian cancer
models. J Biol Chem. 279:23477–23485. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jones RG and Thompson CB: Tumor
suppressors and cell metabolism: A recipe for cancer growth. Genes
Dev. 23:537–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi J and Wei PK: Low-dose interleukin-8
induces the adhesion, migration and invasion of the gastric cancer
SGC-7901 cell line. Oncol Lett. 10:2871–2877. 2015.PubMed/NCBI
|