Introduction
Breast cancer is a common malignant tumor in women.
According to statistics, breast cancer accounts for 10% of
malignant tumors and its incidence is second to that of uterine
endometrial carcinoma (1). There are
several causes of breast cancer. Early detection is difficult,
women aged 40–60 years are at high risk for breast cancer, and its
incidence is highest during the peri-menopausal period (2). Since breast cancer relapses and
metastasizes easily, and has poor prognosis, there are great
challenges in diagnosis and treatment of breast cancer.
In recent years, obesity was shown to increase the
risk of a variety of diseases (3).
Additional studies revealed that obesity related genes, such as fat
mass and obesity-associated (FTO) are widely expressed in the human
body (4). The FTO gene was found to
be overexpressed in prostate cancer, pancreatic cancer, endometrial
cancer and liver cancer. Its overexpression affected the energy
metabolism of cancer cells, and was closely related to the
occurrence and development of cancer (5). To our knowledge, there are no studies on
the effect of FTO gene expression on breast cancer cell energy
metabolism. In the present study, we used breast cancer cells as a
model to explore the relationship between FTO gene expression and
energy metabolism, and performed preliminary studies on its
mechanism of action, to provide a new potential target for the
treatment and diagnosis of breast cancer.
Materials and methods
Cells
The human breast cancer cell lines (MCF-7 and
MDA-MB-231), and human breast cells (HCC1937) purchased from Cell
Bank of Chinese Academy of Sciences (Shanghai, China) were used.
Additional instruments and reagents used are shown in Table I. MCF-7, MDA-MB-231 and HCC1937 cells
were removed from storage in liquid nitrogen and thawed in a water
bath set to 37°C. Cells were then added to culture medium
[Dulbecco's modified Eagle's medium/F12 (DMEM/F12) supplemented
with 5% fetal bovine serum (FBS), 0.1 mg/ml streptomycin, 100 U/ml
penicillin and 2 mmol/l glutamine]. Cells were grown in culture
bottles in an incubator (37°C, 5% CO2) for 48 h. Culture
medium was removed when cells reached 90% confluence. Cells were
trypsinized in 0.25% trypsin, and centrifuged at 800 × g for 10 min
at room temperature. Cells were washed with DMEM/F12 and then
seeded again in cell culture bottles.
 | Table I.Major instruments and reagents. |
Table I.
Major instruments and reagents.
| Instruments and
reagents | Sources |
|---|
| Enzyme-labeled
instrument | Nanjing Detie
Laboratory Equipment Co., Ltd., Nanjing, China |
| Ultraviolet
spectrophotometer | Thermo Fisher
Scientific, Inc., Waltham, MA, USA |
| CO2
incubator | Sanyo, Tokyo,
Japan |
| Laminar flow
cabinet | Suzhou Purification
Equipment Co., Ltd., Suzhou, China |
| Inverted
microscope | Nikon, Tokyo,
Japan |
| PCR instrument | Beckman Coulter,
Inc., Brea, CA, USA |
| Centrifuge | Hunan Hengnuo
Instrument Equipment Co., Ltd., Changsha, China |
| RevertAid First
Strand cDNA Synthesis kit | Beyotime Institute of
Biotechnology, Haimen, China |
| DMEM/F12 culture
medium | Sigma-Aldrich, St.
Louis, MO, USA |
| Lactic acid test
kit | Sigma-Aldrich, St.
Louis, MO, USA |
| ATP content test
kit | Sigma-Aldrich, St.
Louis, MO, USA |
| Pyruvate kinase test
kit | Sigma-Aldrich, St.
Louis, MO, USA |
| Hexokinase test
kit | Sigma-Aldrich, St.
Louis, MO, USA |
| Real-time fluorescent
quantitative PCR kit | Thermo Fisher
Scientific, Inc., Waltham, MA, USA |
| Agarose | Thermo Fisher
Scientific, Inc., Waltham, MA, USA |
| Antibody
dilution | MultiSciences
(Lianke) Biotech Co., Ltd., Hangzhou, China |
| FBS | Hangzhou Sijiqing
Biology Engineering Materials Co., Ltd., Hangzhou, China |
| Protein concentration
test kit | Beyotime Institute of
Biotechnology, Hangzhou, China |
| Mycillin | Sigma-Aldrich, St.
Louis, MO, USA |
| Trypsin | Sigma-Aldrich, St.
Louis, MO, USA |
| RNA isolating reagent
kit | Beyotime Institute of
Biotechnology, Haimen, China |
| Cell total protein
extraction kit | Beyotime Institute of
Biotechnology, Haimen, China |
| PBS | SinoBio Biotech Co.,
Ltd., Shanghai, China |
| P13K monoclonal
antibody | Abcam, Cambridge, MA,
USA |
| p-P13K monoclonal
antibody | Abcam, Cambridge, MA,
USA |
| AKT monoclonal
antibody | Abcam, Cambridge, MA,
USA |
| p-AKT monoclonal
antibody | Abcam, Cambridge, MA,
USA |
|
HRP-anti-antibody | Abcam, Cambridge, MA,
USA |
RT-PCR
TRIzol was added to cell lysis buffer for lysis of
MCF-7, MDA-MB-231 and HCC1937 cells that were in the logarithmic
growth phase. After 5 min of digestion, lysates were placed in new
Eppendorf (EP) tubes (Corning, Inc., Corning, NY, USA), and 200 µl
chloroform was added. Solutions were shaken up and down 15 times,
and placed at room temperature for 5 min. Samples were then
centrifuged (8,500 × g, 4°C, 10 min). RNA in the supernatant was
transferred to new EP tubes. Next, 75% ethanol was added and
samples were centrifuged (6,500 × g, 4°C, 5 min). The supernatant
was discarded, and solutions were placed on a super-clean worktable
to air dry. DEPC water (Biosharp, Hefei, China) was then added and
mixed well. The concentration and purity of RNA were determined by
UV spectrophotometry. According to the instructions of the reverse
transcription kit, RNA was reverse transcribed into cDNA. Real-time
PCR amplification of cDNA was performed to measure the expression
of FTO mRNA in each group of cells.
Transfection
After trypsinization of MDA-MB-231 cells in the
logarithmic growth phase, cells were washed with DMEM/F12, cell
growth medium was added, and the cell concentration was adjusted to
2×105/ml. Cells were then seeded in 6-well culture
plates, and placed at 37°C, 5% CO2 for 48 h. When cells
were 50% confluent, growth medium was replaced with incomplete
culture medium (without FBS), and placed at 37°C, 5% CO2
for 1 h. The incomplete, serum-free medium was mixed with miFTO
inhibitor or inhibitor control, and incubated at 37°C for 5 min
(solution A). The serum-free medium was mixed with Lipofectamine
2000 (solution B) (KeyGen, Nanjing, China). Next, solutions A and B
were mixed, and left to incubate at room temperature for 20 min.
The cell culture medium was discarded, and cells were repeatedly
washed with phosphate-buffered saline (PBS). The transfection
reagent and miRNA were added to the cell culture plates, and cells
were placed at 37°C, 5% CO2 for 6 h. The medium was
replaced with complete culture medium, and cells were left in the
incubator for an additional 48 h.
Measurement of lactic acid content in
culture medium of breast cancer cells after transfection
MDA-MB-231 cells were transfected with miFTO
inhibitor, or inhibitor control and left to incubate at 37°C, 5%
CO2 for 48 h. Culture supernatant was harvested and
transferred to EP tubes, and centrifuged (800 × g, 5 min). The
content of lactic acid in supernatant was measured according to the
instructions of the Sigma Lactic Acid Test kit (Sigma-Aldrich, St.
Louis, MO, USA).
Detection of ATP content in
transfected cells
Transfected MDA-MB-231 cells grown in the incubator
(37°C, 5% CO2) for 48 h were trypsinized and washed with
PBS. Cell culture medium was added to resuspend the cells, then
cells were transferred to EP tubes and centrifuged (800 × g, 5
min). The supernatant was discarded. Cells were washed twice with
PBS and the supernatant was discarded after centrifugation at 800 ×
g for 3 min. The cells were mixed homogeneously with ultrapure
water. The cell homogenates were transferred to EP tubes and heated
in a water bath (100°C, 10 min). The content of ATP was determined
according to the instructions of the ATP test kit.
Detection of hexokinase and pyruvate
kinase activity in breast cancer cells
MDA-MB-231 cells were transfected with miFTO
inhibitor or inhibitor control for 48 h. The activity of hexokinase
and pyruvate kinase were detected according to the instructions of
the hexokinase and pyruvate kinase test kits.
Western blot analysis
After 48 h culture, culture medium from transfected
MDA-MB-231 cells was discarded, cells were washed with PBS and
lysed on ice for 30 min. Protein extracts were mixed with loading
buffer, and boiled at 100°C for 5 min. A total volume of 50 µl of
the denatured protein samples were loaded on gels (12% separation
gel and 5% spacer gel). A voltage of 80 V was applied to samples
and then adjusted to 120 V when protein reached the separation gel.
When bromophenol blue entered the separation gel, electrophoresis
was stopped. Protein was transferred to PVDF membranes overnight at
4°C. PVDF membranes were washed with TBST 3 times, and blocked with
skim milk for 2 h at 37°C. Membranes were treated with primary
antibody overnight at 4°C, washed with TBST, and incubated with
secondary antibody for 1.5 h at room temperature. Membranes were
washed and developed. Protein expression was analyzed with the
Odyssey scanning system (LI-COR, Inc., Lincoln, NE, USA). Primary
rabbit polyclonal AKT antibody (dilution, 1:500; cat. no. ab38449);
rabbit monoclonal p-AKT antibody (dilution, 1:500; cat. no.
ab81283), rabbit monoclonal PI3K antibody (dilution, 1:500; cat.
no. ab86714), rabbit polyclonalp-PI3K antibody (dilution, 1:500;
cat. no. ab182651) and secondary goat anti-rabbit (HRP) IgG
antibody (dilution, 1:2,000; cat. no. ab6721) were all purchased
from Abcam (Cambridge, MA, USA).
Statistical analysis
Data were analyzed with SPSS 16.0 statistical
software (SPSS, Inc., Chicago, IL, USA). Comparisons between groups
were by t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
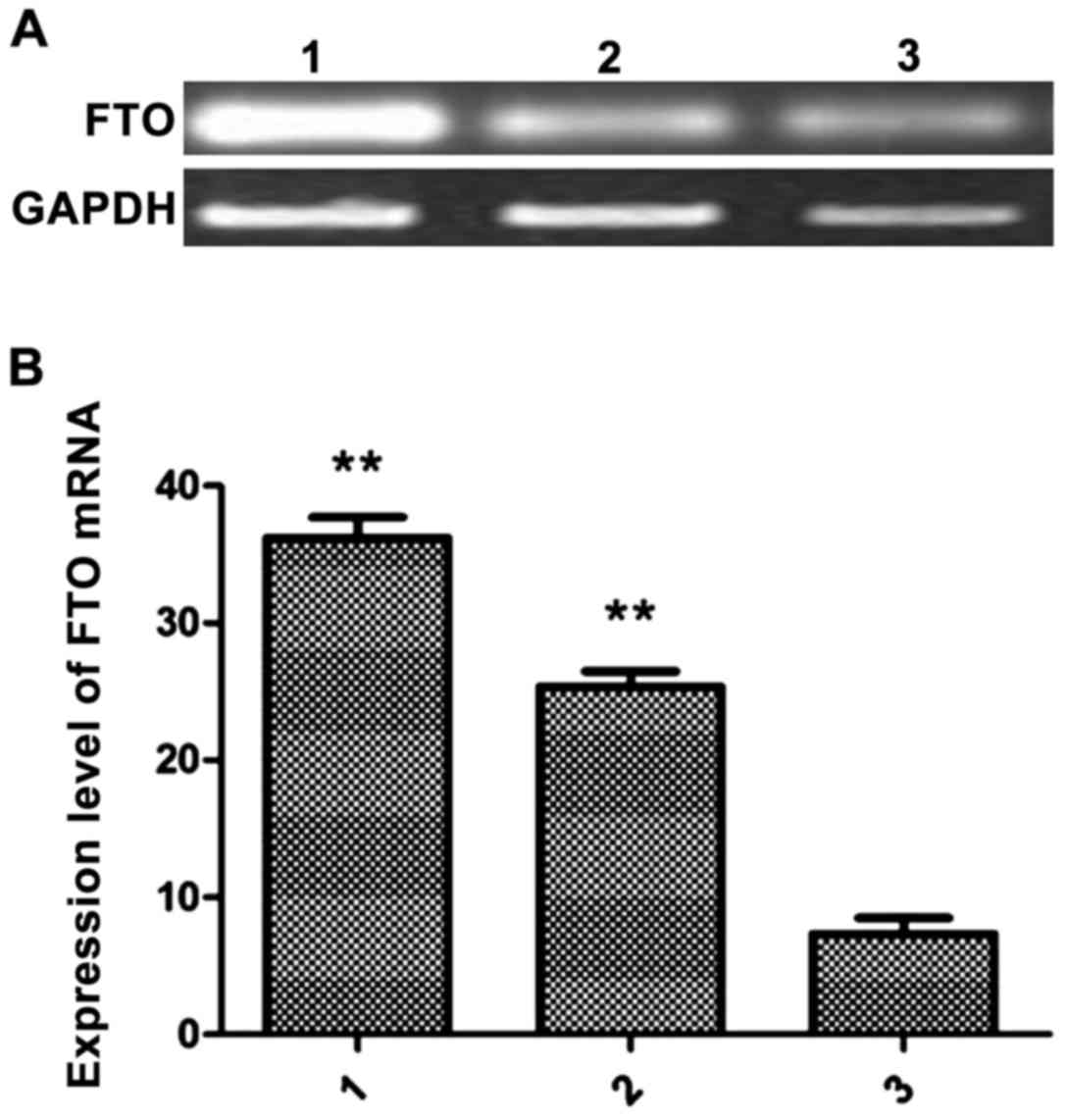
Measurement of FTO mRNA expression in
breast cancer cells and breast cells by RT-PCR
In the logarithmic growth phase, MCF-7, MDA-MB-231
and HCC1937 cells were harvested for extraction of total RNA. The
expression levels of FTO in the 3 groups of cells were detected by
real-time quantitative PCR. The relative expression levels of FTO
mRNA in MCF-7 cells was 26.89±2.31, 36.23±2.91 in MDA-MB-231 cells
and 8.96±3.01 in HCC1937 cells. The levels of FTO mRNA in MCF-7 and
MDA-MB-231 cells were significantly higher than in HCC1937 cells
(P<0.01) (Fig. 1).
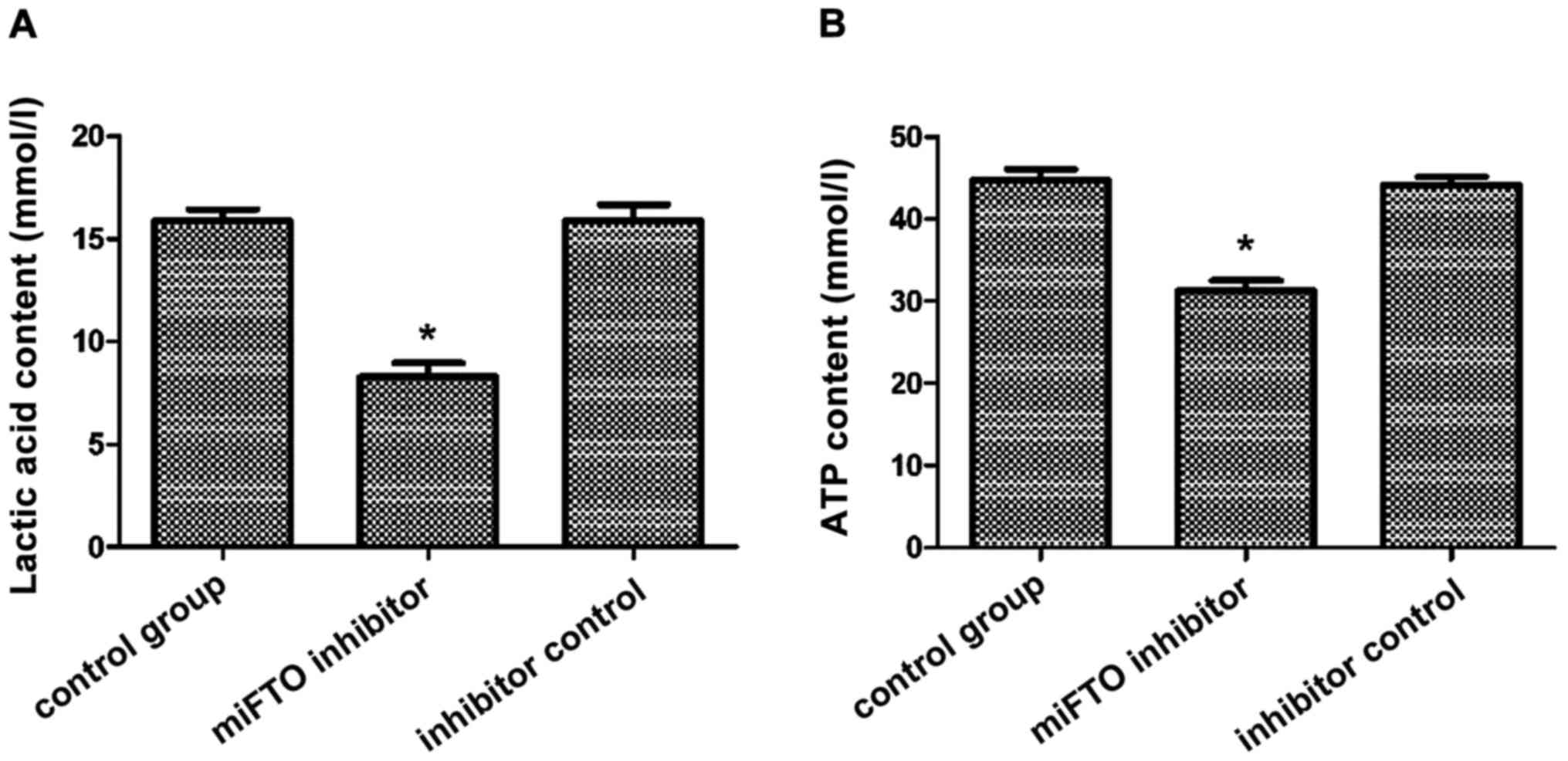
Measurement of lactic acid and ATP
content
The content of lactic acid and ATP in MDA-MB-231
cells transfected with the miFTO inhibitor or inhibitor control
were detected according to the lactic acid test kit and ATP test
kit. After 48 h culture, the lactic acid content of the miFTO
inhibitor group was 8.97±0.25 mmol/l, the lactic acid content of
the inhibitor control group was 17.11±1.02 mmol/l, and the lactic
acid content of the blank control group was 17.08±1.32 mmol/l. The
lactic acid content of breast cancer cells transfected with miFTO
inhibitor was significantly lower compared with the control group
and inhibitor control group. FTO mRNA inhibitors can inhibit the
production of lactic acid in breast cancer cells (Fig. 2A). After 48 h culture, the ATP content
of the miFTO inhibitor group was 31.45±1.58 mmol/l, the ATP content
of the inhibitor control group was 44.12±3.12 mmol/l, and the ATP
content of the blank control group was 44.56±2.45 mmol/l. The ATP
content of breast cancer cells transfected with the miFTO inhibitor
was significantly lower compared with the control group and
inhibitor control group (P<0.05). FTO mRNA inhibitors can
inhibit the production of ATP in breast cancer cells (Fig. 2B).
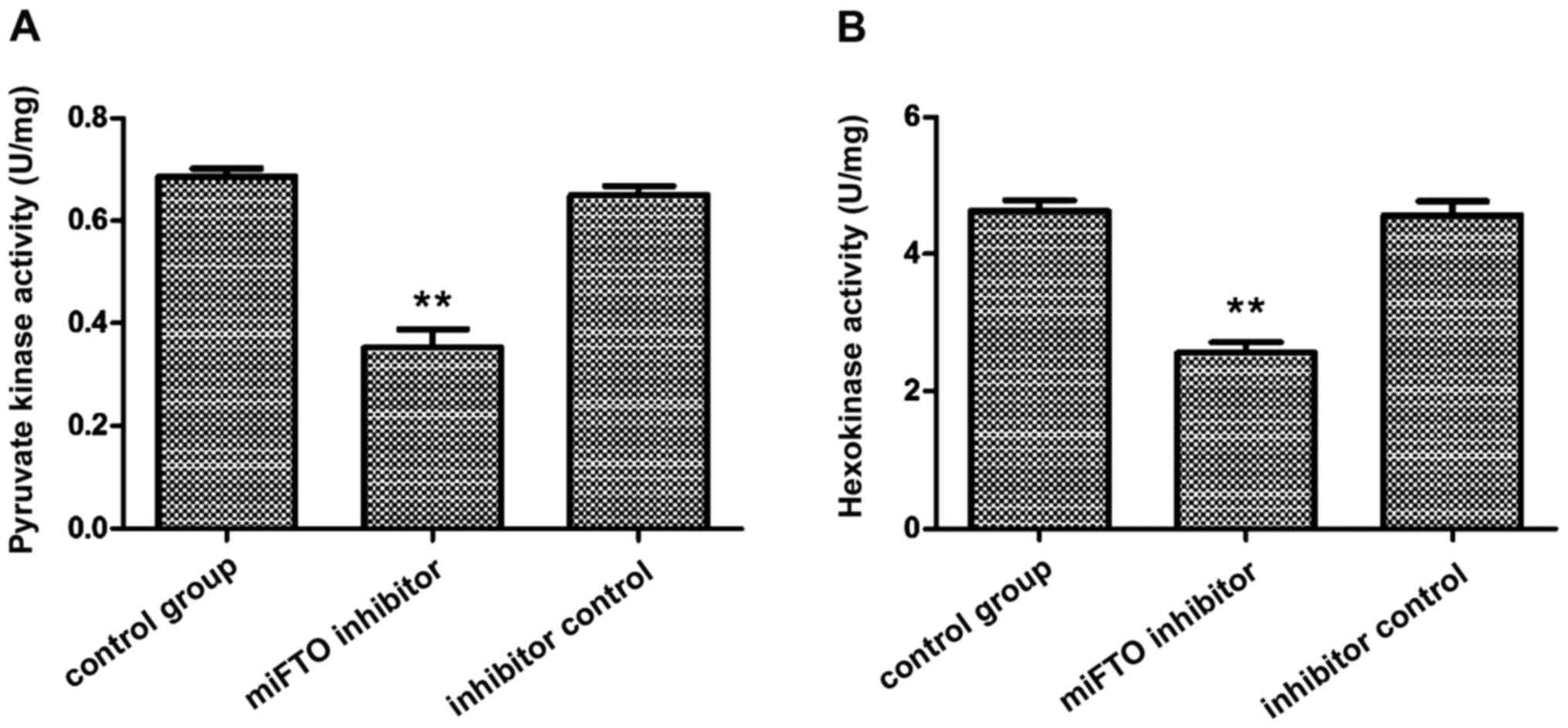
Detection of pyruvate kinase and
hexokinase activity
The activity of hexokinase and pyruvate kinase in
MDA-MB-231 cells transfected with miFTO inhibitor or inhibitor
control were detected according to the instructions of the
hexokinase and pyruvate kinase test kits. The results showed that
pyruvate kinase activity in breast cancer cells transfected with
miFTO inhibitor and inhibitor control were 0.39±0.01 and 0.68±0.02,
respectively, and 0.71±0.03 in the nontransfected cells. Pyruvate
kinase activity of breast cancer cells transfected with miFTO
inhibitor was significantly lower compared with the control group
and inhibitor control group (P<0.01). Hexokinase activity in
breast cancer cells transfected with the miFTO inhibitor and
inhibitor control were 2.54±0.21 and 4.86±0.25, respectively, and
4.84±0.20 in the nontransfected cells. Hexokinase activity of
breast cancer cells transfected with the miFTO inhibitor was
significantly lower compared with the control group and inhibitor
control group (P<0.01). Therefore, miFTO inhibitors can reduce
the activity of hexokinase and pyruvate kinase in breast cancer
cells (Fig. 3).
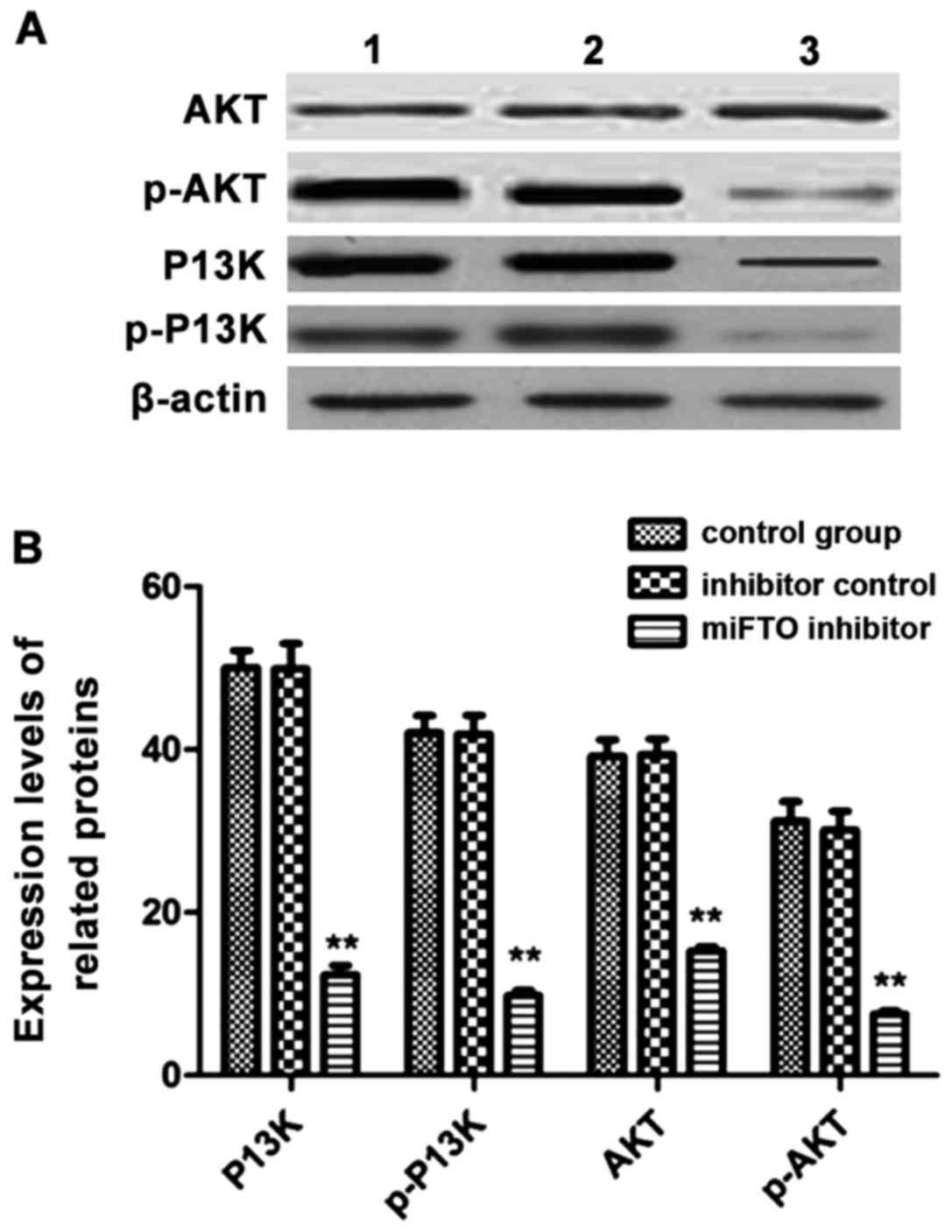
Western blot analysis to detect the
expression levels of related proteins in cells
MDA-MB-231 cells transfected with the miFTO
inhibitor or inhibitor control for 48 h were harvested, and lysates
were used to analyze the expressions of phosphatidylinositol
3-kinase (PI3K), p-PI3K, protein kinase B (Akt), and p-AKT by
western blot analysis. After transfection with the miFTO inhibitor,
the expressions of PI3K, p-PI3K, AKT and p-AKT were significantly
lower compared with the control group and inhibitor control group.
The phosphorylated forms of PI3K and AKT decreased significantly
(Fig. 4).
Discussion
Breast cancer is a common malignant tumor in women.
According to statistics, the incidence of breast cancer accounts
for 10% of all malignant tumors. There are 1.3 million newly
diagnosed cases of breast cancer worldwide every year, and ~50
million people die from breast cancer each year. Therefore, breast
cancer is a serious threat to the health of women. The occurrence
of breast cancer has regional differences. The morbidity of breast
cancer in developed countries is higher than that in developing
countries (6). There are several
causes of breast cancer and it is difficult to detect in the early
stage. Research on the pathogenesis of breast cancer is important
for treatment and diagnosis.
In recent years, studies have shown that obesity can
increase the risk of breast cancer. Obese women were 3 times more
likely to have breast cancer than nonobese women (3). The lipid metabolism gene FTO, has been
found to be closely related to obesity. The FTO gene contains nine
exons and is located on chromosome 16. It is widely expressed in
adults and in the fetus. FTO is most highly expressed in the
pituitary, pancreatic islets, hypothalamus and adrenal glands
(7,8).
FTO is overexpressed in prostate cancer, pancreatic cancer,
hepatocellular carcinoma and endometrial carcinoma. It therefore
has a close relationship with the occurrence and development of
cancer. In this study, we used human breast cancer cells (MCF-7 and
MDA-MB-231) and human breast cells (HCC1937) to determine the
levels of FTO mRNA by real-time fluorescence quantitative PCR. The
results showed that the FTO mRNA levels in breast cancer cells were
significantly higher than in normal breast cells, suggesting that
FTO is an oncogene, which represents a potential new marker for the
early diagnosis of breast cancer.
Cell energy metabolism is the process of
transforming organic matter into energy. In normal cells, ATP is
produced by oxidative decomposition of glucose, which can be
divided into aerobic oxidation and glycolysis (9,10). Glucose
can be oxidized to produce ATP under aerobic conditions, while
under anoxic conditions, ATP can be generated by glycolysis
(11). The energy metabolism of tumor
cells is different from that of normal cells. In cases where tumor
cells receive sufficient oxygen, energy is also produced by
glycolysis which converts pyruvate to lactic acid (the ‘Warburg
effect’) (12). Studies have shown
that aerobic glycolysis can be found in lung cancer, breast cancer,
colon cancer and renal cancer cells (13). Pyruvate kinase and hexokinase play a
key role in glycolysis. In tumor cells, hexokinase exists as
isozymes. The expression of hexokinase is related to the occurrence
and development of colon cancer and renal cell carcinoma.
Downregulated expression of pyruvate kinase can inhibit the
production of lactic acid by glycolysis (14). In this study, after cells were
transfected with the FTO mRNA inhibitor, the ATP levels in breast
cancer cells decreased, pyruvate kinase and hexokinase activity
decreased significantly, and the content of lactic acid in the
medium decreased significantly. These results demonstrate that
overexpression of the FTO gene could promote glycolysis in breast
cancer cells.
The role of the PI3K/AKT signaling pathway in tumor
cells is an area of intense study (15,16). The
PI3K/AKT signaling pathway is related to the proliferation and
apoptosis of cancer cells, and can regulate the activity of
caspase-9 (17), p53 (18), Bad (19), and other proteins, and inhibit
apoptosis. The PI3K/AKT signaling pathway is active in several
types of cells, and it plays an important role when cells are under
hypoxic conditions. Under hypoxic conditions, the PI3K/AKT
signaling pathway can upregulate insulin, epidermal growth factor
and cytokine expression. It can deliver messages to protein
tyrosine kinases via transmembrane receptors, activate PI3K, and
catalyze the generation of PIP3, which then delivers messages to
Akt, and activates the Ras-MAPK signaling pathway, which
consequently causes a series of complex reactions in the body
(20,21). In this study, through transcriptional
inhibition of the FTO gene, the protein expression of PI3K, p-PI3K,
Akt and p-Akt in cells increased significantly according to western
blot analysis, demonstrating that expression of the FTO gene
affected the energy metabolism of breast cancer cells through the
PI3K/AKT signaling pathway.
In conclusion, overexpression of FTO in breast
cancer cells can result in upregulation of pyruvate kinase and
hexokinase activity, increase the amount of ATP generation in
cells, and promote glycolysis and lactic acid production. FTO
overexpression affects the energy metabolism of breast cancer
cells, and the mechanism is related to the PI3K/AKT signaling
pathway. Our results represent a potential new therapeutic option
for the treatment and diagnosis of breast cancer.
References
|
1
|
Piccart-Gebhart MJ, Procter M,
Leyland-Jones B, Goldhirsch A, Untch M, Smith I, Gianni L, Baselga
J, Bell R, Jackisch C, et al: Herceptin Adjuvant (HERA) Trial Study
Team: Trastuzumab after adjuvant chemotherapy in HER2-positive
breast cancer. N Engl J Med. 353:1659–1672. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dowsett M, Forbes JF, Bradley R, Ingle J,
Aihara T, Bliss J, Boccardo F, Coates A, Coombes RC, Cuzick J, et
al: Early Breast Cancer Trialists' Collaborative Group (EBCTCG):
Aromatase inhibitors versus tamoxifen in early breast cancer:
patient-level meta-analysis of the randomised trials. Lancet.
386:1341–1352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gallagher EJ and LeRoith D: Obesity and
diabetes: the increased risk of cancer and cancer-related
mortality. Physiol Rev. 95:727–748. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sevgi M, Rigoux L, Kühn AB, Mauer J,
Schilbach L, Hess ME, Gruendler TO, Ullsperger M, Stephan KE,
Brüning JC, et al: An obesity-predisposing variant of the FTO gene
regulates D2R-dependent reward learning. J Neurosci.
35:12584–12592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Milagro FI, Moreno-Aliaga MJ and Martinez
JA: FTO obesity variant and adipocyte browning in humans. N Engl J
Med. 374:190–191. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuchenbaecker KB, Ramus SJ, Tyrer J, Lee
A, Shen HC, Beesley J, Lawrenson K, McGuffog L, Healey S, Lee JM,
et al: EMBRACE; GEMO Study Collaborators; Breast Cancer Family
Registry; HEBON; KConFab Investigators; Australian Cancer Study
(Ovarian Cancer Investigators); Australian Ovarian Cancer Study
Group; Consortium of Investigators of Modifiers of BRCA1 and BRCA2:
Identification of six new susceptibility loci for invasive
epithelial ovarian cancer. Nat Genet. 47:164–171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hasstedt SJ, Coon H, Xin Y, Adams TD and
Hunt SC: APOH interacts with FTO to predispose to healthy thinness.
Hum Genet. 135:201–207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Salgado-Montilla J, Rodríguez-Caban J,
Gonzalez-Sepulveda L, Sanchez-Ortiz R and Irizarry-Ramirez M:
Presence of FTO rs9939609 and rs9930506 and severity of prostate
cancer in Puerto Ricans. Cancer Res (106th Annual Meeting
Abstracts). pp. 48362015;
|
|
9
|
Barbier-Torres L, Delgado TC,
García-Rodríguez JL, Zubiete-Franco I, Fernández-Ramos D, Buqué X,
Cano A, Gutiérrez-de Juan V, Fernández-Domínguez I, Lopitz-Otsoa F,
et al: Stabilization of LKB1 and Akt by neddylation regulates
energy metabolism in liver cancer. Oncotarget. 6:2509–2523. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Roberts DJ and Miyamoto S: Hexokinase II
integrates energy metabolism and cellular protection: akting on
mitochondria and TORCing to autophagy. Cell Death Differ.
22:248–257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Giménez-Cassina A and Danial NN:
Regulation of mitochondrial nutrient and energy metabolism by BCL-2
family proteins. Trends Endocrinol Metab. 26:165–175. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Valenti D, Vacca RA and de Bari L:
3-Bromopyruvate induces rapid human prostate cancer cell death by
affecting cell energy metabolism, GSH pool and the glyoxalase
system. J Bioenerg Biomembr. 47:493–506. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gatenby RA and Gillies RJ: Why do cancers
have highaerobic glycolysis? Nat Rev Cancer. 4:891–899. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vincent EE, Sergushichev A, Griss T,
Gingras MC, Samborska B, Ntimbane T, Coelho PP, Blagih J, Raissi
TC, Choinière L, et al: Mitochondrial phosphoenolpyruvate
carboxykinase regulates metabolic adaptation and enables
glucose-independent tumor growth. Mol Cell. 60:195–207. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang M, Liu J, Ji H, Chen M, Zhao Y, Li
S, Zhang X and Li J: A Aconitum coreanum polysaccharide fraction
induces apoptosis of hepatocellular carcinoma (HCC) cells via
pituitary tumor transforming gene 1 (PTTG1)-mediated suppression of
the P13K/Akt and activation of p38 MAPK signaling pathway and
displays antitumor activity in vivo. Tumour Biol. 36:7085–7091.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang CZ, Wang XD, Wang HW, Cai Y and Chao
LQ: Sorafenib inhibits liver cancer growth by decreasing mTOR, AKT,
and PI3K expression. J BUON. 20:218–222. 2015.PubMed/NCBI
|
|
17
|
Zhu JJ, Cui Y, Cui K, Li X and Zhang ZY:
Distinct roles of parafibromin in the extracellular environment,
cytoplasm and nucleus of osteosarcoma cells. Am J Transl Res.
8:2426–2431. 2016.PubMed/NCBI
|
|
18
|
Mock CD, Jordan BC and Selvam C: Recent
advances of curcumin and its analogues in breast cancer prevention
and treatment. RSC Advances. 5:75575–75588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Z, Xie C, Xu W, Liu G, Cao X, Li W,
Chen J, Zhu Y, Luo S, Luo Z, et al: Phosphorylation and
inactivation of PTEN at residues Ser380/Thr382/383 induced by
Helicobacter pylori promotes gastric epithelial cell survival
through PI3K/Akt pathway. Oncotarget. 6:31916–31926.
2015.PubMed/NCBI
|
|
20
|
Pande M, Bondy ML, Do KA, Sahin AA, Ying
J, Mills GB, Thompson PA and Brewster AM: Association between
germline single nucleotide polymorphisms in the PI3K-AKT-mTOR
pathway, obesity, and breast cancer disease-free survival. Breast
Cancer Res Treat. 147:381–387. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bains M and Roberts JL: Estrogen protects
against dopamine neuron toxicity in primary mesencephalic cultures
through an indirect P13K/Akt mediated astrocyte pathway. Neurosci
Lett. 610:79–85. 2016. View Article : Google Scholar : PubMed/NCBI
|