Introduction
Tumor suppressor breast cancer susceptibility gene 2
(BRCA2) is responsible for a large percentage of familial breast
cancer cases (1,2). In addition to breast cancer, BRCA2
mutations are also linked to other types of cancer, including
ovarian, hepatocellular, pancreatic, prostate and gastric tumors
(3–5). The protein encoded by this gene is
involved in the repair of chromosomal damage and has an essential
role in the repair of DNA double-strand breaks (DSBs) through
homologous recombination (HR) (6–8). In
support of this theory, mammalian cells lacking functional BRCA2
have been shown to be sensitive to DNA damaging agents (9,10),
exhibit genomic instability (11–13)
and are deficient in homology-directed DNA repair (8,14).
BRCA2 interacts with a number of DNA repair
proteins, including γ-H2AX and RAD51 (15–17).
γ-H2AX foci formation functions to recruit DNA repair factors to
the damaged sites, enforcing the HR of DNA DSBs and linking the
process of chromatin remodeling to DNA repair (16,18,19).
RAD51 is a DNA recombinase that is essential in initiating the HR
process by mediating DNA strand exchange during recombination.
BRCA2 is required for RAD51 foci assembly in response to ionizing
radiation (IR)-induced DNA DSBs (15,17,20,21).
N-nitroso compounds (NOCs) and their precursors
exist extensively in the environment, certain occupational
settings, diets, tobacco products, cosmetics and pharmaceutical
products, and are endogenously formed in the human body from
dietary components (22,23). Many NOCs have been identified as
carcinogenic (23–27) and the International Agency for
Research on Cancer (IARC) has classified four NOCs as probably
carcinogenic to humans and another 15 as possibly carcinogenic
(23,28–30).
The carcinogenic effect of NOCs is usually attributed to their DNA
damaging and genotoxic properties (23,31,32).
Certain studies have shown that NOCs are able to
induce DNA single-strand breaks and DSBs (31,32),
suggesting that DNA repair by the HR pathway may function in
repairing the DNA damage induced by NOCs. However, there have been
no studies on the role of HR in the repair of DNA damage induced by
NOCs. We hypothesized that, as a DNA damage response, the
BRCA2-mediated HR pathway may be involved in DNA damage repair
induced by the NOCs and that this may contribute to their
carcinogenic effect. Three NOCs, N-nitrosodiethylamine (NDEA),
N-nitrosodiethanolamine (NDELA) and N-nitrosodipropylamine (NDPA),
with similar chemical structures and varying carcinogenic risks,
which were classified into differing carcinogenic classes in humans
according to the IARC, were investigated in the present study
(23,24,28–30).
The aim of the present study was to characterize the formation and
repair of the DNA damage caused by NDEA, NDELA and NDPA in gastric
cancer SGC7901 cells and investigate whether BRCA2 was involved in
the DNA damage response to these NOCs.
Materials and methods
Cells and reagents
Epidemiological studies indicate that NOCs are
positively associated with stomach cancer, therefore, the human
gastric cancer SGC7901 cell line was used in the present study. The
SGC7901 cell line was established from an untreated patient with
progressive adenocarcinoma of the stomach. The cells were cultured
in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine
serum. The SGC7901 cells stably transfected with the vector and
BRCA2 siRNA (siBRCA2) were cultured in DMEM containing 200
μg/ml of G418 (Invitrogen, Carlsbad, CA, USA). The NDEA,
NDELA, NDPA and dimethyl sulfoxide (DMSO) were purchased from Sigma
(St. Louis, MO, USA). All chemicals and solvents were of the
highest grade commercially available. The NOCs were dissolved in
sterile DMSO (0.1%) and freshly prepared each time prior to use.
Logarithmically growing SGC7901 cells were treated with NOCs at
appropriate concentrations where indicated.
Comet assay
The comet assay (Trevigen, Inc., Gaithersburg, MD,
USA) was performed as described previously, using neutral
conditions to detect the DSBs (33). In brief, the cells were harvested,
washed with ice-cold PBS and combined with molten LMP agarose, then
75 μl (500–1,000 cells) was immediately added to the comet
slide. Subsequent to being hardened, the slides were incubated for
30 min in lysis solution at 4°C, then rinsed with 1X
Tris/borate/EDTA prior to electrophoresis for 60 min at 30 V. The
slides were rinsed with distilled H2O, placed in 70%
ethanol for 10 min and then air-dried. To visualize the DNA, 50
μl of a 1:1,000 dilution of SYBR Green (Molecular Probes,
Invitrogen) in PBS was added to each slide. The slides were
visually scored using fluorescence microscopy (Leica DMS 4000B;
Leica, Mannheim, Germany). The comet tail to head ratios or tail
lengths were determined using the software package ‘Comet Assay II’
(Perceptive Instruments, Haverhill, Suffolk, UK). A minimum of 50
cells per experiment were analyzed. All the experiments were
performed at least three times independently and in triplicate.
Western blotting
The cells were harvested and lysed with a lysis
buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM
MgCl2, 100 μg/ml phenylmethylsulfonyl fluoride
and 1% Triton X-100 for 30 min on ice. Total cellular extracts (50
μg) were separated by SDS-PAGE and transferred onto
nitrocellulose membranes. The membranes were probed with specific
primary antibodies, followed by incubation with IRDye680-conjugated
secondary antibodies (Rockland, Inc., Gilbertsville, PA, USA).
Detection was performed using an Odyssey IR imaging system (LI-COR
Biotechnology, Lincoln, NE, USA). The following antibodies were
used for the immunoblotting studies: mouse anti-γ-H2AX (serine 139;
Upstate, Charlottesville, VA, USA), mouse anti-BRCA2 (Cell
Signaling Technology, Beverly, MA, USA), rabbit polyclonal
anti-RAD51 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
and anti-β-actin (Sigma).
Immunofluorescence studies
The cells were plated onto coverslips and treated
with 1X IC50 concentrations of NDEA, NDELA and NDPA
(0.76, 1.09 and 0.39 mM, respectively) for 1 h. The cells were then
fixed and stained with monoclonal anti-γ-H2AX (Upstate). Subsequent
to being stained with Alexa Fluor 488-conjugated goat anti-mouse
secondary antibodies (Invitrogen), the slides were mounted with
Vectashield mounting medium (Vector Laboratories, Burlingame, CA,
USA) containing 5 ng/ml 4′,6-diamidino-2-phenylindole (DAPI; Vector
Laboratories). The staining images were captured using fluorescence
microscopy (Leica DMS 4000B) and a Spot digital camera (Spot
Imaging Solutions, Sterling Heights, MI, USA).
Clonogenic survival assay
To determine the cytotoxicity and IC50
concentrations of NDEA, NDELA and NDPA, a clonogenic survival assay
was performed on 60-mm cell culture dishes as described previously
(34). The SGC-7901 cells were
treated with various concentrations of NDEA, NDELA or NDPA for 1 h,
followed by drug-free incubation for 10 days. The colonies were
stained with crystal violet and counted if ≥50 cells were present.
The IC50 concentration was calculated as the
concentration of NDEA, NDELA or NDPA that killed 50% of the
untreated control colonies.
Small interfering RNA (siRNA)
transfection assays
BRCA2 siRNA (SiBRCA2) oligos containing the target
sequences of 5′-AAGACACGCTGCAACAAAGCA-3′ were designed and
synthesized. Following annealing, the double-stranded siBRCA2
fragment was inserted into a pSilencer 2.1-U6-neo vector (Ambion,
Austin, TX, USA) and transfected into the SGC7901 cells with
Lipofectamine-2000 (Invitrogen). The pSilencer 2.1-U6-neo vector
containing a scrambled sequence (Ambion) was transfected as a
nonspecific control. Stable cell lines were established by
performing selection in a medium containing G418.
Statistical analyses
Images of 50 randomly selected cells were evaluated
per treatment and the test was performed three times. The Student’s
t-test was used to provide the statistical comparisons and
P<0.05 was considered to indicate a statistically significant
difference.
Results
NDEA, NDELA and NDPA-induced DNA
DSBs
First, the effects of NDEA, NDELA and NDPA on the
DNA damage in the SGC7901 cells were examined. DNA damage was
evaluated using the comet assay under neutral electrophoresis
conditions to predominantly detect the DSBs. Logarithmically
growing SGC-7901 cells were treated with 1X IC50
concentrations of NDEA (0.76 mM), NDELA (1.09 mM) or NDPA (0.39 mM)
for 15, 30 and 60 min. The comet assays revealed that NDEA, NDELA
and NDPA induced apparent DNA damage in the SGC7901 cells as
evidenced by the presence of DNA comet tails (Fig. 1A). Analysis of the tail to head
ratio, which reflected the degree of DNA damage in the cells,
showed that each of the three compounds induced a time-dependent
increase in the extent of the DNA damage (Fig. 1B).
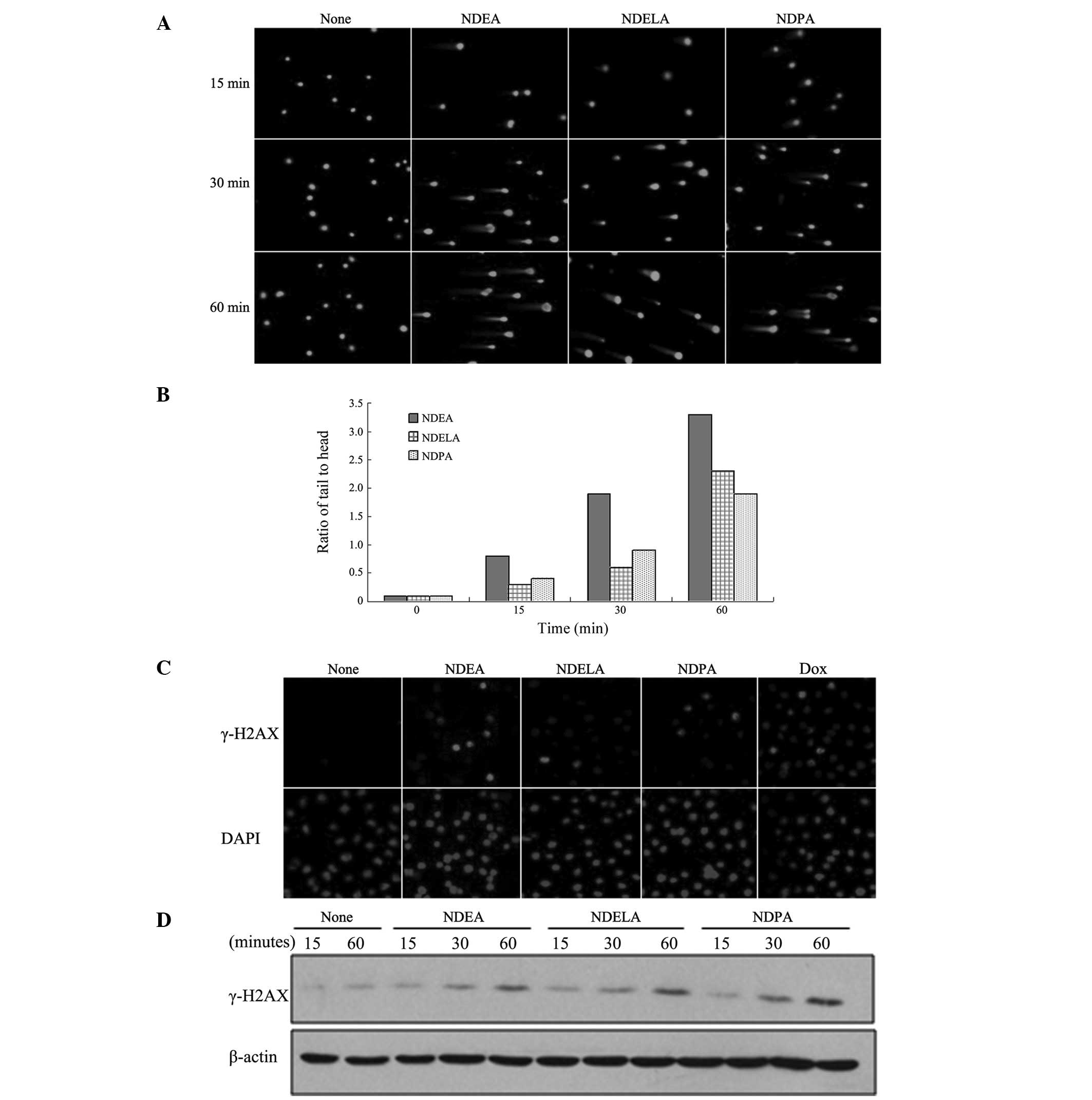 | Figure 1NDEA, NDELA and NDPA-induced DNA
double-strand breaks (DSBs). SGC-7901 cells were treated with 1X
IC50 concentrations of NDEA (0.76 mM), NDELA (1.09 mM)
or NDPA (0.39 mM) for 15, 30 and 60 min. (A) DNA damage was
determined using a neutral comet assay. Three compounds induced
apparent DNA damage in SGC7901 cells. (B) The 50 cells per slide
from (A) were analyzed to calculate the tail to head ratio of the
SGC-7901 cells. Three compounds induced a time-dependent increase
in extent of DNA damage. (C) The SGC-7901 cells were treated with
1X IC50 concentrations of NDEA (0.76 mM), NDELA (1.09
mM) or NDPA (0.39 mM) for 1 h. Following treatment, the cells were
stained with antibodies against γ-H2AX and counter-stained with
DAPI. The cells treated with Dox (1 mM) for 1 h were used as
positive controls. NOC treatment lead to formation of γ-H2AX. (D)
The SGC-7901 cells were treated with 1X IC50
concentrations of NDEA (0.76 mM), NDELA (1.09 mM) or NDPA (0.39 mM)
for 15, 30 and 60 min. Following the treatment, the cells were
collected and the cellular protein was prepared for western
blotting using antibodies against γ-H2AX. Three compounds induced
expression of γ-H2AX. NDEA, N-nitrosodiethylamine; NDELA,
N-nitrosodiethanolamine; NDPA, N-nitrosodipropylamine; DAPI,
4′,6-diamidino-2-phenylindole; Dox, Doxorubicin. |
Upon the induction of a DSB, the histone variant,
H2AX, is rapidly phosphorylated (γ-H2AX) and forms discrete nuclear
foci. γ-H2AX foci formation also allows the sensitive detection of
DSBs (16,18,35,36).
To further confirm that NOCs induce the generation of DSBs, the
SGC-7901 cells were treated with 1X IC50 concentrations
of NDEA (0.76 mM), NDELA (1.09 mM) or NDPA (0.39 mM) for 1 h and
then immunofluorescently stained using γ-H2AX antibodies. An
examination of the results showed that the NOC treatment led to the
formation of γ-H2AX (Fig. 1C), thus
indicating the presence of DSBs. Immunoblotting analysis further
demonstrated that NDEA, NDELA and NDPA induced the expression of
γ-H2AX in a time-dependent manner (Fig.
1D).
Differential efficiency of DNA damage
repair in response to NDEA, NDELA and NDPA treatment
The comet assay and the data on the expression of
γ-H2AX demonstrated that the three compounds, NDEA, NDELA and NDPA,
were able to induce a time-dependent increase in the extent of DNA
damage. These observations indicated that these three NOCs
similarly induced the generation of DSBs. As with the varying
carcinogenic potentials, it was unclear whether these NOCs induced
similar DNA repair. We hypothesized that the repair mechanism of
these NOC-induced DSBs may be different. To examine this
hypothesis, the SGC7901 cells were treated with 1X IC50
concentrations of NDEA (0.76 mM), NDELA (1.09 mM) or NDPA (0.39 mM)
for 1 h, followed by a 12-h drug-free incubation to allow DNA
repair. The formation and resolution of the DNA damage were then
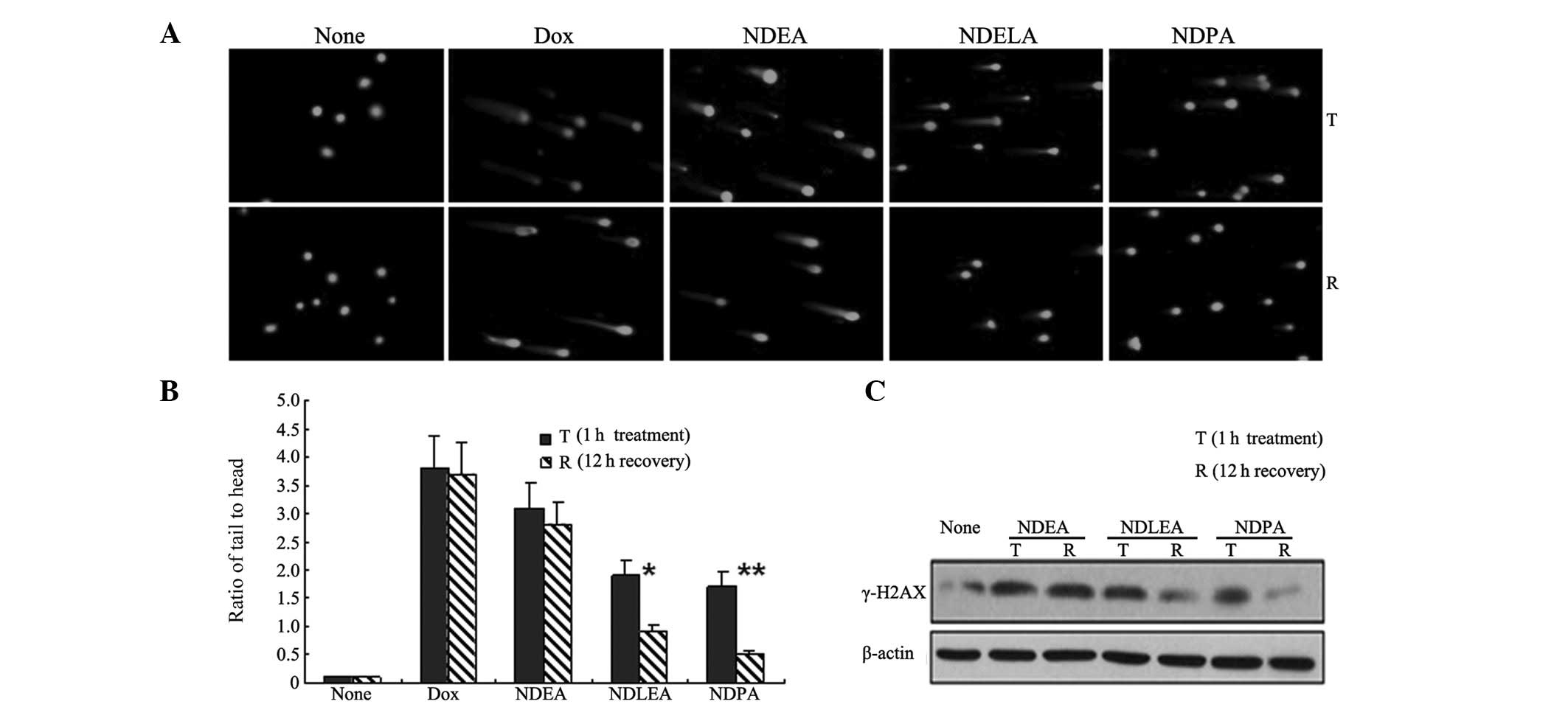
analyzed using the comet assay. Following a drug-free incubation of
12 h, the tail to head ratio of the comet DNA in the cells treated
with NDELA (P<0.05) or NDPA (P<0.01) was observed to be
significantly reduced (Fig. 2A and
B). However, there was no apparent reduction in the tail to
head ratio of the comet DNA in the cells treated with NDEA
(P>0.05). The levels of γ-H2AX expression were also studied and
it was observed that following a drug-free incubation of 12 h,
there was a significant reduction in the expression of γ-H2AX in
the SGC7901 cells treated with NDELA or NDPA (Fig. 2C). However, there was no clear
change in the γ-H2AX levels in the cells treated with NDEA. These
observations suggested that the DSBs induced by NDELA or NDPA were
more effectively repaired than those induced by NDEA.
BRCA2-mediated HR contributes to the
differential repair of NOC-induced DSBs in SGC7901 cells
In order to determine whether HR was involved in the
repair of NOC-induced DSBs, the expression levels of BRCA2 and
RAD51, two key proteins in HR (4,8,20),
were observed in response to the NOC treatment. The SGC7901 cells
were treated with 1X IC50 concentrations of NDEA, NDELA
and NDPA for 15 and 60 min. The expression of BRCA2 and RAD51 was
reduced in the SGC7901 cells treated with NDEA (Fig. 3A). However, the expression levels of
BRCA2 and RAD51 were notably upregulated in a time-dependent manner
in the cells treated with NDELA or NDPA (Fig. 3A). The results suggest that the
BRCA2-RAD51-mediated HR pathway may be significant in the DNA
damage repair induced by NDELA and NDPA.
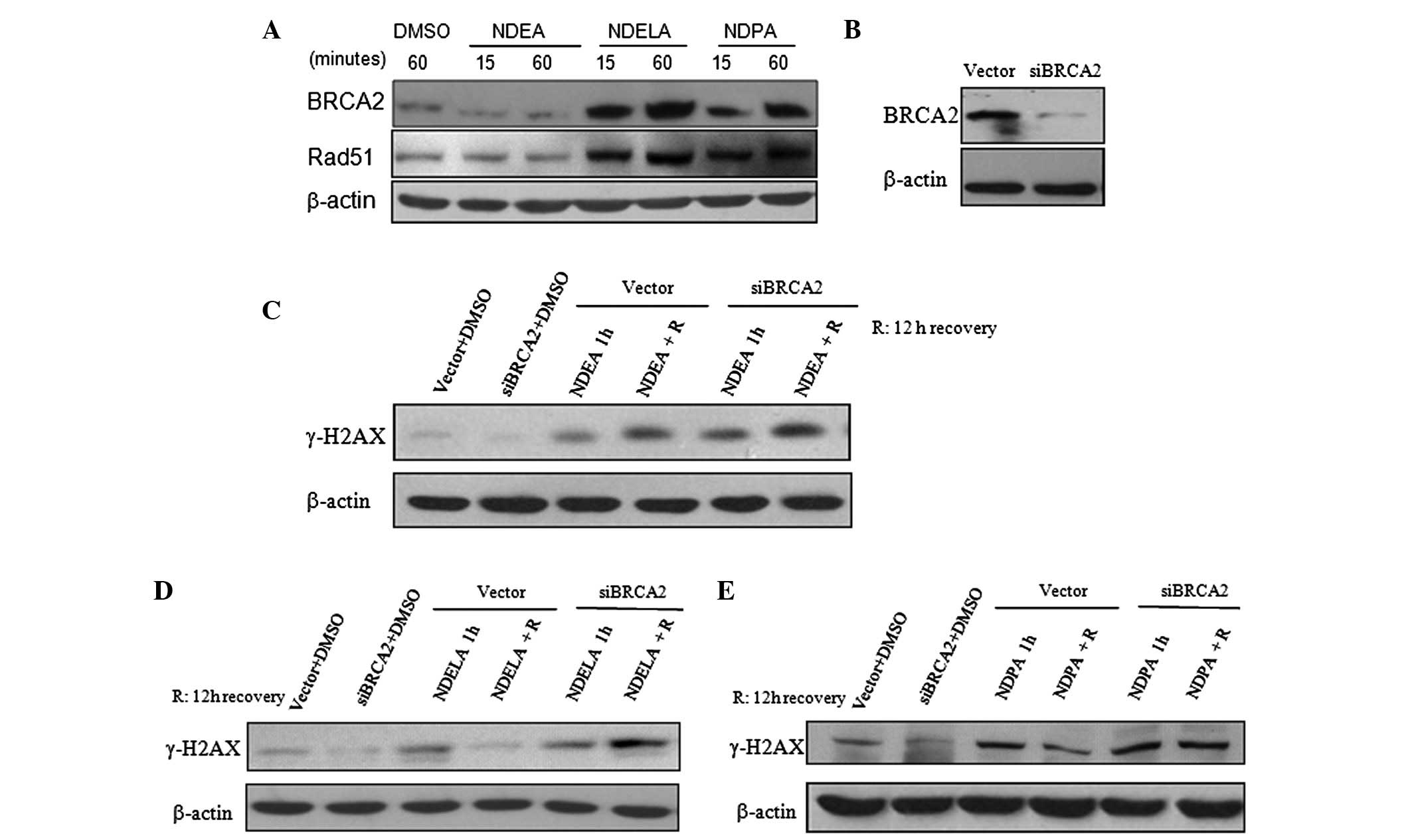 | Figure 3Involvement of the BRCA2-mediated
pathway in the DNA repair of NOC-induced damage. (A) The SGC7901
cells were treated with 1X IC50 concentrations of NDEA
(0.76 mM), NDELA (1.09 mM) or NDPA (0.39 mM) for 15, and 60 min.
Following an incubation in drug-free medium for 12 h, the cells
were harvested and the expression of BRCA2 and RAD51 was determined
by western blotting. The expression of BRCA2 and RAD51 was reduced
in SGC7901 cells treated with NDEA, but were notably upregulated in
cells treated with NDELA or NDPA. (B) Expression of BRCA2 was
knocked down in siBRCA2-transfected SGC7901 cells. BRCA2-targeted
siRNA and a control vector plasmid were stably transfected into the
SGC7901 cells. (C–E) Differential expression levels of γ-H2AX
induced by NOCs in vector- and siBRCA2-transfected cells following
a 12-h drug-free incubation. The vector- and siBRCA2-transfected
SGC7901 cells were treated with 1X IC50 concentrations
of (C) NDEA, (D) NDELA and (E) NDPA for 1 h, followed by a 12-h
drug-free incubation. The expression levels of of γ-H2AX were
determined. The DNA damage induced by NDELA or NDPA was not
repaired as effectively as in the vector cells. DMSO (1 mM)
treatment was used as a negative control. BRCA2, breast cancer
susceptibity gene 2; NOC, N-nitroso compound; NDEA,
N-nitrosodiethylamine; NDELA, N-nitrosodiethanolamine; NDPA,
N-nitrosodipropylamine; siRNA, small interfering RNA. |
The role of BRCA2 in the repair of NOC-induced DNA
damage was further investigated. A BRCA2-targeted siRNA was
designed (as described in the Materials and methods) and stably
transfected into the SGC7901 cells. The effectiveness of the
siBRCA2 construct in knocking down the endogenous BRCA2 level was
demonstrated by western blotting. As shown in Fig. 3B, the expression of BRCA2 was
effectively knocked down using BRCA2 siRNA transfected into the
SGC7901 cells. The control group consisted of SGC7901 cells stably
transfected with the control vector plasmid. The vector- and
siBRCA2-transfected SGC7901 cells were treated with 1X
IC50 concentrations of NDEA, NDELA or NDPA for 1 h,
followed by a 12-h drug-free incubation. Similar to the cells
treated with NDEA (Fig. 3C), the
level of γ-H2AX expression in the BRCA2-knockdown cells treated
with NDELA (Fig. 3D) or NDPA
(Fig. 3E) was not reduced to the
level of the vector cells following the drug-free incubation. The
results showed that in the BRCA2-knockdown cells, the DNA damage
induced by NDELA or NDPA was not repaired as effectively as in the
vector cells. This also suggested that BRCA2 contributes to the
variations in DNA repair in response to these NOC-induced DSBs.
BRCA2 confers sensitivity to NOC
treatment
Based on the previous observations that BRCA2 is
significant for the repair of DNA damage induced by NOCs, we
hypothesized that BRCA2 may affect the sensitivity to NOCs. To
determine whether BRCA2 affects the sensitivity to NDEA, NDELA and
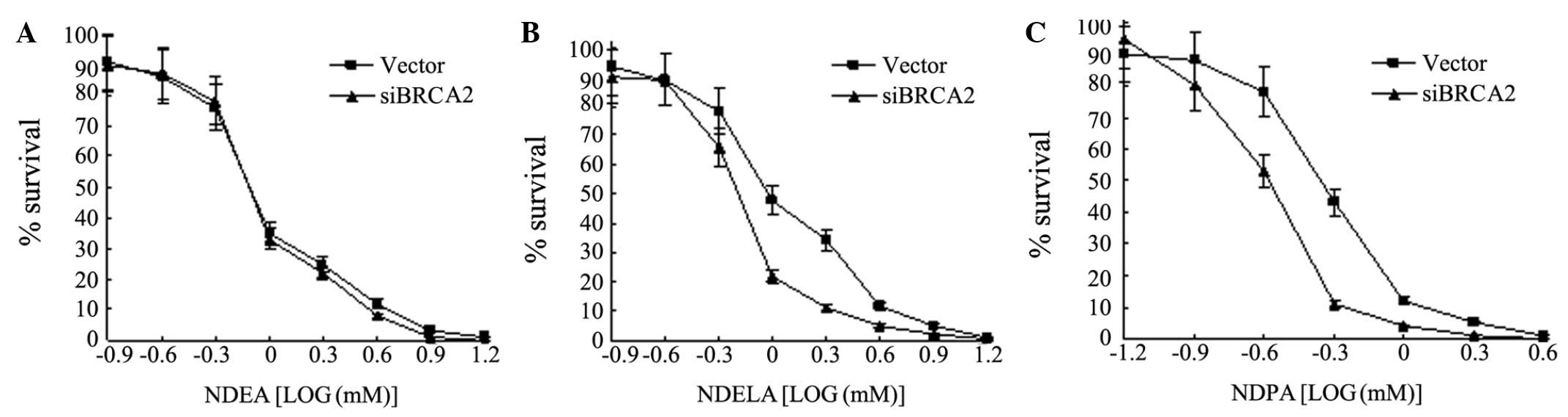
NDPA, a clonogenic survival assay was performed. The vector- and
siBRCA2-transfected SGC7901 cells were treated with various
concentrations of NDEA, NDELA or NDPA for 1 h, followed by
drug-free incubations. When the IC50 concentrations were
compared, the results showed that the cells with the endogenous
knock down of BRCA2, caused by RNA interference, exhibited a
>2-fold increase in sensitivity to NDELA (Fig. 4B) or NDPA (Fig. 4C) compared with the vector cells. No
significant change was observed in the sensitivity to NDEA
(Fig. 4A). These results show that
BRCA2 confers sensitivity to NOCs.
Discussion
NOCs are widely distributed in the environment and
are recognized as genotoxic agents and possible chemical
carcinogens. Their carcinogenicity mainly depends on their
genotoxicity (23–27). NDEA, NDELA and NDPA belong to
carcinogen groups 2A, 2B and 3, respectively (28–30).
In the present study, it was observed that these three NOCs were
able to similarly induce time-dependent DSBs in the SGC7901 cells
and that the induced DSB repair varied. The DNA damage induced by
NDEA, the most potent carcinogen, was observed to not be repaired
as efficiently as that caused by NDELA or NDPA. The results
suggested that the pathways of NOC-induced DNA damage repair differ
and that this may contribute significantly to an NOC’s
genotoxicity.
DNA DSBs are the most lethal DNA lesions, posing an
almost insurmountable challenge to the cells’ DNA repair machinery.
HR is a DSB repair pathway. The HR pathway uses one sister DNA
strand as the repair template and repairs DNA DSBs with high
fidelity (11,14,37).
In order to understand why the repair of NOC-induced DSBs varied
and to determine the role of HR in this process, the expression
levels of of BRCA2 and RAD51, two key proteins in HR, were
evaluated. The SGC7901 cells exhibited a defective HR in response
to NDEA treatment and an increased HR due to NDELA or NDPA
treatment. The results show that HR has a significant role in the
process of NOC-induced DSB repair, which may explain why the DNA
damage induced by NDEA is not repaired as efficiently as that
caused by NDELA or NDPA and why NDEA possesses the most potent
genotoxic-carcinogenicity among the NOCs.
Nonhomologous end joining (NHEJ) is an alternative
mechanism for the repair of DSBs (38). This process joins broken chromosome
ends in a manner that does not depend on sequence homology and so
may not be error free. Although the NHEJ pathway frequently results
in minor changes in DNA sequence at the break site, and
occasionally the joining of previously unlinked DNA molecules, it
is a major contributor to cell survival following the exposure of
mammalian cells to agents that cause DSBs. Since the HR repair of
the DNA damage induced by NDELA or NDPA did not vary significantly,
it is possible that a differential NHEJ response was induced.
However, this hypothesis requires further study.
BRCA2 is a key protein involved in HR. Mammalian
cells lacking functional BRCA2 are sensitive to DNA damaging agents
and are deficient in homology-directed DNA repair (6–8).
Genotoxic agents such as mitomycin C have been associated with
decreased BRCA2 protein expression (39). BRCA2 is also degraded during the
alkyltransferase-mediated DNA repair of DNA adducts (40). The present study showed that the
expression of BRCA2 was inhibited by NDEA treatment, but
upregulated with NDELA or NDPA. The knock down of BRCA2 impaired
the DNA damage repair induced by NDELA or NDPA. The cells with this
knock down showed an increased sensitivity to NDELA or NDPA,
suggesting that BRCA2 may have a particularly significant role in
differential DSB repair in response to NOC-induced DSBs.
NDEA, NDELA and NDPA are three NOCs with similar
chemical structures, but different carcinogenic risks. The present
study demonstrated that these NOCs had similar effects on DNA
damage. NDELA- or NDPA-induced DNA damage was observed to be more
effectively repaired than that induced by NDEA. NDELA and NDPA
upregulated the expression of BRCA2 and RAD51, but NDEA did not.
Furthermore, it was observed that the knock down of BRCA2 blocked
NDELA- or NDPA-induced DNA damage repair and also that cells with
this knock down showed an increased sensitivity to NDELA or NDPA.
Taken together, these observations suggest that the BRCA2-mediated
DNA repair pathway may have a significant role in NOC-induced DNA
damage repair and that this may be associated with the differential
carcinogenicity of these NOCs.
Acknowledgements
The present study was supported by
grants from the Natural Science Foundation of China (Grant No.
81170385), the Shanghai Pujiang Talent Plan (07pj14003), the
Chinese Ministry of Science and Technology (2007BAC27B02) and the
Chinese Academy of Sciences (KSCX2-YW-R-114).
References
|
1
|
Wooster R, Bignell G, Lancaster J, et al:
Identification of the breast cancer susceptibility gene BRCA2.
Nature. 378:789–792. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tavtigian SV, Simard J, Rommens J, et al:
The complete BRCA2 gene and mutations in chromosome 13q-linked
kindreds. Nat Genet. 12:333–337. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jakubowska A, Nej K, Huzarski T, Scott RJ
and Lubiński J: BRCA2 gene mutations in families with aggregations
of breast and stomach cancers. Br J Cancer. 87:888–891. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Venkitaraman AR: Cancer susceptibility and
the functions of BRCA1 and BRCA2. Cell. 108:171–182. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liede A, Karlan BY and Narod SA: Cancer
risks for male carriers of germline mutations in BRCA1 or BRCA2: a
review of the literature. J Clin Oncol. 22:735–742. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patel KJ, Yu VP, Lee H, et al: Involvement
of Brca2 in DNA repair. Mol Cell. 1:347–357. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moynahan ME, Pierce AJ and Jasin M: BRCA2
is required for homology-directed repair of chromosomal breaks. Mol
Cell. 7:263–272. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
O’Donovan PJ and Livingston DM: BRCA1 and
BRCA2: breast/ovarian cancer susceptibility gene products and
participants in DNA double-strand break repair. Carcinogenesis.
31:961–967. 2010.PubMed/NCBI
|
|
9
|
Bryant HE, Schultz N, Thomas HD, et al:
Specific killing of BRCA2-deficient tumours with inhibitors of
poly(ADP-ribose) polymerase. Nature. 434:913–917. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang W and Figg WD: Secondary BRCA1 and
BRCA2 alterations and acquired chemoresistance. Cancer Biol Ther.
7:1004–1005. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thompson LH and Schild D: Homologous
recombinational repair of DNA ensures mammalian chromosome
stability. Mutat Res. 477:131–153. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pellegrini L and Venkitaraman A: Emerging
functions of BRCA2 in DNA recombination. Trends Biochem Sci.
29:310–316. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shivji MK and Venkitaraman AR: DNA
recombination, chromosomal stability and carcinogenesis: insights
into the role of BRCA2. DNA Repair (Amst). 3:835–843. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ohnishi T, Mori E and Takahashi A: DNA
double-strand breaks: their production, recognition, and repair in
eukaryotes. Mutat Res. 669:8–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davies AA, Masson JY, McIlwraith MJ, et
al: Role of BRCA2 in control of the RAD51 recombination and DNA
repair protein. Mol Cell. 7:273–282. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Foster ER and Downs JA: Histone H2A
phosphorylation in DNA double-strand break repair. FEBS J.
272:3231–3240. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Esashi F, Galkin VE, Yu X, et al:
Stabilization of RAD51 nucleoprotein filaments by the C-terminal
region of BRCA2. Nat Struct Mol Biol. 14:468–474. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rogakou EP, Pilch DR, Orr AH, Ivanova VS
and Bonner WM: DNA double-stranded breaks induce histone H2AX
phosphorylation on serine 139. J Biol Chem. 273:5858–5868. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van Attikum H, Fritsch O, Hohn B and
Gasser SM: Recruitment of the INO80 complex by H2A phosphorylation
links ATP-dependent chromatin remodeling with DNA double-strand
break repair. Cell. 119:777–788. 2004.PubMed/NCBI
|
|
20
|
Pellegrini L, Yu DS, Lo T, et al: Insights
into DNA recombination from the structure of a RAD51-BRCA2 complex.
Nature. 420:287–293. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Powell SN and Kachnic LA: Roles of BRCA1
and BRCA2 in homologous recombination, DNA replication fidelity and
the cellular response to ionizing radiation. Oncogene.
22:5784–5791. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Filho PJ, Rios A, Valcárcel M and Caramao
EB: Development of a new method for the determination of
nitrosamines by micellar electrokinetic capillary chromatography.
Water Res. 37:3837–3842. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brambilla G and Martelli A: Genotoxic and
carcinogenic risk to humans of drug-nitrite interaction products.
Mutat Res. 635:17–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hecht SS: Approaches to cancer prevention
based on an understanding of N-nitrosamine carcinogenesis. Proc Soc
Exp Biol Med. 216:181–191. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Williams GM, Iatropoulos MJ, Jeffrey AM,
Luo FQ, Wang CX and Pittman B: Diethylnitrosamine
exposure-responses for DNA ethylation, hepatocellular
proliferation, and initiation of carcino-genesis in rat liver
display non-linearities and thresholds. Arch Toxicol. 73:394–402.
1999. View Article : Google Scholar
|
|
26
|
Pinto LF, Moraes E, Albano R, et al: Rat
oesophageal cytochrome P450 (CYP) monooxygenase system: comparison
to the liver and relevance in N-nitrosodiethylamine carcinogenesis.
Carcinogenesis. 22:1877–1883. 2001. View Article : Google Scholar
|
|
27
|
Visoni S, Lang M and Ribeiro Pinto LF:
Hamster exhibits major differences in organ-specific metabolism of
the esophageal carcinogen N-nitrosodiethylamine. Toxicol Lett.
183:90–94. 2008.PubMed/NCBI
|
|
28
|
International Agency for Research on
Cancer (IARC): Overall evaluations of carcinogenicity: an updating
of IARC Monographs volumes 1 to 42. IARC Monogr Eval Carcinog Risks
Hum. Suppl 7:1–440. 1987.PubMed/NCBI
|
|
29
|
International Agency for Research on
Cancer (IARC): Some N-Nitroso Compounds. IARC Monogr Eval Carcinog
Risks Hum. 17:1–365. 1978.
|
|
30
|
International Agency for Research on
Cancer (IARC): Tobacco habits other than smoking; betel-quid and
areca-nut chewing; and some related nitrosamines. IARC Working
Group Lyon, 23–30 October 1984. IARC Monogr Eval Carcinog Risk Chem
Hum. 37:1–268. 1985.PubMed/NCBI
|
|
31
|
Bradley MO, Dysart G, Fitzsimmons K,
Harbach P, Lewin J and Wolf G: Measurements by filter elution of
DNA single- and double-strand breaks in rat hepatocytes: effects of
nitrosamines and gamma-irradiation. Cancer Res. 42:2592–2597.
1982.PubMed/NCBI
|
|
32
|
Loeppky RN, Ye Q, Goelzer P and Chen Y:
DNA adducts from N-nitrosodiethanolamine and related beta-oxidized
nitrosamines in vivo: (32)P-postlabeling methods for glyoxal- and
O(6)-hydroxyethyldeoxyguanosine adducts. Chem Res Toxicol.
15:470–482. 2002. View Article : Google Scholar
|
|
33
|
Wiltshire T, Senft J, Wang Y, et al: BRCA1
contributes to cell cycle arrest and chemoresistance in response to
the anticancer agent irofulven. Mol Pharmacol. 71:1051–1060. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Wiltshire T, Senft J, et al:
Fanconi anemia D2 protein confers chemoresistance in response to
the anticancer agent, irofulven. Mol Cancer Ther. 5:3153–3161.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fillingham J, Keogh MC and Krogan NJ:
GammaH2AX and its role in DNA double-strand break repair. Biochem
Cell Biol. 84:568–577. 2006.PubMed/NCBI
|
|
36
|
Dickey JS, Redon CE, Nakamura AJ, et al:
H2AX: functional roles and potential applications. Chromosoma.
118:683–692. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Phillips ER and McKinnon PJ: DNA
double-strand break repair and development. Oncogene. 26:7799–7808.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hefferin ML and Tomkinson AE: Mechanism of
DNA double-strand break repair by non-homologous end joining. DNA
Repair (Amst). 4:639–648. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schoenfeld AR, Apgar S, Dolios G, Wang R
and Aaronson SA: BRCA2 is ubiquitinated in vivo and interacts with
USP11, a deubiquitinating enzyme that exhibits prosurvival function
in the cellular response to DNA damage. Mol Cell Biol.
24:7444–7455. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Philip S, Swaminathan S, Kuznetsov SG, et
al: Degradation of BRCA2 in alkyltransferase-mediated DNA repair
and its clinical implications. Cancer Res. 68:9973–9981. 2008.
View Article : Google Scholar : PubMed/NCBI
|