Introduction
Tangeretin is a natural polymethoxyflavonoid
extracted from citrus fruits (1),
which exhibits antiproliferative, anti-invasive, antimetastatic and
antioxidant activities (2–4). Recently, many studies have shown that
polymethoxylated flavonoids such as tangeretin and nobiletin are
more potent inhibitors of tumor cell growth than free hydroxylated
flavonoids (3,5,6).
Tangeretin inhibits the growth of many cell lines such as breast
cancer cell lines MDA-MB-435 and MCF-7, human colon cancer cell
line HT-29 and leukaemic HL-60 cells through cell cycle arrest or
apoptosis (2,7,8).
Tangeretin activates the unfolded protein response and synergizes
with imatinib in the erythroleukemia cell line K562 and suppresses
IL-1β-induced cyclooxygenase (COX)-2 expression through inhibition
of p38 MAPK, JNK and AKT activation in human lung carcinoma cells
(9,10). In addition, tangeretin was reported
to sensitize cisplatin-resistant human ovarian cancer cells through
downregulation of the phosphoinositide 3-kinase/Akt signaling
pathway (11). Thus, tangeretin may
be a potential plant-derived agent for use in the treatment of
gastric cancer.
Gastric cancer is the second leading cause of
cancer-related death in the world (12). AGS is a type of human gastric
adenocarcinoma cell line with wild-type p53, which has been used in
numerous studies on antineoplastic drugs (13). Wild-type p53 is a tumor-suppressor
gene, which plays an important role in cancer cell growth
inhibition and mediates either cell cycle arrest or apoptosis
(14,15). In response to various forms of
stress, p53 is activated and accumulates in the nucleus, where it
regulates the transcription of numerous target genes using specific
DNA response elements. Beyond its nuclear role, p53 regulates a
number of mitochondrial functions. However, the mutant p53 existing
in some tumor cells loses its activity and has no antitumor
function (16). Thus, as a gastric
cancer cell line with wild-type p53, AGS is suitable for
investigation of the role of p53 in the anticancer action of
tangeretin.
Our previous study showed that tangeretin inhibits
the proliferation of human AGS gastric cancer cells and induces
cell cycle arrest in the S and G2/M phases through inhibition of
ERK phosphorylation and upregulation of cyclin B1 protein (17). In the present study, we further
examined the effects of tangeretin on the apoptosis of AGS cells
and explored the related mechanisms.
Materials and methods
Reagents and antibodies
Tangeretin was obtained from Shanghai Tongtian
Biotechnology Co., Ltd. (Shanghai, China).
Methylthiazolyldiphenyl-tetrazolium bromide (MTT) was obtained from
Shanghai Richu Biotechnology Ltd. Propidium iodide (PI) was from
Sigma-Aldrich, Inc. (St. Louis, MO, USA). ECL Plus Western Blotting
detection reagents were from GE Healthcare (Wauwatosa, WI, USA).
Annexin V-FITC apoptosis kit and caspase-3, -8, -9 colorimetric
assay kits were obtained from BioVision (Mountain View, CA, USA).
All antibodies and secondary antibodies were from Cell Signaling
Technology (Beverly, MA, USA).
Cell culture
Human gastric cancer cell line AGS from the American
Type Culture Collection (ATCC, Rockville, MD, USA) was cultured in
HAM’s/F12 medium (HyClone, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS) at 37°C in a 5% CO2 air
atmosphere.
MTT assay for cell viability
Cells were seeded in 96-well plates and treated with
different concentrations of tangeretin for 24, 48 and 72 h,
respectively. Then, MTT solution was added to each well at a final
concentration of 1 mg/ml, and the plates were incubated at 37°C for
another 4 h. After incubation, 150 μl dimethyl sulfoxide (DMSO) was
added to each well and the absorbance was read at 570 nm using an
absorbance microplate reader (SpectraMax 190; Molecular Devices,
Sunnyvale, CA, USA). The experiments were performed in triplicate
and at least repeated three times. Inhibition rates = (1 − sample
OD/control OD) × 100%.
Analysis of apoptosis
Apoptosis was evaluated using an Annexin V-FITC and
PI double staining kit. After treatment with 10, 30 and 60 μM
tangeretin or 0.05% DMSO for 48 h, AGS cells were harvested and
incubated with 500 μl Annexin V binding buffer containing 5 μg/ml
PI and 1 μl FITC-labeled Annexin V for 30 min on ice. Then cells
were analyzed by a FACSCalibur flow cytometer (BD Biosciences, San
Jose, CA, USA), and the results were indicated as percentages of
apoptotic cells. Images of cells were captured under an inverted
fluorescence microscope (IX71, Olympus, Tokyo, Japan).
Hoechst 33258 staining
After treatment with tangeretin for 48 h, cells were
fixed in 4% paraformaldehyde for 2 h and stained with 20 μM Hoechst
33258 for 20 min after washing with PBS. Cells were then observed
and images were captured under an inverted fluorescence
microscope.
Caspase-3, -8 and -9 activity assay
Caspase activity was examined by a caspase-3, -8, -9
colorimetric assay kit respectively according to the manufacturer’s
protocol. Sample readings at 405 nm were conducted using a
SpectraMax 190 microplate reader. Fold-increase in activity was
determined by comparing the results with the level of the
control.
Mitochondrial transmembrane potential
assay
The cells treated with tangeretin were collected by
centrifugation at 3,000 rpm for 10 min and suspended in 500 μl PBS.
The cells were incubated with Rhodamine 123 (1 μg/ml) for 30 min
and examined by a FACSCalibur flow cytometer.
Western blot analysis
After a 48-h exposure to tangeretin at different
doses, cells were washed twice with ice-cold PBS and lysed in
protein lysis buffer. Protein concentrations were tested by the
Bradford method. Proteins were subjected to electrophoresis on 12%
polyacrylamide gels and then transferred to polyvinylidene
difluoride (PVDF) membranes by electrotransfer (Bio-Rad
Laboratories, CA, USA). After blocking with 5% non-fat milk in PBS
buffer containing 0.5% Tween-20 and washed three times, the
membranes were incubated with various primary antibodies,
respectively. Membranes were washed and incubated with the
corresponding secondary antibodies, and then the protein bands were
visualized with ECL Plus Western blotting detection reagents.
Statistical analysis
Data are presented as means ± SEM. Statistical
analysis of data was carried out with GraphPad Prism 5.0 (Hearne
Scientific Software, Chicago, IL, USA). Multiple comparisons were
carried out by one-way analysis of variance (ANOVA). A value of
P<0.05 was considered to indicate a statistically significant
result.
Results
Effects of tangeretin on the viability of
AGS cells
The anti-proliferative effect of tangeretin on the
human gastric cancer AGS cell line was examined by MTT assay. The
inhibitory rates of the cells exposed to 5, 10, 30, 60, 120 and 240
μM tangeretin for 24, 48 and 72 h were observed, respectively. The
results showed that tangeretin decreased the viability rates of AGS
cells in a dose- and time-dependent manner (Fig. 1).
Tangeretin induces AGS cell
apoptosis
Flow cytometric assay showed that the early
apoptosis rates and late apoptosis (necrosis) rates of AGS cells
treated with tangeretin for 48 h increased in a dose-dependent
manner (Fig. 2A; P<0.05,
P<0.01). Fig. 2B also shows that
the percentage of Annexin V-FITC (green)-labeled early apoptotic
cells and PI (red)-labeled late apoptotic or necrotic cells
increased along with the increase in the concentration of
tangeretin. In addition, typical apoptotic morphological changes
including cell nuclear shrinkage, chromatin condensation and
apoptotic bodies were observed after exposure to different doses of
tangeretin by Hoechst 33258 staining (Fig. 2C). These results confirmed that
tangeretin induced the apoptosis of AGS cells.
Involvement of caspases in
tangeretin-induced apoptosis
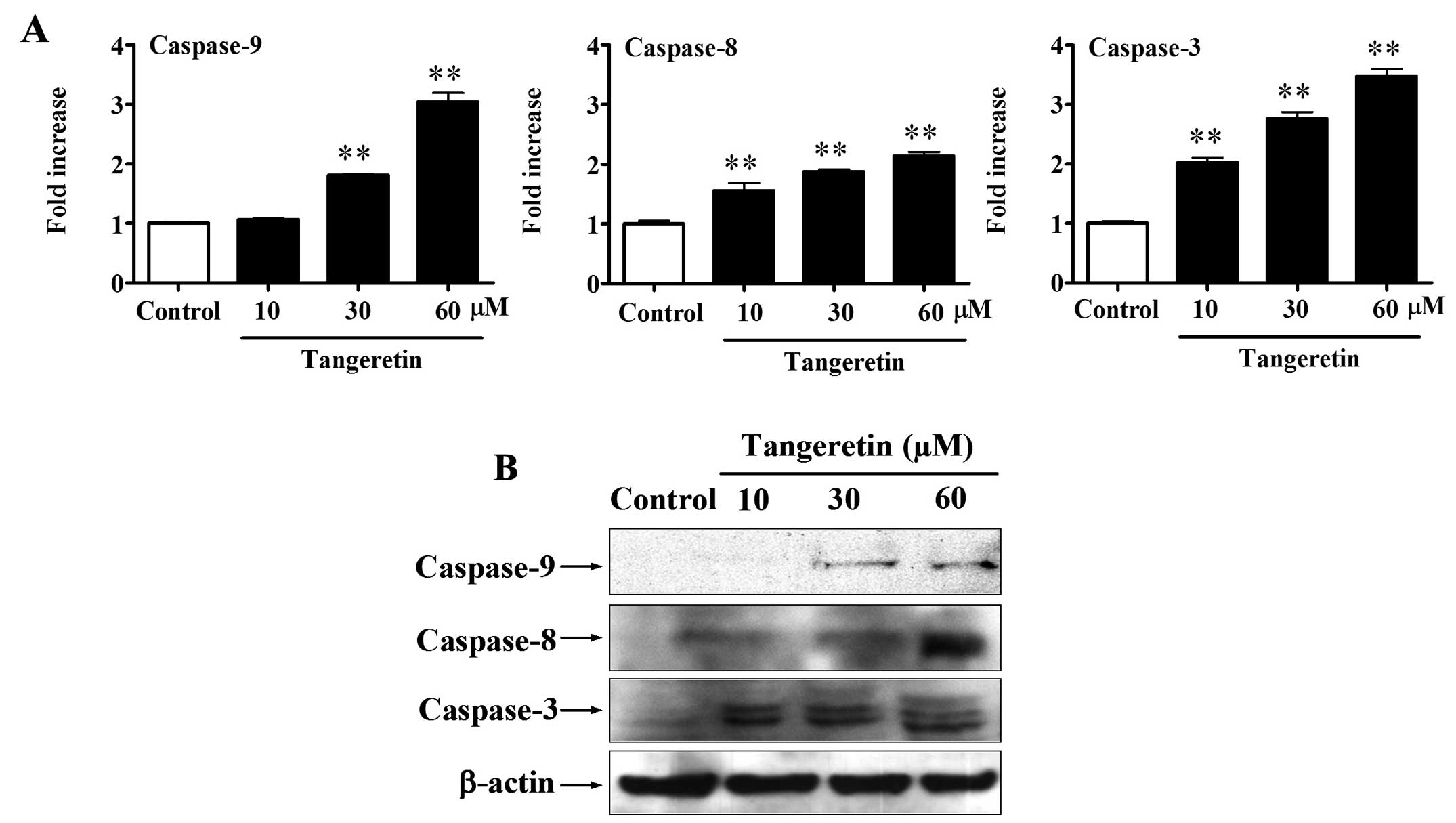
As shown in Fig. 3A,
the results showed that the activities of caspase-3, -8 and -9 were
increased by tangeretin in a dose-dependent manner after a 48-h
exposure (P<0.01). Treatment of cells with 60 μM tangeretin
increased the caspse-3 activity ~3.5-fold, caspase-8 ~1.8-fold and
caspase-9 ~3.0-fold. Western blot assay showed that the expression
levels of cleaved caspase-3, -8 and -9 proteins were upregulated by
tangeretin in a dose-dependent manner (Fig. 3B). These results indicate that
tangeretin induces both the extrinsic and intrinsic apoptotic
pathways with the activation of the caspase cascade.
Fas/FasL is involved in
tangeretin-induced apoptosis
To examine the extrinsic signaling pathway through
Fas, the expression levels of death receptor Fas and its ligand
FasL were assessed. As shown in Fig.
4, the expression of Fas and FasL was increased in a
dose-dependent manner (P<0.05, P<0.01). Thus, the binding of
FasL to Fas induced Fas trimerization, which recruited and
activated initiator caspase-8. Activated caspase-8 stimulates
apoptosis via two parallel cascades; it directly cleaves and
activates caspase-3, or it cleaves the pro-apoptotic Bcl-2 family
protein Bid. Cleaved/truncated Bid (tBid) translocates to the
mitochondria, inducing cytochrome c release, which
sequentially activates caspase-9 and caspase-3.
Mitochondrial signaling pathway in
tangeretin-induced apoptosis
Decrease and breakage of mitochondrial membrane
potential (MMP) is an important sign of mitochondrial damage. Flow
cytometric assay after Rhodamine 123 staining showed that the
percentages of cells with high MMP decreased in a dose-dependent
manner (P<0.01), suggesting that tangeretin reduced the levels
of MMP and induced mitochondrial damage (Fig. 5). In addition, the protein levels of
cytochrome c, Bax, Bid and tBid increased as the dose of
tangeretin increased (Fig. 6;
P<0.05, P<0.01). These results indicate that Bax and tBid
activated by upstream proteins mediated mitochondrial dysfunction,
induced the release of cytochrome c and activation of
caspase-9 and caspase-3. In addition, the extrinsic signaling
pathway interacted with the mitochondrial signaling pathway through
induction of cleavage of Bid by caspase-8.
Tangeretin-induced apoptosis is dependent
on p53 activation
As shown in Fig. 7A,
significant elevations in the p53 and p21 protein levels were noted
in a dose-dependent manner (P<0.01). To determine whether
tangeretin-induced apoptosis is p53-dependent, a p53 inhibitor
Pifithrin-α (PFT-α) was used. The results showed that 20 μM PFT-α
reduced the apoptotic rates induced by tangeretin from 15.8±0.1 to
7.6±1.1% (Fig. 7B; P<0.01,
P<0.001). In addition, western blot analysis showed that 20 μM
PFT-α reduced the expression of p53, p21, caspase-3 and caspase-9
proteins significantly increased by 60 μM tangeretin (Fig. 7C). These results suggest that
tangeretin-induced apoptosis of AGS cells was p53-dependent.
Discussion
The present study showed that tangeretin inhibited
the viability and induced the apoptosis of human gastric cancer AGS
cells through both extrinsic and intrinsic signaling pathways
through the activation of the caspase cascade. The present study
provides important data to support the finding that tangeretin as a
natural component widely existing in many fruits or fruit juices
exhibits antineoplastic activity in human gastric cancer.
Caspases play a crucial role in apoptosis, and the
activation of caspases amplifies apoptotic signaling (18). In the present study, tangeretin
increased the activities of caspase-3, -8 and -9 and upregulated
the expression of cleaved caspase-3, -8 and -9 proteins. The active
forms of caspase-8 and -9 can activate downstream effectors
caspase-3, -6 and -7, resulting in the cleavage of several
intracellular polypeptides (e.g., PARP-1) as well as activation of
DNase, which leads to DNA fragmentation (19).
Apoptosis is mainly mediated by the death
receptor-triggered extrinsic pathway, the mitochondrial-initiated
intrinsic pathway and the endoplasmic reticulum pathway (20–22).
The dissipation of MMP marks the point of no return during the
apoptotic program and occurs prior to DNA fragmentation. Thus, the
evaluation of MMP depolarization is of critical importance for the
assessment of apoptosis (23,24).
In the present study, MMP was observed to be reduced by tangeretin
in a dose-dependent manner, which suggests that mitochondrial
dysfunction contributed to the tangeretin-induced apoptosis. In
addition, western blot analysis showed that the expression levels
of Fas, FasL, Bid and tBid were increased by tangeretin. It was
previously reported that Bid is activated through cleavage by
caspase-8 to become cleaved Bid (tBid) in the Fas-induced process
of apoptosis, and tBid is necessary for Bax oligomerization
(25–27). Therefore, in the present study,
tangeretin may have activated caspase-8 through Fas/FasL, and then
the activated caspase-8 cleaved Bid into tBid, which was then
transferred to the mitochondria where it probably combined with
Bax. Then cytochrome c was released to activate caspase-9
and caspase-3 (Fig. 8).
p53 is a transcription factor which plays an
important role in the cellular response to DNA damage through
either cell apoptosis or cell cycle arrest (13,28,29).
In our previous study, tangeretin induced S phase arrest at 24 h
and G2/M arrest at 48 h in AGS cells (17,30).
In the present study, the results showed that p53 expression was
elevated by tangeretin in a dose-dependent manner associated with
upregulation of p21waf1/cip1 and Bax in AGS cells. p21
plays an essential role in growth arrest after DNA damage, and its
overexpression leads to G1 and G2 phase cell cycle arrest (31). Upregulation of
p21waf1/cip1 is also involved in the modulation of
apoptosis in gastric cancer cells (13). Bax expression can be regulated by
p53 (32). To investigate the
function of p53 in tangeretin-treated AGS cells, we observed the
effect of a p53 inhibitor (PFT-α) on the rate of apoptosis of
tangeretin-treated AGS cells. The results showed that PFT-α reduced
the apoptotic rate of AGS cells following treatment with
tangeretin, which confirmed that the apoptosis induced by
tangeretin was p53-dependent. Western blot analysis showed that
PFT-α inhibited the expression of p21, Bax, caspase-3 and
caspase-9, which indicated that p53 was located upstream of these
proteins in the apoptotic pathway. Thus, these results suggest that
tangeretin induces the apoptosis of AGS cells by p53-activated Bax.
Then Bax acts in the mitochondria to cause the release of
cytochrome c, leading to the activation of caspase-9, and
the subsequent activation of caspase-3.
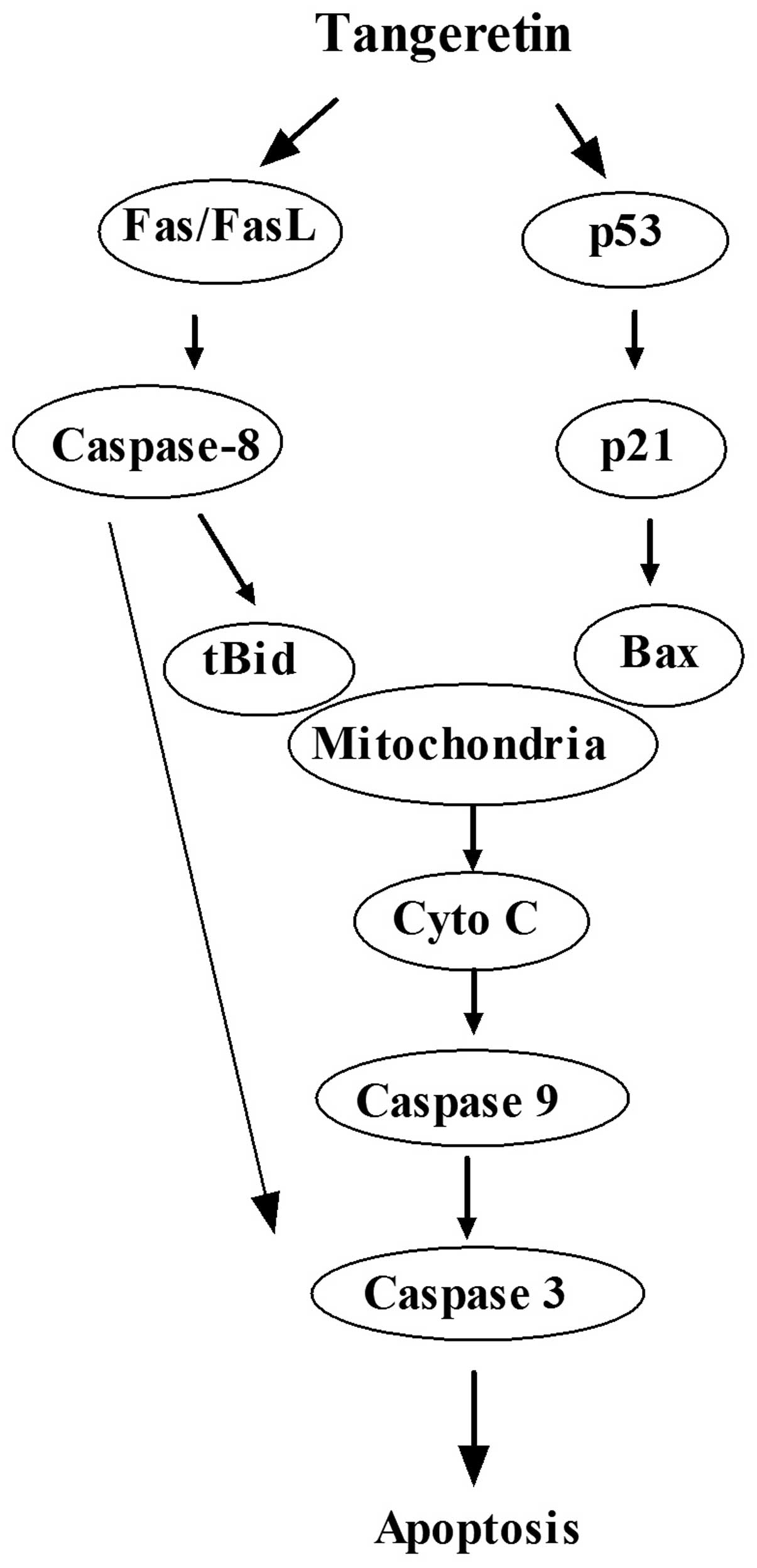
In summary, as shown in Fig. 8, tangeretin induced the apoptosis of
AGS cells through extrinsic and intrinsic signaling pathways.
Activation of p53 induced mitochondrial-mediated apoptosis through
the upregulation of Bax, which contributed to the activation of
caspase-9, leading to the activation of downstream caspases in the
process. Moreover, the Fas/FasL-mediated death receptor pathway may
interact with the mitochondrial signaling pathway through
caspase-8-cleaved Bid.
Acknowledgements
The present study was supported by the National
Science and Technology Major Project of China (2009ZX09311-003);
the Project of Shanghai Leading Academic Discipline, Shanghai
Education Committee (J50305, J50301), China; and the E-Institute of
Traditional Chinese Medicine Internal Medicine, Shanghai Municipal
Education Commission (E 03008), China.
References
|
1
|
Wang D, Wang J, Huang X, Tu Y and Ni K:
Identification of polymethoxylated flavones from green tangerine
peel (Pericarpium Citri Reticulatae Viride) by
chromatographic and spectroscopic techniques. J Pharm Biomed Anal.
44:63–69. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirano T, Abe K, Gotoh M and Oka K: Citrus
flavone tangeretin inhibits leukaemic HL-60 cell growth partially
through induction of apoptosis with less cytotoxicity on normal
lymphocytes. Br J Cancer. 72:1380–1388. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rooprai HK, Kandanearatchi A, Maidment SL,
et al: Evaluation of the effects of swainsonine, captopril,
tangeretin and nobiletin on the biological behaviour of brain
tumour cells in vitro. Neuropathol Appl Neurobiol. 27:29–39. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martinez Conesa C, Vicente Ortega V, Yanez
Gascon MJ, et al: Treatment of metastatic melanoma B16F10 by the
flavonoids tangeretin, rutin, and diosmin. J Agric Food Chem.
53:6791–6797. 2005.PubMed/NCBI
|
|
5
|
Manthey JA and Guthrie N:
Antiproliferative activities of citrus flavonoids against six human
cancer cell lines. J Agric Food Chem. 50:5837–5843. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ishii K, Tanaka S, Kagami K, et al:
Effects of naturally occurring polymethyoxyflavonoids on cell
growth, P-glycoprotein function, cell cycle, and apoptosis of
daunorubicin-resistant T lymphoblastoid leukemia cells. Cancer
Invest. 28:220–229. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morley KL, Ferguson PJ and Koropatnick J:
Tangeretin and nobiletin induce G1 cell cycle arrest but not
apoptosis in human breast and colon cancer cells. Cancer Lett.
251:168–178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan MH, Chen WJ, Lin-Shiau SY, Ho CT and
Lin JK: Tangeretin induces cell-cycle G1 arrest through inhibiting
cyclin-dependent kinases 2 and 4 activities as well as elevating
Cdk inhibitors p21 and p27 in human colorectal carcinoma cells.
Carcinogenesis. 23:1677–1684. 2002. View Article : Google Scholar
|
|
9
|
Chen KH, Weng MS and Lin JK: Tangeretin
suppresses IL-1beta-induced cyclooxygenase (COX)-2 expression
through inhibition of p38 MAPK, JNK, and AKT activation in human
lung carcinoma cells. Biochem Pharmacol. 73:215–227. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lust S, Vanhoecke B, Van Gele M, Philippe
J, Bracke M and Offner F: The flavonoid tangeretin activates the
unfolded protein response and synergizes with imatinib in the
erythroleukemia cell line K562. Mol Nutr Food Res. 54:823–832.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arafa el SA, Zhu Q, Barakat BM, et al:
Tangeretin sensitizes cisplatin-resistant human ovarian cancer
cells through downregulation of phosphoinositide 3-kinase/Akt
signaling pathway. Cancer Res. 69:8910–8917. 2009.PubMed/NCBI
|
|
12
|
Gao J, Senthil M, Ren B, et al: IRF-1
transcriptionally upregulates PUMA, which mediates the
mitochondrial apoptotic pathway in IRF-1-induced apoptosis in
cancer cells. Cell Death Differ. 17:699–709. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang XH, Wong BC, Lin MC, et al:
Functional p53 is required for triptolide-induced apoptosis and
AP-1 and nuclear factor-κB activation in gastric cancer cells.
Oncogene. 20:8009–8018. 2001.PubMed/NCBI
|
|
14
|
Linke SP, Clarkin KC and Wahl GM: p53
mediates permanent arrest over multiple cell cycles in response to
γ-irradiation. Cancer Res. 57:1171–1179. 1997.PubMed/NCBI
|
|
15
|
Midgley CA, Owens B, Briscoe CV, Thomas
DB, Lane DP and Hall PA: Coupling between gamma irradiation, p53
induction and the apoptotic response depends upon cell type in
vivo. J Cell Sci. 108:1843–1848. 1995.PubMed/NCBI
|
|
16
|
Goldstein I, Marcel V, Olivier M, Oren M,
Rotter V and Hainaut P: Understanding wild-type and mutant p53
activities in human cancer: new landmarks on the way to targeted
therapies. Cancer Gene Ther. 18:2–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong Y, Ji G, Shi JR, Cao AL, Xie JQ and
Wu DZ: Arrest of human gastric cancer AGS cells by tangeretin via
cyclinB1 upregulation and inhibition of ERK phosphorylation. Chin
Pharmacol Bull. 28:52012.
|
|
18
|
Cohen GM: Caspases: the executioners of
apoptosis. Biochem J. 326:1–16. 1997.
|
|
19
|
Budihardjo I, Oliver H, Lutter M, Luo X
and Wang X: Biochemical pathways of caspase activation during
apoptosis. Annu Rev Cell Dev Biol. 15:269–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dias N and Bailly C: Drugs targeting
mitochondrial functions to control tumor cell growth. Biochem
Pharmacol. 70:1–12. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schulze-Osthoff K, Ferrari D, Los M,
Wesselborg S and Peter ME: Apoptosis signaling by death receptors.
Eur J Biochem. 254:439–459. 1998. View Article : Google Scholar
|
|
22
|
Rao RV, Ellerby HM and Bredesen DE:
Coupling endoplasmic reticulum stress to the cell death program.
Cell Death Differ. 11:372–380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zamzami N, Marchetti P, Castedo M, et al:
Sequential reduction of mitochondrial transmembrane potential and
generation of reactive oxygen species in early programmed cell
death. J Exp Med. 182:367–377. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zamzami N, Marchetti P, Castedo M, et al:
Reduction in mitochondrial potential constitutes an early
irreversible step of programmed lymphocyte death in vivo. J Exp
Med. 181:1661–1672. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kudla G, Montessuit S, Eskes R, et al: The
destabilization of lipid membranes induced by the C-terminal
fragment of caspase 8-cleaved bid is inhibited by the N-terminal
fragment. J Biol Chem. 275:22713–22718. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Roucou X, Montessuit S, Antonsson B and
Martinou JC: Bax oligomerization in mitochondrial membranes
requires tBid (caspase-8-cleaved Bid) and a mitochondrial protein.
Biochem J. 368:915–921. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schug ZT, Gonzalvez F, Houtkooper RH, Vaz
FM and Gottlieb E: BID is cleaved by caspase-8 within a native
complex on the mitochondrial membrane. Cell Death Differ.
18:538–548. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vousden KH and Lu X: Live or let die: the
cell’s response to p53. Nat Rev Cancer. 2:594–604. 2002.
|
|
29
|
Cui Q, Yu JH, Wu JN, et al: P53-mediated
cell cycle arrest and apoptosis through a caspase-3-independent,
but caspase-9-dependent pathway in oridonin-treated MCF-7 human
breast cancer cells. Acta Pharmacol Sin. 28:1057–1066. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu L, Dong Y, Shi JR and Wu DZ: Tangeretin
enhances the proliferation inhibition of 5-fluorouracil in human
gastric cancer AGS cells. Chin Pharmacol Bull. 27:760–763.
2011.
|
|
31
|
Bunz F, Dutriaux A, Lengauer C, et al:
Requirement for p53 and p21 to sustain G2 arrest after DNA damage.
Science. 282:1497–1501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kurland JF and Meyn RE: Protease
inhibitors restore radiation-induced apoptosis to Bcl-2-expressing
lymphoma cells. Int J Cancer. 96:327–333. 2001. View Article : Google Scholar : PubMed/NCBI
|