Introduction
An extensive body of clinical data and experimental
research has confirmed Stephen Paget's original 'seed and soil'
hypothesis that proposed that the organ-preference patterns of
tumor metastasis are the product of favorable interactions between
metastatic tumor cells (the 'seed') and their organ
microenvironment (the 'soil') (1,2). The
liver is the second most common site of metastasis (spread of
cancer), after the lymph nodes. Liver metastases have been found in
30–70% of patients who are dying of cancer (3). Breast cancer, gastrointestinal cancer,
melanoma and pancreatic cancer all might spread to the liver,
especially colorectal cancer. Unfortunately, only 20% of patients
with colorectal liver metastases present liver-confined resectable
disease and/or are candidates for major surgical operation
(depending on comorbidities) (4,5).
Importantly, 5-year survival rates are reported to be in the range
of 40–58% (6). Based on the above
analysis, it is important to investigate the liver environment and
establish co-culture models for cancer research. Discovery of an
efficient isolating method is definitely the first step, not only
for hepatocytes but also for non-parenchymal cells (NPCs) such as
liver sinusoid endothelial cells (LSECs), hepatic stellate cells
(HSCs) and liver resident immune cells, including Kupffer cells
(KCs), invariant natural killer T (iNKT) cells as well as dendritic
cells (DCs). However, successful and consistent isolation of
primary human hepatic cells remains a challenge for both cell-based
therapeutics and laboratory research (7). Herein, a relatively synchronous (7–14
days for each cell type) and efficient method for isolating primary
human hepatocytes (PHHs) and liver NPCs from surgical specimens is
described.
Materials and methods
Tissue specimens
Liver wedges were obtained from non-diseased tissue
removed at surgical resection for liver tumor, liver metastasis
from colorectal carcinoma or fresh transplanted specimens (n=21;
weight, 20–83 g). The surgical procedures were carried out at the
University Hospital of Munich and Shanghai Jiao Tong University
School of Medicine. All patients provided written documentation of
informed consent. The human liver pieces used in this study were
acquired with consent for governance through the Human Tissue and
Cell Research Foundation (http://www.htcr.de/english/home.html), a
state-controlled non-profit foundation (ref. no. Antrags-Nr.
2014-01). After a pathologist removed what was required for
diagnosis, liver pieces were collected from the remaining tissue.
The tissue sectioned off by the pathologist was morphologically a
healthy tissue obtained from resection margins after liver
resection. The study conforms to the ethical guidelines of the
Declaration of Helsinki and was approved by the Human Tissue and
Cell Research Foundation and the Ethics Committee of the University
Hospital of Munich and Shanghai Jiao Tong University School of
Medicine.
Isolation of PHHs and NPCs
Liver cells were prepared according to a modified
'two-stage' perfusion technique described by Seglen (8) and isolated as previously described
(9). Following 'two-stage'
collagenase (batch no. NB 4G, cat. no. 17465; SERVA Electrophoresis
GmbH, Germany) procedure and manual dissociation of the liver
wedge, the suspension was passed through a sterile nylon mesh of
210 µm (neoLab 4–1413) followed by a sterile nylon mesh of
70 µm (neoLab 4–1419) (both from neoLab Migge GmbH,
Germany). Suspensions were then washed once at 50 × g for 5 min at
4°C in cold storage solution (CSS; Hepacult GmbH, Germany). The
pellet was washed two more times for hepatocyte enrichment. The
NPC-containing cell suspension, as the supernatant obtained after
the first wash, was further used to isolate KCs, LSECs, HSCs, DCs
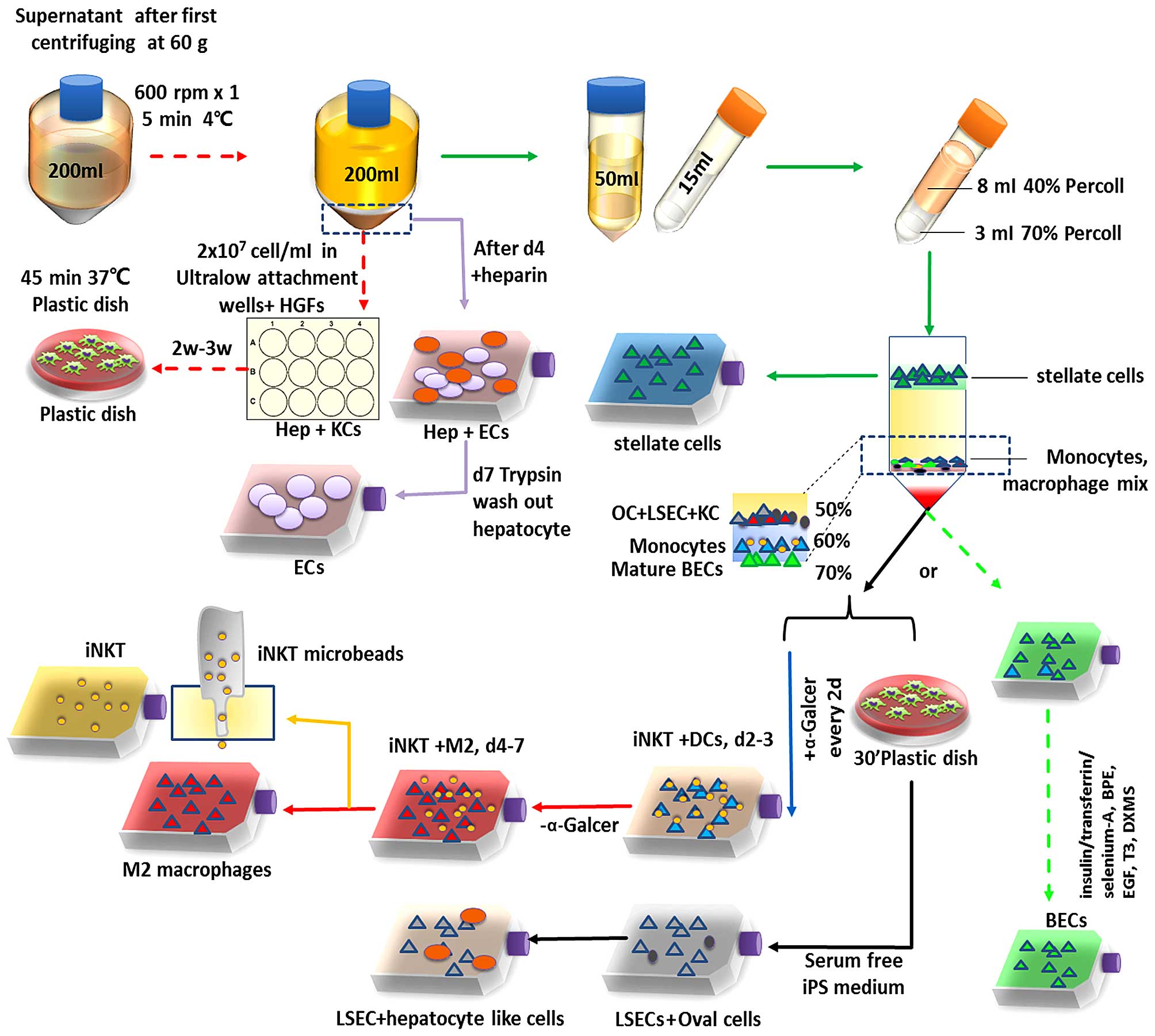
and iNKT cells. The procedure is schematically illustrated in
Fig. 1. In detail, the supernatant
was collected in 200 ml polypropylene centrifuge tubes (cat. no.
352075; BD Biosciences) and pelleted by 600 rpm centrifugation
(Eppendorf 5810R; Eppendorf) for 5 min at 4°C.
The pellet was then resuspended and seeded in
12-well ultra-low attachment plates (Sigma-Aldrich, Germany) at a
density of 1.5–2×107 viable cells/cm2 by
using Dulbecco's modified Eagle's medium (DMEM)/Ham's F-12 (Gibco
Life Technologies, Germany). This pellet contained the remaining
hepatocytes and KCs; the mixture was co-cultured for 2–3 weeks with
10 µg/ml HGF (red arrow, Fig.
1). The cells were collected at 14–21 days from each well and
washed three times, and then the cells were transferred into 100-mm
non-tissue culture grade plastic dishes. Incubation was carried out
for 30 min at 37°C in an atmosphere of 5% CO2 and ~95%
air. Macrophages readily attached to the non-tissue culture grade
plastic dishes under this incubation process, whereas other types
of cells did not. Two milliliters of trypsin solution was added
into the dish and incubated for 10 min at 37°C in an atmosphere of
5% CO2 and ~95% air. Then 10 ml of the growth medium
were added and the cells were gently scraped. The collected KCs
were then centrifuged at 1,500 rpm for 5 min, and propagated in
75-cm2 flasks (red dashed line in Fig. 1).
The supernatant washed with CSS was collected in a
50-ml polystyrene conical Falcon tube (BD Biosciences) and pelleted
at 1,500 rpm for 7 min at 4°C. After three times washing, the
pellet was seeded in collagen I-coated empty tissue culture
75-cm2 flasks or 6-well Corning BioCoat Collagen I
Multiwell Plates (both from Corning, Inc.) with DMEM/F-12 and EGF,
10% FBS, 100 U/ml penicillin, 0.1 mg/ml streptomycin, and 2 mM
L-glutamine for hepatic microvascular endothelial cell (EC)
isolation (purple arrow, Fig. 1).
The assessment of yield and viability was made by using a
hemocytometer-based trypan blue exclusion assay. DMEM/F-12 plus 0.1
ng/ml recombined human EGF and 90 µg/ml heparin (10 ng/ml
insulin-like growth factor was also optimal) were used from day 4
which lasted for ~7–8 days. On day 7, the hepatocytes were detached
by trypsin (PAA) for 4–5 min and the procedure was observed by
microscopy. Hepatocytes were then washed out, and the rest were
microvascular epithelial cells (Fig.
2).
Or the pellet was processed as mentioned in step 1.
The supernatant was placed in 50-ml Falcon tubes after washing once
with RPMI-1640 medium by centrifugation at 800 × g for 5 min. The
cells were then resuspended in 8 ml 30% Percoll™ PLUS/Percoll (GE
Healthcare Life Sciences, Freiburg, Germany), placed in 15-ml
polystyrene conical centrifuge tube (BD Biosciences) and overlaid
with 3 ml 70% Percoll solution. After being centrifuged at 2,500
rpm for 20 min at room temperature with decreased acceleration and
without breaks, the various cell types were arranged according to
their density. HSCs were enriched in an upper cell layer, whereas
macrophages/KCs, oval cells and LSEC fractions were floating
between 50 and 60% Percoll. DCs, monocytes and lymphocytes were
enriched in the interphase of 60 and 70% Percoll. Mature
intrahepatic biliary epithelial cells were floating on the 70%
Percoll. Cell fractions were collected separately by pipetting
(10).
For the immune cell enrichment, the monocyte
fraction was pelleted, resuspended in 1 ml of 10X RBC lysis buffer
and transfered to a new tube. The cell suspension was allowed to
stay for 3 min at room temperature to lyse RBCs, and was then
diluted with 9 ml RPMI-1640 medium, centrifuged at 800 × g for 5
min at 4°C and the cells were washed twice with 10 ml of RPMI-1640
medium. The live cell number was counted and adjusted to a cell
concentration of 1×106 cells/ml and placed in
75-cm2 flasks after being washed three times. A total of
100 ng/ml α-galactosylceramide (α-Galcer; Abcam, UK) was added 8 h
after seeding. The medium containing α-Galcer was changed every 2–3
days. DCs were induced and enriched in the first 48 h, being
replaced by monocyte-induced macrophages after 72 h. On day 7–10,
the supernatant was collected and washed three times. After being
adjusted to a cell concentration of 1×106 cells/ml by
adding FACS staining buffer, the cells were labeled with iNKT
(clone 6B–11) microbeads (human TCR Vα24-Jα18 combined with Vβ11;
Miltenyi Biotec GmbH, Germany) according to the manufacturer's
instructions. Cells were applied onto LS magnetic-activated cell
sorting (MACS) columns (Miltenyi Biotec GmbH) twice, which were
placed within the magnetic field of a MACS separator and washed
three times with MACS buffer (Miltenyi Biotec GmbH). Vα24-Jα18
combined with Vβ11-positive iNKT cells were eluted from the
columns, and the viable iNKT cells were counted and propagated in
flasks.
DCs were enriched on day 2 or 3 as attached cells.
However, other monocytes were in the suspension. These cell
suspensions were collected and were propagated in another flask.
The remaining cells were detached by trypsin (PAA) and were
incubated with human IgG (1 µg/106 ml; BioLegend,
Inc, San Diego, CA, USA) to block non-specific binding to Fcγ
receptors for 15 min. The cells were then labeled with CD1c
microbeads. CD1c+ DCs were propagated in culture plates
(blue arrow, Fig. 1) after magnetic
sorting.
Cell suspension as mentioned in step 4 was collected
and propagated in another flask as step 4. The former flasks which
contained the attached cells were refilled with 20 ml completed
growth medium. M2-differentiated macrophages were enriched on day
4–7, iNKT cells was also separated here (orange arrow, red arrow,
Fig. 1). Alternatively, culture
plates coated by the attached cells from the last step were
cultivated with DMEM containing IL-4 and GM-CSF for 5 days, and
M2-differentiated macrophages were harvested.
The macrophage/KC fraction was pelleted and KCs were
also labeled with APC-conjugated anti-Clec4f antibody (LS-C233943,
LifeSpan Biosciences, Inc., USA) for 45 min at 10 µl/100
µl after a 15-min incubation of human IgG blocking, followed
by 10 µl/100 µl anti-APC-microbead incubation
(Miltenyi Biotec GmbH) and magnetic cell sorting according to the
manufacturer's instructions. Cells were resuspended in growth
medium and seeded at a sufficient density of 1–2×106
cells/cm2.
For the stem cell enrichment, the fraction between
50 and 60% Percoll was pelleted and seeded in a 75-cm2
collagen I-coated flask after being washed three times and
cultivated in a plastic dish for 30 min to eliminate KCs.
Serum-free iPSC medium plus EGF was placed for the cell culture.
ECs were proliferated from day 3. Hepatic progenitor cell-like
cells differentiated to hepatocytes surrounding the EC niches from
day 5.
For the intrahepatic biliary epithelial cell
isolation and purification, the fraction floating on 70% Percoll
was seeded in 75-cm2 collagen I-coated empty tissue
culture flasks (Corning, Inc.). Cells were grown in medium
containing DMEM/F-12 (Sigma-Aldrich, St. Louis, MO, USA)
supplemented with fetal bovine serum (Cellgro; Mediatech, Inc.
Manassas, VA, USA), penicillin/streptomycin, vitamin solution, MEM
solution, CD lipid concentrate, L-glutamine,
insulin/transferrin/selenium-A, bovine pituitary extract, epidermal
growth factor, 3,3′,5-triiodo-L-thyronine and dexamethasone.
Identification of cell populations
Morphological characteristics were determined by
phase contrast images acquired with an Olympus IX50 microscope
(Olympus, Germany). Cells cultured in 8-well chamber slides
(Falcon; BD Biosciences, Germany) and culture inserts (Ibidi GmbH,
Martinsried, Germany) were washed twice with cold Tris-buffered
saline (TBS), pH 7.4, fixed with 4% paraformaldehyde for 10 min,
permeabilized with 0.1% Triton X-100 for 5 min (for the staining of
extracellular markers, permeabilization was not necessary), blocked
with 5% BSA, incubated with the indicated primary antibodies:
rabbit anti-human Clec4f (Sigma-Aldrich, Germany), mouse anti-human
CD68 (Sigma-Aldrich), rabbit anti-human epithelial cell adhesion
molecule (EpCAM, Cell Signaling Technology, Inc., UK), anti-GFAP
and anti-desmin (Sigma-Aldrich), anti-α-SMA (Abcam), anti-OV-6
(Santa Cruz Biotechnology, Inc., Paso Robles, CA, USA),
cytokeratin-19 (CK-19; Santa Cruz Biotechnology, Inc.), rabbit
anti-human VE-cadherin (Cell Signaling Technology, Inc.) and
APC-iNKT (Miltenyi Biotec GmbH) at 4°C overnight followed by
anti-rabbit Alexa Fluor 488 and anti-mouse Alexa Fluor
568-conjugated secondary antibodies (Life Technologies, Germany).
The cells were then stained with anti-fade DAPI (Life Technologies)
for nuclear staining, and the images were acquired with an Olympus
Axion microscope (Olympus, Japan).
3D cell culture
Two 3D cell culture models were used in this study,
ultra-low attachment plates (Sigma-Aldrich) and HDP1096
Perfecta3D® 96-Well Hanging Drop Plates (3D Biomatrix,
Inc.). For the hanging drop plates, the pellet collected from step
1 was washed three times and a 100 cell/µl cell suspension
was prepared. Hanging drops were formed by pipetting 45 µl
of cell suspension into each well from the top side of the plate.
Hanging drops were formed on and confined to the bottom of the
plate. The lid was placed on the plate and the assembly was placed
into a tissue culture incubator. Within hours, individual cells
started to aggregate and eventually formed into spheroids. Media
exchange followed the protocol supplied by 3D Biomatrix, Inc.
DMEM/F-12 with HGF, 10% FBS, 100 U/ml penicillin, 0.1 mg/ml
streptomycin, and 2 mM L-glutamine was used for culture
maintenance.
Statistical analysis
All continuous values are expressed as means ± SD
and all experiments were repeated three times. Immunofluorescence
staining of cell type-specific markers was performed in five
independent experiments. For quantitative analysis, DAPI-stained,
marker-positive and -negative cells were counted in 10 independent
images per cell type at ×20 magnification. The results were
subjected to a non-parametric Mann-Whitney U test. A paired
Student's t-test was also used to analyze the intragroup
differences. All statistical analyses were conducted using StatView
5.0 for Windows (SAS Institute, Inc., Cary, NC, USA). Student's
t-test was also used to test differences in cell viability assays.
P<0.05 was considered statistically significant.
Results
Hepatic (microvascular) EC isolation,
primary culture and subculture based on the co-culture system
The supernatant from the first washing was
centrifuged, the pellet was resuspended and plated in collagen
I-coated flasks and processed as detailed in 'Materials and
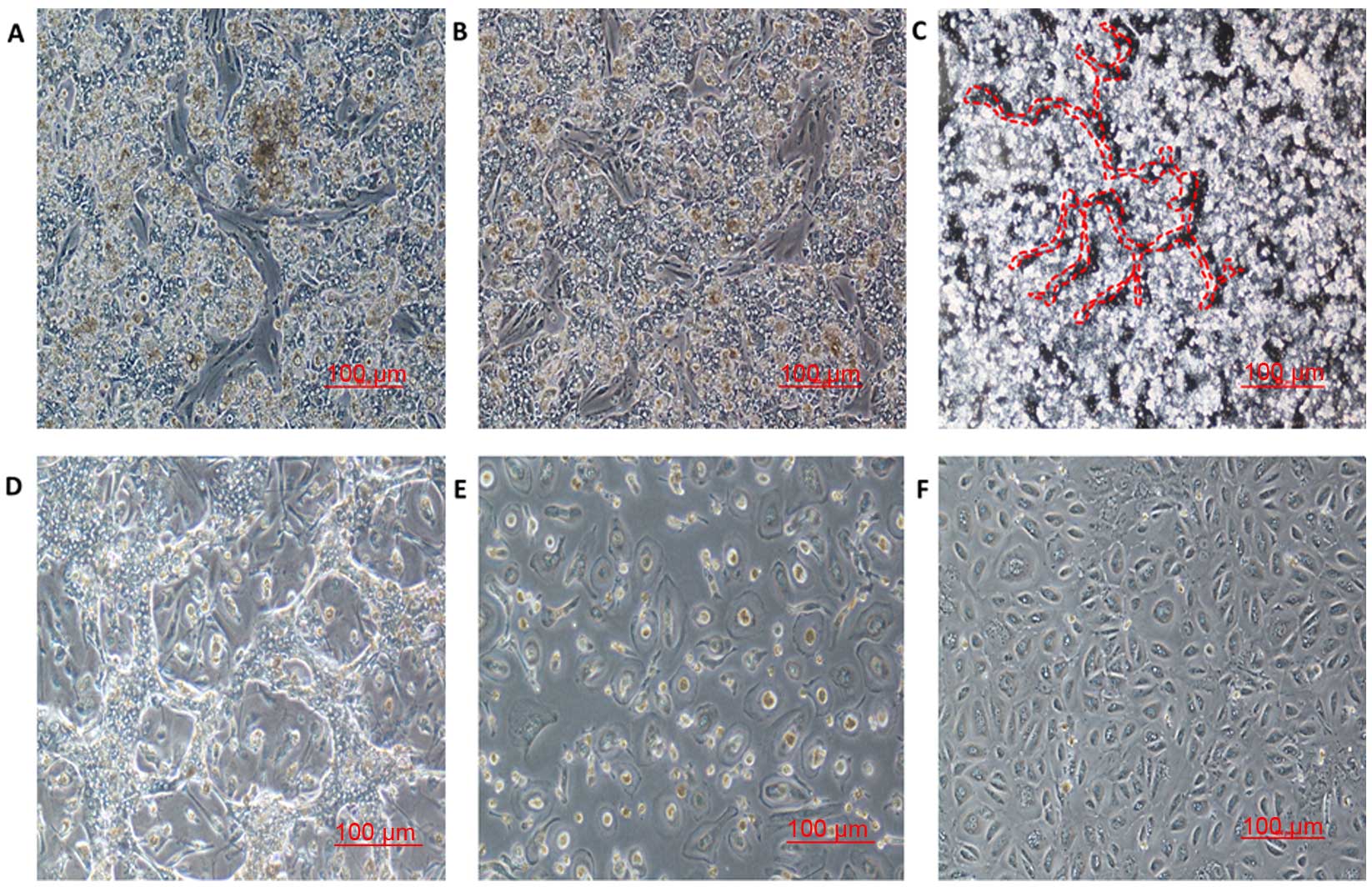
methods'. Fig. 2A and B show
representative images of PHHs and microvascular ECs, in culture on
days 4 and 5 after isolation. The hepatocytes formed an island-like
shape with an irregular outline. In the rest space proliferated ECs
were noted until day 7–9 (Fig. 2B).
Fig. 2C and D shows the
morphological features of the hepatocyte and EC co-culture system 6
and 7 days after isolation. The ECs exhibited cobblestone
morphologic characteristics, which resembled the shape of
microvascular epithelial cells (11). The cultured ECs were harvested as
early as day 8, by detachment of hepatocytes. After a 4–5-min
trypsin treatment, the hepatocytes were soon detached, however ECs
were still attached (Fig. 2E). The
number of ECs reached maximal levels on day 12–14, when the cells
formed a flat cell sheet and tight conjunction on the plate surface
(Fig. 2F). When the colonies of ECs
reached 80% confluency, the cells were detached by trypsin for 8–10
min and subcultured in new collagen I-coated tissue culture flasks.
The purity of the ECs was 92±1.2% as characterized by VE-cadherin
and EpCAM.
KC isolation and purification based on
the 3D co-culture system
Similar to what was observed in the 2D co-culture
system for EC isolation, the island-like shape of hepatocytes was
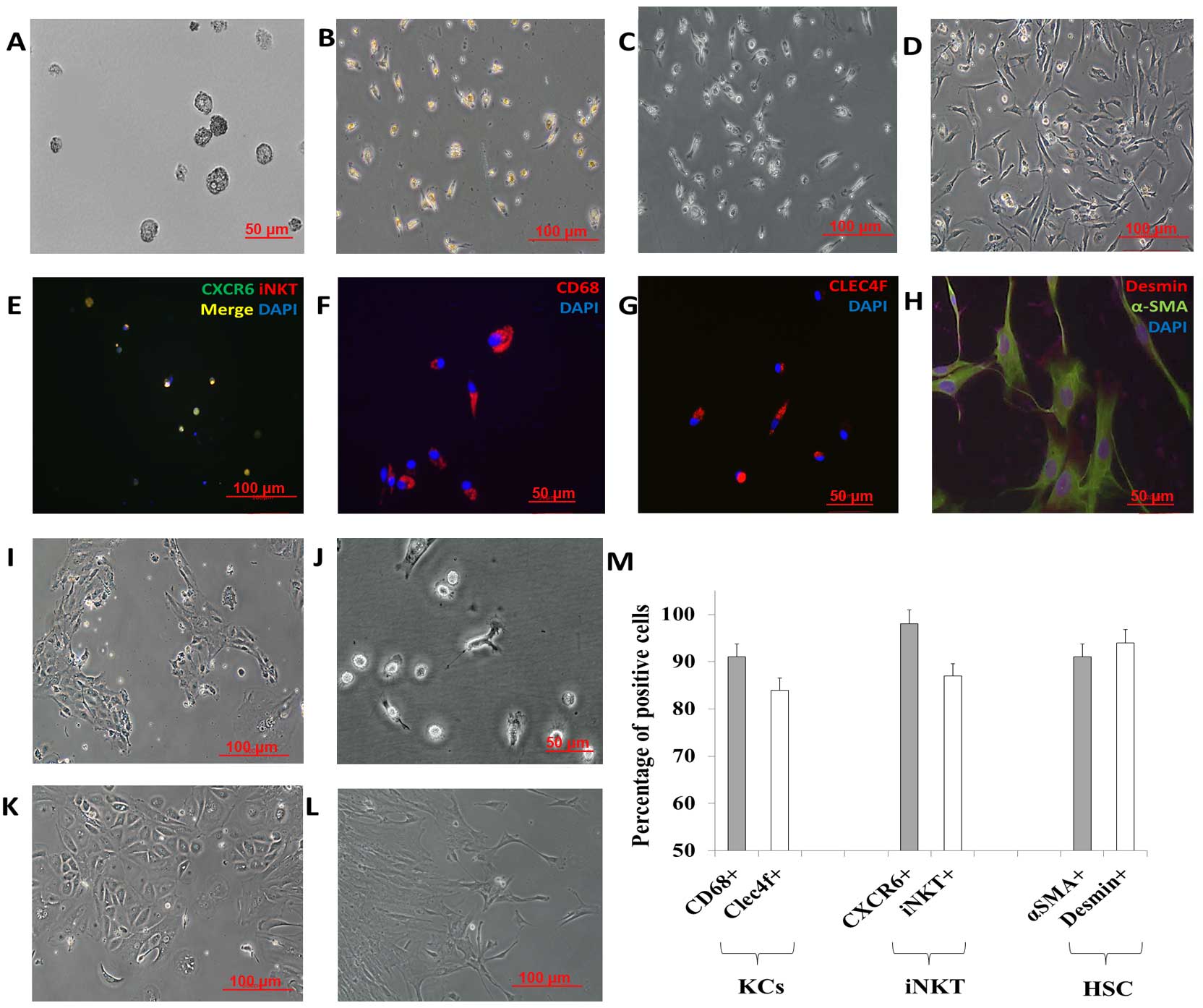
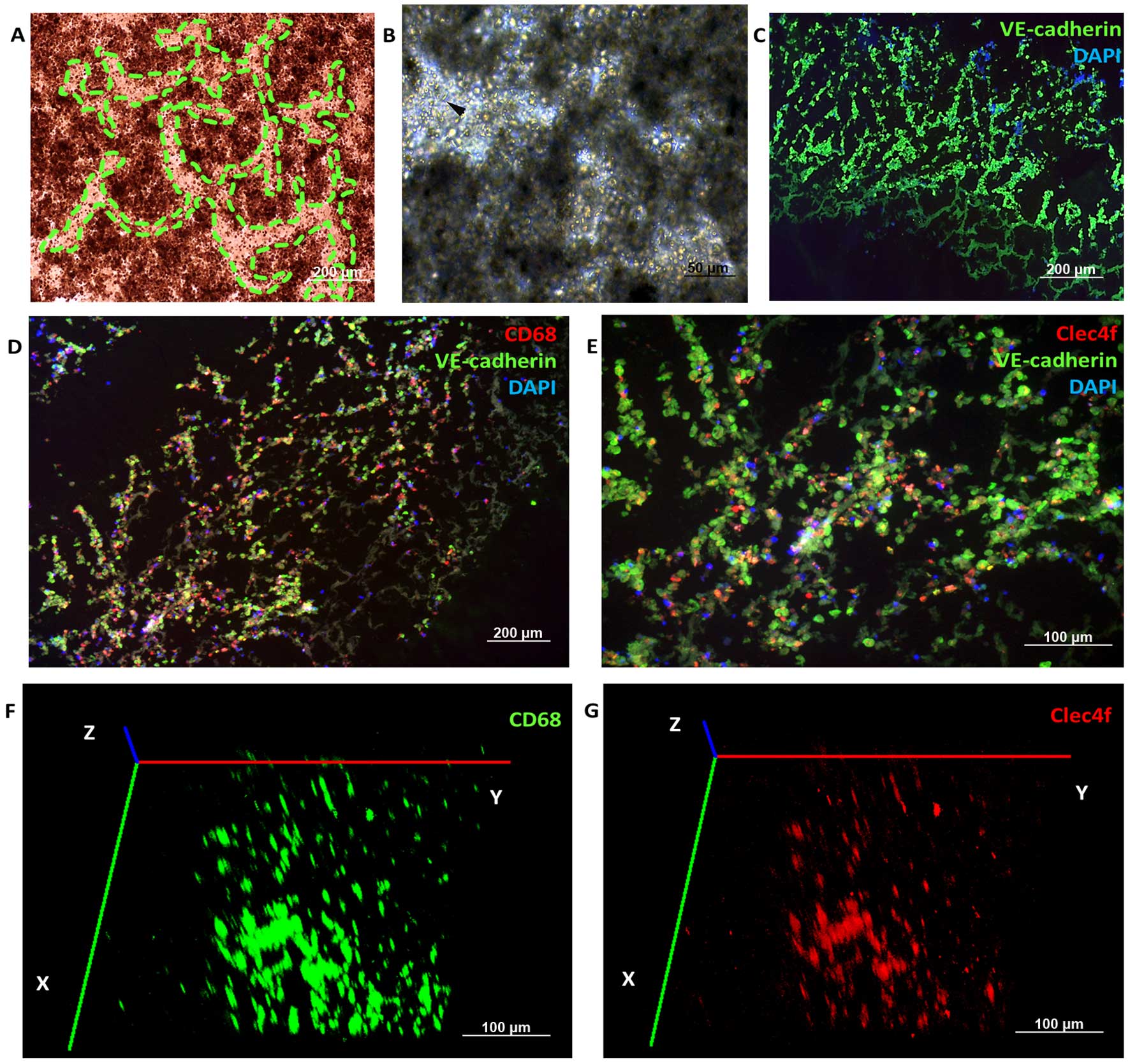
also observed in the 12- or 96-well 3D co-culture system (Fig. 3A). Spindle cells were found in the
rest space (Fig. 3B). Primary KCs,
prepared from the 3D co-culture system and plastic dish attachment
as described in step 1, formed typical irregular morphology of both
a stretched and an oval shape (12). Immunofluorescence staining was
performed using the same methods as below. The medium in each well
was carefully removed and gently refilled with embedding medium for
cryosectioning (Leica, Germany), followed by freezing of the
plates. The 3D co-culture model then underwent the
immunofluorescence analysis. A net structure was found, and the
connecting tissue was positively stained with VE-cadherin, CD68 or
Clec4f (Fig. 3C–E), which were ECs
and KCs/macrophages. Clec4f+ KCs were less than the
CD68+ macrophages in the 3D organotypic co-culture
models after whole-mount staining by two-photon microscope (Carl
Zeiss, Germany). In addition, the cells collected from the other
wells were collected, centrifuged and washed three times. After
that, KCs were enriched using the isolation method above (step 1).
A total of 87±6.3% of these cells was Clec4f-positive (Fig. 6G and M), which were KCs rather than
peripheral derived macrophages (13).
LSEC isolation and heterogeneous
endothelium populations
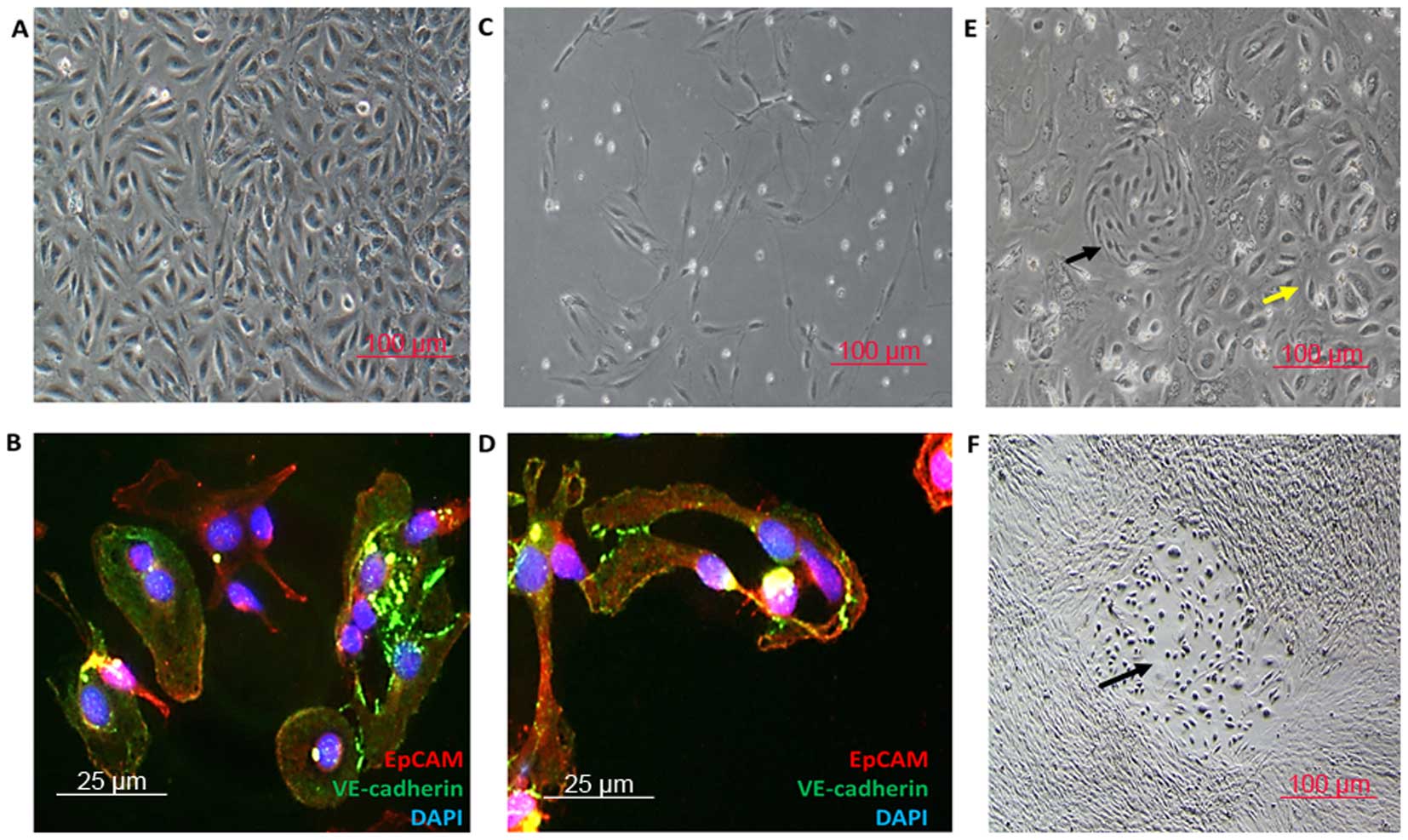
The morphology of LSECs was different from the
microvascular ECs (14,15) (Fig. 4A
and C), although both were VE-cadherin+
EpCAM+ (Fig. 4B and D),
especially when observed in the co-culture system of ECs and LSECs
(Fig. 4E). In addition, the LSECs
tended to form hepatic EC niche when co-cultivated with hepatocytes
or NPCs (Fig. 4F). However, it was
difficult to tell the differences between the hepatic microvascular
ECs and ECs from the hepatic lymphatic vessels (Fig. 6K) according to the morphology.
Lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1)
unequivocally distinguishes lymphatic from blood microvascular
endothelium (16), as assessed by
immunostaining with the anti-LYVE-1 antibody.
Hanging drop models for 3D hepatocyte
culture
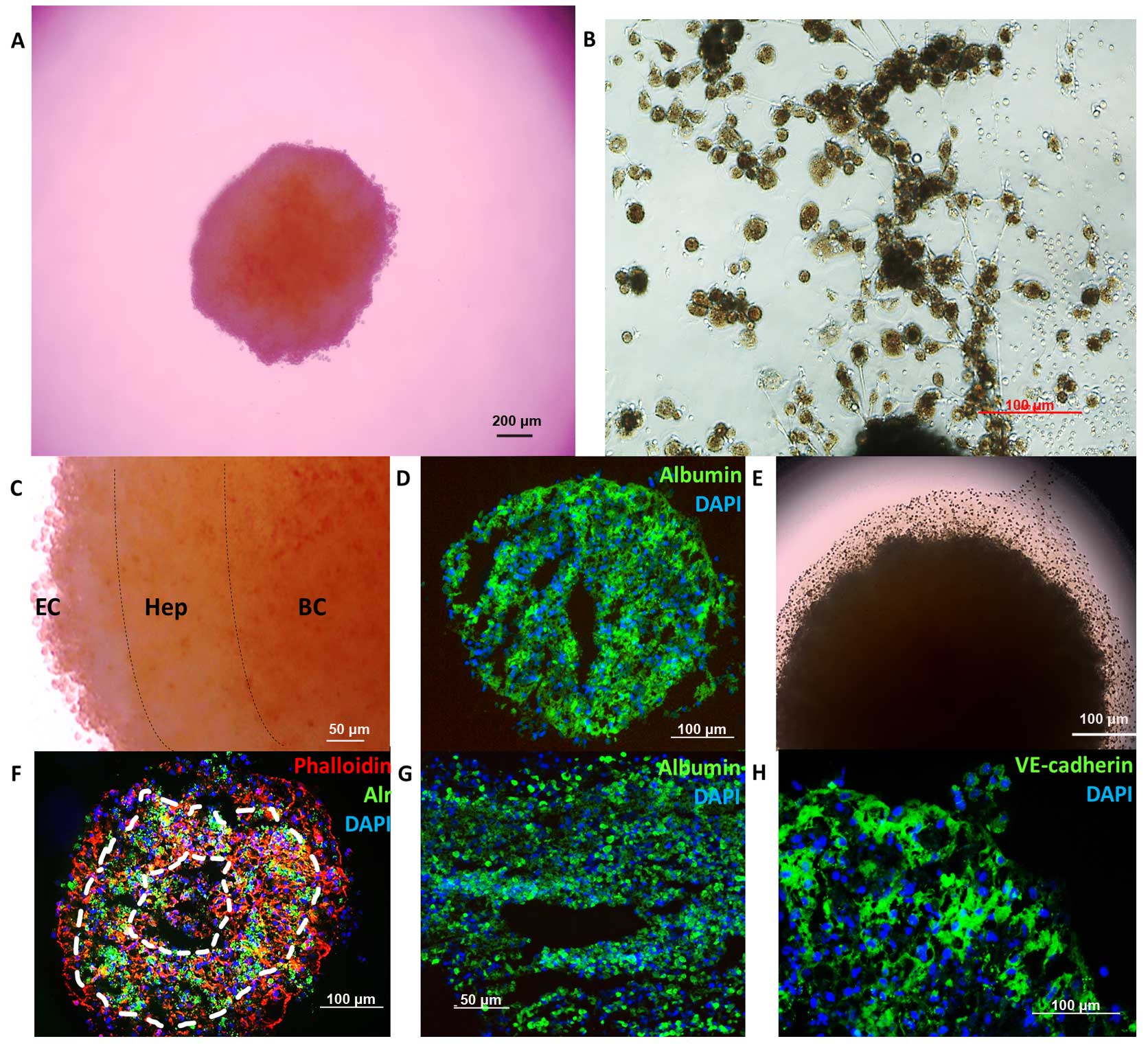
Spheroid formation time was 48–72 h and lasted for
9–12 days. Sponges were observed from day 10–12, which were formed
by ECs and hepatocytes. VE-cadherin+ ECs not only were
detected in the sponges but also in the spheroid. According to the
color variations and immunofluorescence, the spheroid was formed by
three layers. Blood cells were the major source of the core,
hepatocytes formed the second layer and maintained the capability
of regeneration. However, ECs and other NPCs constituted the cover
of the spheroid (Fig. 5).
Characterization of other NPCs
The complete yield of the HSCs, separated from the
NPC suspension by density gradient centrifugation, was seeded into
a 75-cm2 tissue culture flask using stellate cell growth
medium (cat. no. 5301; ScienCell Research Laboratories, Inc.). Once
90% confluency was reached, the cells were detached by trypsin and
seeded into plastic dishes using DMEM supplemented with 10% FBS,
100 U/ml penicillin, 0.1 mg/ml streptomycin, and 2 mM L-glutamine.
Freshly isolated HSCs exhibited the phenotypically star-shaped
morphology and were transformed into an activated state, as
identified by a myofibroblast-like phenotype within 10 days of cell
culture. In the present study, HSCs were stained for α-SMA, desmin
and GFAP, well-known markers of HSCs to distinguish HSCs from
myofibroblasts as described elsewhere (17). Imaging revealed high protein
expression of desmin and GFAP in the HSCs. Most of the HSCs were
α-SMA-positive cells, which confirmed the identity of the activated
HSCs. Intrahepatic human biliary epithelial cells (hBECs) were
semi-purified by centrifugation on a Percoll gradient and were
further immunopurified. hBECs were cytokeratin-7
(CK-7+), CK-19+ (18,19)
and α-SMA− cells.
Hepatic progenitor cells were driven to
differentiate into hepatoblasts
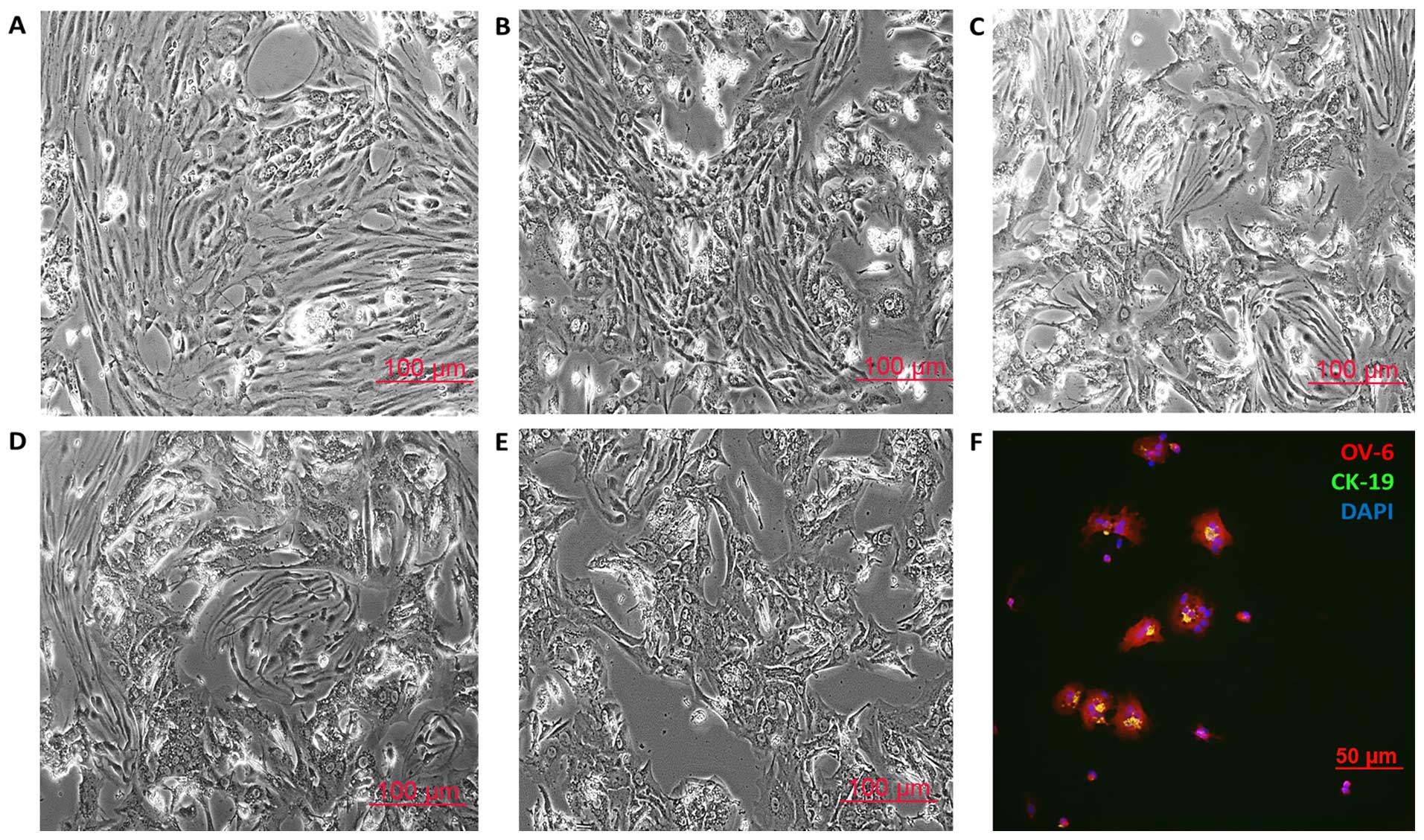
The oval cells were collected as described above.
Cells were cultured using iPSC cell growth medium (Miltenyi Biotec
GmbH) with 10 ng/ml HGF. On day 0, the oval cells (hepatic
progenitor cells) displayed small oval morphology with a dark
nucleus and few cytoplasma. Immunostaining revealed that most of
the cells were positive for the definitive markers CK-19 and OV-6
(Fig. 7F) and the hepatic
progenitor cell marker EpCAM (data not shown) (20–22).
On day 2 the oval cells started to display typical LSEC morphology,
and LSECs were enriched on day 3–4. The number of LSECs reached
maximal levels on day 4–6. Five to six days of iPSC cell growth
medium and HGF-induction treatment resulted in the appearance of
hepatocyte-like cells, which are reported to be hepatoblasts
(Fig. 7A) (23). Fig.
7B–E shows the enrichment of the hepatoblasts on day 7–9.
Discussion
In order to improve the research platform for cancer
research, especially for in vitro liver metastatic model
establishment, functional and reproducible human liver cell
co-culture models as well as high quality and quantity hepatocytes
and non-parenchymal hepatic cells are essential. Recently, Pfeiffer
et al isolated human hepatocytes and NPCs from tissue
obtained by partial liver resection by a two-step EDTA/collagenase
perfusion technique. The obtained cell fractions were purified by
Percoll density gradient centrifugation. KCs, LSECs and HSCs were
further separated using selective adherence to plastic. LSECs were
separated by MACS using CD31 microBeads (24). Most recently, Werner et al
established a three layer iodixanol density gradient using 9 and
12.6% iodixanol to separate NPCs. The advantage of the method of
Werner et al is that LSECs and KCs can be separated as one
fraction from the HSC population. These two methods were reported
and approved to be available for high quality and quantity
(25). In order to increase the
purity, magnetic beads specific for CD31 or CD146 were used to
label LSECs. Both methods provide cell yields comparable to those
obtained by single and multiple cell type isolation methods.
In the present study, we established an efficient
and co-culture isolation method based on 3D cell co-culture,
long-term cell co-culture, density gradient centrifugation and
MACS. The presented method provided high purities and comparable
cell yields to the methods above.
Most recently, Lavin et al identified 12,743
macrophage-specific enhancers and established that tissue-resident
macrophages have distinct enhancer landscapes beyond what can be
explained by developmental origin. Clec4f was reported to be the
biomarker of liver-resident macrophages, as known KCs, which would
be a specific marker for KC characteristics (13). In the present study, we found less
Clec4f+ KCs than CD68+ macrophages. However,
in the co-culture model, 87±6.3% of the isolated macrophages were
Clec4f-positive, which might also be differentiated tissue-resident
macrophages that were reprogrammed by the 3D cultured liver
microenvironment.
The liver has a dual blood supply. The hepatic
artery and portal vein both drain into the hepatic sinusoids, which
represent the capillary network in the liver. The endothelial
lining of liver sinusoids demonstrates microheterogeneity between
periportal and centrilobular regions (26). Nolan et al identified LSECs
as
VEGFR3+IsolectinGSIB4+CD34dim/−IgG−
cells (27).
VE-cadherin+ ESC− ECs (embryonic stem cell
differentiation into ECs) were found to consist of ~60% of the
vasculature in the regenerating liver in the same report. In line
with this study, our study showed that both LSECs and microvascular
ECs expressed VE-cadherin. To increase the purity of the LSECs,
VEGFR3+IsolectinGSIB4 +CD34dim/−
cell sorting would be recommended as an available strategy. Many
groups used CD31 as a sorting strategy for LSECs (28,29),
however, it was reported that purified ESC-ECs maintained 99.3%
VE-cadherin and CD31 positivity for at least 4 weeks after
purification (25).
With only <1% of the NPC population as hepatic
DCs, isolating a sufficient amount of hepatic DCs is a timely and
costly task. Several methods using cytokine stimulation have been
developed to increase the total hepatic DC population in
vitro and in vivo (30).
Joyee et al provide direct evidence that iNKTs
preferentially promote the functional development of DCs (31). Bollyky et al also found that
iNKT cell activation with α-Galcer induced mature DCs (23). In this study, α-Galcer, as a
specific ligand for human and mouse natural killer T (NKT) cells,
was added into the culture system of monocytes. We supposed that
iNKT cells could be selectively activated so that the DCs, mostly
monocyte-derived dendritic cells (moDCs) would be induced. However,
we only separated CD1c+ DCs in this study, which was the
major subset of classic human DCs as well as inflammatory DCs
(32). The subgroups of DCs need to
be further analyzed by flow cytometry.
Concerning the biliary epithelial cells, their
limited number (3% of the total liver mass) and their intrahepatic
localization have limited the development of in vitro cell
models in order to gain molecular insights into their function
(33). In the present study, we
isolated hBECs from chronic liver injury, most of which were liver
metastasis, as it was reported that chronic cell injury such as
chronic hepatitis elicited mobilization of hepatic endodermal stem
cells or oval cells, which can give rise both to hepatocytes and
cholangiocytes (34).
Reports have suggested that a 3D liver cell
co-culture system is a promising approach to mimic the tissue
architecture in vivo (35,36).
Kostadinova et al (37) and
Messner et al (35) reported
the earliest investigation of 3D co-culture liver model. Messner
et al showed that the accumulation of hepatocytes and NPCs
in hanging drops resulted in microtissue formation within 3 days.
Immunohistochemical staining showed hepatocytes, CD68+
KCs as well as CD31+ ECs. In addition, the functionality
of liver microtissues was reported to be preserved for more than 5
weeks, as indicated by persistent albumin secretion. However, the
PHHs in our hanging drop model did not attach after a 2-week
cultivation (27). And the
functionality of the PHHs was not as sufficient as reported. This
might be because in our study, the cell number (>5,000 cells)
and diameter of the spheroids were larger than these parameters in
the previous studies. It is recommended to investigate the
functionality of NPCs and PHHs on organotypic co-culture
models.
In conclusion, we developed an efficient and 3D cell
co-culture isolation method. Isolated KCs, sinusoid ECs, ECs,
biliary epithelial cells, DCs, stellate cells, hepatic progenitor
cell-like cells and iNKT cells can be further used as co-cultures
for cancer research in particular tumor immunology and liver
metastasis.
Acknowledgments
We would like to thank Wolfgang E. Thasler, and
Hepacult GmbH for their assistance with the hepatocyte isolation.
This study was supported, in part, by the China Scholarship Council
(CSC) (201306230127) and the German Academic Exchange Service
scholarship.
References
|
1
|
Paget S: The distribution of secondary
growths in cancer of the breast. 1889 Cancer Metastasis Rev.
8:98–101. 1989.
|
|
2
|
Langley RR and Fidler IJ: Tumor cell-organ
microenvironment interactions in the pathogenesis of cancer
metastasis. Endocr Rev. 28:297–321. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wanebo HJ, Chu QD, Avradopoulos KA and
Vezeridis MP: Current perspectives on repeat hepatic resection for
colorectal carcinoma: A review. Surgery. 119:361–371. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Malafosse R, Penna C, Sa Cunha A and
Nordlinger B: Surgical management of hepatic metastases from
colorectal malignancies. Ann Oncol. 12:887–894. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bentrem DJ, Dematteo RP and Blumgart LH:
Surgical therapy for metastatic disease to the liver. Annu Rev Med.
56:139–156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pawlik TM, Schulick RD and Choti MA:
Expanding criteria for resectability of colorectal liver
metastases. Oncologist. 13:51–64. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhogal RH, Hodson J, Bartlett DC, Weston
CJ, Curbishley SM, Haughton E, Williams KT, Reynolds GM, Newsome
PN, Adams DH, et al: Isolation of primary human hepatocytes from
normal and diseased liver tissue: A one hundred liver experience.
PLoS One. 6:e182222011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seglen PO: Preparation of rat liver cells.
3 Enzymatic requirements for tissue dispersion. Exp Cell Res.
82:391–398. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhogal RH, Hodson J, Bartlett DC, Weston
CJ, Curbishley SM, Haughton E, Williams KT, Reynolds GM, Newsome
PN, Adams DH, et al: Isolation of primary human hepatocytes from
normal and diseased liver tissue: A one hundred liver experience.
PloS one. 6:e182222011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rothe GM: Electrophoresis of Enzymes:
Laboratory Methods (Springer Lab Manuals). 1st edition.
Springer-Verlag; Berlin, Heidelberger: pp. 39–45. 1994
|
|
11
|
Holmén C, Christensson M, Pettersson E,
Bratt J, Stjärne P, Karrar A and Sumitran-Holgersson S: Wegener's
granulomatosis is associated with organ-specific antiendothelial
cell antibodies. Kidney Int. 66:1049–1060. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Isidro RA, Bonilla FJ, Pagan H, Cruz ML,
Lopez P, Godoy L, Hernandez S, Loucil-Alicea RY, Rivera-Amill V,
Yamamura Y, et al: The probiotic mixture VSL#3 alters the
morphology and secretion profile of both polarized and unpolarized
human macrophages in a polarization-dependent manner. J Clin Cell
Immunol. 5:10002272014.PubMed/NCBI
|
|
13
|
Lavin Y, Winter D, Blecher-Gonen R, David
E, Keren-Shaul H, Merad M, Jung S and Amit I: Tissue-resident
macrophage enhancer landscapes are shaped by the local
microenvironment. Cell. 159:1312–1326. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Daneker GW, Lund SA, Caughman SW, Swerlick
RA, Fischer AH, Staley CA and Ades EW: Culture and characterization
of sinusoidal endothelial cells isolated from human liver. In Vitro
Cell Dev Biol Anim. 34:370–377. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elvevold K, Smedsrød B and Martinez I: The
liver sinusoidal endothelial cell: A cell type of controversial and
confusing identity. Am J Physiol Gastrointest Liver Physiol.
294:G391–G400. 2008. View Article : Google Scholar
|
|
16
|
Podgrabinska S, Braun P, Velasco P, Kloos
B, Pepper MS and Skobe M: Molecular characterization of lymphatic
endothelial cells. Proc Natl Acad Sci USA. 99:16069–16074. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yin C, Evason KJ, Asahina K and Stainier
DY: Hepatic stellate cells in liver development, regeneration, and
cancer. J Clin Invest. 123:1902–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Massani M, Stecca T, Fabris L, Caratozzolo
E, Ruffolo C, Furlanetto A, Morton S, Cadamuro M, Strazzabosco M
and Bassi N: Isolation and characterization of biliary epithelial
and stromal cells from resected human cholangiocarcinoma: A novel
in vitro model to study tumorstroma interactions. Oncol Rep.
30:1143–1148. 2013.PubMed/NCBI
|
|
19
|
Muff MA, Masyuk TV, Stroope AJ, Huang BQ,
Splinter PL, Lee SO and Larusso NF: Development and
characterization of a cholangiocyte cell line from the PCK rat, an
animal model of Autosomal Recessive Polycystic Kidney Disease. Lab
Invest. 86:940–950. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duret C, Gerbal-Chaloin S, Ramos J, Fabre
JM, Jacquet E, Navarro F, Blanc P, Sa-Cunha A, Maurel P and
Daujat-Chavanieu M: Isolation, characterization, and
differentiation to hepatocyte-like cells of nonparenchymal
epithelial cells from adult human liver. Stem Cells. 25:1779–1790.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Terrace JD, Currie IS, Hay DC, Masson NM,
Anderson RA, Forbes SJ, Parks RW and Ross JA: Progenitor cell
characterization and location in the developing human liver. Stem
Cells Dev. 16:771–778. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Xin J, Zhang L, Wu J, Jiang L, Zhou
Q, Li J, Guo J, Cao H and Li L: Human hepatic progenitor cells
express hematopoietic cell markers CD45 and CD109. Int J Med Sci.
11:65–79. 2013. View Article : Google Scholar
|
|
23
|
Bollyky PL and Wilson SB: CD1d-restricted
T-cell subsets and dendritic cell function in autoimmunity. Immunol
Cell Biol. 82:307–314. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pfeiffer E, Kegel V, Zeilinger K,
Hengstler JG, Nüssler AK, Seehofer D and Damm G: Isolation,
characterization, and cultivation of human hepatocytes and
non-parenchymal liver cells. Exp Biol Med (Maywood). 240:645–656.
2015. View Article : Google Scholar
|
|
25
|
Werner M, Driftmann S, Kleinehr K, Kaiser
GM, Mathé Z, Treckmann JW, Paul A, Skibbe K, Timm J, Canbay A, et
al: All-in-one: Advanced preparation of human parenchymal and
non-parenchymal liver cells. PloS One. 10:e01386552015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aird WC: Phenotypic heterogeneity of the
endothelium: II. Representative vascular beds. Circ Res.
100:174–190. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nolan DJ, Ginsberg M, Israely E, Palikuqi
B, Poulos MG, James D, Ding BS, Schachterle W, Liu Y, Rosenwaks Z,
et al: Molecular signatures of tissue-specific microvascular
endothelial cell heterogeneity in organ maintenance and
regeneration. Dev Cell. 26:204–219. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Asahara T, Murohara T, Sullivan A, Silver
M, van der Zee R, Li T, Witzenbichler B, Schatteman G and Isner JM:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Duncan GS, Andrew DP, Takimoto H, Kaufman
SA, Yoshida H, Spellberg J, de la Pompa JL, Elia A, Wakeham A,
Karan-Tamir B, et al: Genetic evidence for functional redundancy of
platelet/endothelial cell adhesion molecule-1 (PECAM-1):
CD31-deficient mice reveal PECAM-1-dependent and
PECAM-1-independent functions. J Immunol. 162:3022–3030.
1999.PubMed/NCBI
|
|
30
|
Hsu W, Shu SA, Gershwin E and Lian ZX: The
current immune function of hepatic dendritic cells. Cell Mol
Immunol. 4:321–328. 2007.PubMed/NCBI
|
|
31
|
Joyee AG, Uzonna J and Yang X: Invariant
NKT cells preferentially modulate the function of CD8α+
dendritic cell subset in inducing type 1 immunity against
infection. J Immunol. 184:2095–2106. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Collin M, McGovern N and Haniffa M: Human
dendritic cell subsets. Immunology. 140:22–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dianat N, Dubois-Pot-Schneider H, Steichen
C, Desterke C, Leclerc P, Raveux A, Combettes L, Weber A, Corlu A
and Dubart-Kupperschmitt A: Generation of functional
cholangiocyte-like cells from human pluripotent stem cells and
HepaRG cells. Hepatology. 60:700–714. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Forbes S, Vig P, Poulsom R, Thomas H and
Alison M: Hepatic stem cells. J Pathol. 197:510–518. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Messner S, Agarkova I, Moritz W and Kelm
JM: Multi-cell type human liver microtissues for hepatotoxicity
testing. Arch Toxicol. 87:209–213. 2013. View Article : Google Scholar :
|
|
36
|
Schyschka L, Sánchez JJ, Wang Z, Burkhardt
B, Müller-Vieira U, Zeilinger K, Bachmann A, Nadalin S, Damm G and
Nussler AK: Hepatic 3D cultures but not 2D cultures preserve
specific transporter activity for acetaminophen-induced
hepatotoxicity. Arch Toxicol. 87:1581–1593. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kostadinova R, Boess F, Applegate D, Suter
L, Weiser T, Singer T, Naughton B and Roth A: A long-term three
dimensional liver co-culture system for improved prediction of
clinically relevant drug-induced hepatotoxicity. Toxicol Appl
Pharmacol. 268:1–16. 2013. View Article : Google Scholar : PubMed/NCBI
|