Introduction
Many chemical mutagens were proven to cause mutation
and inactivation of the cancer suppressor genes such as P53, BRCA1
and BRCA2. One of the most influential chemical compounds that
could cause a mutation in these genes are the mycotoxins (1). Mycotoxins are carcinogenic toxins that
are produced by many Aspergillus and Penicillium
species growing on food commodities (1). Among the different mycotoxins,
aflatoxin B1 has been reported as the highest carcinogenic
mycotoxin (2) and aflatoxin B1 can
penetrate the cell membrane and attach to its DNA where it causes
irreversible mutations (2). The
chemical nature of aflatoxins makes it a highly liposoluble
compound that can be absorbed from the site of exposure such as
from the gastrointestinal and respiratory tract to the blood stream
where it can move throughout the body (3). The exposure to these carcinogenic
mycotoxins can be through the ingestion of the contaminated food
that ends up in the stomach where it gets absorbed or by inhaling
the dust particles of aflatoxins B1 from the food that is
contaminated with these aflatoxins (3). Once the aflatoxins enter the cells,
they are metabolized by the action of cytochrome P450, a microsomal
enzyme to aflatoxin-8, 9-epoxide in the detoxification metabolic
pathway for aflatoxins. It is highly reactive and unstable and
requires the binding to a DNA or to the protein molecule in order
to become more stable (3). Once the
highly unstable aflatoxin-8, 9-epoxide binds to the DNA molecule
with high affinity, it forms aflatoxin-N7-guanine that cause a
guanine (G) to thymine (T) transversion mutations which will
directly affect the cell cycle by affecting the P53 gene which
carries the codes for tumor suppressor proteins, meant to inhibit
the development of tumors and cancers (3).
The role of aflatoxin B1 in causing mutation in the
humans genome was extensively studied (2), many other mycotoxins are yet to be
investigated to understand their method of action once they are in
contact with the humans genome. This is particularly crucial
because many mycotoxins have a closely related chemical composition
and similar functions inside the human body. The researchers
decided on two research questions in this review that accurately
describe the objectives of this research: What are the different
types of mycotoxins that can cause cancer? What is the role of
mycotoxins in causing cancer and what types of cancers can be
caused by mycotoxins?
The reason for this review is due to the fact that
there is an increase in mycotoxin contamination in the world and by
understanding the role of different mycotoxins in the human body
and their levels of toxicity and carcinogenicity, many new measures
could be followed and many human lives and animals alike could be
saved. In addition, there is a gap in cancer studies relating to
mycotoxins as it has been only related to liver cancer (4). Although due to the similarity in
nature and functions, mycotoxins can be related to many types of
cancer. This review will concentrate on the previously published
studies that answer both of our research questions and build on the
arguments that support these questions.
Types of mycotoxins and their natural
commodities
In an experiment done by El-Banna et al
(5) nearly 1400 Penicillium
isolates were collected from several cultures, isolated directly
from food and feed. Each isolate was identified using Pitt's
classification to investigate their mycotoxins production, and 18
different mycotoxins were isolated. Each isolate was allowed to
grow on malt extract agar medium and was incubated for the period
between one to three weeks at 25°C. The mycotoxins were then
extracted using chloroform and filtration, and the produced
mycotoxins were concentrated. Finally, the mycotoxins were
classified and characterized by mycotoxins analysis using TLC
(6–8). This experiment resulted in the
production and the identification of 18 different mycotoxins from
one or more species of Penicillium. The toxins are as
follows: Brevianamid A produced by 3 species, Citreoviridin
produced by 2 species, Citrinin (Fig.
2) produced by 3 species, Cyclopiazonic acid produced by 8
species, Fumitremorgin B produced by 1 species, Griseofulvin
produced by 6 species, luteoskyrin produced by 1 species,
Ochratoxin A produced by 2 species, Patulin produced by 4 species,
Penicillic acid produced by 2 species, Penitrem A produced by 1
species, PR-toxin produced by 1 species, Roquefortine produced by 2
species, rugulosin produced by 2 species, Verrucosidin produced by
1 species, Verruculogen produced by 1 species, Viridicarumtoxin
produced by produced by 2 species and finally, Xanthomegnin
produced by 2 species (5).
Another group of mycotoxins is produced by
Aspergillus species, and these mycotoxins are called
Aflatoxins (12). They were found
to be extremely toxic and highly cariogenic. Aflatoxins are
chemical compounds that are released by the fungi Asergillus
Flavus and Aspergillus Parasiticus as secondary
metabolites, and the different laboratory investigations confirmed
the high carcinogenicity of these chemical compounds and how they
are associated with many cancers such as liver and kidney (9–12). The
different types of aflatoxins include aflatoxin B1 and B2, produced
by Aspergillus Flavus and Aspergillus parasiticus,
Aflatoxin G1 and G2, produced by Aspergillus parasiticus
(13–16). Among these toxins, it was found the
B1 and G1 are the most toxic with the highest carcinogenic effect
on animal cells (9). Food products
that can be contaminated with aflatoxins include cereal (maize,
sorghum, pearl millet, rice, and wheat), oil seeds (groundnut,
soybean, sunflower, and cotton), spices (chilies, black pepper,
coriander, turmeric, and ginger), tree nuts (almonds, pistachio,
walnuts, and coconut) and milk (17–22).
Moreover, aflatoxins are found in the contaminated tobacco leaves
(23). Aflatoxins can also be
present in the dry soil, which can move through underground water
with rain and be ingested by humans and animals through drinking
water (24).
Another group of mycotoxins that is produced by
Aspergillus, Penicillium and Fusarium were
classified and characterized based on the frequency of their
accordance with nature and their severity of the diseases they
cause. These mycotoxins are Deoxynivalenol (Fig. 8), Fumonisins (Fig. 4), Zearalenone (Fig. 10), T-2 toxin (Fig. 7), and finally certain types of ergot
alkaloids. Found mainly on grain crops (29–31),
they were known to develop a set of diseases called Mycotoxicoses,
and these diseases include blood toxicity, food poisoning and
sometimes cancer (25–27).
Another group of mycotoxins is produced by the
Fusarium species and when culturing the Fusarium
moniliforme cultures on corn, a mycotoxin called Fumonisins was
found to be produced (28). When
analyzing fumonisins using liquid chromatography, three types of
fumonisins were found namely B1 (FB1), B2 (FB2), and B3 (FB3) and
all three types are isolated when allowing the fusarium to grow on
corn and the concentrations of fumonisins will range from 800–12800
ng total fumonisins/g (29).
In addition to the aflatoxins, another group of
mycotoxins were found to have a similar action on humans and
animals, grouped as the Ochratoxins (30–32).
The Ochratoxins are mycotoxins produced by some Aspergillus
species such as A. Ochraceus and A. Niger, whereas
some Penicillium species such as P. Verrucosum and
P. Carbonarius are also found to produce Ochratoxins
(32). Among all the types
mentioned, Ochratoxin A is the most common and it has the highest
degree of toxicity (33–35). The Ochratoxins can be found in
cereals, coffee, dried fruit, and red wine. It can be accumulated
in the meat of animals and thus, meat and meat products can be
contaminated with this toxin (36–39).
As previous literature indicates, fungal strains can
be found in many terrestrial habitats which make the availability
of different mycotoxins a very common phenomenon. Since many
different fungal strains such as Penicillium,
Aspergillus and Fusarium species grow very well on
commodities such as crops, grains and stored animal feed, the
chances of mycotoxins entering the humans or animal biological
system are very high, mostly accompanied with many health concerns
and medical complications (40).
The chemical and physical nature of
mycotoxins
Researched confirmed that the most important
mycotoxins are Citrine, Aflatoxin B1, Ergot, fumonisins akaloids,
Patulin, Trichothecene, Zearalenone and Ochratoxin A and the reason
behind their importance is the fact that they are involved in many
human and animal diseases which sometimes leads to death (41).
Aflatoxin B1
Aflatoxins are the most important mycotoxins that
are produced mainly by Aspergillus Flavus and Aspergillus
Parasiticus (42), and they are
mainly produced by the aspergillus species (43–45).
There are four types of aflatoxins named B1, B2, G1, and G2 and can
be categorized based on two characteristics: Their fluorescence
color under UV light (whether it is blue or green), and their
mobility during TLC (41). The
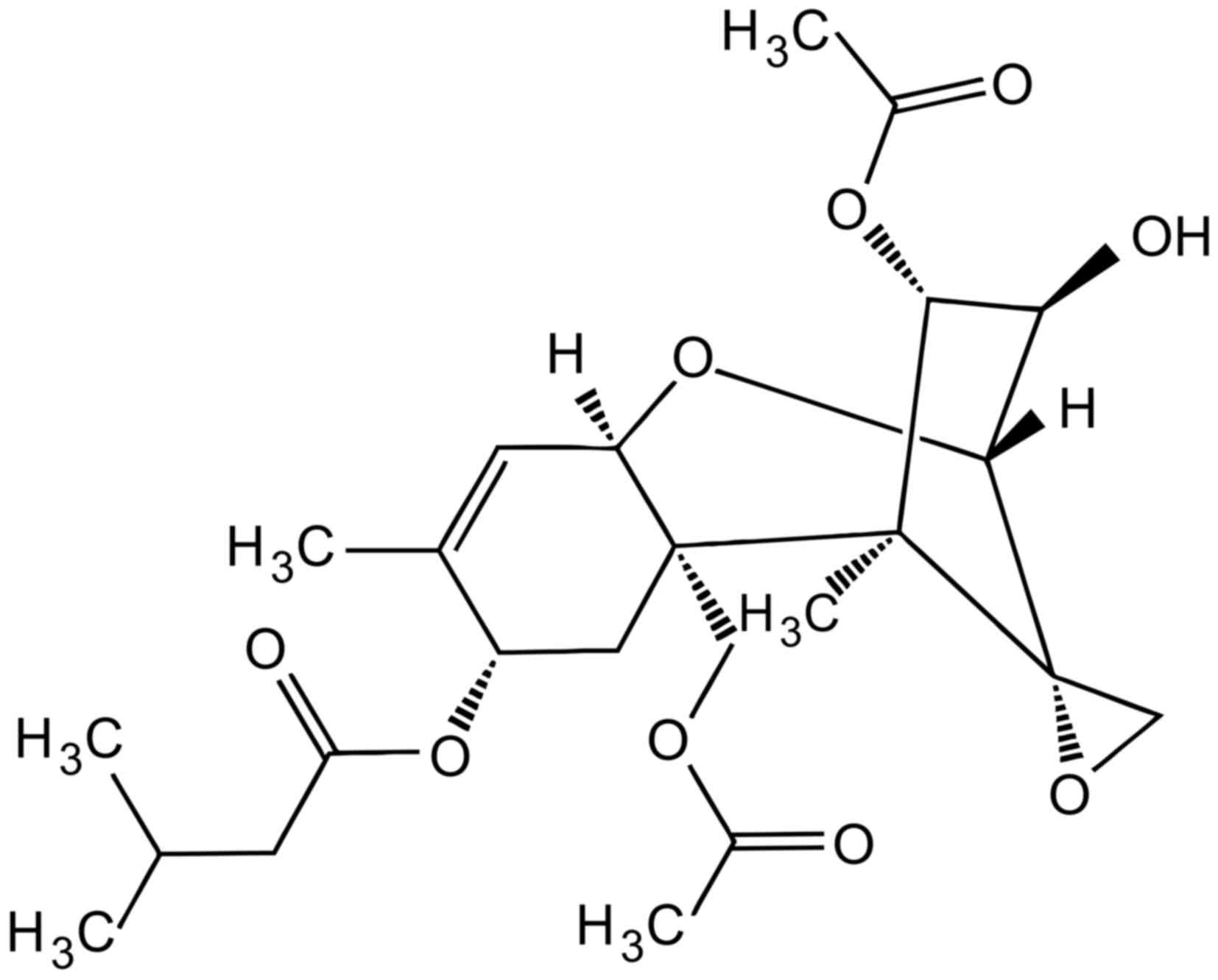
chemical formula of the aflatoxin is
C17H12O6 (Fig. 1) (46). The biosynthesis of aflatoxins starts
with the production of norsolorinic acid which is an anthraquinone
precursor joined together by the action of the enzyme II polyketide
synthase followed by 15 post-polyketide synthase steps which will
yield a series of toxigenic metabolites (47–52).
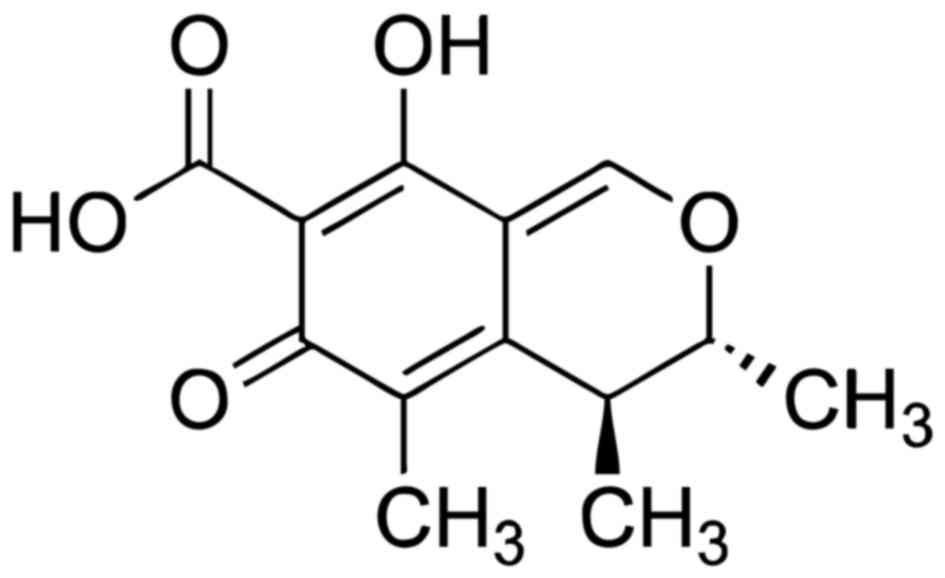
Citrinin
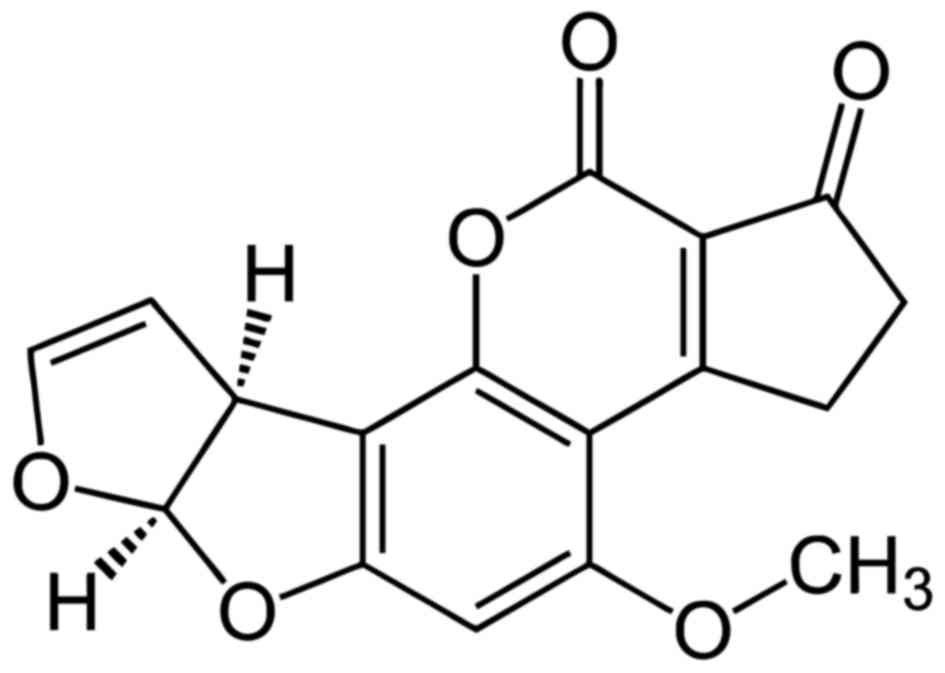
Chemically, citrine is known as
(3R,4S)-8-hydroxy-3,4,5-trimethyl-6-oxo-4,6-dihydro-3H-isochromene-7-carboxylic
acid (53) and its chemical formula
is C13H14O5 (Fig. 2) (54). It is mainly produced by
Penicillium Citrinum (55).
Citrinin is connected to the yellow rice disease in Japan and it is
a potent nephrotoxin in animals. Its acute toxicity varies with
different species (56,57).
Ergot alkaloids
The most important and fascinating Ergot alkaloids
is the Ergotamine, produced mainly by Claviceps Sclerotia and its
chemical formula is C13H14O5
(Fig. 3) (58). Ergotamine is made up of indole
alkaloids that are derived from a tetracyclic ergoline ring system
(59).
Fumonisins
The most important fumonisins are the fumonisin B1
and they are the product of condensation of alanine into
acetate-derived precursor (60).
The chemical formula of the Fumonisin B1 is
C34H59NO15 (Fig. 4) and it is basically produced by the
Fusarium species (61).
Ochratoxins
The most important ochratoxin is the Ochratoxin A,
which is produced mainly by Aspergillus Ochraceus (62) and its chemical formula is
C20H18ClNO6 (Fig. 5) (63).
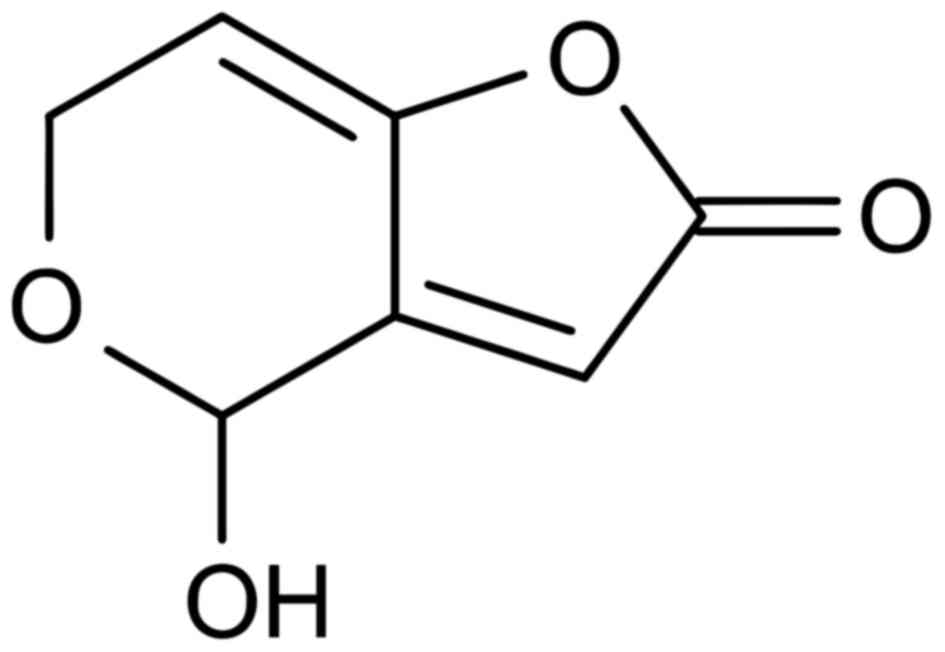
Patulin
This mycotoxin is produced by the Penicillium
pabulum, and its chemical name is 4-hydroxy-4H-furo[3,2c]
pyran-2(6H)-one (64). The chemical
formula is C7H6O4 (Fig. 6) (65). This toxin was once used as an
antibiotic until it was proven toxic in the 1960s and classified as
a mycotoxin (64).
Trichothecenes
This group of mycotoxin contains three major toxins
called T-2 toxin (Fig. 7),
Deoxynivalenol (Fig. 8) and
Satratoxin H (Fig. 9) and they are
produced mainly by Myrothecium, Phomopsis, Trichothecium,
Fusarium, Trichoderma and Stachybotrys (66–68).
Chemically, trichothecenes are made up of 12, 13-epoxytrichothene
skeletons linked up with an olefinic bond with various side chain
substitutions. Trichothecenes can be classified into group A, which
has hydrogen or ester type side chain at the C-8 position and
include T-2 toxin, and group B which has a ketone and include
fusarenon-x, nivalenol, and deoxynivalenol (69). Satratoxin is another important
Trichothecene which is a macrocyclic trichothecene produced mainly
by Stachybotrys Atra. Research has proven its toxicity towards
humans and animals (70).
Zearalenone
This mycotoxin is mainly produced by Fusarium
Graminearum and its chemical name is
6-[10-hydroxy-6-oxo-trans-1-undecenyl]-B-resorcyclic acid lactone
(71). The chemical formula of
Zearalenone is C18H22O5 (Fig. 10) (72).
Mycotoxin chemical composition makes them strong and
stable to a point where they cannot be completely destroyed during
the different food processing procedures. They can contaminate the
final food product which will eventually enter the humans
biological system (73). When the
food that is processed is mainly cereals, the contaminating
mycotoxins most likely to be found are Aflatoxins, Ochratoxin A,
Fumonisins, Deoxynivalenol and Zearalenone (74). Mycotoxins can be affected by
different food processes such as trimming, cooking, baking,
cleaning, canning, alkaline cooking, frying, roasting, flaking,
nixtamalization, sorting, milling, extrusion and brewing (75). With the increase in the temperature,
the effect on the mycotoxin will increase but due to its chemical
nature, the concentration of the mycotoxins will be reduced.
Despite this, mycotoxins cannot be completely eliminated (73). High temperature that is greater than
150°C during the extrusion was needed to reduce zearalenone greatly
and aflatoxins moderately (76). In
the same temperature during the extrusion process, there will be a
low reduction of deoxynivalenol and a high reduction of fumonisins.
Temperatures 160°C and higher with the addition of glucose was
found to reduce fumonisin to even greater levels (73).
In another experiment, the degradation of aflatoxin
was examined in different conditions and it was found that
Aflatoxin can be partially or completely degraded by several
sterilization techniques such as heat and irradiation. Besides
that, aflatoxins can be eliminated by the addition of bisulfate,
strong acids, strong bases or oxidizing agent or treatment with
strong acids or bases, oxidizing agents or bisulfite. Hydrogen
peroxide plus riboflavin denature aflatoxin in milk (77). Chemical stability of Nivalenol,
Deoxynivalenol and Zearalenone related to moisture, pH and heat was
studied by Lauren and Smith (78)
and the results showed that Nivalenol and deoxynivalenol are stable
in buffer solutions that have the range of PH 1–10, but with a PH
higher than 12, high salt concentration and a temperature of 80°C
for a long period of time, both Nivalenol and deoxynivalenol tend
to break down. In the same experiment, both Nivalenol and
deoxynivalenol are reduced to 60–100% by treating the contaminated
food with 10, 20 or 50% of the food dry weight with bicarbonate
solution and raise the temperature up to 80 or 110°C for 2–12 days.
Finally, in the case of zearalenone, the result shows that it is
highly stable and cannot be broken down even when treated with
sodium bicarbonate solution at a temperature of 110°C for more than
12 days (78).
All mycotoxins have at least one benzene ring which
gives them thermodynamic and chemical stability, and that is mainly
why they require a high temperature to desaturate and to be
utilized as less complex compounds (27,54,60,62,65,66,72,79,80).
This stability makes mycotoxins highly resistant when it comes to
decontamination and sterilization and that in turn will increase
the chances of contaminating human food and animal feed (55,72,73,81).
Finally, the chemical and physical nature of mycotoxins makes them
a very strong carcinogenic agent that can be related to different
cancer types in all countries around the world.
The action of mycotoxins on the cellular
genome and its effect on humans
The most dangerous mycotoxins are the aflatoxin B1
(Fig. 1) and Ochratoxin A (Fig. 5) for their lethal effect on humans
and animal. Aflatoxin B1 has the highest carcinogenicity among all
mycotoxins and it is capable of penetrating the cell membrane and
attach to its DNA where it makes changes to the genome in order to
become more stable (2,82,83).
Aflatoxin B1 is considered a liposoluble compound which can be
absorbed from the site of exposure and move into the blood stream
throughout the body (84). As soon
as the aflatoxins enter the cells, they get metabolized by
cytochrome P450, to aflatoxin-8, 9-epoxide which is highly reactive
and unstable and requires the binding to a DNA or to the protein
molecule in order to become more stable (85,86).
Once the highly unstable aflatoxin-8, 9-epoxide binds to the DNA
molecule with high affinity, it forms aflatoxin-N7-guanine that
causes GC to TA transversion mutations. This will directly affect
the cell cycle by affecting the P53 gene which encodes tumor
suppressor protein that inhibits the development of tumors and
cancers (87,88).
Ochratoxin (Fig. 5)
is another mycotoxin which is given a similar importance as
Aflatoxin B1 because of how it targets the kidney (89). Ochratoxin A is considered a
nephrotoxin to animals and humans and is a very strong liver toxin,
potent teratogen, carcinogenic and immune suppressant (90,91).
Ochratoxin A will mainly disturb cellular physiology in several
ways, but the primary effects of it are usually associated with the
enzymes that are involved in phenylalanine metabolism which
inhibits the enzyme involved in the synthesis of the
phenylalanine-tRNA complex (92,93).
Moreover, Ochratoxin A is capable of inhibiting mitochondrial ATP
production and is considered a strong stimulant for lipid
peroxidation (35,94).
Trichothecene mycotoxins are proven to have multiple
inhibition effects on eukaryote cells where they inhibit protein,
DNA and RNA synthesis, mitochondrial function and effects both cell
division and cell membrane of the eukaryotic cells (95). Trichothecenes are capable of
inducing a programmed cell death response within infected cells,
causing a reduction in downstream gene products, starting a
ribotoxic stress response and activating a mitogen-activated
protein kinase (96).
Deoxynivalenol mycotoxin (Fig. 8)
usually mediates an inflammatory response by controlling the
binding activities of some transcription factors which will results
in inducing cytokine gene expression (97). In animal cells, Trichothecenes are
capable of inhibiting protein synthesis, inhibiting RNA and DNA
synthesis; altering cellular membrane structure, interfere with
mitochondria functions, initiating apoptosis, activation of MAPK
and finally, activation of the cytokine (98–104).
Citrinin (Fig. 2) is
considered as one of the strongest nephrotoxins to animals, and the
levels of toxicity differ between species (56). Citrinin can act together with
Ochratoxin A, interfere with RNA synthesis and suppress it in
rodent kidneys (105). Citrinin is
known to inhibit cytokine expression and reduce the interleukin-4
in T-helper type 1 which would result in increasing the risk of
allergies in humans (106). It is
considered genotoxic at high concentrations to cultured human
lymphocytes (107). Finally,
Citrinin is considered a nephrotoxin which causes nephropathy and
impose oxidative stress, or increase the permeability of
mitochondrial membranes (108).
Ergot Alkaloids mycotoxins are a type of Citrinin, and they are
known to induce humans ergotism, low nerve fever in humans and have
a strong influence on fertility (109–111). Although certain cleaning processes
for grains has overcome the problem of ergotism, it is still
considered a significant animal problem which causes symptoms such
as abortion, suppression of lactation, hypersensitivity, ataxia and
gangrene (112,113) and smooth muscle contractions
(114). Despite its toxicity,
Ergot Alkaloids mycotoxins and its derivatives are used widely in
abortifacient and as a drug to accelerate uterine contractions
during labor (114). They are also
used for the treatment of migraine headaches, prolactin inhibitors
for the treatment of Parkinsonism and for the treatment of
cerebrovascular insufficiency (59). Unfortunately, the use of ergot
alkaloids may result in the development of human ergotism (115).
Fumonisin B1 (Fig.
4) is an important Fumonisins mycotoxin which was reported to
be a hepatotoxic and carcinogenic toxin (28,116,117). In addition, fumonisin B1 was
reported to cause apoptosis in the liver of rats (118) and there is a possibility that it
can be linked to esophageal cancer (119). FB1's main action is to disrupt
sphingolipid metabolism which induces lipid peroxidation that will
end up altering the cell membrane, causing cell death through
apoptosis. It also has the potential of inhibiting protein
synthesis and in higher concentration to inhibit DNA synthesis
(120). It was proven to have some
carcinogenic activity by stimulating 3H thymidine
incorporation (121). Fumonisin
mycotoxins are involved in the neural tube defects in animals, and
it was hypothesized that it can have the same effect on humans. A
cluster of anencephaly and spina bifida cases in Texas in the USA
is related to fumonisins mycotoxins infection in corn products
(122,123) and it was evaluated by the
International Agency for Research that fumonisins have a cancer
risk to humans. Its carcinogenicity is classified as group 2B which
means probably carcinogenic (124). Another mycotoxin that affects
humans on a cellular level is Patulin (Fig. 6) which is just as toxic at high
concentration in but there is not enough evidence for natural
poisoning, and its recorded poisonous effects are inconclusive
(125).
Trichothecene mycotoxins were proven to be extremely
potent inhibitors of eukaryotic protein synthesis, and were found
to interfere with initiation, elongation, and termination stages of
protein synthesis. They are the first mycotoxin from the
trichothecene group known to inhibit peptidyl transferase activity
during protein synthesis (126,127). The way trichothecenes inhibit
peptidyl transferase is by binding to the same ribosome-binding
site, leading to correlation with different functional groups and
subsequently affecting the protein synthesis process (128). Among all Trichothecene mycotoxins,
Deoxynivalenol is the most commonly found mycotoxin in different
grains and once they are ingested, are reported to cause nausea,
vomiting, and diarrhea with refusal for food and weight loss in
animals (129). Deoxynivalenol was
found to have a mitogen-activated protein kinase-mediated IL-8
expression in cloned human monocytes and peripheral blood
mononuclear cells where the concentration of deoxynivalenol is from
250 to 1000 ng/ml. This will induce both IL-8 mRNA and IL-8
heteronuclear RNA which is an indicator of IL-8 transcription in
human U937 monocytic cell line in a concentration-dependent manner
(130).
Other Trichothecene mycotoxins that were found to
have a great toxic effect on humans and animals are T-2 (Fig. 7) and diacetoxyscirpenol. Both were
proven to be immunosuppressive in which they reduce resistance
towards different infections and microbes (129,131). T-2 mycotoxin was proven to induce
apoptosis at 10 ng/ml concentration (132) and was reported to inhibit the
platelet aggregation and the release of dense bodies with a change
in the membrane permeability (133). On the other hand,
diacetoxyscirpenol is capable of inhibiting protein synthesis, has
high immunomodulatory effects (134) and was reported to cause alimentary
intoxication, food poisoning and induce hematological symptoms
(135). Moreover, it was proven
that T-2 and diacetoxyscirpenol are capable of causing several
dermatological, gastrointestinal, and neurologic symptoms in humans
and animals (136). Finally, T-2
and diacetoxyscirpenol are hypothesized to be associated with a
humans disease called alimentary toxic aleukia in which the
infected person will have skin inflammation, continuous vomiting,
and damage to hematopoietic tissues that gives rise to blood cells
and in advanced stages, cause central nervous system disorders,
oral cavity, and severe bleeding from the nose, mouth, and vagina
(111).
Zearalenone mycotoxins (F-2) (Fig. 10) are a unique mycotoxin that is
biologically potent but is hardly toxic and resembles
17β-estradiol, which is an ovary hormone which humans produce to
help the ovaries bind to estrogen receptors in the target cells.
Therefore, it can be classified as a non-steroidal estrogen or
mycoestroge (137) and used to
reduce the postmenopausal symptoms in women (138). The major effects of Zearalenone
are on the reproduction in females, where it affects reproductive
organs and their function, eventually leading to a medical
condition called hyperestrogenism (137). Zearalenone in its reduced form,
zearalenol, is known to increase estrogenic activity (139) but it was proven to cause disrupted
conception, abortion, and other problems in female pigs (140) and that can be hypothesized for
humans as well.
Mycotoxins and their carcinogenicity
The effects of mycotoxins on humans and animals will
vary based on the nature of the toxins. While several are
considered to be mutagenic; it was proven that Aflatoxin B1,
Ochratoxin A and Sterigmatocystin are strong carcinogens (17). In a study made to identify the
DNA-attacking ability of carcinogenic mycotoxins, six Penicillium
toxins that include patulin, Citrinin, penicillic acid
negative-luteoskyrin, positive-rugulosin, and PR-toxin, 5
aspergillus toxins including O-acetylsterigmatocystin,
sterigmatocystin, O-acetyldihydrosterigmatocystin aflatoxin B1 and
aflatoxin G1, and two fusarium toxins that include zearalenol-b and
zearalenone, all these mycotoxins were found to have destructive
ability towards genomic DNA (141). Although these 13 mycotoxins have
the ability to attack the genomic DNA, 8 of them were found to be
highly carcinogenic, including penicillic acid, citrinin, patulin,
negative-luteoskyrin, positive-rugulosin, sterigmatocystin,
aflatoxins B1, and G1 (141).
In another experiment conducted on mice, the effect
of luteoskyrin and cyclochlorotine mycotoxins was investigated, and
it was found that both toxins have hepatotoxic and
hepatocarcinogenic activities on rats (142) with a hypothetical possibility to
affect humans in a similar fashion. Fumonisin mycotoxins were found
to be carcinogenic in rats due to their action in inhibiting
ceramide synthase in vitro which in turn would enhance the
production of tumor necrosis factor α (TNFα). This leads to the
initiation of cells apoptosis (143). In addition, an experiment was
conducted using 28 mycotoxins to investigate their action against
hepatocytes of rats and mice. The results of the experiment showed
that unknown carcinogenic mycotoxins such as
5,6-dimethoxysterigmatocystin, versicolorins A and B, averufin,
xanthomegnin, luteosporin, and chrysazin, known carcinogenic
mycotoxins such as aflatoxin B1 (Fig.
1), sterigmatocystin, luteoskyrin, Ochratoxin A (Fig. 5), azaserine, mitomycin C, and
actinomycin D have the tendency to elicit DNA repair synthesis,
which suggest the possibility of these mycotoxins to be genotoxic
carcinogens (144). On the other
hand, some carcinogenic mycotoxins such as penicillic acid,
patulin, griseofulvin, and rugulosin were not found to elicit DNA
repair synthesis and suggest the possibility of these mycotoxins
being nongenotoxic carcinogens (144).
Aflatoxin B1
Aflatoxin B1 has a ubiquitous nature and is
considered a toxic metabolite, making it a serious public health
concern. Furthermore, aflatoxin B1 (Fig. 1) toxin contamination is related to
liver diseases (145). Aflatoxin
B1 is said to have an active role in causing hepatic and
extrahepatic carcinogenesis in both humans and animals by inducing
a DNA single strand break (146)
and through bio activation of Aflatoxin B1 to its epoxide
metabolite. As a result, it will interact with the DNA molecule
which will finally lead to neoplastic transformation of the cells
(147). The activation of
Aflatoxin B1 is accomplished by two approaches, chemical and
enzymatic in which the Aflatoxin B1 will be biologically active.
The chemical activation of the Aflatoxin B1 will be through the
chemical oxidation of dimethyldioxirane whereas the enzymatic
activation will be through the cytochrome P450. Combined, they are
a mixed-function oxidase that will give rise to a mixture of exo
and endo-8, 9-epoxides (148). The
exo-8, 9-epoxides will interact with the DNA molecule by attacking
the nitrogen atom at the 7 position (N7) of guanine at the C8 of
the epoxide by the Sn2 reaction. This process initiates the trans
adduct that will lead to the development of a malignant cell. They
are considered to be carcinogenic agents (149,150). On the other hand, it is not
capable of interacting with DNA molecules. Hence, it is not
considered a carcinogenic agent (148).
It was reported that almost half of the
hepatocellular carcinomas (HCC) were reported in the part of the
world that witnessed high levels of Aflatoxin B1 contamination in
food where a mutation in the 249th codon of the P53 tumor
suppressing gene (151). The
mutation caused in this codon is in the form of transversion in the
third position of this codon and will cause the insertion of serine
at the 249 in the mutant protein (152). In addition to the G-->T
transversion, Aflatoxin B1 will cause G-->T and C-->A
transversions into adjacent codons but with reduced frequency but
it was proven only as G-->T transversion at 249 codon. It turns
out that the mutant serine 249 p53 protein are responsible for the
mutation at p53 in HCC from aflatoxin B1 in the areas that were
reported to have aflatoxin B1 contamination in the food (153). This confirmed that aflatoxin B1 is
hepatocarcinogenic (154). In
another study, there was an increase of the hepatocellular
carcinoma in Mozambique and Transkei, and several parts of Asia
(155) including China (156) and Africa including Kenya (153).
Inhalation of aflatoxin B1 is thought to cause
primary lung cancer due to the activation of it to its biologically
active form 8, 9-epoxide where the rate of 8, 9-epoxide was
measured using AFB1-glutathione (AFB1-GSH) conjugate by HPLC with
aflatoxin b1 at a concentration of 124 µm. Rates of activation were
less than in the liver cells most probably due to the
CYP-containing cells that are less abundant in the lungs than in
the liver (157). In another
study, the X-ray cross-complementing group 1 protein (XRCC1) which
is involved in the repair mechanism of the single strand DNA was
investigated in the presence of aflatoxin. A substitution of
guanine to adenine was found at codon 399 and that in turn was
found to increase the risks of adenocarcinoma of the lung (158). The presence of aflatoxin B1 has
been confirmed in two cases of human lung cancer using thin-layer
chromatography and fluorescence microscopy (159,160). Finally, the expression of GSTM1-1
that is responsible for the production of cytosolic Glutathione
S-transferases helps in detoxifying several carcinogenic
electrophiles in the presence of polycyclic aromatic hydrocarbons
that serves as the main compound, yielding substrates for these
enzymes, and naturally was affected by the presence of Aflatoxin
B1, which leads to inactivation of GSTM1-1 and increased risk
factor for lung cancer (161).
Ochratoxin A
Ochratoxin A (Fig.
5) was reported to be associated with different effects on
humans and animals but their nephrocarcinogenicity was found to be
the most important effect, and that made it a major public health
concern. In an experiment conducted on male Fischer rats that were
fed Ochratoxin B1, a renal tumor was discovered within the first
six months, and the tumor incidents increased by 25%. This is
because the genes responsible for kidney injury and cell
regeneration were significantly influenced by the action of
Ochratoxin A (162). Ochratoxin A
was found to cause an alteration of gene expression that resulted
in affecting calcium homeostasis which disrupts the different
pathways regulated by the HNF4α and Nrf2 in the kidney (163). In addition, Ochratoxin A was found
to have a small influence on the expression of genes involved in
DNA synthesis and repair and ones induced as a result of DNA
damage. There was minimal to no influence at all on the genes that
are responsible for apoptosis (163,164). Ochratoxin was also found to effect
Nrf2-regulated genes in the kidney that is required for the
chemical detoxication and antioxidant defense within the cell. That
in turn affects the cell's defense mechanism while also causing an
increase in the kidney oxidative stress, which contributes to the
Ochratoxin A carcinogenic nature (165). Ochratoxin A is suggested to have a
renal carcinogenesis effect on the kidneys and its mode of action
is by the means of direct genotoxicity through covalent DNA adduct
formation. This information should be taken into consideration for
Ochratoxin A for OTA human risk assessment (166).
The alteration caused by Ochratoxin A could lead to
single cell death in kidney and stimulates cell proliferation and
an enlarged cellular nucleus which indicates blocked nuclear
division during mitosis. Exposure to Ochratoxin A could also lead
to overexpression of mitosis regulators such as mitotic protein
kinases Polo-like kinase 1, Aurora B and cyclin-dependent kinase 1,
cyclins and cyclin-dependent kinase inhibitors, topoisomerase II
and surviving (167). Ochratoxin A
(Fig. 5) is classified as a
ubiquitous nephrotoxic mycotoxin and it was reported to be
responsible for human Balkan endemic nephropathy and involved in
urinary tract tumors. For this reason, it was classified by the
Agency for Research on Cancer as a possible carcinogen belonging to
group 2B (168). Ochratoxin A has
a genotoxic activity on the cell DNA followed by oxidative
metabolism, which plays a major role in Ochratoxin mediated
carcinogenesis. It can be divided into direct modes of action where
Ochratoxin A is involved in covalent DNA adduction and indirect
modes where Ochratoxin A is involved in oxidative DNA damage
(169). Ochratoxin OTA is a
genotoxic carcinogen because of their ability to oxidase DNA
lesions and the direct DNA adducts through quinone formation
(170). Ochratoxin A was proven to
increase phosphorylation of atypical-PKC which in turn leads to a
downstream activation of ERK1 and ERK 2 and their substrate ELK1,
ELK2 and p90RSK where PKC and the MEK-ERK MAP-kinase pathways are
activated by Ochratoxin A involved in cell proliferation, cell
survival, anti-apoptotic activity and most importantly renal cancer
development (171).
A study on rodents supported the involvement of
Ochratoxin A mycotoxin in developing breast cancer through genetic
damage, and the study hypothesized the possibility of the
Ochratoxin A in causing breast cancer. The validity of that
hypothesis relates to data that show an increase in breast cancer
cases when a number of people in a population migrate from one
geographic area to another (172).
Ochratoxin A was hypothesized to be involved in testicular cancer
which is one the most common cancer affecting young men due to
adducts in testicular DNA where the consumption of Ochratoxin
during pregnancy and during young childhood would cause lesions in
the testicular DNA during puberty. These lesions will develop into
testicular cancer (173–175).
Citrinin
Citrinin (Fig. 2)
is considered as nephrotoxic mycotoxin (176), and traces of Citrinin were found
in contaminated food in areas known to have endemic nephropathy,
diseases that are associated with urinary tract cancer. It produced
micronuclei and DNA migration and had the potential of inducing
micronuclei in HepG2 (human-derived liver cell line), and can cause
DNA damage, effectively increasing the risk of cancer in humans
(177). Citrinin was proved to
induce chromosomal aberrations in both humans and rat liver
microsomes. In addition, Citrinin can also cause complex
translocations with the cellular DNA in a high frequency, and is
capable of inducing defects in chromosomal coiling (178), which may lead to the formation of
DNA adducts (179). In an
experiment performed to assess the effect of Citrinin on viability
and DNA fragmentation and chromosome aberrations in mouse bone
marrow cells and cultured Vero cells, Citrinin was found to be a
weak cytotoxic. However, it causes DNA fragmentation and chromosome
aberrations that may lead to cancer, apoptosis, and carcinogenesis
(180). Citrinin was found to
inhibit cell-free microtubule polymerization and it has an
aneuploidogenic activity. Citrinin is also a source of mitotic
arrest and CREST-positive micronuclei where it will induce the
formation of micronuclei containing whole chromosomes or chromatids
(181). Finally, some strong
evidence suggests that Citrinin is involved in the development of
Balkan endemic nephropathy and associated urinary tract tumors
(182).
Fumonisins
Fumonisin (Fig. 4)
is a potential cancer causing agent in humans (183). An experiment was conducted on 25
rats where their food was mixed with Fumonisin B1 for a period
ranged from 6 to 26 months and the result of the experiment
confirmed that Fumonisin B1 is hepatocarcinogenic and hepatotoxic
(184,185). Fumonisins are capable of
inhibiting the biosynthesis of sphingosine and the complex
sphingolipids by inhibiting the enzyme ceramide synthase and that,
in turn, will lead to the increase of free sphinganine and the free
sphingoid bases which are toxic to cells. In cultured cells that
are treated with Fumonisin B1, the accumulation of bioactive
long-chain sphingoid bases and depletion of complex sphingolipids
essential for cell growth and development lead to inhibiting the
growth, increasing cell death, and influence on mitogenicity that
would lead to the development of cancer in fibroblast cell line 3T3
(186).
Fumonisins were found correlated with incidence of
esophageal cancer in both Southern and Northern Africa and China,
neurodegenerative disease (187,188) and leukoencephalomalacia in horses
(28). It was proven that fumonisin
B1 are involved in the development of renal carcinomas in male
rats, and liver cancer in female mice which could occur in humans
as well (189). Finally, Fumonisin
B1 was found to cause inhibition in the production of ceramide
synthase in vitro which in turn enhances tumor necrosis
factor α, production and cause apoptosis (143). Finally, Fumonisins mycotoxins were
found to contribute to the development of neural tube defects among
Mexican-American woman in south Texas, and was characterized by
defects in the brain, spine, or spinal cord of fetus in the first
month of pregnancy (190).
Patulin
Patulin (Fig. 6)
and its precursor 6-methylsalicylic acid are derived from
Acetyl-CoA which makes them polyketides and carcinogenic mycotoxins
(191,192). They are capable of causing gene
mutations in different mammalian cells (193). The administration of patulin
subcutaneously twice a week for 15 months showed the development of
malignant tumor cells in the area of administration which proved
the carcinogenic effect of this mycotoxin. It can by hypothesized
in humans as well (64,194–196) although the carcinogenic effect of
patulin was not confirmed by Osswald at al (197), patulin was confirmed to be toxic
to the mice born to patulin treated mothers and deaths occurred in
both males and females. Patulin is found to be mutagenic,
carcinogenic and teratogenic and it has the tendency to induce
intestinal injuries (198) and it
has the ability to attack cellular DNA in bacteria (199) and humans (141), that can lead to the development of
tumors, and cancer.
It was proposed that the toxicity of patulin for
intestinal cells involves an inactivation of the active site of
protein tyrosine phosphatase (PTP). The protein tyrosine
phosphatase is a key regulator of intestinal epithelial barrier
function, and the active site of PTP contains a cysteine residue
(Cys215) which is important and required for phosphatase activity.
On the other hand, the sulfhydryl-reacting compounds such as
acetaldehyde decrease the transepithelial resistance (TER) through
a covalent modification of cysteine residue Cys215 of the protein
tyrosine phosphatase and this toxicity can lead to damage in the
intestinal cells that eventually may lead to intestinal and stomach
cancer (198) and this evidence
can be used to hypothesize and indicate the involvement of patulin
in the development of intestinal and stomach cancer in humans.
Patulin was found to inhibited the cell-free microtubule
polymerization, it has the ability to bind covalently to reactive
thiol groups of microtubule proteins, and deactivates the protein.
It induces the formation of micronuclei that contains the whole
chromosomes or chromatids, it also induces the formation of
micronuclei containing acentric chromosomal fragments, and that
indicates its clastogenic ability, and finally, Patulin has both
aneuploidogenic and clastogenic properties that could contribute to
the carcinogenicity of this mycotoxin in the long-term studies
which in turn could be hypothesized in humans as well (181).
Trichothecenes
T-2 (Fig. 7) are
mycotoxins that belong to Trichothecene mycotoxin and is considered
as immunosuppressive, cytotoxic and lymphocytic and carcinogenic
mycotoxin in mammalian cells (17)
although the work of Ueno et al (200) on Salmonella typhimurium suggested
that trichothecenes have failed to demonstrate mutagenicity. T-2 is
capable of inducing breaks in the DNA of lymphocytes when
administrated in vivo and in vitro and once
fibroblast cells are treated with this mycotoxin incorporation with
3H thymidine, unscheduled DNA synthesis will take place
(201). The susceptibility of
newborn and immature animals to T-2 are more compared to adults and
it causes a high degree of edema and dermal toxicities through the
direct attack of trichothecenes on the capillary vessels (202). In the presence of aflatoxin B1
(Fig. 1), the trichothecenes
nivalenol, will enhance the aflatoxin B1-induced
hepatocarcinogenesis and T-2 lack any hepatocarcinogenesis ability
and that was confirmed by an investigation in the presence of
GST-P-positive foci that acts as a marker for the presence of
hepatocarcinogenesis (203).
Although it was proposed (204)
that under the experimental conditions in both in vivo and
in vitro, no damage took place in the hepatic DNA, T-2 has
the ability to decrease responses to mitogens in human lymphocytes
in vitro (205).
The exposure to trichothecene mycotoxin can
activate apoptosis in both in vitro and in vivo
within many organs that include the lymphoid organs, the
hematopoietic tissues, with in the liver and the intestine, in the
bone marrow and finally the thymus (95,206,207) and it inhibits the protein
synthesis (206,208) and it was found to induce cellular
necrosis and karyorrhexis in the dividing tissues of the intestinal
mucosa, bone marrow, spleen, testis, and finally, the ovary
(208).
Another trichothecene mycotoxin that has the
ability to increase the frequency of inducing mutation and cancer
is Nivalenol. Nivalenol can also be involved in casing sister
chromatid exchange in Chinese hamster cells and it can case DNA
damage making it a potential genotoxic agent (205) which can be hypothesized in humans
as well. Nivalenol acute toxicity can lead to bone marrow toxicity
and Nivalenol chronic toxicity would lead to leucopenia (209). Unfortunately, there is no clear
evidence of Nivalenol carcinogenicity in experimental animals or
humans and an investigation was carried on cell lines in
vitro (205,210).
Diacetoxyscirpenol mycotoxin is another group of
trichothecene mycotoxin which is known to inhibit the proliferation
and Ig production in the mitogen-stimulated human lymphocytes
(211) and it causes esophageal
hyperplasia but is not capable of inducing cancer in rats (212) and despite its toxicity, it was
used as a chemotherapeutic agent for cancer treatment (132).
Finally, FUS-X in another group of trichothecene
mycotoxin which is capable of causing an immunosuppressive
response, carcinogenicity, cytotoxicity in experimental animals and
possibly in humans as well (208).
FUS-X is toxic to many cell types such as murine thymocytes,
lymphocytes, and gastric epithelial cells but they have a high
degree of toxicity to human hepatoblastoma cells (213). It can initiate apoptosis in mouse
thymocytes in vivo as well as in vitro (214,215) that can be hypothesized in humans
as well. Although FUS-X was confirmed to be carcinogenic in animals
(208), no clear evidence is
available on its carcinogenicity in humans (213). FUS-X is highly cytotoxic to
several type of cells (216,217). It is suggested that it has
teratogenicity or chromosomal effects, but no strong evidence is
available to support this hypothesis (213) although (214) has demonstrated that FUS-X has
genotoxic potentials when administered or exposed at low levels.
Bony et al (210) concluded
that due to the lack of strong evidence on FUS-X carcinogenicity in
both animal and man, FUS-X is not considered as a carcinogenic
agent.
Zearalenone
Zearalenone (Fig.
10) shows hormonal activity in many animal species which makes
it genotoxic, carcinogenic or both at the same time (218), and it can impose changes in the
reproductive organs and systems of many laboratory animals such as
mice, rats and different domestic animals such as horses (205). Due to this ability, it is
hypothesized that Zearalenone can be involved in causing
reproductive organ cancer in both humans and animal. Zearalenone
was found to be carcinogenic in mice and capable of causing
hepatocellular adenomas and pituitary tumors (205). More work is needed in order to
confirm its carcinogenicity in humans. Despite its toxicity and
carcinogenicity, Zearalenone is used to increase the meat
production in cattle in some countries, and it is forbidden in
other countries like some European countries (218). That is most probably due to the
lack of evidence or data that supports its toxicity and
carcinogenicity in humans and animal (219).
A high incident of esophageal cancer was reported
in areas known to have mycotoxin contamination that includes
zearalenone and the ability of this mycotoxin in inducing
hyperkeratotic papillomatous in the forestomach of a rat esophageal
squamous epithelium (220)
suggested the involvement of this mycotoxin in causing and
developing a tumor in the digestive tract (221–224). When cultured Vero cells and mouse
bone marrow were treated with zearalenone, the mycotoxin induces
the formation of micronuclei, the development of clastogenicity and
induces genotoxicity (225).
Zearalenone has a DNA-attacking ability (141).
In an experiment to demonstrate the effect of
estrogen present in the feed of animals, Zearalenone was found to
induce mammary tumor in C3HAvyfB mice and further
studies need to be conducted in order to confirm wether Zearalenone
has cancer inducing ability or carcinogenic effects on target
organs like diethylstilbestrol and the steroidal estrogens
(226). A study was conducted on
NTP mouse where the mice were given a dose of Zearalenone through
food, and the result of this study showed the ability of
Zearalenone in inducing hepatocellular adenomas in the female mice.
Zearalenone was also found to cause DNA damage in Bacillus subtilis
and it showed DNA adducts in female mice kidney and liver (227,228). Finally, it was shown that the
results of this study confirmed the genotoxicity and the ability of
zearalenone to induce hepatocellular adenomas in mice but not in
rats (227). Since there is
evidence to prove that these result can relate to humans, these
results can be used to hypothesize the involvement of Zearalenone
in developing liver, kidney and other types of cancer in
humans.
Mycotoxin research gaps and the recommended
new research areas
Many studies were conducted on the effect of
mycotoxins on the biological system and their
carcinogenicity-toxicity as mentioned before in this study but
unfortunately, many of these studies were done on animal models and
the action of mycotoxins in human biological systems were simply
hypothesized and no solid data were generated for many mycotoxins.
As mentioned earlier, food products that can be contaminated with
mycotoxins include cereal (maize, sorghum, pearl millet, rice, and
wheat), oil seeds (ground nut, soybean, sunflower, cotton), spices
(chilies, black pepper, coriander, turmeric, and ginger), tree nuts
(almonds, pistachio, walnuts, and coconut) and finally milk
(27,229–232). In addition, aflatoxins are found
in the contaminated tobacco leaves (233). Finally, aflatoxins can be present
in dry soil, which can move to the underground water with rain and
be ingested by humans and animals through the drinking water
(234). All these food sources and
food products are the basic food items for people around the world
which make their contamination with mycotoxin considered as a major
health concern for their high carcinogenic-toxic nature and
potential in both animals and humans biological system (40).
It is well known that the availability of safe and
edible food is essential for sustaining humans and animal life and
the fact that food shortage was the reason behind provoking many
wars, people started to store and preserve food on large scale for
longer time to avoid any shortage that aid in storing and
transporting food items in different forms of packages and
containers. That happened right before the Second World War and
continues to grow exponentially after that (235). As the food industry became bigger
after the Second World War (236),
packaging and distribution importance grew exponentially, and both
processes became more susceptible to different weather conditions
and the exposure to different microorganisms that may lead to food
spoilage and contamination with different microorganisms including
fungi and the mycotoxins they produce (237,238). An increase in the rates of
testicular cancer incidences across Europe was observed post Second
World War (239) and the reasons
for it was not clear. It can hypothesized that the reasons for it
can be linked to the increase in the stored food contamination with
mycotoxins that can lead to testicular cancer (173,174).
During and after the 2nd World War, there was an
increase in the incidents of lung cancer in Coastal Georgia
specifically between males working in the shipyards in a warm and
humid conditions (240). Due to
the high humidity, tobacco leaves were contaminated with
Aspergillus Flavus that produces aflatoxin B1 (23) which is a highly carcinogenic
substance that was associated with different cancer types including
lung cancer (161). No solid data
were generated to support the induction of human lung cancer by
aflatoxin B1 but it can be hypothesized that their involvement
induce lung cancer since aflatoxin B1 inactivates GSTM1-1 that is
responsible for the production of cytosolic Glutathione
S-transferases that helps in detoxifying several carcinogenic
electrophiles that cause lung cancer, hence additional lab
experiments need to be conducted to confirm this involvement.
Moreover, aflatoxin B1 was suggested to cause primary lung cancer
also through the activation of GSTM1-1 (161) and this mycotoxin can be inhaled
and cause a direct effect on the lung cells (157) and that can indicate the effect of
mycotoxin in causing lung cancer, but very limited data were
collected in this area and further studies need to be
conducted.
Another study was conducted in Norway where it was
found that the rates of breast cancer increased greatly during and
after World War II and the reasons were not conclusive, and they
linked the increase in the breast cancer rate to the change in the
people's lifestyle (241). The
fact that eating habits and food storage and transportation changed
greatly during and after World War II, the possibility of food
contamination increased greatly and fungal contamination started to
become a common phenomenon in stored food and mycotoxin
contamination could spread widely. Mycotoxins such as Aflatoxin B1
(Fig. 1) and Ochratoxin A (Fig. 5) were confirmed to suppress tumor
suppressor genes BRCA1, BRCA2 and P52 (2,172). A
hypothesis can be made to conclude the involvement of mycotoxin in
inducing breast cancer around the world, but there is a huge gap in
this area and additional laboratory work need to be established to
confirm it. In addition, HER-1, HER-2 and the c-myc oncogenes were
found to be overexpressed in many breast cancer cases (242–244) and that can also be related to the
involvement of mycotoxins since they have the ability to impose
different changes in the human genome but further studies need to
be conducted.
Patulin (Fig. 6)
causes an inactivation of the active site of protein tyrosine
phosphatase (PTP) and this protein is a key regulator of intestinal
epithelial barrier function and the active site of PTP contains a
cysteine residue (Cys215). This is important and required for
phosphatase activity and sulfhydryl-reacting compounds such as
acetaldehyde decrease the transepithelial resistance (TER) through
a covalent modification of cysteine residue Cys215 of the protein
tyrosine phosphatase. This toxicity can lead to damage in the
intestinal cells that eventually may lead to intestinal and stomach
cancer.
Some mycotoxins such as Fumonisins can cause
neurodegenerative disease (187,188). Their mode of action can be further
studied to detect if they are involved in causing brain cancer and
tumors by studying their effect on different genes in the brain.
Fumonisins were also found to be involved in the development of
neural tube defects among Mexican-American woman in south Texas
which is characterized by defects in the brain, spine, or spinal
cord of fetus in the first month of pregnancy (190) this involvement can also be used to
hypothesize the involvement of Fumonisins in causing brain cancer
in humans and further studies need to be conducted to support and
confirm this hypothesis.
Zearalenone mycotoxin (F-2) major effects will be
on the reproduction ability in females where they effects the
reproductive organs, and their functions which will eventually lead
to a medical condition called hyperestrogenism (137) and this phenomenon can be studied
further to uncover if F-2 can be involved in developing ovary
cancer. Finally, Fumonisin B1 was found to cause inhibition in the
production of ceramide synthase which will lead to enhancing tumor
necrosis factor α, production leading to apoptosis (143) and the ability of this mycotoxin to
induce apoptosis can be used to detect the involvement of this
mycotoxin in causing skin cancer.
Conclusions
Many mycotoxins have the ability of causing major
changes to the human genome which may result in the development of
cancer, and many other mycotoxins are capable of causing highly
toxic effects on the biological systems they enter. The different
mycotoxins that can cause cancer are Aflatoxin B1, Citrinin,
Fumonisins, Patulin, Trichothecene and Zearalenone. The role of
mycotoxins in causing cancer will vary with the type of mycotoxin
and it will depend on the mode of action within the biological
system.
Aflatoxin B1 (Fig.
1) can penetrate the cell membrane and attach to its DNA where
it causes irreversible mutations. As soon as aflatoxin B1 enters
the cells, they are metabolized by cytochrome P450, to aflatoxin-8,
9-epoxide which is highly reactive and unstable and requires the
binding to a DNA or to the protein molecule in order to become more
stable and it will form aflatoxin-N7-guanine that cause GC to TA
transversion mutations which will directly affect the cell cycle by
affecting the P53 gene which encodes for tumor suppressor protein
that inhibits the development of cancer.
Ochratoxin A (Fig.
5) can cause an alteration of gene expression that results in
affecting calcium homeostasis which will disrupt the different
pathways regulated by the HNF4α and Nrf2 in the kidney, influence
the expression of genes that are involved in DNA synthesis and
repair, in response to DNA damage and it has a very small influence
on the genes that are responsible for apoptosis. Ochratoxin A can
cause genotoxicity by forming a DNA adduct and single cell death.
Furthermore, Ochratoxin A can increase phosphorylation of a
typical-PKC which in turn will lead to a downstream activation of
ERK1 and ERK 2 and their substrate ELK1, ELK2 and p90RSK where PKC
and the MEK-ERK MAP-kinase pathways activated by Ochratoxin A are
involved in cell proliferation, cell survival, anti-apoptotic
activity and most importantly renal cancer development. Ochratoxin
A is suggested to have a renal carcinogenesis effect. Its mode of
action is by means of direct genotoxicity through covalent DNA
adduct formation. The effects of aflatoxin B1 and Ochratoxin A in
causing breast cancer and lung cancer are demonstrated in only a
few studies and additional studies need to be conducted in order to
confirm the course of action which will help in controlling and
reducing the rates of these cancers which are considered as the
most common cancers in the world.
Citrinin produced micronuclei and DNA migration and
it has the potential of inducing micronuclei. Citrinin would cause
DNA damage which would increase the risk of cancer in humans.
Citrinin can induce chromosomal aberrations in both human and rat
livers and it can cause complex translocations within the cellular
DNA at a high frequency and it is capable of inducing defects in
chromosomal coiling which may lead to the formation of DNA adducts.
Apart from this, it can cause DNA fragmentation and apoptosis and
finally, it can cause a mitotic arrest and CREST-positive
micronuclei where it will induce the formation of micronuclei
containing a whole chromosome or chromatids and will eventually
lead to cancer development.
Fumonisins (Fig. 4)
are capable of inhibiting the biosynthesis of sphingosine and the
complex sphingolipids by inhibiting the enzyme ceramide synthase.
That, in turn, will lead to the increase in free sphinganine and
the free sphingoid bases which are toxic in cells, and that will
inhibit the growth, increase cell death, and initiate involvement
in mitogenicity that would lead to the development of cancer.
Patulin (Fig. 6)
will cause an inactivation of the active site of protein tyrosine
phosphatase (PTP) and this protein is a key regulator of intestinal
epithelial barrier function. The active site of PTP contains a
cysteine residue (Cys215) which is important and required for
phosphatase activity and sulfhydryl-reacting compounds such as
acetaldehyde. Acetaldehyde decreases the transepithelial resistance
(TER) through a covalent modification of cysteine residue Cys215 of
the protein tyrosine phosphatase and this toxicity can lead to
damage in the intestinal cells that may eventually lead to
intestinal and stomach cancer.
T-2, a trichothecene mycotoxin is capable of
inducing breaks in the DNA of lymphocytes when administered in
vivo and in vitro once the fibroblast cells are treated
with this mycotoxin combined with 3H thymidine. In
addition, Nivalenol, a trichothecene mycotoxin is also involved in
causing chromatid exchange in cells and it can cause DNA damage and
that makes it a potential genotoxic agent. Finally, trichothecene
mycotoxin can inhibit the proliferation and Ig production in the
mitogen-stimulated human lymphocytes and it can induce apoptosis,
the migration of sister chromatids as well as cause protein
synthesis inhibition. The action of T-2 and Nivalenol on a cellular
level can be responsible for the development of tumors and
cancer.
Zearalenone (Fig.
10) shows hormonal activity in many animal species which makes
it genotoxic, carcinogenic or both at the same time, and it can
impose changes in the reproductive organs and systems of many
laboratory animals such as mice and rats and different domestic
animals such as horses. Thus, Zearalenone is capable of causing
reproductive organ cancer in both man and animal.
The cancers that can be caused by mycotoxins
include liver cancer, kidney cancer, esophageal cancer, lung
cancer, renal cancer, breast cancer, testicular cancer, stomach
cancer and reproductive organ cancer. Considering the danger caused
by mycotoxins, additional investigations need to be conducted to
confirm the ability of each mycotoxin in inducing cancer, as stated
above.
Acknowledgements
The first author would like to sincerely thank
Universiti Sains Malaysia for the USM Postgraduate Fellowship. All
authors acknowledge the Universiti Sains Malaysia for Research
University Team Grant (RUT) grant nos: 1001/CIPPT/812196 and
304/CIPPT/6313241.
Glossary
Abbreviations
Abbreviations:
|
TLC
|
thin-layer chromatography
|
|
HCC
|
hepatocellular carcinoma
|
References
|
1
|
Sulaiman MR, Chye FY, Hamid AA and Yatim
AM: The occurrence of aflatoxins in raw shelled peanut samples from
three districts of Perak, Malaysia. Electronic Journal of
Environmental and Food Chemistry. 6:2045–2052. 2007.http://eprints.ums.edu.my/3769/
|
|
2
|
Moradi S, Azari H, Anarkoli IJ,
Qasemi-Panahi B, Elhami S and Forouharmehr A: Effect of aflatoxin
B1 on BRCA1 and BRCA2 genes expression under in vitro cultured cell
line of normal Human Mammary Epithelial Cells (HMEC).
Jan;2015.https://www.researchgate.net/publication/268226306_Effect_of_aflatoxin_B1_on_BRCA1_and_BRCA2_genes_expression_under_in_vitro_cultured_cell_line_of_normal_Human_Mammary_Epithelial_Cells_HMEC
|
|
3
|
Bbosa GS, Kitya D, Lubega A, Ogwal-Okeng
J, Anokbonggo WW and Kyegombe DB: Review of the biological and
health effects of aflatoxins on body organs and body systems.
Aflatoxins - Recent Advances and Future Prospects. Razzaghi-Abyaneh
M: InTech. doi: 10.5772/51201.
|
|
4
|
Mohd-Redzwan S, Jamaluddin R, Abd-Mutalib
MS and Ahmad Z: A mini review on aflatoxin exposure in Malaysia:
Past, present and future. Front Microbiol. 4:3342013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
El-banna AA, Pitt J and Leistner L:
Production of mycotoxins by Penicillium species. Syst Appl
Microbiol. 10:42–46. 1987.doi: 10.1016/S0723-2020(87)80008–5.
View Article : Google Scholar
|
|
6
|
Frisvad JC, Filtenborg O and Thrane U:
Analysis and screening for mycotoxins and other secondary
metabolites in fungal cultures by thin-layer chromatography and
high-performance liquid chromatography. Arch Environ Contam
Toxicol. 18:331–335. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin L, Zhang J, Wang P, Wang Y and Chen J:
Thin-layer chromatography of mycotoxins and comparison with other
chromatographic methods. J Chromatogr A. 815:3–20. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
van der Gaag B, Spath S, Dietrich H,
Stigter E, Boonzaaijer G, van Osenbruggen T and Koopal K:
Biosensors and multiple mycotoxin analysis. Food Control.
14:251–254. 2003. View Article : Google Scholar
|
|
9
|
Butler WH, Greenblatt M and Lijinsky W:
Carcinogenesis in rats by aflatoxins B1, G1, and B2. Cancer Res.
29:2206–2211. 1969.PubMed/NCBI
|
|
10
|
Creppy EE: Update of survey, regulation
and toxic effects of mycotoxins in Europe. Toxicol Lett. 127:19–28.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ghoshal AK and Farber E: The induction of
liver cancer by dietary deficiency of choline and methionine
without added carcinogens. Carcinogenesis. 5:1367–1370. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kensler TW, Egner PA, Davidson NE, Roebuck
BD, Pikul A and Groopman JD: Modulation of aflatoxin metabolism,
aflatoxin-N7-guanine formation, and hepatic tumorigenesis in rats
fed ethoxyquin: Role of induction of glutathione S-transferases.
Cancer Res. 46:3924–3931. 1986.PubMed/NCBI
|
|
13
|
Davis ND, Diener UL and Eldridge DW:
Production of aflatoxins B1 and G1 by Aspergillus flavus in a
semisynthetic medium. Appl Microbiol. 14:378–380. 1966.PubMed/NCBI
|
|
14
|
Dorner JW, Cole RJ and Diener UL: The
relationship of Aspergillus flavus and Aspergillus parasiticus with
reference to production of aflatoxins and cyclopiazonic acid.
Mycopathologia. 87:13–15. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Farag RS, Daw ZY and Abo-Raya SH:
Influence of some spice essential oils on Aspergillus parasiticus
growth and production of aflatoxins in a synthetic medium. J Food
Sci. 54:74–76. 1989.doi: 10.1111/j.1365-2621.1989.tb08571.x.
View Article : Google Scholar
|
|
16
|
Mateles RI and Adye JC: Production of
aflatoxins in submerged culture. Appl Microbiol. 13:208–211.
1965.PubMed/NCBI
|
|
17
|
Chu FS: Mycotoxins: food contamination,
mechanism, carcinogenic potential and preventive measures. Mutat
Res. 259:291–306. 1991.doi: 10.1016/0165-1218(91)90124–5.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jelinek CF, Pohland AE and Wood GE:
Worldwide occurrence of mycotoxins in foods and feeds - an update.
J Assoc Off Anal Chem. 72:223–230. 1989.PubMed/NCBI
|
|
19
|
Krogh P: Mycotoxins in food. Academic
Press Ltd.; London: 1987
|
|
20
|
Wang J and Liu XM: Contamination of
aflatoxins in different kinds of foods in China. Biomed Environ
Sci. 20:483–487. 2007.PubMed/NCBI
|
|
21
|
Rustom IY: Aflatoxin in food and feed:
Occurrence, legislation and inactivation by physical methods. Food
Chem. 59:57–67. 1997. View Article : Google Scholar
|
|
22
|
Tabata S, Kamimura H, Ibe A, Hashimoto H,
Iida M, Tamura Y and Nishima T: Aflatoxin contamination in foods
and foodstuffs in Tokyo: 1986–1990. J AOAC Int. 76:32–35.
1993.PubMed/NCBI
|
|
23
|
Lane KS: Aflatoxin, tobacco, ammonia and
the p53 tumor-suppressor gene: Cancer's missing link? MedGenMed.
1:E101999.
|
|
24
|
Hill RA, Blankenship PD, Cole RJ and
Sanders TH: Effects of soil moisture and temperature on preharvest
invasion of peanuts by the Aspergillus flavus group and subsequent
aflatoxin development. Appl Environ Microbiol. 45:628–633.
1983.PubMed/NCBI
|
|
25
|
Desjardins AE: Fusarium mycotoxins:
chemistry, genetics, and biologyAmerican Phytopathological Society.
APS Press; St. Paul, MN: 2006
|
|
26
|
Placinta C, D'mello J and Macdonald A: A
review of worldwide contamination of cereal grains and animal feed
with Fusarium mycotoxins. Anim Feed Sci Technol. 78:21–37. 1999.
View Article : Google Scholar
|
|
27
|
Richard JL: Some major mycotoxins and
their mycotoxicoses - an overview. Int J Food Microbiol. 119:3–10.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gelderblom WC, Jaskiewicz K, Marasas WF,
Thiel PG, Horak RM, Vleggaar R and Kriek NP: Fumonisins - novel
mycotoxins with cancer-promoting activity produced by Fusarium
moniliforme. Appl Environ Microbiol. 54:1806–1811. 1988.PubMed/NCBI
|
|
29
|
Sydenham EW, Shephard GS, Thiel PG,
Stockenström S, Snijman PW and Van Schalkwyk DJ: Liquid
chromatographic determination of fumonisins B1, B2, and B3 in corn:
AOAC-IUPAC Collaborative Study. J AOAC Int. 79:688–696.
1996.PubMed/NCBI
|
|
30
|
Gimeno A: Thin layer chromatographic
determination of aflatoxins, ochratoxins, sterigmatocystin,
zearalenone, citrinin, T-2 toxin, diacetoxyscirpenol, penicillic
acid, patulin, and penitrem A. J Assoc Off Anal Chem. 62:579–585.
1979.PubMed/NCBI
|
|
31
|
Petzinger E and Weidenbach A: Mycotoxins
in the food chain: The role of ochratoxins. Livest Prod Sci.
76:245–250. 2002. View Article : Google Scholar
|
|
32
|
Trenk HL, Butz ME and Chu FS: Production
of ochratoxins in different cereal products by Aspergillus
ochraceus. Appl Microbiol. 21:1032–1035. 1971.PubMed/NCBI
|
|
33
|
Cavin C, Delatour T, Marin-Kuan M,
Holzhäuser D, Higgins L, Bezençon C, Guignard G, Junod S,
Richoz-Payot J, Gremaud E, et al: Reduction in antioxidant defenses
may contribute to Ochratoxin A toxicity and carcinogenicity.
Toxicol Sci. 96:30–39. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fink-Gremmels J, Jahn A and Blom MJ:
Toxicity and metabolism of Ochratoxin A. Nat Toxins. 3:214–220,
discussion 221. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rahimtula AD, Béréziat J-C,
Bussacchini-Griot V and Bartsch H: Lipid peroxidation as a possible
cause of Ochratoxin A toxicity. Biochem Pharmacol. 37:4469–4477.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Battilani P, Giorni P and Pietri A:
Epidemiology of toxin-producing fungi and Ochratoxin A occurrence
in grapeEpidemiology of Mycotoxin Producing Fungi. Springer; pp.
715–722. 2003, doi: 10.1007/978-94-017-1452-5_6. View Article : Google Scholar
|
|
37
|
Czerwiecki L, Czajkowska D and
Witkowska-Gwiazdowska A: On Ochratoxin A and fungal flora in Polish
cereals from conventional and ecological farms. Part 2: Occurrence
of Ochratoxin A and fungi in cereals in 1998. Food Addit Contam.
19:1051–1057. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Duarte SC, Pena A and Lino CM: A review on
Ochratoxin A occurrence and effects of processing of cereal and
cereal derived food products. Food Microbiol. 27:187–198. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shotwell OL, Hesseltine CW and Goulden ML:
Ochratoxin A: Occurrence as natural contaminant of a corn sample.
Appl Microbiol. 17:765–766. 1969.PubMed/NCBI
|
|
40
|
López-García R: Mycotoxin management: an
international challengeEnsuring Global Food Safety: Exploring
Global Harmonization. Boisrobert C, et al: Elsevier; pp. 209–216.
2009
|
|
41
|
Bennett JW and Klich M: Mycotoxins. Clin
Microbiol Rev. 16:497–516. 2003.doi: 10.1128/CMR.16.3.497-516.2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Squire RA: Ranking animal carcinogens: A
proposed regulatory approach. Science. 214:877–880. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Goto T, Wicklow DT and Ito Y: Aflatoxin
and cyclopiazonic acid production by a sclerotium-producing
Aspergillus tamarii strain. Appl Environ Microbiol. 62:4036–4038.
1996.PubMed/NCBI
|
|
44
|
Klich MA, Mullaney EJ, Daly CB and Cary
JW: Molecular and physiological aspects of aflatoxin and
sterigmatocystin biosynthesis by Aspergillus tamarii and A.
ochraceoroseus. Appl Microbiol Biotechnol. 53:605–609. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Peterson SW, Ito Y, Horn BW and Goto T:
Aspergillus bombycis, a new aflatoxigenic species and genetic
variation in its sibling species, A. nomius. Mycologia. 93:689–703.
2001. View Article : Google Scholar
|
|
46
|
Heathcote JG and Hibbert J: Aflatoxins:
Chemical and Biological aspects. Elsevier Scientific Publishing
Co.; Amsterdam: 1978
|
|
47
|
Bennett JW, Chang PK and Bhatnagar D: One
gene to whole pathway: The role of norsolorinic acid in aflatoxin
research. Adv Appl Microbiol. 45:1–15. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cleveland TE and Bhatnagar D: Molecular
strategies for reducing aflatoxin levels in crops before
harvestMolecular Approaches to Improving Food Quality and Safety.
Springer; pp. 205–228. 1992, doi: 10.1007/978-1-4684-8070-2_9.
View Article : Google Scholar
|
|
49
|
Hicks JK, Shimizu K and Keller NP:
Genetics and biosynthesis of aflatoxins and sterigmatocystin. The
Mycota XIAgricultural Applications. Kempken F: Springer-Verlag;
Berlin: pp. 55–69. 2002, View Article : Google Scholar
|
|
50
|
Payne GA and Brown MP: Genetics and
physiology of aflatoxin biosynthesis. Annu Rev Phytopathol.
36:329–362. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Townsend C: Progress toward a biosynthetic
rationale of the aflatoxin pathway. Pure Appl Chem. 58:227–238.
1986. View Article : Google Scholar
|
|
52
|
Trail F, Mahanti N and Linz J: Molecular
biology of aflatoxin biosynthesis. Microbiology. 141:755–765. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hackbart H, Prietto L, Primel EG,
Garda-Buffon J and Badiale-Furlong E: Simultaneous extraction and
detection of Ochratoxin A and citrinin in rice. J Braz Chem Soc.
23:103–109. 2012. View Article : Google Scholar
|
|
54
|
Aleo MD, Wyatt RD and Schnellmann RG: The
role of altered mitochondrial function in citrinin-induced toxicity
to rat renal proximal tubule suspensions. Toxicol Appl Pharmacol.
109:455–463. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Manabe M: Fermented foods and mycotoxins.
J Jpn Assoc Mycotoxicol. 51:25–29. 2001. View Article : Google Scholar
|
|
56
|
Carlton WW and Tuite J: Metabolites of P.
viridicatum toxicologyMycotoxins in Human and Animal Health.
Mehlman MA: Pathotox Publications, Inc.; Park Forest South, IL: pp.
525–555. 1977
|
|
57
|
Saito M, Enomoto M, Tatsuno T and Uraguchi
K: Yellowed Rice Toxins: Luteoskyrin and Related Compounds,
Chlorine-Containing Compounds, and Citrinin. Ajl, Samuel J Microb
Toxins. 1971.
|
|
58
|
Richard JL: Mycotoxins - an overview.
Romer Labs' Guide To Mycotoxins. 2012.
|
|
59
|
Bennett J and Bentley R: Pride and
prejudice: The story of ergot. Perspect Biol Med. 42:333–355. 1999.
View Article : Google Scholar
|
|
60
|
Sweeney MJ and Dobson AD: Molecular
biology of mycotoxin biosynthesis. FEMS Microbiol Lett.
175:149–163. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lia Y, Lou L, Cerny RL, Butchko RA,
Proctor RH, Shen Y and Du L: Tricarballylic ester formation during
biosynthesis of fumonisin mycotoxins in Fusarium verticillioides.
Mycology. 4:179–186. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Van der Merwe KJ, Steyn PS, Fourie L,
Scott DB and Theron JJ: Ochratoxin A, a toxic metabolite produced
by Aspergillus ochraceus Wilh. Nature. 205:1112–1113. 1965.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Huang B, Tao W, Shi J, Tang L and Jin J:
Determination of Ochratoxin A by polyclonal antibodies based
sensitive time-resolved fluoroimmunoassay. Arch Toxicol.
80:481–485. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ciegler A, Detroy RW and Lillehoj EB:
Patulin, penicillic acid, and other carcinogenic lactonesMicrobial
Toxins. Ciegler A, Kadis S and Ajl SJ: 6. Academic Press; New York:
pp. 409–434. 1971
|
|
65
|
Sekiguchi J and Gaucher GM: Identification
of phyllostine as an intermediate of the patulin pathway in
Penicillium urticae. Biochemistry. 17:1785–1791. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cole RJ and Cox RH: The
trichothecenesHandbook of Toxic Fungal Metabolites. Academic Press;
New York: pp. 152–263. 1981, View Article : Google Scholar
|
|
67
|
Scott PM: The natural occurrence of
trichothecenesTrichothecene mycotoxicosis: Pathophysiologic
effects. Beasley VR: 1. CRC Press; Boca Raton, FL: pp. 1–26.
1989
|
|
68
|
Ueno Y: TrichothecenesChemical, Biological
and Toxicological Aspects. Elsevier; Amsterdam: 1983
|
|
69
|
Marasas WFO, Nelson PE and Toussoun TA:
Toxigenic Fusarium speciesIdentity and mycotoxicology. University
Park: Pennsylvania State University Press; 1984
|
|
70
|
Hinkley SF and Jarvis BB: 15
Chromatographic method for Stachybotrys toxinsMycotoxin Protocols.
157. Humana Press; pp. 173–194. 2001, View Article : Google Scholar
|
|
71
|
Urry W, Wehrmeister H, Hodge E and Hidy P:
The structure of zearalenone. Tetrahedron Lett. 7:3109–3114. 1966.
View Article : Google Scholar
|
|
72
|
Cousin MA, Riley RT and Pestka JJ:
Foodborne Mycotoxins: Chemistry, Biology, Ecology, and Toxicology.
Caister Academic Press; Norfolk: 2005
|
|
73
|
Bullerman LB and Bianchini A: Stability of
mycotoxins during food processing. Int J Food Microbiol.
119:140–146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Visconti A and Pascale M: REVIEW: An
overview on Fusarium mycotoxins in the durum wheat pasta production
chain. Cereal Chem. 87:21–27. 2010. View Article : Google Scholar
|
|
75
|
Kabak B: The fate of mycotoxins during
thermal food processing. J Sci Food Agric. 89:549–554. 2009.
View Article : Google Scholar
|
|
76
|
Scott P: Effects of food processing on
mycotoxins. J Food Prot. 47:489–499. 1984. View Article : Google Scholar
|
|
77
|
Doyle M, Applebaum R, Brackett R and Marth
E: Physical, chemical and biological degradation of mycotoxins in
foods and agricultural commodities. J Food Prot. 45:964–971. 1982.
View Article : Google Scholar
|
|
78
|
Lauren DR and Smith WA: Stability of the
fusarium mycotoxins nivalenol, deoxynivalenol and zearalenone in
ground maize under typical cooking environments. Food Addit Contam.
18:1011–1016. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Gund P: Guanidine, trimethylenemethane,
and Y-delocalization. Can acyclic compounds have aromatic
stability? J Chem Educ. 49:1001972.
|
|
80
|
Silverstein RM and Bassler GC:
Spectrometric identification of organic compounds. J Chem Educ.
39:5461962. View Article : Google Scholar
|
|
81
|
Abbas HK: Aflatoxin and Food Safety. CRC
Press; 2005, doi: 10.1201/9781420028171. View Article : Google Scholar
|
|
82
|
Jarvis BB and Miller JD: Mycotoxins as
harmful indoor air contaminants. Appl Microbiol Biotechnol.
66:367–372. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Serra R, Braga A and Venâncio A:
Mycotoxin-producing and other fungi isolated from grapes for wine
production, with particular emphasis on Ochratoxin A. Res
Microbiol. 156:515–521. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Agag B: Mycotoxins in foods and feeds:
1-aflatoxins. Assiut Univ Bull Environ Res. 7:173–205. 2004.
|
|
85
|
Eaton DL and Groopman JD: Toxicology of
Aflatoxins: Human Health, Veterinary, and Agricultural
Significance. Elsevier; 2013
|
|
86
|
He XY, Tang L, Wang SL, Cai QS, Wang JS
and Hong JY: Efficient activation of aflatoxin B1 by cytochrome
P450 2A13, an enzyme predominantly expressed in human respiratory
tract. Int J Cancer. 118:2665–2671. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Baertschi SW, Raney KD, Stone MP and
Harris TM: Preparation of the 8, 9-epoxide of the mycotoxin
aflatoxin B1: The ultimate carcinogenic species. J Am Chem Soc.
110:7929–7931. 1988. View Article : Google Scholar
|
|
88
|
Yates MS, Kwak M-K, Egner PA, Groopman JD,
Bodreddigari S, Sutter TR, Baumgartner KJ, Roebuck BD, Liby KT,
Yore MM, et al: Potent protection against aflatoxin-induced
tumorigenesis through induction of Nrf2-regulated pathways by the
triterpenoid
1-[2-cyano-3-,12-dioxooleana-1,9(11)-dien-28-oyl]imidazole. Cancer
Res. 66:2488–2494. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Boorman GA, McDonald MR, Imoto S and
Persing R: Renal lesions induced by Ochratoxin A exposure in the
F344 rat. Toxicol Pathol. 20:236–245. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Chu Y-M, Jeon J-J, Yea S-J, Kim Y-H, Yun
S-H, Lee Y-W and Kim KH: Double-stranded RNA mycovirus from
Fusarium graminearum. Appl Environ Microbiol. 68:2529–2534. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kuiper-Goodman T and Scott PM: Risk
assessment of the mycotoxin Ochratoxin A. Biomed Environ Sci.
2:179–248. 1989.PubMed/NCBI
|
|
92
|
Bunge I, Heller K and Röschenthaler R:
Isolation and purification of Ochratoxin A. Zeitschrift
fürLebensmitteluntersuchung und-Forschung. 168:457–458. 1979.doi:
10.1007/BF01479260. View Article : Google Scholar
|
|
93
|
Marquardt RR and Frohlich AA: A review of
recent advances in understanding ochratoxicosis. J Anim Sci.
70:3968–3988. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Meisner H and Meisner P: Ochratoxin A, an
in vivo inhibitor of renal phosphoenolpyruvate carboxykinase. Arch
Biochem Biophys. 208:146–153. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Rocha O, Ansari K and Doohan FM: Effects
of trichothecene mycotoxins on eukaryotic cells: A review. Food
Addit Contam. 22:369–378. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Desmond OJ, Manners JM, Stephens AE,
Maclean DJ, Schenk PM, Gardiner DM, Munn AL and Kazan K: The
Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide
production, programmed cell death and defence responses in wheat.
Mol Plant Pathol. 9:435–445. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Pestka JJ: Deoxynivalenol-induced IgA
production and IgA nephropathy-aberrant mucosal immune response
with systemic repercussions. Toxicol Lett. 140–141, 287–295.
2003.
|
|
98
|
Bunner DL and Morris ER: Alteration of
multiple cell membrane functions in L-6 myoblasts by T-2 toxin: An
important mechanism of action. Toxicol Appl Pharmacol. 92:113–121.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Diamond M, Reape TJ, Rocha O, Doyle SM,
Kacprzyk J, Doohan FM and McCabe PF: The fusarium mycotoxin
deoxynivalenol can inhibit plant apoptosis-like programmed cell
death. PLoS One. 8:e695422013. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Hansen B: Inhibition of protein synthesis
by trichothecenesMycotoxins in Human and Animal Health. Meky FA,
Hardie LJ, Evans SW and Wild CP: Pathotox Publications; Park Forest
South, IL: pp. 263–273. 2001
|
|
101
|
Thompson WL and Wannemacher RW Jr:
Detection and quantitation of T-2 mycotoxin with a simplified
protein synthesis inhibition assay. Appl Environ Microbiol.
48:1176–1180. 1984.PubMed/NCBI
|
|
102
|
Wannemacher RW, Wiener SL, Sidell FR,
Takafuji ET and Franz DR: Trichothecene mycotoxins. Medical Aspects
of Chemical and Biological Warfare. 655–676. 1997.
|
|
103
|
Yang G-H, Jarvis BB, Chung Y-J and Pestka
JJ: Apoptosis induction by the satratoxins and other trichothecene
mycotoxins: Relationship to ERK p38 MAPK, and SAPK/JNK activation.
Toxicol Appl Pharmacol. 164:149–160. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhou H-R, Islam Z and Pestka JJ: Rapid,
sequential activation of mitogen-activated protein kinases and
transcription factors precedes proinflammatory cytokine mRNA
expression in spleens of mice exposed to the trichothecene
vomitoxin. Toxicol Sci. 72:130–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Sansing GA, Lillehoj EB, Detroy RW and
Miller MA: Synergistic toxic effects of citrinin, Ochratoxin A and
penicillic acid in mice. Toxicon. 14:213–220. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wichmann G, Herbarth O and Lehmann I: The
mycotoxins citrinin, gliotoxin, and patulin affect interferon-γ
rather than interleukin-4 production in human blood cells. Environ
Toxicol. 17:211–218. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Dönmez-Altuntas H, Dumlupinar G, Imamoglu
N, Hamurcu Z and Liman BC: Effects of the mycotoxin citrinin on
micronucleus formation in a cytokinesis-block genotoxicity assay in
cultured human lymphocytes. J Appl Toxicol. 27:337–341. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Flajs D and Peraica M: Toxicological
properties of citrinin. Arh Hig Rada Toksikol. 60:457–464. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Bhat R, Rai RV and Karim AA: Mycotoxins in
food and feed: present status and future concerns. Compr Rev Food
Sci Food Saf. 9:57–81. 2010.doi: 10.1111/j.1541-4337.2009.00094.x.
View Article : Google Scholar
|
|
110
|
Matossian MK: Mold poisoning: An
unrecognized English health problem, 1550–1800. Med Hist. 25:73–84.
1981. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Matossian MK: Poisons of the Past: Molds,
Epidemics, and History. Yale University Press; New Haven and
London: 1989, Arch Nat Hist. 18. pp. 145Available online: Jul 2010.
http://dx.doi.org/10.3366/anh.1991.18.1.145
|
|
112
|
Binder EM: Managing the risk of mycotoxins
in modern feed production. Anim Feed Sci Technol. 133:149–166.
2007. View Article : Google Scholar
|
|
113
|
Lorenz K and Hoseney RC: Ergot on cereal
grains. Crit Rev Food Sci Nutr. 11:311–354. 1979. View Article : Google Scholar
|
|
114
|
Riddle JM: Eve's Herbs: A History of
Contraception and Abortion in the West. Harvard University Press;
1999
|
|
115
|
Caballero-Granado FJ, Viciana P, Cordero
E, Gómez-Vera MJ, del Nozal M and López-Cortés LF: Ergotism related
to concurrent administration of ergotamine tartrate and ritonavir
in an AIDS patient. Antimicrob Agents Chemother.
41:12071997.PubMed/NCBI
|
|
116
|
Alberts JF, Gelderblom WC, Thiel PG,
Marasas WF, Van Schalkwyk DJ and Behrend Y: Effects of temperature
and incubation period on production of fumonisin B1 by Fusarium
moniliforme. Appl Environ Microbiol. 56:1729–1733. 1990.PubMed/NCBI
|
|
117
|
Gelderblom WC, Smuts CM, Abel S, Snyman
SD, Cawood ME, van der Westhuizen L and Swanevelder S: Effect of
fumonisin B1 on protein and lipid synthesis in primary rat
hepatocytes. Food Chem Toxicol. 34:361–369. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Pozzi CR, Corrêa B, Xavier JG, Direito GM,
Orsi RB and Matarazzo SV: Effects of prolonged oral administration
of fumonisin B1 and aflatoxin B1 in rats. Mycopathologia.
151:21–27. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Sydenham EW, Shephard GS, Thiel PG,
Marasas WF and Stockenstrom S: Fumonisin contamination of
commercial corn-based human foodstuffs. J Agric Food Chem.
39:2014–2018. 1991. View Article : Google Scholar
|
|
120
|
Kouadio JH, Mobio TA, Baudrimont I, Moukha
S, Dano SD and Creppy EE: Comparative study of cytotoxicity and
oxidative stress induced by deoxynivalenol, zearalenone or
fumonisin B1 in human intestinal cell line Caco-2. Toxicology.
213:56–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Tolleson WH, Melchior WB Jr, Morris SM,
McGarrity LJ, Domon OE, Muskhelishvili L, James SJ and Howard PC:
Apoptotic and anti-proliferative effects of fumonisin B1 in human
keratinocytes, fibroblasts, esophageal epithelial cells and
hepatoma cells. Carcinogenesis. 17:239–249. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Hendricks KA, Simpson JS and Larsen RD:
Neural tube defects along the Texas-Mexico border, 1993–1995. Am J
Epidemiol. 149:1119–1127. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Missmer S, Hendricks K, Suarez L, Larsen R
and Rothman I: Fumonisins and neural tube defects. Epidemiology.
11:183–184. 2000.
|
|
124
|
Rheeder JP, Marasas WF and Vismer HF:
Production of fumonisin analogs by Fusarium species. Appl Environ
Microbiol. 68:2101–2105. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Trucksess MW and Tang Y: Solid-phase
extraction method for patulin in apple juice and unfiltered apple
juice. J AOAC Int. 82:1109–1113. 1999.PubMed/NCBI
|
|
126
|
Stafford ME and McLaughlin CS:
Trichodermin, a possible inhibitor of the termination process of
protein synthesis. J Cell Physiol. 82:121–128. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wei C-M, Campbell IM, McLaughlin CS and
Vaughan MH: Letter: Binding of trichodermin to mammalian ribosomes
and its inhibition by other 12,13-epoxytrichothecenes. Mol Cell
Biochem. 3:215–219. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Bennett JW and Klich M: Mycotoxins. Clin
Microbiol Rev. 16:497–516. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Rotter BA: Invited review: Toxicology of
deoxynivalenol (vomitoxin). J Toxicol Environ Health A. 48:1–34.
1996. View Article : Google Scholar
|
|
130
|
Islam Z, Gray JS and Pestka JJ: p38
Mitogen-activated protein kinase mediates IL-8 induction by the
ribotoxin deoxynivalenol in human monocytes. Toxicol Appl
Pharmacol. 213:235–244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zain ME: Impact of mycotoxins on humans
and animals. J Saudi Chem Soc. 15:129–144. 2011. View Article : Google Scholar
|
|
132
|
Ueno Y, Umemori K, Niimi E, Tanuma S,
Nagata S, Sugamata M, Ihara T, Sekijima M, Kawai K, Ueno I, et al:
Induction of apoptosis by T-2 toxin and other natural toxins in
HL-60 human promyelotic leukemia cells. Nat Toxins. 3:129–137.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Yarom R, More R, Eldor A and Yagen B: The
effect of T-2 toxin on human platelets. Toxicol Appl Pharmacol.
73:210–217. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Sudakin DL: Trichothecenes in the
environment: Relevance to human health. Toxicol Lett. 143:97–107.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Lautraite S, Parent-Massin D, Rio B and
Hoellinger H: Comparison of toxicity induced by T-2 toxin on human
and rat granulo-monocytic progenitors with an in vitro model. Hum
Exp Toxicol. 14:672–678. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Kuhn DM and Ghannoum MA: Indoor mold,
toxigenic fungi, and Stachybotrys chartarum: Infectious disease
perspective. Clin Microbiol Rev. 16:144–172. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Kuiper-Goodman T, Scott PM and Watanabe H:
Risk assessment of the mycotoxin zearalenone. Regul Toxicol
Pharmacol. 7:253–306. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Utian WH: Comparative trial of P1496, a
new non-steroidal oestrogen analogue. BMJ. 1:579–581. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Shier WT, Shier AC, Xie W and Mirocha CJ:
Structure-activity relationships for human estrogenic activity in
zearalenone mycotoxins. Toxicon. 39:1435–1438. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Kanora A and Maes D: The role of
mycotoxins in pig reproduction: a review. Vet Med Czech.
54:565–576. 2009.
|
|
141
|
Ueno Y and Kubota K: DNA-attacking ability
of carcinogenic mycotoxins in recombination-deficient mutant cells
of Bacillus subtilis. Cancer Res. 36:445–451. 1976.PubMed/NCBI
|
|
142
|
Uraguchi K, Saito M, Noguchi Y, Takahashi
K, Enomoto M and Tatsuno T: Chronic toxicity and carcinogenicity in
mice of the purified mycotoxins, luteoskyrin and cyclochlorotine.
Food Cosmet Toxicol. 10:193–207. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Voss KA, Howard PC, Riley RT, Sharma RP,
Bucci TJ and Lorentzen RJ: Carcinogenicity and mechanism of action
of fumonisin B1: A mycotoxin produced by Fusarium moniliforme (= F.
verticillioides). Cancer Detect Prev. 26:1–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Mori H, Kawai K, Ohbayashi F, Kuniyasu T,
Yamazaki M, Hamasaki T and Williams GM: Genotoxicity of a variety
of mycotoxins in the hepatocyte primary culture/DNA repair test
using rat and mouse hepatocytes. Cancer Res. 44:2918–2923.
1984.PubMed/NCBI
|
|
145
|
Newberne PM: Mycotoxins: Toxicity,
carcinogenicity, and the influence of various nutritional
conditions. Environ Health Perspect. 9:1–32. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Gradelet S, Astorg P, Le B, on A-M, Bergès
R and Suschetet M: Modulation of aflatoxin B1 carcinogenicity,
genotoxicity and metabolism in rat liver by dietary carotenoids:
Evidence for a protective effect of CYP1A inducers. Cancer Lett.
114:221–223. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Massey TE, Stewart RK, Daniels JM and Liu
L: Biochemical and molecular aspects of mammalian susceptibility to
aflatoxin B1 carcinogenicity. Proc Soc Exp Biol Med. 208:213–227.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Iyer RS, Coles BF, Raney KD, Thier R,
Guengerich FP and Harris TM: DNA adduction by the potent carcinogen
aflatoxin B1: Mechanistic studies. J Am Chem Soc. 116:1603–1609.
1994. View Article : Google Scholar
|
|
149
|
Johnson WW and Guengerich FP: Reaction of
aflatoxin B1 exo-8,9-epoxide with DNA: Kinetic analysis of covalent
binding and DNA-induced hydrolysis. Proc Natl Acad Sci USA.
94:6121–6125. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Wilson SC, Howard PW, Forrow SM, Hartley
JA, Adams LJ, Jenkins TC, Kelland LR and Thurston DE: Design,
synthesis, and evaluation of a novel sequence-selective
epoxide-containing DNA cross-linking agent based on the
pyrrolo[2,1-c][1,4]benzodiazepine system. J Med Chem. 42:4028–4041.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Hollstein M, Sidransky D, Vogelstein B and
Harris CC: p53 mutations in human cancers. Science. 253:49–53.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Chen J, Goetchius MP, Campbell TC and
Combs GF Jr.: Effects of dietary selenium and vitamin E on hepatic
mixed-function oxidase activities and in vivo covalent binding of
aflatoxin B1 in rats. J Nutr. 112:324–331. 1982.PubMed/NCBI
|
|
153
|
Peers FG and Linsell CA: Dietary
aflatoxins and liver cancer - a population based study in Kenya. Br
J Cancer. 27:473–484. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Aguilar F, Hussain SP and Cerutti P:
Aflatoxin B1 induces the transversion of G->T in codon 249 of
the p53 tumor suppressor gene in human hepatocytes. Proc Natl Acad
Sci USA. 90:8586–8590. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Van Rensburg SJ, Cook-Mozaffari P, Van
Schalkwyk DJ, van der Watt JJ, Vincent TJ and Purchase IF:
Hepatocellular carcinoma and dietary aflatoxin in Mozambique and
Transkei. Br J Cancer. 51:713–726. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Sun G, Wang S, Hu X, Su J, Huang T, Yu J,
Tang L, Gao W and Wang JS: Fumonisin B1 contamination of home-grown
corn in high-risk areas for esophageal and liver cancer in China.
Food Addit Contam. 24:181–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Kelly JD, Eaton DL, Guengerich FP and
Coulombe RA Jr.: Aflatoxin B1 activation in human lung. Toxicol
Appl Pharmacol. 144:88–95. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Divine KK, Gilliland FD, Crowell RE,
Stidley CA, Bocklage TJ, Cook DL and Belinsky SA: The XRCC1 399
glutamine allele is a risk factor for adenocarcinoma of the lung.
Mutat Res. 461:273–278. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Dvorackova I, Stora C and Ayraud N:
Evidence of aflatoxin B1 in two cases of lung cancer in man. J
Cancer Res Clin Oncol. 100:221–224. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Zhang X, Miao X, Liang G, Hao B, Wang Y,
Tan W, Li Y, Guo Y, He F, Wei Q, et al: Polymorphisms in DNA base
excision repair genes ADPRT and XRCC1 and risk of lung cancer.
Cancer Res. 65:722–726. 2005.PubMed/NCBI
|
|
161
|
Ketterer B, Harris JM, Talaska G, Meyer
DJ, Pemble SE, Taylor JB, Lang NP and Kadlubar FF: The human
glutathione S-transferase supergene family, its polymorphism, and
its effects on susceptibility to lung cancer. Environ Health
Perspect. 98:87–94. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Marin-Kuan M, Nestler S, Verguet C,
Bezençon C, Piguet D, Mansourian R, Holzwarth J, Grigorov M,
Delatour T, Mantle P, et al: A toxicogenomics approach to identify
new plausible epigenetic mechanisms of Ochratoxin A carcinogenicity
in rat. Toxicol Sci. 89:120–134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Marin-Kuan M, Cavin C, Delatour T and
Schilter B: Ochratoxin A carcinogenicity involves a complex network
of epigenetic mechanisms. Toxicon. 52:195–202. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Dörrenhaus A, Flieger A, Golka K, Schulze
H, Albrecht M, Degen GH and Föllmann W: Induction of unscheduled
DNA synthesis in primary human urothelial cells by the mycotoxin
Ochratoxin A. Toxicol Sci. 53:271–277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Boesch-Saadatmandi C, Loboda A, Jozkowicz
A, Huebbe P, Blank R, Wolffram S, Dulak J and Rimbach G: Effect of
Ochratoxin A on redox-regulated transcription factors, antioxidant
enzymes and glutathione-S-transferase in cultured kidney tubulus
cells. Food Chem Toxicol. 46:2665–2671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Pfohl-Leszkowicz A and Manderville RA: An
update on direct genotoxicity as a molecular mechanism of
Ochratoxin A carcinogenicity. Chem Res Toxicol. 25:252–262. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Adler M, Müller K, Rached E, Dekant W and
Mally A: Modulation of key regulators of mitosis linked to
chromosomal instability is an early event in Ochratoxin A
carcinogenicity. Carcinogenesis. 30:711–719. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
González-Salgado A, Patiño B, Vázquez C
and González-Jaén MT: Discrimination of Aspergillus niger and other
Aspergillus species belonging to section Nigri by PCR assays. FEMS
Microbiol Lett. 245:353–361. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Lioi MB, Santoro A, Barbieri R, Salzano S
and Ursini MV: Ochratoxin A and zearalenone: A comparative study on
genotoxic effects and cell death induced in bovine lymphocytes.
Mutat Res. 557:19–27. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Pfohl-Leszkowicz A and Manderville RA:
Ochratoxin A: An overview on toxicity and carcinogenicity in
animals and humans. Mol Nutr Food Res. 51:61–99. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Marin-Kuan M, Nestler S, Verguet C,
Bezençon C, Piguet D, Delatour T, Mantle P, Cavin C and Schilter B:
MAPK-ERK activation in kidney of male rats chronically fed
Ochratoxin A at a dose causing a significant incidence of renal
carcinoma. Toxicol Appl Pharmacol. 224:174–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Dunnick JK, Elwell MR, Huff J and Barrett
JC: Chemically induced mammary gland cancer in the National
Toxicology Program's carcinogenesis bioassay. Carcinogenesis.
16:173–179. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Bray F, Ferlay J, Devesa SS, McGlynn KA
and Møller H: Interpreting the international trends in testicular
seminoma and nonseminoma incidence. Nat Clin Pract Urol. 3:532–543.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Jennings-Gee JE, Tozlovanu M, Manderville
R, Miller MS, Pfohl-Leszkowicz A and Schwartz GG: Ochratoxin A: In
utero exposure in mice induces adducts in testicular DNA. Toxins
(Basel). 2:1428–1444. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Schwartz GG: Hypothesis: Does Ochratoxin A
cause testicular cancer? Cancer Causes Control. 13:91–100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Vrabcheva T, Usleber E, Dietrich R and
Märtlbauer E: Co-occurrence of Ochratoxin A and citrinin in cereals
from Bulgarian villages with a history of Balkan endemic
nephropathy. J Agric Food Chem. 48:2483–2488. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Knasmüller S, Cavin C, Chakraborty A,
Darroudi F, Majer BJ, Huber WW and Ehrlich VA: Structurally related
mycotoxins Ochratoxin A ochratoxin B, and citrinin differ in their
genotoxic activities and in their mode of action in human-derived
liver (HepG2) cells: Implications for risk assessment. Nutr Cancer.
50:190–197. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Thust R and Kneist S: Activity of citrinin
metabolized by rat and human microsome fractions in clastogenicity
and SCE assays on Chinese hamster V79-E cells. Mutat Res.
67:321–330. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Ostry V, Malir F and Ruprich J: Producers
and important dietary sources of Ochratoxin A and citrinin. Toxins
(Basel). 5:1574–1586. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Bouslimi A, Bouaziz C, Ayed-Boussema I,
Hassen W and Bacha H: Individual and combined effects of Ochratoxin
A and citrinin on viability and DNA fragmentation in cultured Vero
cells and on chromosome aberrations in mice bone marrow cells.
Toxicology. 251:1–7. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Pfeiffer E, Gross K and Metzler M:
Aneuploidogenic and clastogenic potential of the mycotoxins
citrinin and patulin. Carcinogenesis. 19:1313–1318. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Castegnaro M, Chernozemsky IN, Hietanen E
and Bartsch H: Are mycotoxins risk factors for endemic nephropathy
and associated urothelial cancers? Arch Geschwulstforsch.
60:295–303. 1990.PubMed/NCBI
|
|
183
|
Fink-Gremmels J: Mycotoxins: Their
implications for human and animal health. Vet Q. 21:115–120. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Gelderblom WC, Kriek NP, Marasas WF and
Thiel PG: Toxicity and carcinogenicity of the Fusarium moniliforme
metabolite, fumonisin B1, in rats. Carcinogenesis. 12:1247–1251.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Howard PC, Eppley RM, Stack ME, Warbritton
A, Voss KA, Lorentzen RJ, Kovach RM and Bucci TJ: Fumonisin b1
carcinogenicity in a two-year feeding study using F344 rats and
B6C3F1 mice. Environ Health Perspect. 109 Suppl 2:277–282. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Riley RT, Wang E, Schroeder JJ, Smith ER,
Plattner RD, Abbas H, Yoo HS and Merrill AH Jr.: Evidence for
disruption of sphingolipid metabolism as a contributing factor in
the toxicity and carcinogenicity of fumonisins. Nat Toxins. 4:3–15.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Chu FS and Li GY: Simultaneous occurrence
of fumonisin B1 and other mycotoxins in moldy corn collected from
the People's Republic of China in regions with high incidences of
esophageal cancer. Appl Environ Microbiol. 60:847–852.
1994.PubMed/NCBI
|
|
188
|
Huang C, Dickman M, Henderson G and Jones
C: Repression of protein kinase C and stimulation of cyclic AMP
response elements by fumonisin, a fungal encoded toxin which is a
carcinogen. Cancer Res. 55:1655–1659. 1995.PubMed/NCBI
|
|
189
|
Dragan YP, Bidlack WR, Cohen SM,
Goldsworthy TL, Hard GC, Howard PC, Riley RT and Voss KA:
Implications of apoptosis for toxicity, carcinogenicity, and risk
assessment: Fumonisin B(1) as an example. Toxicol Sci. 61:6–17.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Hendricks K: Fumonisins and neural tube
defects in South Texas. Epidemiology. 10:198–200. 1999.PubMed/NCBI
|
|
191
|
Chang P-K, Cary JW, Yu J, Bhatnagar D and
Cleveland TE: The Aspergillus parasiticus polyketide synthase gene
pksA, a homolog of Aspergillus nidulans wA, is required for
aflatoxin B1 biosynthesis. Mol Gen Genet. 248:270–277. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Gaucher GM: Mycotoxins-their biosynthesis
in fungi: Patulin and related carcinogenic lactones. J Food Prot.
42:810–814. 1979. View Article : Google Scholar
|
|
193
|
Schumacher DM, Metzler M and Lehmann L:
Mutagenicity of the mycotoxin patulin in cultured Chinese hamster
V79 cells, and its modulation by intracellular glutathione. Arch
Toxicol. 79:110–121. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Stewart SG: Carcinogenesis: A broad
critique. J Clin Pathol. 21:5341968. View Article : Google Scholar
|
|
195
|
Dickens F and Jones HE: Carcinogenic
activity of a series of reactive lactones and related substances.
Br J Cancer. 15:85–100. 1961. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Dickens F and Jones HE: The carcinogenic
action of aflatoxin after its subcutaneous injection in the rat. Br
J Cancer. 17:691–698. 1963. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Osswald H, Frank HK, Komitowski D and
Winter H: Long-term testing of patulin administered orally to
Sprague-Dawley rats and Swiss mice. Food Cosmet Toxicol.
16:243–247. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Mahfoud R, Maresca M, Garmy N and Fantini
J: The mycotoxin patulin alters the barrier function of the
intestinal epithelium: Mechanism of action of the toxin and
protective effects of glutathione. Toxicol Appl Pharmacol.
181:209–218. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Lee KS and Röschenthaler RJ: DNA-damaging
activity of patulin in Escherichia coli. Appl Environ Microbiol.
52:1046–1054. 1986.PubMed/NCBI
|
|
200
|
Ueno Y, Kubota K, Ito T and Nakamura Y:
Mutagenicity of carcinogenic mycotoxins in Salmonella typhimurium.
Cancer Res. 38:536–542. 1978.PubMed/NCBI
|
|
201
|
Schoental R: Trichothecenes, zearalenone,
and other carcinogenic metabolites of Fusarium and related
microfungi. Adv Cancer Res. 45:217–290. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Ueno Y: Toxicological features of T-2
toxin and related trichothecenes. Fundam Appl Toxicol. 4:S124–S132.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Ueno Y, Yabe T, Hashimoto H, Sekijima M,
Masuda T, Kim DJ, Hasegawa R and Ito N: Enhancement of
GST-P-positive liver cell foci development by nivalenol, a
trichothecene mycotoxin. Carcinogenesis. 13:787–791. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Lafarge-Frayssinet C, Decloitre F, Mousset
S, Martin M and Frayssinet C: Induction of DNA single-strand breaks
by T2 toxin, a trichothecene metabolite of Fusarium: Effect on
lymphoid organs and liver. Mutat Res Genet Toxicol. 88:115–123.
1981. View Article : Google Scholar
|
|
205
|
Schothorst RC and van Egmond HP: Report
from SCOOP task 3.2.10 ‘collection of occurrence data of Fusarium
toxins in food and assessment of dietary intake by the population
of EU member states’. Subtask: trichothecenes. Subtask:
Trichothecenes. Toxicol Lett. 153:133–143. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Eriksen GS and Pettersson H: Toxicological
evaluation of trichothecenes in animal feed. Anim Feed Sci Technol.
114:205–239. 2004. View Article : Google Scholar
|
|
207
|
Pestka JJ: Deoxynivalenol: Toxicity,
mechanisms and animal health risks. Anim Feed Sci Technol.
137:283–298. 2007. View Article : Google Scholar
|
|
208
|
Yazar S and Omurtag GZ: Fumonisins,
trichothecenes and zearalenone in cereals. Int J Mol Sci.
9:2062–2090. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Ryu J-C, Ohtsubo K, Izumiyama N, Nakamura
K, Tanaka T, Yamamura H and Ueno Y: The acute and chronic
toxicities of nivalenol in mice. Fundam Appl Toxicol. 11:38–47.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Bony S, Olivier-Loiseau L, Carcelen M and
Devaux A: Genotoxic potential associated with low levels of the
Fusarium mycotoxins nivalenol and fusarenon X in a human intestinal
cell line. Toxicol In Vitro. 21:457–465. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Thuvander A, Wikman C and Gadhasson I: In
vitro exposure of human lymphocytes to trichothecenes: Individual
variation in sensitivity and effects of combined exposure on
lymphocyte function. Food Chem Toxicol. 37:639–648. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Craddock VM, Hill RJ and Henderson AR:
Stimulation of DNA replication in rat esophagus and stomach by the
trichothecene mycotoxin diacetoxyscirpenol. Cancer Lett.
38:199–208. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Wijnands LM and van Leusden FM: An
overview of adverse health effects caused by mycotoxins and
bioassays for their detection. Research for man and environment.
RIVM report 257852,004 2000. http://rivm.openrepository.com/rivm/bitstream/10029/9410/1/257852004.pdf
|
|
214
|
Bony S, Olivier-Loiseau L, Carcelen M and
Devaux A: Genotoxic potential associated with low levels of the
Fusarium mycotoxins nivalenol and fusarenon X in a human intestinal
cell line. Toxicol In Vitro. 21:457–465. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Miura K, Aminova L and Murayama Y:
Fusarenon-X induced apoptosis in HL-60 cells depends on caspase
activation and cytochrome c release. Toxicology. 172:103–112. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Bondy GS, McCormick SP, Beremand MN and
Pestka JJ: Murine lymphocyte proliferation impaired by substituted
neosolaniols and calonectrins - Fusarium metabolites associated
with trichothecene biosynthesis. Toxicon. 29:1107–1113. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Carter CJ and Cannon M: Inhibition of
eukaryotic ribosomal function by the sesquiterpenoid antibiotic
fusarenon-X. Eur J Biochem. 84:103–111. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Moss MO: Mycotoxin review-2. Fusarium.
Mycologist. 16:158–161. 2002. View Article : Google Scholar
|
|
219
|
Yazar S and Omurtag GZ: Fumonisins,
trichothecenes and zearalenone in cereals. Int J Mol Sci.
9:2062–2090. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Marasas WF, van Rensburg SJ and Mirocha
CJ: Incidence of Fusarium species and the mycotoxins,
deoxynivalenol and zearalenone, in corn produced in esophageal
cancer areas in Transkei. J Agric Food Chem. 27:1108–1112. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Richard JL: Some major mycotoxins and
their mycotoxicoses - An overview. Int J Food Microbiol. 119:3–10.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Schoental R and Cavanagh JB: Mechanisms
involved in the ‘dying-back’ process - an hypothesis implicating
coenzymes. Neuropathol Appl Neurobiol. 3:145–157. 1977. View Article : Google Scholar
|
|
223
|
Schoental R and Joffe AZ: Lesions induced
in rodents by extracts from cultures of Fusarium poae and F.
sporotrichioides. J Pathol. 112:37–42. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Schoental R, Joffe AZ and Yagen B:
Irreversible depigmentation of dark mouse hair by T-2 toxin (a
metabolite of Fusarium sporotrichioides) and by calcium
pantothenate. Experientia. 34:763–764. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Ouanes Z, Abid S, Ayed I, Anane R, Mobio
T, Creppy EE and Bacha H: Induction of micronuclei by Zearalenone
in vero monkey kidney cells and in bone marrow cells of mice:
Protective effect of Vitamin E. Mutat Res. 538:63–70. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Schoental R: Letter: Role of
podophyllotoxin in the bedding and dietary zearalenone on incidence
of spontaneous tumors in laboratory animals. Cancer Res.
34:2419–2420. 1974.PubMed/NCBI
|
|
227
|
Pfohl-Leszkowicz A, Chekir-Ghedira L and
Bacha H: Genotoxicity of zearalenone, an estrogenic mycotoxin: DNA
adduct formation in female mouse tissues. Carcinogenesis.
16:2315–2320. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Program NT: National Toxicology Program:
Carcinogenesis Bioassay of Zearalenone (CAS No. 17924–92-4) in
F344/N Rats and B6C3F1 Mice (Feed Study). Natl Toxicol Program Tech
Rep Ser. 235:1–155. 1982.PubMed/NCBI
|
|
229
|
Cho S-H, Lee C-H, Jang M-R, Son Y-W, Lee
S-M, Choi I-S, Kim S and Kim D: Aflatoxins contamination in spices
and processed spice products commercialized in Korea. Food Chem.
107:1283–1288. 2008. View Article : Google Scholar
|
|
230
|
Samson RA, van Reenen-Hoekstra ES and Van
Oorschot CAN: Introduction to food-borne fungiCentraalbureau voor
Schimmelcultures. Institute of the Royal Netherlands Academy of
Arts and Sciences; 1981
|
|
231
|
Jürgen R: Mycotoxins in foodstuffs. Appl
Microbiol Biotechnol. 1:183–190. 1975.
|
|
232
|
van Egmond HP, Schothorst RC and Jonker
MA: Regulations relating to mycotoxins in food: Perspectives in a
global and European context. Anal Bioanal Chem. 389:147–157. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Binder EM: Managing the risk of mycotoxins
in modern feed production. Anim Feed Sci Technol. 133:149–166.
2007. View Article : Google Scholar
|
|
234
|
Paterson RRM and Lima N: How will climate
change affect mycotoxins in food? Food Res Int. 43:1902–1914. 2010.
View Article : Google Scholar
|
|
235
|
Beckeman M and Skjöldebrand C:
Clusters/networks promote food innovations. J Food Eng.
79:1418–1425. 2007. View Article : Google Scholar
|
|
236
|
Wansink B: Changing eating habits on the
home front: Lost lessons from World War II research. J Public
Policy Mark. 21:90–99. 2002. View Article : Google Scholar
|
|
237
|
Kabak B, Dobson AD and Var I: Strategies
to prevent mycotoxin contamination of food and animal feed: A
review. Crit Rev Food Sci Nutr. 46:593–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Magan N and Aldred D: Post-harvest control
strategies: Minimizing mycotoxins in the food chain. Int J Food
Microbiol. 119:131–139. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Bergström R, Adami H-O, Möhner M, Zatonski
W, Storm H, Ekbom A, Tretli S, Teppo L, Akre O and Hakulinen T:
Increase in testicular cancer incidence in six European countries:
A birth cohort phenomenon. J Natl Cancer Inst. 88:727–733. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Blot WJ, Harrington JM, Toledo A, Hoover
R, Heath CW Jr and JF Jr Fraumeni: Lung cancer after employment in
shipyards during World War II. N Engl J Med. 299:620–624. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Tretli S and Gaard M: Lifestyle changes
during adolescence and risk of breast cancer: An ecologic study of
the effect of World War II in Norway. Cancer Causes Control.
7:507–512. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Birner P, Oberhuber G, Stani J, Reithofer
C, Samonigg H, Hausmaninger H, Kubista E, Kwasny W,
Kandioler-Eckersberger D, Gnant M, et al: Austrian Breast &
Colorectal Cancer Study Group: Evaluation of the United States Food
and Drug Administration-approved scoring and test system of HER-2
protein expression in breast cancer. Clin Cancer Res. 7:1669–1675.
2001.PubMed/NCBI
|
|
243
|
Lu SH, Hsieh LL, Luo FC and Weinstein IB:
Amplification of the EGF receptor and c-myc genes in human
esophageal cancers. Int J Cancer. 42:502–505. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
244
|
Ross JS, Slodkowska EA, Symmans WF,
Pusztai L, Ravdin PM and Hortobagyi GN: The HER-2 receptor and
breast cancer: Ten years of targeted anti-HER-2 therapy and
personalized medicine. Oncologist. 14:320–368. 2009. View Article : Google Scholar : PubMed/NCBI
|