Introduction
Gastric cancers continue to pose a significant
burden of morbidity and mortality globally (1). It is estimated that gastric cancer is
the fifth most common cancer worldwide, and the third leading cause
of cancer-related death (2). While
important advances have been made in the early detection and
treatment of gastric cancers, the majority of patients continue to
present with clinical evidence of regional or distant disease
(3). While treatment for gastric
cancers has traditionally involved combinations of aggressive
surgical resection, chemotherapy, and radiotherapy, the percentage
of patients achieving 5-year survival remains under 20% (4). Moreover, the detection rate of early
gastric cancers is low due to the lack of specific symptoms of
early gastric cancer, and therefore, most patients (>70%)
develop advanced-stage disease (5).
Some patients even lose the opportunity to undergo surgical
resection. Metastatic potential may also exist in advanced gastric
cancer, thus the overall prognosis is poor. Thereby, due to the
intractable nature of these diseases, it is not surprising that
attention has turned towards the mechanisms underlying cancer cell
survival and migration.
Mitophagy, is an essential housekeeping process that
is required to maintain tumor homeostasis. Studies suggest that
mitophagy is important for eliminating impaired mitochondria both
under baseline conditions and in response to stress. Mitophagy
activation may reduce mitochondrial oxidative stress injury,
alleviate mitochondrial calcium overload, block mitochondrial
pro-apoptotic factor leakage and promote cellular ATP generation
(6–8). These data hint that mitophagy is vital
for cellular homeostasis. However, research has linked impaired
mitophagy to cancer suppression in hepatocellular carcinoma, renal
carcinoma, cervical cancer and colorectal cancer (9–11). As
for gastric cancers, little evidence is available concerning the
role of mitophagy in gastric cancer progression involving cellular
migration and survival. Therefore, it is necessary to establish the
regulatory role of mitophagy in the biological function of gastric
cancers.
Ample evidence has focused on the anti-apoptotic
effect of mitophagy on cancer survival. However, there is little
research evidence to suggest whether mitophagy has the ability to
manipulate cancer migration. Cellular migration involves drastic
structural changes, a process that demands high levels of energy
and fully functional mitochondria whose quality and quantity are
balanced by mitophagy (12).
Moreover, several studies have proposed the direct regulatory
effect of mitophagy on cellular migration via stress fibers such as
F-actin and tubulin (13).
Moreover, excessive reactive oxygen species (ROS) or oxidative
stress have been found to influence F-actin homeostasis in cancers
(14). This information indicates
the possible relationship between mitophagy and F-actin-based
migration.
Several factors have been found to be related to
activation of mitophagy. In response to hypoxic stress, FUNDC1 is
dephosphorylated and mediates mitophagy activation, which provides
a survival advantage for cardiomyocytes (15). Moreover, under high glucose attack,
Bnip3 is activated and offers beneficial actions on β-cells
(16). Recently, mitofusin 2 (Mfn2)
is a novel controller for mitophagy activation (17). In damaged mitochondria with lower
mitochondrial membrane potential, Mfn2 is phosphorylated at Thr-111
and Ser-442 and selectively activates mitophagy (18), resulting in the removal of bad
mitochondria by lysosomes. This process could be enhanced by JNK
pathways (19). In contrast, loss
of Mfn2 is associated with enhanced mitochondrial dysfunction. For
example, Mfn2 deficiency in the heart leads to cardiomyopathy
(20). The age-induced reduction in
Mfn2 levels is involved in musculoskeletal disorders (17). However, whether Mfn2 is involved in
gastric cancer cell survival and migration remains unknown.
Yes-associated protein (Yap) is overexpressed and
has an oncogenic role in several types of tumors including gastric
cancer (21). Yap inhibition by
siRNA inhibits gastric carcinoma growth and tumor metastasis,
suggesting the vital role of Yap in cancer cell migration and
survival (22). Moreover, recent
studies have indicated that the Hippo pathway controls the
mitochondrial function via Bcl-xL (23), demonstrating the possible
relationship between Yap and mitochondria. Whether mitophagy is
signaled by Yap and contributes to cancer progression is
incompletely understood. In the present study, we explored the role
of Yap in gastric cancer migration and survival, particularly
focusing on Mfn2-mediated mitophagy.
Materials and methods
Ethics statement
The present study was conducted in accordance with
the Declaration of Helsinki and the guidelines of the Ethics
Committee of Shanghai Jiao Tong University Institutional Animal
Care and Use Committee. All experimental protocols were approved by
the Ethics Committee of Shanghai Jiao Tong University Institutional
Animal Care and Use Committee. The animals were cared for in
accordance with the Guide to the Care and Use of Experimental
Animals (Vol. 1, 2nd edition, 1993, and Vol. 2, 1984, available
from the Canadian Council on Animal Care, Constitution Square,
Tower 2, Suite 315, 350 Albert Street, Ottawa, ON K1R 1B1, Canada,
or on their Web site at www.ccac.ca), or
the Guide for the Care and Use of Laboratory Animals (1996,
published by National Academy Press, 2101 Constitution Ave. NW,
Washington, DC 20055, USA).
Cell culture
The human normal gastric mucosal cell line GES-1 and
human gastric cancer cell line AGS were all purchased from the
American Type Culture Collection (ATCC) (Manassas, VA, USA)
(24). Next, the cell lines were
incubated in the Roswell Park Memorial Institute (RPMI)-1640 medium
containing 10% fetal bovine serum (FBS) in an incubator with 5%
CO2 at 37°C.
ROS detection and NAO staining
To detect cellular ROS production, DCFH-DA (Beyotime
Institute of Biotechnology, Jiangsu, China) was used. In brief,
cells were incubated in FBS-free DMEM containing DCFH-DA (10 µM)
for 30 min (25). Then, the cells
were washed with PBS. Flow cytometric analyses were performed using
a BD FACSCalibur™ flow cytometer (BD Biosciences, San Jose, CA,
USA).
To measure the oxidation of cardiolipin,
10-N-nonyl acridine orange (NAO; 2 mmol/l; Molecular Probes,
Eugene, OR, USA) was used. In normal cells, NAO interacts with
non-oxidized cardiolipin and produces a characteristic green
fluorescence (26). However, after
cardiolipin is oxidized, NAO cannot bind to it. We evaluate the
fluorescence intensity to reflect the cardiolipin oxidation.
Image-Pro Plus 6.0; Media Cybernetics, Rockville, MD, USA) was used
to obtain the mean densities of the region of interest (27), which was normalized to that of the
control group.
Transwell assay
Cells in all groups were digested and added into a
24-well Transwell plate where the chambers were then transferred in
a manner that may avoid the formation of bubbles. A total of 0.5 ml
cell suspension was added into the chambers and they were then
incubated in an incubator for 24 h (28). Subsequently, the extra liquids in
the upper and lower chambers were discarded and cells in the upper
chamber above the Transwell membrane was carefully wiped off using
a cotton swab. Cells transferred in the lower chambers were fixed
using pre-cooled formalin for 30 min, stained using 1% crystal
violet for 10 min and washed in running water until they were
totally clean (29). Cells were
dried naturally and observed under the microscope. Images were
captured and observation results were recorded.
Scratch assay
When cell attachment was observed at the plate
bottom, a sterile spearhead was used to slightly draw a line in the
well, ensuring that the widths of all scratches were the same.
Images were captured and the cover of 6-well plate was marked
properly to guarantee the same observation field (30). After 24 h, an Olympus inverted
microscope was employed for observation of 6 fixed fields of
vision.
Immunofluorescence
The cells were washed in PBS and permeabilized for
10 min at 4°C in a solution of 0.1% Triton X-100 and 0.1% sodium
citrate in PBS. Cells were washed 3 times in PBS and incubated
overnight at 4°C with the primary antibody (30). The primary antibodies used in the
present study were as follows: Tomm20 (1:1,000, #ab78547), LAMP1
(1:1,000, #ab24170), F-actin (1:1,000, #ab205) and cyt-c (1:1,000,
#ab133504) (all from Abcam, Cambridge, MA, USA), Sirt1 (1:500,
#2310), Mfn2 (1:1,000, #11925) (both from Cell Signaling
Technology, Inc., Danvers, MA, USA).
Immunoblotting
Immunoblotting was carried out as described
previously (31,32). Briefly, the cells were washed with
PBS and lysed in RIPA buffer containing 1.0% Triton X-100, 0.5%
sodium deoxycholate, 0.1% SDS, 150 mM NaCl, 50 mM Tris-HCl (pH
8.0), 10 mg/ml pepstatin A, 10 mg/ml leupeptin and 1 mM
phenylmethylsulfonyl fluoride (PMSF). After centrifugation for 10
min, supernatant with 20 mg protein for each sample was subjected
to immunoblotting analysis (33).
Primary antibodies used for immunoblotting were as follows:
pro-caspase-3 (1:1,000; #9662), cleaved caspase-3 (1:1,000; #9664;
both from Cell Signaling Technology), Bad (1:2,000; #ab90435;
Abcam), Bax (1:2,000; #5023; Cell Signaling Technology), caspase-9
(1:1,000; #ab32539), Bnip3 (1:1,000; #ab109362; both from Abcam),
LC3II (1:1,000; #3868; Cell Signaling Technology), p62 (1:1,000;
#ab56416; Abcam), Beclin1 (1:1,000; #3495), Atg5 (1:1,000; #12994),
Sirt1 (1:1,000; #2310), Mfn2 (1:1,000; #11925), FAK (1:1,000;
#3285), integrin β1 (1:1,000; #34971), Talin-1 (1:1,000; #4021)
(all from Cell Signaling Technology).
RNA isolation and quantitative RT-PCR
(qPCR)
Total RNA was isolated from cells using TRIzol
reagent (Invitrogen, Carlsbad, CA, USA). RNA (1 µg) from each
sample was reversely transcribed into cDNA using a Reverse
Transcription kit (Eurogentec, Fremont, CA, USA). qPCR was
performed with primers and matched probes from the Universal
Fluorescence-labeled Probe Library (Roche, Basel, Switzerland)
(34). The primers used in the
present study were as follows: FAK forward primer,
5′-GACTGGCCCAGTGTTCTTCGCTTC and reverse primer,
5′-GCTTCTGACAGAAGGAAAGCCAAC; integrin β1 forward primer,
5′-TCAAACAACTTCATGGTCCCAGTGCTC and reverse primer,
5′-TAAACCTGACTGGTTCGGGGGATTTCT; Talin-1 forward primer,
5′-CGACATGACCAAGTGGGAGCAGAAGTA and reverse primer,
5′-TGGAGCACCTTTAACCTGCTTTCCATC.
JC-1 staining and ATP detection
The mitochondrial potential was assessed using the
probe JC-1, a sensitive fluorescent dye used to detect changes in
mitochondrial potential (35).
Briefly, after treatment, cells were incubated with 10 mg/ml JC-1
for 10 min at 37°C in the dark and monitored with a fluorescence
microscope. Red-orange fluorescence was attributable to a
potential-dependent aggregation in the mitochondria. Green
fluorescence, reflecting the monomeric form of JC-1, appeared in
the cytosol after mitochondrial membrane depolarization.
The level of ATP in cells was determined using the
ATP Bioluminescence Assay kit. Briefly, harvested cultured cells
were lysed with a lysis buffer, followed by centrifugation for 2
min at 4°C. Finally, the level of ATP was determined by mixing 50
µl of the supernatant with 50 µl of luciferase reagent (36), which catalyzed the light production
from ATP and luciferin. The emitted light was linearly related to
the ATP concentration and measured using a microplate
luminometer.
MTT and TUNEL assay
MTT experiments were performed in 96-well plates.
The cells were applied to the scaffold at a density of
104 cells/well. After 2 or 3 days of culture, the
samples were washed 3 times with PBS, and then 50 µl of MTT was
added to each well (37). The
samples were then incubated for 4 h in a humid atmosphere. The MTT
solution was removed and 200 µl DMSO was added to each sample and
incubated for 10 min. After the addition of Sorensen's buffer, the
absorbance was determined at a wavelength of 570 nm.
A TUNEL assay to detect DNA fragmentation in cell
nuclei (a marker for apoptosis in testicular tissue), was performed
using an In Situ Cell Death Detection kit (Roche, Mannheim,
Germany), according to the manufacturer's protocol. DAPI was used
to label the nuclei. The results are expressed as apoptotic
cells/tubule cross-section (38).
Small RNA interference assay
To evaluate the functional role of Yap, siRNA was
used to reduce its expression. For Yap, the sense strand siRNA,
5′-GCGACATTCAGGGUGACUAUU-3′ and antisense strand siRNA,
3′-TCGCUGUUCCTCCCACUGAUAAU-5′ were used (39). Briefly, cultured cells were washed
with Opti-Minimal Essential Medium without serum or antibiotics and
seeded in 6-well plates to 30–40% confluence. The transfection
reagent and siRNA were diluted separately in serum-free media,
mixed and incubated for 10 min at room temperature to form the
siRNA/lipid complex. This complex was then added to each well at a
final concentration of 70 nM/well of siRNA. At 48 h after
transfection, cells were collected to determine Yap protein
expression levels by western blot analysis (40).
Statistical analysis
Data analysis was conducted using SPSS 19.0
statistical software (SPSS, Inc., Chicago, IL, USA). Measurement
data are presented as mean ± standard deviation (SD). The one-way
analysis of variance (ANOVA) was conducted to analyze comparisons
among multi-groups. P<0.05 was assigned as indicative of
statistical significance.
Results
Yap is upregulated in gastric cancer
and activates mitophagy
To determine the expression of Yap in gastric cancer
in the present study, we carried out qRT-PCR and western blot
analysis in normal gastric mucosal cell line GES-1 and gastric
cancer cell line AGS. We found abundant Yap expression in AGS but
not in the normal gastric mucosal cells (Fig. 1A and B), suggesting that Yap may be
involved in the development of gastric cancer. To explain the role
of Yap in gastric cancer, we stably knocked down Yap expression
using the siRNA technique in AGS cells. The knockdown efficiency
was confirmed by western blotting (Fig.
1A and B). Since mitophagy is vital for cancer biological
function, we observed the change in mitophagy under Yap deficiency.
Loss of Yap reduced mitophagy as evidenced by a lower content of
mito-LC3II, p62, Beclin1 and Atg5 (Fig.
1C-G). These data indicated that Yap sustained mitophagy
activity. To further provide more evidence for the role of Yap in
mitophagy, we conducted the co-staining of mitochondria and
lysosomes. As shown in Fig. 1H,
loss of Yap reduced the overlap of mitochondria and lysosomes.
However, there was no change concerning the mitochondria and
lysosomes in the siRNA-control group. Altogether, these data
confirmed that Yap is a vital signal for mitophagy activation.
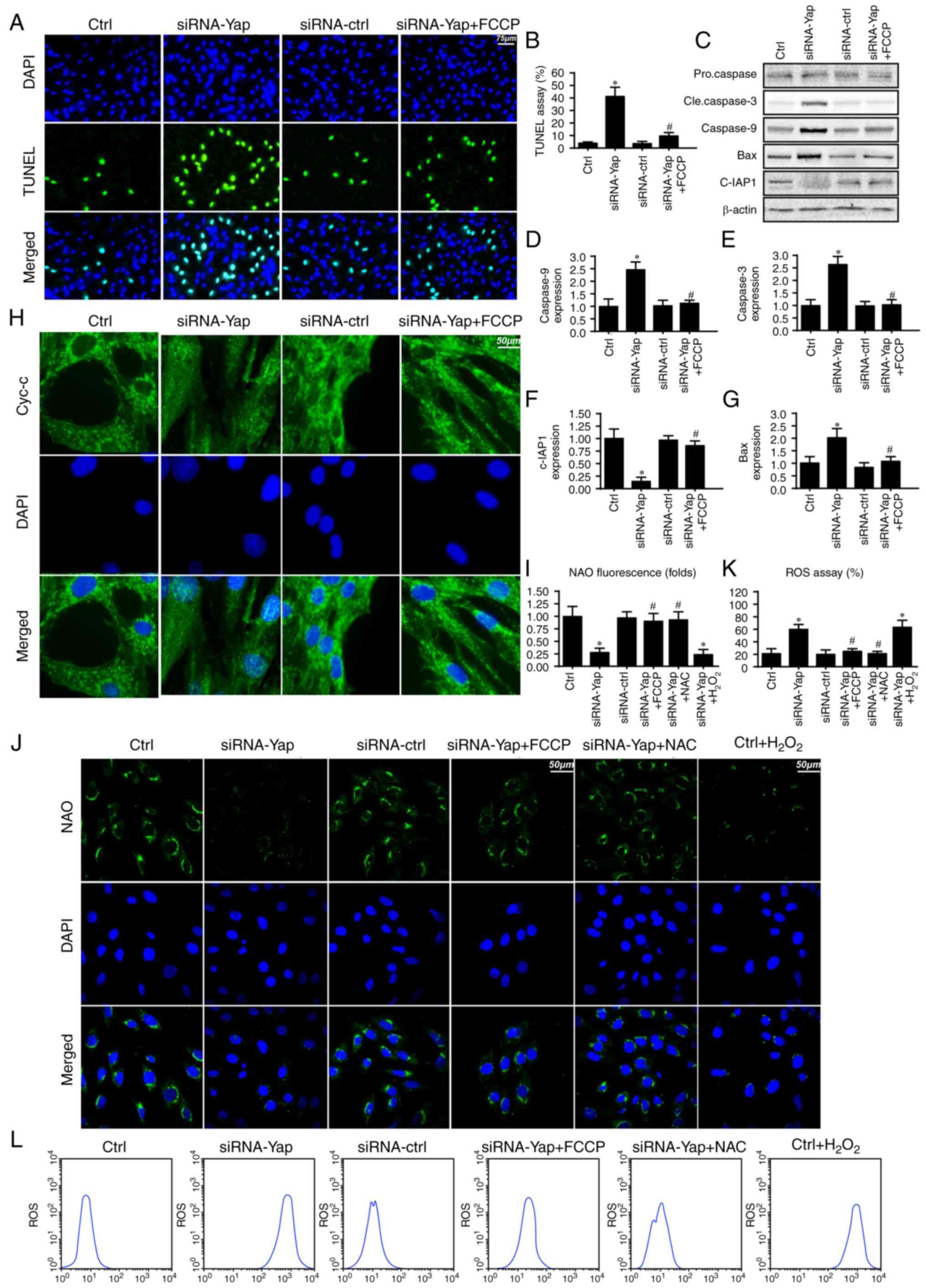
 | Figure 1.Yap is upregulated in gastric cancer
and sustains mitophagy activity. (A and B) Expression of Yap in
normal gastric mucosal cell line GES-1 and gastric cancer cell line
AGS. Yap was upregulated in gastric cancer when compared to the
normal cells. Meanwhile, siRNA was used to knockdown the expression
of Yap in gastric cancer in order to conduct loss-of-function assay
for Yap. The knockdown efficiency was examined via western blot
analyses. (C-G) The change in expression of mitophagy markers such
as mitochondrially contained LC3II, Beclin1, p62 and Atg5. Loss of
Yap was associated with mitophagy inhibition. Meanwhile, FCCP, an
activator of mitophagy, was used to conduct gain-of-function assay
for mitophagy. (H) The co-staining of mitochondria and lysosomes.
The overlap of mitochondria and lysosomes indicated mitophagy which
was demonstrated by orange fluorescence. (I) MTT assay was used to
detect cellular viability. (J) The change in caspase-3 activity. (K
and L) Wound healing assay was used to detect cellular
mobilization. After 24 h, the distance was measured and is
expressed as the unmigrated distance. *P<0.05 vs. the Ctrl
group, #P<0.05 vs. the siRNA-Yap group. Yap,
Yes-associated protein; Atg5, autophagy protein 5. |
Next, we used MTT, caspase-3 activity and wound
healing assay to examine the influence of mitophagy on gastric
cancer viability and migration. Meanwhile, FCCP, an activator of
mitophagy, was used as the gain-of-function assay in Yap-deleted
AGS cells. Compared to the control group, loss of Yap reduced
cellular viability as revealed by a lower OD value and increased
caspase-3 activity (Fig. 1I and J).
However, re-activation of mitophagy by FCCP under Yap deficiency
reversed cellular viability (Fig. 1I
and J). Similarly, results from the wound healing assay also
uncovered that Yap promoted cancer mobilization (Fig. 1K and L). Loss of Yap reduced the
distance of cancer mobilization and this tendency was reversed by
FCCP (Fig. 1K and L). Collectively,
these data indicated that Yap maintained gastric cancer viability
and mobilization via mitophagy.
Mitophagy inhibits gastric cancer
death via blocking the caspase-9-related apoptosis pathway
To further investigate the mechanism by which
mitophagy sustains cellular viability, we firstly detected the
cellular apoptotic rate under Yap deficiency. Loss of Yap increased
the number of TUNEL-positive cells (Fig. 2A and B). However, this harmful
effect of Yap deficiency was reversed by re-activation of
mitophagy. These data indicated that Yap preserved gastric cancer
cell viability via blocking cellular apoptosis.
Since mitophagy is a self-clearing system for
mitochondria; therefore, we proposed that Yap-regulated mitophagy
has the ability to reduce the caspase-9-related mitochondrial
apoptosis pathway. Firstly, we measured the change in proteins
related to the caspase-9 apoptosis pathway. Loss of Yap increased
caspase-3 and −9 and Bax expression but reduced c-IAP1 content
(Fig. 2C-G). These data indicated
that Yap deletion initiated caspase-9-related cellular apoptosis.
However, upon reactivation of mitophagy by FCCP, the apoptotic
markers were abated (Fig.
2C-G).
Apart from the downstream protein alteration, we
also observed that Yap loss aggravated cytochrome c (cyt-c) leakage
from mitochondria via immunofluorescence (Fig. 2H). However, this trend was reversed
by mitophagy activation by FCCP (Fig.
2H). The cyt-c release was derived from mitochondrial oxidative
stress. In healthy cells, mitochondrial cardiolipin retains cyt-c
in the inner membrane of mitochondria. However, upon oxidation,
cardiolipin may liberate cyt-c contributing to cyt-c leakage into
the cytoplasm via mPTP. Therefore, we used NAO to observe
cardiolipin oxidation. As shown in Fig.
2I and J, loss of Yap increased the expression of oxidized
cardiolipin (CL). However, activation of mitophagy may reduce
cardiolipin oxidation. Since cardiolipin oxidation is derived from
excessive ROS, we aimed to ascertain whether Yap-regulated
mitophagy had the ability to alleviate cellular oxidative stress.
As shown in Fig. 2K and L, Yap
deletion caused ROS overproduction via flow cytometric analysis.
Upon re-activation of mitophagy in Yap-deleted gastric cells by
FCCP, the ROS content was reduced. Notably, we used the exogenous
H2O2 to mimic the ROS outburst in the control
gastric cancer cells, which was used as the positive control group.
Meanwhile, NAC, a ROS scavenger, was used to remove the oxidative
stress in Yap-deleted cells, which was used as the negative control
group. Exogenous H2O2 not only increased the
cellular ROS content (Fig. 2L) but
also induced cardiolipin (Fig. 2J).
However, NAC had the ability to reduce oxidative stress (Fig. 2L) and (Fig. 2J) in Yap-deleted cells. Altogether,
these data indicated that Yap was the housekeeper for gastric
cancer cell survival via activation of mitophagy which blocked the
caspase-9-related apoptotic pathways.
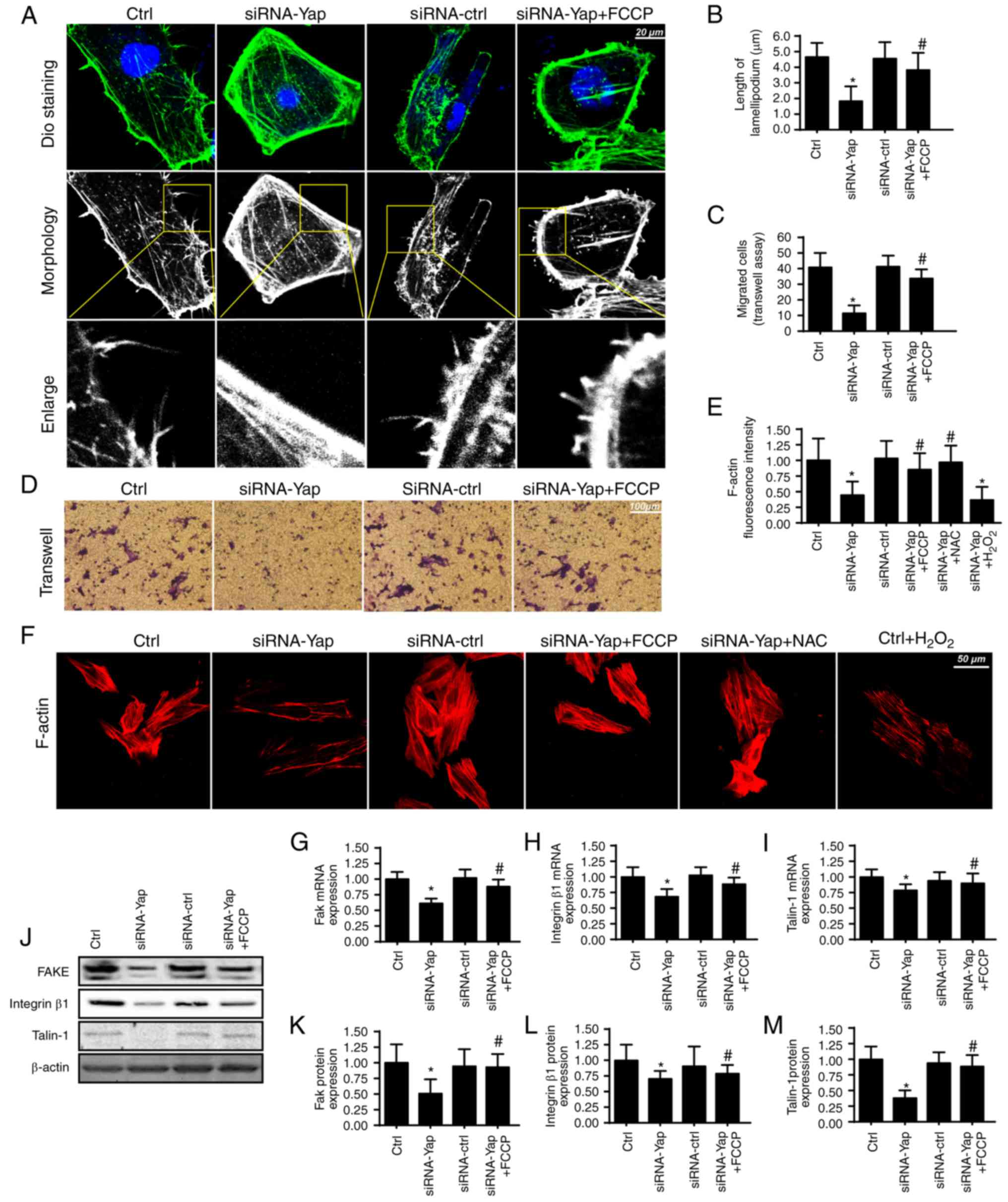
Mitophagy handles F-actin-based
cellular migration via reducing oxidative stress
Apart from cellular apoptosis, cellular migration is
another factor for the development of gastric cancer. Since cancer
migration is related to the F-actin-mediated cytoskeletal
rearrangement, F-actin may contribute to the formation of
lamellipodia which directs the cancer cells to move in some
direction. In the present study, we used the cell membrane dye DiO
to observe the change in lamellipodium. We found that loss of Yap
impaired the formation of lamellipodia (Fig. 3A and B). The shorter and less
lamellipodium appeared in response to the Yap loss (Fig. 3A and B), which was accompanied by a
lower number of migrated cells via Transwell assay (Fig. 3C and D). However, activation of
mitophagy reversed the formation of lamellipodium and therefore
facilitated cellular migration. These data indicated that Yap
regulated lamellipodium-based migration via mitophagy.
The lamellipodium consists of F-actin, a kind of
stress fiber (41). We found that
Yap loss caused F-actin downregulation via immunofluorescence assay
(Fig. 3E and F). However,
activation of mitophagy reversed the F-actin expression despite
deletion of Yap. To ascertain the underlying mechanism underlying
F-actin collapse under Yap inhibition, we focused on the
ROS-induced oxidative stress. Previous research suggests that
F-actin is regulated by oxidative stress and excessive ROS may
impair F-actin synthesis (42). In
the present study, we found that exogenous
H2O2 treatment reduced the F-actin
immunofluorescence intensity which was similar to the results noted
in the Yap-deleted cells (Fig. 3E and
F). However, removal of ROS via NAC in Yap-deleted cells,
restored the F-actin content, comparable to the effect of mitophagy
reactivation in Yap-deleted cells.
Apart from cytoskeletal rearrangement, adhesive
proteins are also implicated in cellular migration (43). The expression of the adhesive
markers FAK, integrin β1 and Talin-1 were analyzed using qPCR
(Fig. 3G-I) and western blot
analyses (Fig. 3J-M). The results
revealed that Yap inhibition reduced FAK, Integrin β1 and Talin-1
expression at the mRNA and protein levels. In contrast, mitophagy
activation by FCCP exerted opposite effects in relation to Yap
deletion. Altogether, these results indicatedan important role of
Yap in modulating gastric cancer migration via mitophagy.
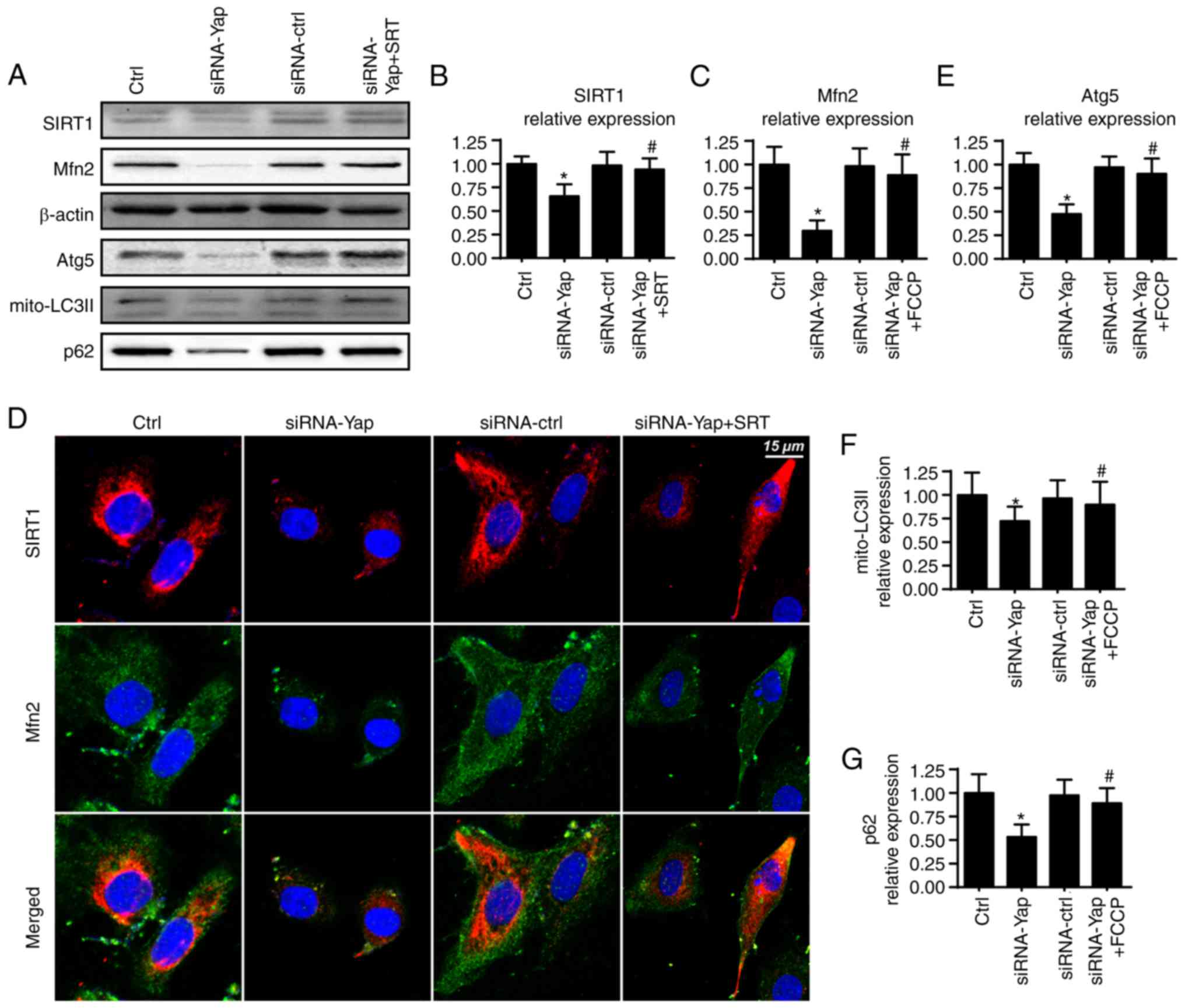
Yap modulates mitophagy via the
SIRT1/Mfn2 pathway
To ascertain the mechanism by which Yap sustains
mitophagy activity, we focused on Mfn2 expression which is a novel
receptor of mitophagy (44). We
demonstrated that loss of Yap expression reduced Mfn2 expression
when compared to the control group (Fig. 4A and C). Furthermore, several
studies have suggested that Sirtuin 1 (SIRT1) is the upstream
inducer of Mfn2 expression (45),
and therefore, we aimed to ascertain whether SIRT1 is involved in
Mfn2 regulation by Yap. Firstly, we found that loss of Yap was
associated with the downregulation of SIRT1 (Fig. 4A and B). Furthermore, SIRT1
activator SRT1720 was used and it reversed SIRT1 expression albeit
Yap deletion (Fig. 4A and B).
Moreover, application of SRT1720 further restored siRNA-Yap-reduced
Mfn2 expression (Fig. 4A-C). These
data established the regulatory role of SIRT1 in Mfn2
expression.
To provide further evidence for the role of SIRT1 in
Mfn2 expression, we co-stained for SIRT1 and Mfn2. As shown in
Fig. 4D, compared to the control
group, loss of Yap reduced the SIRT1 and Mfn2 expression. However,
SIRT1 activator SRT1720 reversed the SIRT1 expression and
subsequent upregulated the Mfn2 content. Next, to demonstrate
whether the SIRT/Mfn2 pathway was responsible for mitophagy
activation, we examined the proteins involved in mitophagy. As
shown in Fig. 4A and E-G, compared
to the Yap-silenced group, reactivation of SIRT1 reversed the
mitophagy activity as evidenced by increased mito-LC3II, Atg5 and
p62. Altogether, these findings demonstrated that Yap signaled
mitophagy via the SIRT1/Mfn2 pathway.
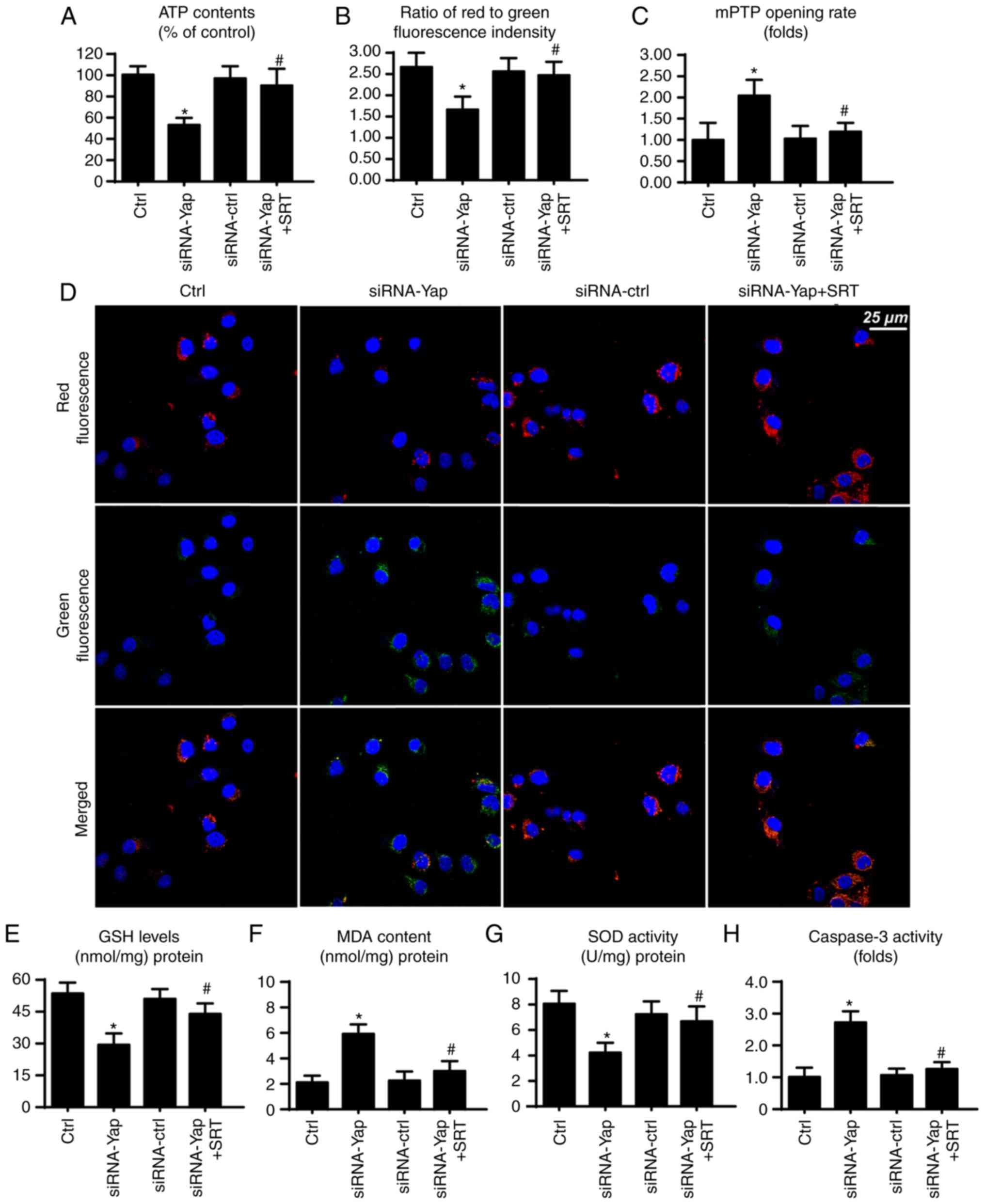
SIRT1/Mfn2 pathway is also involved in
mitochondrial protection
Finally, to ascertain whether SIRT1 and Mfn2 are
involved in gastric cancer viability, we observed mitochondrial
function under SIRT1 activation. Firstly, mitochondria are the
center for cellular energy metabolism. However, loss of Yap reduced
ATP production (Fig. 5A). This
harmful effect was reversed by SIRT1 reactivation. Since the
mitochondrial membrane potential is the source of ATP production
(33), we consequently assessed the
change in mitochondrial potential. Compared to the control group,
loss of Yap reduced the membrane potential, which was restored via
SIRT1 activator (Fig. 5B and D).
Furthermore, loss of Yap is associated with increased mPTP opening
rate. In contrast, reactivation of SIRT1 suppressed the extensive
mPTP opening (Fig. 5C). The mPTP
opening provides a channel for proton leakage into the cytoplasm,
finally leading to membrane potential collapse.
Apart from energy production, mitochondria also
produce excessive ROS to induce cellular apoptosis (46). In the present study, we found that
cellular anti-oxidant factors such as GSH and SOD were
downregulated in response to the loss of Yap (Fig. 5E and G). In contrast, MDA, an end
product of lipid peroxidation, was upregulated (Fig. 5F), indicative of cellular oxidative
stress. However, recovery of SIRT1 exerted an opposite effect and
reduced the levels of these markers of cellular oxidative stress
(Fig. 5E-G). Furthermore, we also
assessed caspase-3 activity to ascertain whether SIRT1/Mfn2 is
involved in cellular damage. Restoration of SIRT1 via its activator
reduced the caspase-3 activity when compared to the Yap deficiency
group (Fig. 5H). Collectively,
these data indicate that the SIRT1/Mfn2 pathway regulated by Yap is
vital for mitochondrial function and cellular viability.
Discussion
Gastric cancer is a multifactorial process, and many
molecular alterations have been shown to influence tumor initiation
and development through abnormal gene expression or protein
alterations (1). Gastric cancer is
the fifth leading cause of cancer-related death in both males and
females worldwide and accounts for 8.8% of all such deaths
(33). Over the past several
decades, studies concerning gastric cancer have provided knowledge
about the genetic, epigenetic, transcriptome and metabolic
alterations underlying this disease. Although the exact cause of
gastric cancer is unclear, its pathogenesis is the same as that of
other malignant tumors. it is a multi-step, multi-factorial
comprehensive disease. Gastric cancer cases can be divided into
early- and advanced-stage gastric cancer. In recent years, the
5-year mortality rate has significantly decreased for early gastric
cancer patients given the development of enteroscopy and surgical
techniques. However, for advanced gastric cancer, the 5-year
mortality remains 30–50% (3).
Early-stage gastric cancers are limited to the mucosa or submucosa,
regardless of the size of the lesion and the presence of lymph node
metastasis. Cancer that extends beyond the submucosa to invade the
gastric muscular layer is middle-stage gastric cancers (47), whereas tumors that infiltrate into
or beyond the subserosa to nearby organs or metastasize are
advanced-stage gastric cancers. In brief, advanced-stage gastric
cancer patients have a reduced survival rate and the tumors exhibit
a more aggressive or invasive property. Since the stage of the
tumor determines treatment effectiveness and treatment strategy, it
is worthwhile to explore the mechanism of gastric cancer
progression such as survival, migration, invasion and
apoptosis.
In the present study, we identified that Yap is a
necessary factor for maintaining gastric cancer migration and
viability. Yes-associated protein (Yap) is a 65 kDa proline-rich
phosphoprotein, which is located at locus 11q22 (48). A previous study reported that YAP
may have oncogenic functions, and elevated YAP expression is
associated with malignant tumors (21,49).
In the present study, we found that Yap expression is increased in
gastric cancer when compared to that noted in normal gastric
mucosal cells. Moreover, Yap expression was associated with gastric
cancer cell viability and migration, suggesting the tumor-promotive
effect of Yap. To the best of our knowledge, this is the first
study to describe the role of Yap in gastric cancer migration and
survival, which supports the mechanism involved in the tumor
development and progression by Yap. More importantly, our data
offer a potential therapeutic target to intervene against gastric
cancer involving cancer cell survival and migration. However, more
clinical data are needed to support our theory.
We determined that Yap promoted cancer cell survival
and mobilization via mitophagy. Loss of Yap abated the level of
mitophagy, leading to the activation of the caspase-9-related
apoptotic pathways (50). Moreover,
mitophagy inactivation was also related to excessive oxidative
stress which caused F-actin degradation and lamellipodium collapse,
finally leading to impairment in cancer cell motility. Although the
majority of studies have uncovered the anti-apoptotic action of
mitophagy on cell fate regardless of cancer cells or normal cells
(51,52), little attention has focused on the
relationship between cellular migration and mitophagy. We
demonstrated that mitophagy contributed to the cellular migration
by reducing ROS. Previous studies have found that oxidative injury
may induce F-actin degradation and collapse, which eventually
blunts the formation of lamellipodium. These findings were similar
to our results. Based on this, we confirmed the core role of
mitophagy in cellular migration (53). Notably, we also found that mitophagy
had the ability to increase ATP production. Considering that the
cellular migration is dependent on energy production, the
ATP-promotive effect of mitophagy could be another factor to
enhance cellular migration.
We found that Yap was the upstream controller of
mitophagy, which elucidated the regulatory machinery of mitophagy.
Mechanistically, Yap preserved Mfn2 expression which enhanced
mitophagy activity. Furthermore, SIRT1 was signaled by Yap and
sustained Mfn2-required mitophagy. These data hint at the possible
links between Yap and mitochondria. Recent research has found that
Yap preserves mitochondrial function via regulation of JNK pathways
(54). In the present study, we
confirmed that the protective effect of Yap on mitochondria was
attributed to mitophagy activation. Accordingly, our findings
provide more information to the role of Yap in tumorigenesis
involving mitochondrial homeostasis.
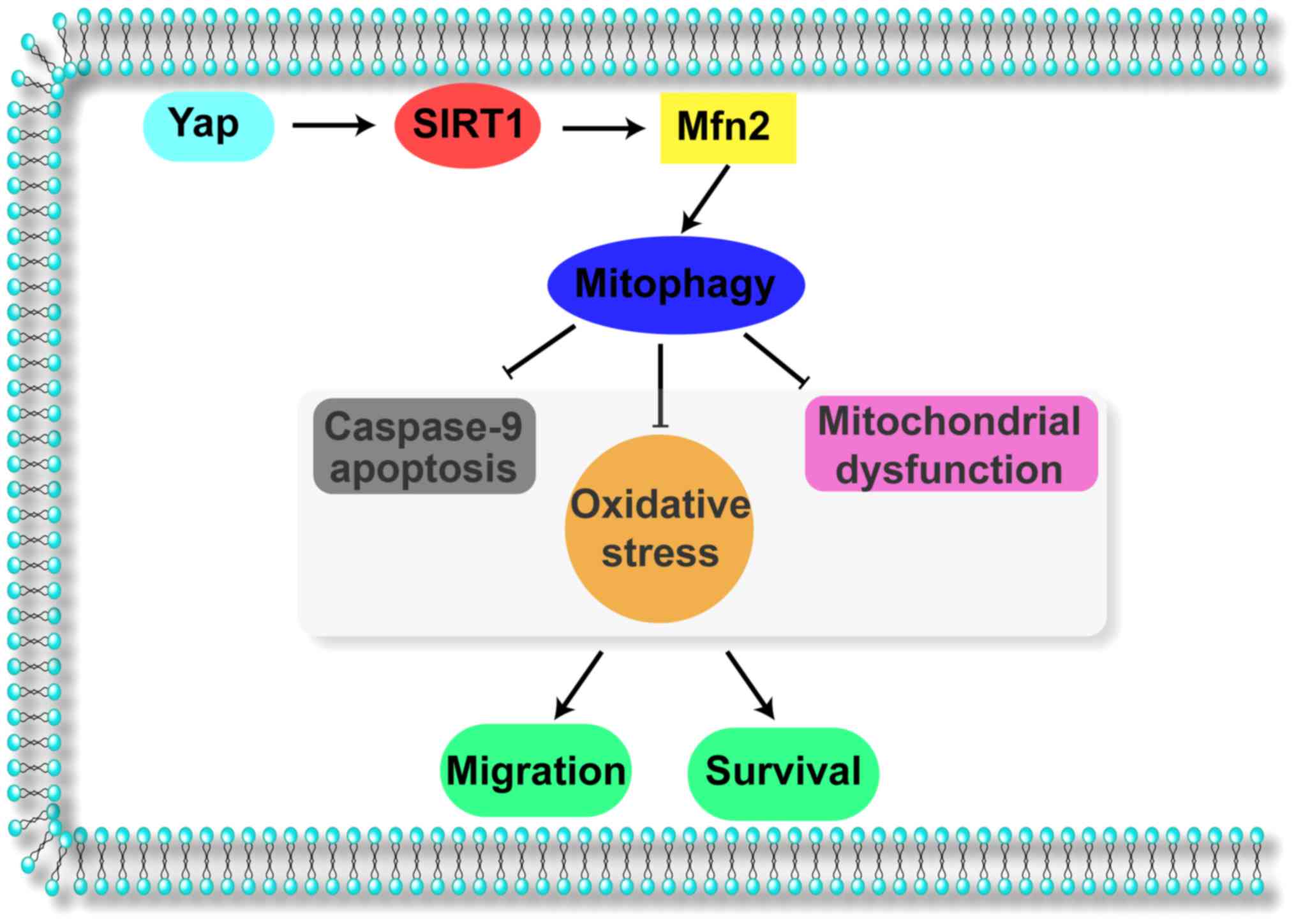
Collectively, the results of our study illustrate
the important role of Yap in gastric cancer migration and survival
(Fig. 6). Yap signals Sirt1 which
activates Mfn2-mediated mitophagy, contributing to mitochondrial
homeostasis. The Yap/SIRT1/Mfn2/mitophagy pathways block the
caspase-9-related apoptotic axis and enhance F-actin-based cellular
migration, responsible for gastric cancer cell migration and
survival. These data identify Yap/SIRT1/Mfn2/mitophagy pathways as
a potential target for the treatment of gastric cancer, and
highlight a new strategy for treating gastric cancer involving Yap
protein and Mfn2-mediated mitophagy.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
project of Songjiang District Science and Technology Commission
(no. 15SJGG29).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HZY, CMQ and WWS were involved in conception and
design, performance of experiments, data analysis and
interpretation, as well as manuscript writing. MMG, FX, JZ and LZ
were involved in data analysis and interpretation.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Authors' information
Not applicable.
References
|
1
|
Procaccio L, Schirripa M, Fassan M,
Vecchione L, Bergamo F, Prete AA, Intini R, Manai C, Dadduzio V,
Boscolo A, et al: Immunotherapy in gastrointestinal cancers. Biomed
Res Int. 2017:43465762017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garattini SK, Basile D, Cattaneo M,
Fanotto V, Ongaro E, Bonotto M, Negri FV, Berenato R, Ermacora P,
Cardellino GG, et al: Molecular classifications of gastric cancers:
Novel insights and possible future applications. World J
Gastrointest Oncol. 9:194–208. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McLean MH and El-Omar EM: Genetics of
gastric cancer. Nat Rev Gastroenterol Hepatol. 11:664–674. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee YC, Chiang TH, Chou CK, Tu YK, Liao
WC, Wu MS and Graham DY: Association between Helicobacter pylori
eradication and gastric cancer incidence: a systematic review and
meta-analysis. Gastroenterology. 150:1113–1124 e5. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Veitch AM, Uedo N, Yao K and East JE:
Optimizing early upper gastrointestinal cancer detection at
endoscopy. Nat Rev Gastroenterol Hepatol. 12:660–667. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rizzo NR, Hank NC and Zhang J: Detecting
presence of cardiovascular disease through mitochondria respiration
as depicted through biophotonic emission. Redox Biol. 8:11–17.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du K, Ramachandran A and Jaeschke H:
Oxidative stress during acetaminophen hepatotoxicity: Sources,
pathophysiological role and therapeutic potential. Redox Biol.
10:148–156. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou H, Wang S, Zhu P, Hu S, Chen Y and
Ren J: Empagliflozin rescues diabetic myocardial microvascular
injury via AMPK-mediated inhibition of mitochondrial fission. Redox
Biol. 15:335–346. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin YW, Lee LM, Lee WJ, Chu CY, Tan P,
Yang YC, Chen WY, Yang SF, Hsiao M and Chien MH: Melatonin inhibits
MMP-9 transactivation and renal cell carcinoma metastasis by
suppressing Akt-MAPKs pathway and NF-κB DNA-binding activity. J
Pineal Res. 60:277–290. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pariente R, Pariente JA, Rodriguez AB and
Espino J: Melatonin sensitizes human cervical cancer HeLa cells to
cisplatin-induced cytotoxicity and apoptosis: Effects on oxidative
stress and DNA fragmentation. J Pineal Res. 60:55–64. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Prieto-Dominguez N, Ordóñez R, Fernández
A, Méndez-Blanco C, Baulies A, Garcia-Ruiz C, Fernández-Checa JC,
Mauriz JL and González-Gallego J: Melatonin-induced increase in
sensitivity of human hepatocellular carcinoma cells to sorafenib is
associated with reactive oxygen species production and mitophagy. J
Pineal Res. 61:396–407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burridge K: Focal adhesions: A personal
perspective on a half century of progress. FEBS J. 284:3355–3361.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou H, Zhu P, Guo J, Hu N, Wang S, Li D,
Hu S, Ren J, Cao F and Chen Y: Ripk3 induces mitochondrial
apoptosis via inhibition of FUNDC1 mitophagy in cardiac IR injury.
Redox Biol. 13:498–507. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi X and Sun X: Regulation of paclitaxel
activity by microtubule-associated proteins in cancer chemotherapy.
Cancer Chemother Pharmacol. 80:909–917. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou H, Li D, Zhu P, Hu S, Hu N, Ma S,
Zhang Y, Han T, Ren J, Cao F and Chen Y: Melatonin suppresses
platelet activation and function against cardiac
ischemia/reperfusion injury via PPARγ/FUNDC1/mitophagy pathways. J
Pineal Res. 63:2017. View Article : Google Scholar :
|
|
16
|
Tol MJ, Ottenhoff R, van Eijk M, Zelcer N,
Aten J, Houten SM, Geerts D, van Roomen C, Bierlaagh MC, Scheij S,
et al: A PPARγ-bnip3 axis couples adipose mitochondrial
fusion-fission balance to systemic insulin sensitivity. Diabetes.
65:2591–2605. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sebastián D, Sorianello E, Segalés J,
Irazoki A, Ruiz-Bonilla V, Sala D, Planet E, Berenguer-Llergo A,
Muñoz JP, Sánchez-Feutrie M, et al: Mfn2 deficiency links
age-related sarcopenia and impaired autophagy to activation of an
adaptive mitophagy pathway. EMBO J. 35:1677–1693. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leboucher GP, Tsai YC, Yang M, Shaw KC,
Zhou M, Veenstra TD, Glickman MH and Weissman AM: Stress-induced
phosphorylation and proteasomal degradation of mitofusin 2
facilitates mitochondrial fragmentation and apoptosis. Mol Cell.
47:547–557. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng J, Ren KD, Yang J and Luo XJ:
Mitochondrial E3 ubiquitin ligase 1: A key enzyme in regulation of
mitochondrial dynamics and functions. Mitochondrion. 28:49–53.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao T, Huang X, Han L, Wang X, Cheng H,
Zhao Y, Chen Q, Chen J, Cheng H, Xiao R and Zheng M: Central role
of mitofusin 2 in autophagosome-lysosome fusion in cardiomyocytes.
J Biol Chem. 287:23615–23625. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ahmed AA, Mohamed AD, Gener M, Li W and
Taboada E: YAP and the Hippo pathway in pediatric cancer. Mol Cell
Oncol. 4:e12951272017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qiao Y, Chen J, Lim YB, Finch-Edmondson
ML, Seshachalam VP, Qin L, Jiang T, Low BC, Singh H, Lim CT and
Sudol M: YAP regulates actin dynamics through ARHGAP29 and promotes
metastasis. Cell Rep. 19:1495–1502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakamura M, Zhai P, Del Re DP, Maejima Y
and Sadoshima J: Mst1-mediated phosphorylation of Bcl-xL is
required for myocardial reperfusion injury. JCI Insight. 1(pii):
e862172016.PubMed/NCBI
|
|
24
|
Griffiths HR, Gao D and Pararasa C: Redox
regulation in metabolic programming and inflammation. Redox Biol.
12:50–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin C, Chao H, Li Z, Xu X, Liu Y, Hou L,
Liu N and Ji J: Melatonin attenuates traumatic brain injury-induced
inflammation: A possible role for mitophagy. J Pineal Res.
61:177–186. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oliver PM, Crooks JA, Leidl M, Yoon EJ,
Saghatelian A and Weibel DB: Localization of anionic phospholipids
in Escherichia coli cells. J Bacteriol. 196:3386–3398. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu S, Wang X, Geng P, Tang X, Xiang L, Lu
X, Li J, Ruan Z, Chen J, Xie G, et al: Melatonin regulates PARP1 to
control the senescence-associated secretory phenotype (SASP) in
human fetal lung fibroblast cells. J Pineal Res. 63:2017.
View Article : Google Scholar
|
|
28
|
Quintana C, Cabrera J, Perdomo J, Estévez
F, Loro JF, Reiter RJ and Quintana J: Melatonin enhances
hyperthermia-induced apoptotic cell death in human leukemia cells.
J Pineal Res. 61:381–395. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mao L, Dauchy RT, Blask DE, Dauchy EM,
Slakey LM, Brimer S, Yuan L, Xiang S, Hauch A, Smith K, et al:
Melatonin suppression of aerobic glycolysis (Warburg effect),
survival signalling and metastasis in human leiomyosarcoma. J
Pineal Res. 60:167–177. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fuhrmann DC and Brüne B: Mitochondrial
composition and function under the control of hypoxia. Redox Biol.
12:208–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jin Q, Li R, Hu N, Xin T, Zhu P, Hu S, Ma
S, Zhu H, Ren J and Zhou H: DUSP1 alleviates cardiac
ischemia/reperfusion injury by suppressing the Mff-required
mitochondrial fission and Bnip3-related mitophagy via the JNK
pathways. Redox Biol. 14:576–587. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi C, Cai Y, Li Y, Li Y, Hu N, Ma S, Hu
S, Zhu P, Wang W and Zhou H: Yap promotes hepatocellular carcinoma
metastasis and mobilization via governing
cofilin/F-actin/lamellipodium axis by regulation of
JNK/Bnip3/SERCA/CaMKII pathways. Redox Biol. 14:59–71. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li M, Pi H, Yang Z, Reiter RJ, Xu S, Chen
X, Chen C, Zhang L, Yang M, Li Y, et al: Melatonin antagonizes
cadmium-induced neurotoxicity by activating the transcription
factor EB-dependent autophagy-lysosome machinery in mouse
neuroblastoma cells. J Pineal Res. 61:353–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nduhirabandi F, Lamont K, Albertyn Z, Opie
LH and Lecour S: Role of toll-like receptor 4 in melatonin-induced
cardioprotection. J Pineal Res. 60:39–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu Z, Gan L, Xu Y, Luo D, Ren Q, Wu S and
Sun C: Melatonin alleviates inflammasome-induced pyroptosis through
inhibiting NF-κB/GSDMD signal in mice adipose tissue. J Pineal Res.
63:2017. View Article : Google Scholar
|
|
36
|
Lin S, Hoffmann K, Gao C, Petrulionis M,
Herr I and Schemmer P: Melatonin promotes sorafenib-induced
apoptosis through synergistic activation of JNK/c-jun pathway in
human hepatocellular carcinoma. J Pineal Res. 62:2017. View Article : Google Scholar
|
|
37
|
Zhou H, Du W, Li Y, Shi C, Hu N, Ma S,
Wang W and Ren J: Effects of melatonin on fatty liver disease: The
role of NR4A1/DNA-PKcs/p53 pathway, mitochondrial fission, and
mitophagy. J Pineal Res. 64:e124502018. View Article : Google Scholar
|
|
38
|
Zhang Y, Zhou H, Wu W, Shi C, Hu S, Yin T,
Ma Q, Han T, Zhang Y, Tian F and Chen Y: Liraglutide protects
cardiac microvascular endothelial cells against
hypoxia/reoxygenation injury through the suppression of the
SR-Ca2+-XO-ROS axis via activation of the
GLP-1R/PI3K/Akt/survivin pathways. Free Radic Biol Med. 95:278–292.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou H, Hu S, Jin Q, Shi C, Zhang Y, Zhu
P, Ma Q, Tian F and Chen Y: Mff-Dependent mitochondrial fission
contributes to the pathogenesis of cardiac microvasculature
ischemia/reperfusion injury via induction of mROS-mediated
cardiolipin oxidation and HK2/VDAC1 disassociation-involved mPTP
opening. J Am Heart Assoc. 6:e0053282017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Poku RA, Salako OO, Amissah F, Nkembo AT,
Ntantie E and Lamango NS: Polyisoprenylated cysteinyl amide
inhibitors induce caspase 3/7- and 8-mediated apoptosis and inhibit
migration and invasion of metastatic prostate cancer cells. Am J
Cancer Res. 7:1515–1527. 2017.PubMed/NCBI
|
|
41
|
Zhou H, Zhang Y, Hu S, Shi C, Zhu P, Ma Q,
Jin Q, Cao F, Tian F and Chen vY: Melatonin protects cardiac
microvasculature against ischemia/reperfusion injury via
suppression of mitochondrial fission-VDAC1-HK2-mPTP-mitophagy axis.
J Pineal Res. 63:e124132017. View Article : Google Scholar :
|
|
42
|
Wang C, Blough ER, Arvapalli R, Dai X,
Paturi S, Manne N, Addagarla H, Triest WE, Olajide O and Wu M:
Metabolic syndrome-induced tubulointerstitial injury: Role of
oxidative stress and preventive effects of acetaminophen. Free
Radic Biol Med. 65:1417–1426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Verma NK and Kelleher D: Not Just an
adhesion molecule: LFA-1 contact tunes the T lymphocyte program. J
Immunol. 199:1213–1221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sebastián D and Zorzano áA: When MFN2
(mitofusin 2) met autophagy: A new age for old muscles. Autophagy.
12:2250–2251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sooyeon L, Go KL and Kim JS: Deacetylation
of mitofusin-2 by sirtuin-1: A critical event in cell survival
after ischemia. Mol Cell Oncol. 3:e10874522015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Quijano C, Trujillo M, Castro L and
Trostchansky A: Interplay between oxidant species and energy
metabolism. Redox Biol. 8:28–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tan DX, Hardeland R, Back K, Manchester
LC, Alatorre-Jimenez MA and Reiter RJ: On the significance of an
alternate pathway of melatonin synthesis via 5-methoxytryptamine:
Comparisons across species. J Pineal Res. 61:27–40. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eldred M: YAP is crucial for lung
branching morphogenesis. Science. 356:150–151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sundar R, Chenard-Poirier M, Collins DC
and Yap TA: Imprecision in the Era of precision medicine in
non-small cell lung cancer. Front Med. 4:392017. View Article : Google Scholar
|
|
50
|
de Souza P, Guarido KL, Scheschowitsch K,
da Silva LM, Werner MF, Assreuy J and da Silva-Santos JE: Impaired
vascular function in sepsis-surviving rats mediated by oxidative
stress and Rho-Kinase pathway. Redox Biol. 10:140–147. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Doskey CM, Buranasudja V, Wagner BA,
Wilkes JG, Du J, Cullen JJ and Buettner GR: Tumor cells have
decreased ability to metabolize H2O2: Implications for
pharmacological ascorbate in cancer therapy. Redox Biol.
10:274–284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mailloux RJ and Treberg JR: Protein
S-glutathionlyation links energy metabolism to redox signaling in
mitochondria. Redox Biol. 8:110–118. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Han J, Weisbrod RM, Shao D, Watanabe Y,
Yin X, Bachschmid MM, Seta F, Janssen-Heininger YMW, Matsui R, Zang
M, et al: The redox mechanism for vascular barrier dysfunction
associated with metabolic disorders: Glutathionylation of Rac1 in
endothelial cells. Redox Biol. 9:306–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kitsati N, Mantzaris MD and Galaris D:
Hydroxytyrosol inhibits hydrogen peroxide-induced apoptotic
signaling via labile iron chelation. Redox Biol. 10:233–242. 2016.
View Article : Google Scholar : PubMed/NCBI
|