Over the past century, human life expectancy has
increased significantly, settling at approximately 76.3 years for
males and 81.2 for females (1). Such
an increment in lifespan has been associated with a huge surge in
the most diffused life-threatening diseases, including
cardiovascular, neurodegenerative and oncological pathologies
(2).
Although aging represents a common risk factor, the
physiological and molecular mechanisms behind the development of
age-related disorders differ completely, particularly when
comparing cancer with neurodegenerative disorders (3,4).
Supporting this evidence, recent epidemiological data have
demonstrated that there is an inverse association between cancer
and neurodegeneration (5,6).
Despite this inverse association between the
incidence of tumors and the rise of neurodegenerative disorders,
some conditions, such as inflammation (7–9), the
alteration of the intestinal microbiota (10–13),
diet-related diseases (14–18) and risks related to the exposure to
environmental pollutants (19–25) are
involved in the development of both pathologies. Consistently, the
association between the development of neurodegenerative diseases
and the onset of tumors may be either direct or indirect. As a
direct consequence, it has been shown that several benign and
malignant types of cancer are associated with neurological, as well
as neurodegenerative disorders (26–28).
In particular, it has been demonstrated that
although glioblastoma multiforme (GBM) and Alzheimer's disease (AD)
share the same molecular pathways, substantial differences exist in
their modulation (29,30). In fact, while rapid cell proliferation
and apoptotic cell arrest are typical features of GBM, cellular
damage and subsequent cell death are common consequences in AD
(31,32).
To date, the only available treatments for AD are
only palliative, as they are capable of delaying memory and
cognitive function impairment, without actually blocking neuronal
loss. The main drugs used in the treatment of AD are cholinesterase
inhibitors and glutamatergic N-methyl D-aspartate (NMDA) receptors
antagonists, such as memantine (Namenda), respectively able to
improve neuropsychiatric symptoms and neuronal cell-to-cell
communication (33). Importantly, the
benefits of such therapies are not long-lasting and are coupled
with adverse impairing effects.
GBM is a widely diffused brain malignancy, as well
as the most aggressive tumor of the central nervous system
(34,35). Currently, GBM treatment options are
limited, typically represented by surgical resection of the tumor
mass (when the lesion does not involve vascular and nerve
structures), followed by radiotherapy and chemotherapy (36). Despite advancements being made in
anticancer treatments (37), the
therapeutic approaches available for GBM are often ineffective,
given the high rate of GBM relapse and drug resistance (36). Recently, in vitro studies have
demonstrated that treatment with nitric oxide-releasing HIV
protease inhibitors previously adopted for other tumors (38,39), is
effective in reducing the proliferation of GBM cancer cells
(40,41).
Notably, the diagnostic and therapeutic approaches
that are normally used to recognize and to treat cognitive deficits
and dementia symptoms, appear to be slightly effective in the early
detection and treatment of brain precancerous lesions. Although
they need to be further validated in a larger cohort of patients,
transcranial magnetic stimulation (TMS) and transcranial Doppler
ultrasonography, normally used for vascular cognitive impairments
(42–47), AD (48),
restless leg syndrome (49–52), and other neurological syndromes
(53–56), appear to be promising approaches
suitable for the diagnosis and cure of precancerous brain
lesions.
A concern about GBM management is relative to the
lack of effective biomarkers. In particular, a number of biomarkers
have been proposed to solve this issue. Several studies have
highlighted the possible application of extracellular protein
biomarkers, such as extracellular matrix proteins, vascular
endothelial growth factor (VEGF), angiogenesis-associated proteins,
matrix metalloproteinases (MMPs; MMP-2, MMP-9) and astrocyte
elevated gene-1 (AEG-1), macrophage migration inhibitory factor
(MIF) and functionally-related genes (DD-T; CD74, CD44, CXCR2 and
CXCR4) (57–59). Other proteins can be also used for the
treatment or prognostic evaluation of tumor development (60,61).
Given the lack of effective diagnostic strategies,
as well as treatments able to effectively cure both GBM and AD,
there is an urgent need for the identification of novel diagnostic
biomarkers and therapeutic targets for the effective treatment of
such pathologies. Moreover, the understanding of the expression
patterns of such biomarkers may prove to be useful in order to
further demonstrate the existence of an inverse association between
GBM and AD. In this context, several studies have demonstrated that
the evaluation of microRNA (miRNA or miR) expression levels in
patients compared with their healthy controls, may provide
information on the development of several diseases, including
cancer and neurodegenerative disorders (62–65).
Indeed, miRNAs are involved in both physiological and pathological
processes; therefore studying their alterations in GBM and AD may
prove to be helpful in detecting early the onset of such
pathologies.
On this ground, the aim of this study was to analyze
miRNA expression profiling datasets of GBM and AD obtained from the
Gene Expression Omnibus (GEO) DataSets portal in order to identify
specific miRNAs de-regulated in both diseases. Once the presence of
altered miRNAs shared between GBM and AD would be established, the
second aim of this study was to determine whether their expression
levels are inversely associated.
The selection of both GBM and AD miRNA expression
profiling datasets was performed using the publicly available GEO
DataSets database, as previously reported (66,67).
Briefly, for the selection of GBM miRNA datasets the following
search terms were used: ‘{[‘non coding rna profiling by
array’(DataSet Type)] AND glioblastoma} AND ‘Homo
sapiens’[porgn:__txid9606]’; while for the selection of AD
datasets the search terms used were as follows: ‘{[‘non coding rna
profiling by array’(DataSet Type)] AND Alzheimer} AND ‘Homo
sapiens’[porgn:__txid9606]’. These search criteria allowed the
identification of different miRNA expression datasets of which only
those with >10 samples (total of normal and pathological
samples) were selected for the further computational analyses.
Given the low number of AD miRNA microarray platforms, datasets
containing <10 samples were considered. Datasets including the
expression data of GBM or AD within in vitro models were not
considered for the analyses.
Following dataset selection, the data matrices were
downloaded and differential analyses were performed between normal
and pathological samples using the GEO2R tool available on GEO
DataSets. Since different miRNA microarray platforms were adopted,
the differentially expressed miRNAs of each dataset were annotated
using the last version of miRBase (miRBase version 22) (68). The miRNA expression fold change (FC)
was expressed as base-2 logarithm of FC (log2FC) to normalize the
miRNA expression values obtained from different microarray
platforms.
All the miRNAs with a value of P≤0.01 were
considered for the merging analyses and the following
identification of miRNAs involved in GBM and AD.
The lists of differentially expressed miRNAs
obtained from the GBM and AD datasets were merged through a Venn
diagram calculating tool, in order to obtain miRNAs shared with
>1 dataset (http://bioinformatics.psb.ugent.be/webtools/Venn/). In
particular, only the miRNAs contained at least in the 50% of GBM
and AD datasets were considered. The log2FC levels of each miRNA
are reported in a graphic table indicating the level of
upregulation and downregulation, using red boxes and blue boxes,
respectively with different gradient.
Schemes of de-regulated miRNAs were generated for
the GBM datasets, AD datasets and for the GBM and AD datasets
together, in order to establish the existence of an inverse
association between the expression levels of GBM- and AD-related
miRNAs.
To establish the involvement of the identified
miRNAs in the modulation of glioma- and AD-related pathways and
their relative target genes, a pathway prediction analysis was
performed using the prediction tool DIANA-mirPath version 3
(69), as previously described
(70). The DIANA-mirPath analysis was
performed comparing the identified miRNAs with the panel of miRNAs
involved in both glioma- (hsa05214) and AD-related (hsa05010)
pathways.
The GO analysis and the functional roles of the
miRNAs-targeted genes related to GBM and AD were evaluated using
the enrichment software ‘STRING: Functional protein association
networks’ (https://string-db.org/) (71). In particular, for the genes related to
GBM and AD the ‘Biological process’, ‘Molecular function’ and
‘Cellular component’ were determined. Furthermore, the interaction
network between genes was determined for both GBM and AD. The
STRING analyses were performed for the 13 selected miRNAs resulting
from the comparison of GBM and AD miRNA expression profiling
datasets.
The GEO2R software already normalized the miRNA
expression data derived from GEO DataSets. Only miRNAs with a value
of P≤0.01 were considered for further analyses. Furthermore, the
GEO2R software automatically calculated the GEO DataSets data
P-values. The P-values obtained from the prediction pathway
analysis were already calculated using DIANA-mirPath software
(V.3.0).
The research of miRNA expression profiling datasets
performed with GEO2R using specific search terms allowed for the
identification of 51 and 9 miRNA profiling datasets for GBM and AD,
respectively. The datasets either relative to in vitro
studies or built with less than 8 samples, including normal and
pathological specimens, were excluded from the analysis. Following
this filtering, five datasets for GBM and two datasets for AD, were
selected for the study. The information of all selected datasets is
reported in Table I.
In particular, for GBM, two datasets were developed
by Affymetrix (Affymetrix miRNA Array), two developed by Illumina
(Illumina Human MicroRNA expression beadchip) and one developed by
Exiqon (Exiqon miRCURY LNA microRNA array, 7th generation). For the
AD datasets, one was developed by 3D-Gene (3D-Gene Human miRNA
V21_1.0.0) and the other one was a custom platform (USC/XJZ Human
0.9 K miRNA-940-v1.0) (Table I).
Merging the lists of differentially expressed miRNAs
in GBM datasets allowed the identification of a set of miRNAs
strictly involved in the development and progression of GBM. In
particular, among all miRNAs, 35 were found to be de-regulated with
concordant expression levels in at least 3 out of 5 GBM miRNA
expression datasets (downregulated or upregulated in all datasets)
(Fig. 1A). In particular, 14 miRNAs
were upregulated and 21 downregulated. Of these miRNAs, the most
upregulated miRNAs were the following: hsa-miR-21, hsa-miR-18a,
hsa-miR-19a, hsa-miR-25, hsa-miR-16-1, hsa-miR-106a and
hsa-miR-106b; and the most downregulated miRNAs were the following:
hsa-miR-128, hsa-miR-129, hsa-miR-7, hsa-miR-873, hsa-miR-218,
hsa-miR-139 and hsa-miR-770.
Similarly, following the merging of the 2 AD miRNA
lists, 7 miRNAs were uncovered. All of them were related to the
development of AD. Among these, 5 were upregulated (hsa-miR-134,
hsa-miR-185, hsa-miR-198, hsa-miR-659 and hsa-miR-671) and 2
downregulated (hsa-miR-29c and hsa-miR-494) (Fig. 1B).
By comparing the de-regulated miRNAs in both the GBM
and AD datasets, it was observed that the expression levels of 12
miRNAs were inversely associated between GBM and AD. Of these, 3
were upregulated in GBM and downregulated in AD (hsa-miR-106a,
hsa-miR-20b and hsa-miR-424) and 9 were downregulated in GBM and
upregulated in AD (hsa-miR-1224, hsa-miR-129, hsa-miR-139,
hsa-miR-330, hsa-miR-433, hsa-miR-485, hsa-miR-487b, hsa-miR-584
and hsa-miR-885). Additionally, hsa-miR-29c was downregulated in
both the GBM and AD datasets, suggesting its involvement in both
pathologies (Fig. 2).
To elucidate the role of the de-regulated miRNAs
shared between the GBM and AD datasets, DIANA-mirPath analysis was
performed. The analysis revealed that all the 13 selected miRNAs,
apart from hsa-miR-433, were involved in the regulation of GBM by
modulating the expression levels of 34 different genes (Fig. 3). As regards AD, the pathway
prediction analysis revealed that, with the exclusion of
hsa-miR-433, hsa-miR-485 and hsa-miR-487b, all the remaining
identified miRNAs were able to interact with the AD pathway and to
target 51 different genes. Notably, the analysis revealed that the
miRNAs, hsa-miR-106a, hsa-miR-424 and hsa-miR-330, were able to
modulate the expression levels of MAPK1 in both GBM and AD. In
particular, hsa-miR-106a and hsa-miR-424 were able to induce the
downregulation and upregulation of MAPK1 in GBM and AD,
respectively. This could be explained by the inverse association of
miR-106a expression levels in GBM and AD. Similarly, hsa-miR-330
was able to induce the upregulation of MAPK1 in GBM, while in AD,
this gene was downregulated. In the same manner, the miRNAs,
hsa-miR-424, hsa-miR-885 and hsa-miR-29c, were involved in the
regulation of several genes belonging to the family of calmodulins
(CALM family). In detail, hsa-miR-424 and hsa-miR-885 exhibited an
inverse association in modulating the expression of CALM genes,
while hsa-miR-29c was downregulated in both the GBM and AD
datasets, thus determining the upregulation of CALM3 in both
pathologies (Fig. 3).
In addition, the pathway prediction analysis
identified the most targeted genes in GBM as IGF1R (targeted by 6
miRNAs), CCND1, CDKN1A, MDM2 (targeted by 5 miRNAs), AKT3, CDK6,
E2F1, MAPK1, PIK3R1, PIK3R3 (targeted by 4 miRNAs). Notably, the
mostly targeted gene families were PI3K (targeted by 11 miRNAs),
CDK (targeted by 10 miRNAs) and E2F (targeted by 5 miRNAs) (data
not shown). As regards the AD pathway, the 13 selected miRNAs were
able to target mainly the MAPK1 (targeted by 4 miRNAs), APH1A, APP,
GSK3B (targeted by 3 miRNAs), ADAM17, ATP2A2, CALM2 and CALM3
(targeted by 2 miRNAs). Of note, the most altered gene families
were the ATP (9 miRNAs), NDUF (8 miRNAs), CALM (5 miRNAs) and ADAM
(3 miRNAs) families, strictly involved in the development of AD
(data not shown).
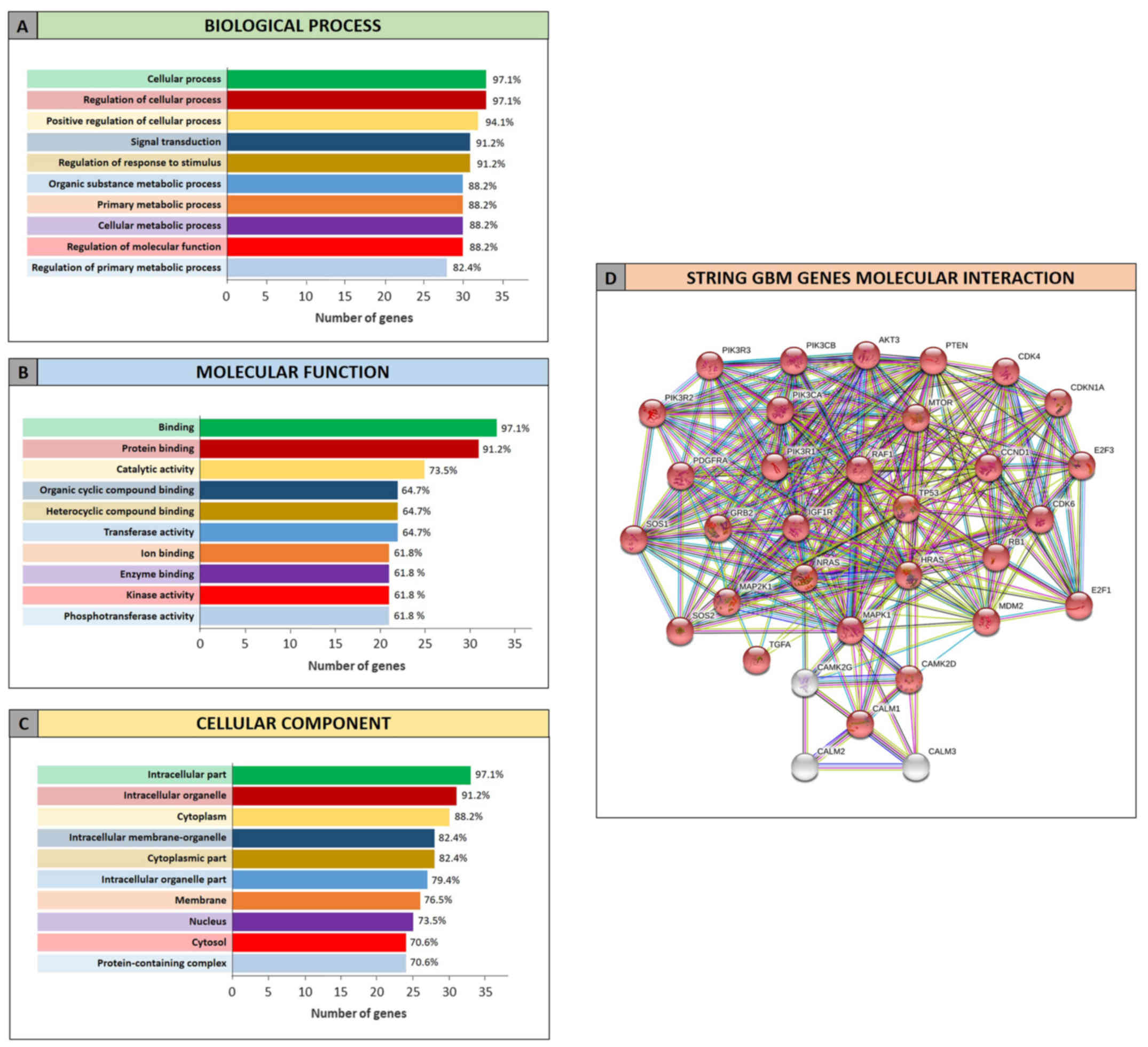
The GO analysis performed by STRING allowed to
determine the functional roles of the genes related to GBM and AD
and targeted by the selected miRNAs. The STRING analysis performed
on GBM miRNA-targeted genes revealed that of the 34 genes, 33 were
recognized as proteins. Among these 33 proteins, 30 were directly
involved in the glioma pathway (Fig.
4D). The clustering of proteins according to the ‘Biological
process’, ‘Molecular function’ and ‘Cellular component’ categories
revealed that the majority of proteins were involved in the
regulation of cellular processes and metabolic processes (Fig. 4A), in the binding of molecules and in
catalytic activities (Fig. 4B).
Moreover they belong to intracellular organelle and cytoplasm
(Fig. 4C).
Similarly, the STRING analysis performed on the list
of miRNA-targeted genes obtained for the AD highlighted that 47 of
the 51 identified genes were involved in AD pathway (Fig. 5D). As described for GBM, regarding the
‘Biological process’ category, the identified genes were involved
in the cellular and metabolic processes (metabolism of different
compounds), but also in the response to stimuli (Fig. 5A). The AD proteins were also
significantly involved in the binding of several molecules and less
in the catalytic activity, in contrast to what it was observed for
the GBM proteins clustered in the ‘Molecular function’ category
(Fig. 5B). Finally, the analyzed
proteins were part of the intracellular organelle, as well as the
cell membrane, therefore playing a fundamental role in the
regulation of cellular homeostasis (Fig.
5C).
Despite tremendous advancements being made in the
characterization of the clinicopathological features typical of
tumors and neurodegenerative diseases, little is known about the
association between the molecular mechanisms responsible for the
development and progression of brain cancer and neurodegeneration
(72–74). In detail, an indirect association in
the incidence rates of GBM and AD has been widely reported, thus
suggesting that this inverse association may be coupled with an
inverse regulation of the same molecular mechanisms involved in the
development of GBM and AD (29,30).
To shed light on the molecular mechanisms
potentially responsible for the rise of these pathologies, and to
demonstrate the existence of an inverse association between the
molecular alterations in GBM and AD, the present study identified
the miRNAs found altered in GBM and AD. In particular, in this
study, we analyzed the potential involvement of these miRNAs in the
onset of both diseases although oppositely expressed. For the first
time, at least to the best of our knowledge, the existence of a
strong inverse association was demonstrated in human samples
between selected miRNA expression levels in GBM and AD through the
comparison of altered miRNAs.
The choice of analyzing the expression levels of
miRNAs in both pathologies depends on the numerous profiling data
collected during the years for GBM and AD, and the increasing
number of studies coupling the analysis of miRNAs in cancer and
neurodegenerative disorders (75–78).
The independent analysis of GBM miRNA expression
datasets revealed that 35 miRNAs were de-regulated in tumor samples
compared to the normal controls. Almost all of these de-regulated
miRNAs have been widely associated with the development and
progression of GBM. In particular, several studies have already
described the role of hsa-miR-21 and hsa-miR-155 overexpression in
the promotion of the development of glioblastoma (79–81). Other
studies have demonstrated the therapeutic potential of the
overexpression of several miRNAs normally downregulated in
glioblastoma, such as hsa-miR-7, hsa-miR-93 and hsa-miR-139
(82–84). As regards the most downregulated
miRNAs, hsa-miR-128 and hsa-miR-129, found in the GBM dataset
analysis, some studies have highlighted their possible use in the
context of novel therapeutic strategies aimed at inhibiting the
molecular pathways involved in GBM aggressive phenotypes (85–87).
Concerning the analysis performed on the 2 AD miRNA
datasets, the results revealed significant data for the
downregulated miRNA, hsa-miR-29c. Several studies have demonstrated
that the downregulation of this miRNA may play a potential role
either as biomarker or as therapeutic agent in AD in vitro
models and in patients (88–91). Of note, the downregulation of
hsa-miR-29c has also been observed in GBM where it was observed
that its induced overexpression led to the suppression of glioma
(92).
However, the most robust and interesting data
derived from the comparison between miRNAs de-regulated in both the
GBM AD datasets, was that all miRNAs identified, with the exception
of hsa-miR-29c, exhibited inverse patterns of expression in GBM
compared with AD. This result is in accordance with the hypothesis
of the existence of an inverse regulation of molecular pathways in
GBM and AD, as postulated by Liu et al (30) and Sánchez-Valle et al (29). In line with this theory, miRNAs were
found inversely regulated in the two pathologies by our analyses.
Noteworthy, the inverse regulation of miRNAs in GBM and AD is
coupled with the inverse regulation of targeted genes by selected
miRNAs. Such inverse miRNAs and genes regulatory patterns may
explain the inverse comorbidity existing between neurological
disorders and cancers (6).
The preliminary data obtained comparing miRNA
expression levels in GBM and AD were further confirmed by the
pathway prediction and gene ontology enrichment analyses. In
particular, the DIANA-mirPath analysis revealed that the 13
selected miRNAs were in common between GBM and AD and they were
able to modulate several genes within the glioma pathway
(hsa05214). In particular, the miRNAs were involved in the
regulation of genes, such as MAPK1, IGF1R, and genes belonging to
the PIK3 and RAS families, known to be involved in the development
of GBM and other tumors (93–97).
Conversely, the same miRNAs were shown to also
target fundamental genes involved in AD, such as APP, responsible
for the β-amyloid plaque formation (98); GSK3B, hyperactivated in Alzheimer's
neurons (99); NDUF family,
responsible for mitochondrial alterations in AD (100); LRP1 able to regulate the metabolism
of amyloid-β peptides thus maintaining brain homeostasis (101) and other genes.
Therefore, it is clear how miRNA de-regulation and
other epigenetic mechanisms may lead to the modulation of these
genes and, in turn, to the acquisition of a more aggressive tumor
phenotype or neurodegenerative disorder susceptibility (102,103).
In addition, STRING analysis revealed that the
selected miRNAs were able to modulate genes involved in both GBM
and AD and those genes performed, in general, the same function and
processes within the two pathologies. Therefore, differential
expression levels of miRNAs in GBM and AD may be responsible for
the onset of GBM rather than the AD, and vice versa.
Overall, the identification of altered miRNAs in
both GBM and AD, as well as the definition of the inverse patterns
of expression, may pave the way for new studies to better elucidate
the involvement of these miRNAs in GBM and AD. In particular,
further studies are required to examine the therapeutic potential
of such identified miRNAs at the early stages of disease, since
several treatments for AD are administered only when the pathology
is at an advanced stage (104–106).
On this matter, the results of the present study and the potential
applications of this research methodology also to other brain
diseases, may improve the diagnostic and therapeutic strategies
mainly based on the analysis of low-specific biomarkers and on the
use of low-sensitive and low-efficacy instrumental procedures
(107–111).
In conclusion, in the present study, at least to the
best of our knowledge, for the first time a set of de-regulated
miRNAs in both GBM and AD was identified, demonstrating the
existence of an inverse association between the expression levels
of miRNAs in GBM and AD. The findings of the present study may pave
the way for other functional in vitro and in vivo
studies to validate the diagnostic or prognostic significance of
the identified miRNAs, as well as to depict their possible use as
novel therapeutic approaches for GBM and AD.
The authors would like to thank the Italian League
Against Cancer (LILT) for its support.
No funding was received.
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request. The analyzed datasets
are publicly available on the GEO DataSet database.
LF, DAS and ML conceived and designed the study. SC,
LF, MP and GG performed all the analyses. GL, CDA and MSB provided
all the information useful to describe the features of Glioblastoma
Multiforme, while MP was involved in the analysis of AD datasets
and gave information about neurodegenerative disorders and
Alzheimer's disease. MP, MCP and SV were involved in the pathway
prediction analysis and in the definition of the functional roles
of selected miRNAs. LF, SC, MCP, MP and SV were involved in the
interpretation of all data. GG, MCP and LF were involved in the
preparation of the figures and tables. SV, LF and ML were involved
in the preparation of the original draft of the manuscript, while
MCP, SC, SV, GB, CDA and MP reviewed and edited the article. All
authors have read and approved the final version of the
manuscript.
All patient data were derived from publicly
available datasets.
Not applicable.
DAS is the Editor-in-Chief for the journal, but had
no influence in the reviewing process, or any involvement in terms
of adjudicating on the final decision, for this article. The other
authors declare that there are not competing interests.
|
1
|
Crimmins EM and Zhang YS: Aging
populations, mortality, and life expectancy. Annu Rev Sociol.
45:annurev–soc-073117-041351. 2019. View Article : Google Scholar
|
|
2
|
Franceschi C, Garagnani P, Morsiani C,
Conte M, Santoro A, Grignolio A, Monti D, Capri M and Salvioli S:
The Continuum of Aging and Age-Related Diseases: Common Mechanisms
but Different Rates. Front Med (Lausanne). 5:612018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ganguli M: Cancer and dementia: It's
complicated. Alzheimer Dis Assoc Disord. 29:177–182.
2015.PubMed/NCBI
|
|
4
|
Behrens MI, Lendon C and Roe CM: A common
biological mechanism in cancer and Alzheimer's disease? Curr
Alzheimer Res. 6:196–204. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Snyder HM, Ahles T, Calderwood S, Carrillo
MC, Chen H, Chang CH, Craft S, De Jager P, Driver JA, Fillit H, et
al: Exploring the nexus of Alzheimer's disease and related
dementias with cancer and cancer therapies: A convening of the
Alzheimer's Association & Alzheimer's Drug Discovery
Foundation. Alzheimers Dement. 13:267–273. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ibáñez K, Boullosa C, Tabarés-Seisdedos R,
Baudot A and Valencia A: Molecular evidence for the inverse
comorbidity between central nervous system disorders and cancers
detected by transcriptomic meta-analyses. PLoS Genet.
10:e10041732014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Trovato Salinaro A, Pennisi M, Di Paola R,
Scuto M, Crupi R, Cambria MT, Ontario ML, Tomasello M, Uva M,
Maiolino L, et al: Neuroinflammation and neurohormesis in the
pathogenesis of Alzheimer's disease and Alzheimer-linked
pathologies: Modulation by nutritional mushrooms. Immun Ageing.
15:82018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pennisi M, Crupi R, Di Paola R, Ontario
ML, Bella R, Calabrese EJ, Crea R, Cuzzocrea S and Calabrese V:
Inflammasomes, hormesis, and antioxidants in neuroinflammation:
Role of NRLP3 in Alzheimer disease. J Neurosci Res. 95:1360–1372.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cornelius C, Trovato Salinaro A, Scuto M,
Fronte V, Cambria MT, Pennisi M, Bella R, Milone P, Graziano A,
Crupi R, et al: Cellular stress response, sirtuins and UCP proteins
in Alzheimer disease: Role of vitagenes. Immun Ageing. 10:412013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vivarelli S, Falzone L, Basile MS,
Nicolosi D, Genovese C, Libra M and Salmeri M: Benefits of using
probiotics as adjuvants in anticancer therapy (Review). World J
Acad Sci June. 1:125–135. 2019.
|
|
11
|
Vivarelli S, Salemi R, Candido S, Falzone
L, Santagati M, Stefani S, Torino F, Banna GL, Tonini G and Libra
M: Gut microbiota and cancer: From pathogenesis to therapy. Cancers
(Basel). 11:E382019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang T, Han Y, Wang J, Hou D, Deng H,
Deng YL and Song Z: Comparative epidemiological Investigation of
Alzheimer's disease and colorectal cancer: The possible role of
gastrointestinal conditions in the pathogenesis of AD. Front Aging
Neurosci. 10:1762018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Banna GL, Torino F, Marletta F, Santagati
M, Salemi R, Cannarozzo E, Falzone L, Ferraù F and Libra M:
Lactobacillus rhamnosus GG: An overview to explore the
rationale of its use in cancer. Front Pharmacol. 8:6032017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lanza G, Bella R, Cantone M, Pennisi G,
Ferri R and Pennisi M: Cognitive impairment and celiac disease: Is
transcranial magnetic stimulation a trait d'Union between gut and
brain? Int J Mol Sci. 19:E22432018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fotakopoulos G, Brotis AG, Kotlia P and
Fountas K: Glioblastoma multiforme in a patient with celiac
disease: Management of seizures after gross total tumor resection.
World Neurosurg. 118:209–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pennisi M, Bramanti A, Cantone M, Pennisi
G, Bella R and Lanza G: Neurophysiology of the ‘Celiac Brain’:
Disentangling gut-brain connections. Front Neurosci. 11:4982017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bella R, Lanza G, Cantone M, Giuffrida S,
Puglisi V, Vinciguerra L, Pennisi M, Ricceri R, D'Agate CC,
Malaguarnera G, et al: Effect of a gluten-free diet on cortical
excitability in adults with celiac disease. PLoS One.
10:e01292182015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pennisi G, Lanza G, Giuffrida S,
Vinciguerra L, Puglisi V, Cantone M, Pennisi M, D'Agate CC, Naso P,
Aprile G, et al: Excitability of the motor cortex in de novo
patients with celiac disease. PLoS One. 9:e1027902014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rapisarda V, Ledda C, Matera S, Fago L,
Arrabito G, Falzone L, Marconi A, Libra M and Loreto C: Absence of
t(14;18) chromosome translocation in agricultural workers after
short-term exposure to pesticides. Mol Med Rep. 15:3379–3382. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garozzo A, Falzone L, Rapisarda V, Marconi
A, Cinà D, Fenga C, Spandidos DA and Libra M: The risk of HCV
infection among health-care workers and its association with
extrahepatic manifestations (Review). Mol Med Rep. 15:3336–3339.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fenga C, Gangemi S, Di Salvatore V,
Falzone L and Libra M: Falzone L amd Libra M: Immunological effects
of occupational exposure to lead (Review). Mol Med Rep.
15:3355–3360. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rapisarda V, Salemi R, Marconi A, Loreto
C, Graziano AC, Cardile V, Basile MS, Candido S, Falzone L,
Spandidos DA, et al: Fluoro-edenite induces fibulin-3
overexpression in non-malignant human mesothelial cells. Oncol
Lett. 12:3363–3367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Falzone L, Marconi A, Loreto C, Franco S,
Spandidos DA and Libra M: Occupational exposure to carcinogens:
Benzene, pesticides and fibers (Review). Mol Med Rep. 14:4467–4474.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pennisi M, Malaguarnera G, Puglisi V,
Vinciguerra L, Vacante M and Malaguarnera M: Neurotoxicity of
acrylamide in exposed workers. Int J Environ Res Public Health.
10:3843–3854. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alessandria I, Pennisi M, Cataudella E,
Frazzetto PM, Malaguarnera M and Rampello L and Rampello L:
Neurotoxicity in cadmium-exposed workers. Acta Med Mediter.
28:253–526. 2012.
|
|
26
|
Pennisi M, Lanza G, Cantone M, Schepis C,
Ferri R, Barone R and Bella R: Unusual neurological presentation of
nevoid basal cell carcinoma syndrome (Gorlin-Goltz syndrome). J
Clin Neurol. 13:439–441. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cantone M, Lanza G, Pennisi M, Bella R,
Schepis C, Siragusa M, Barone R and Ferri R: Prominent neurological
involvement in Dercum disease. J Neurol. 264:796–798. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uccello M, Vacante M, Giordano M,
Malaguarnera M, Biondi A, Basile F, Malaguarnera G, Pennisi M and
Motta M: Osteoblastoma of cervical spine causing an unusual neck
pain. Eur Rev Med Pharmacol Sci. 16 (Suppl 4):17–20.
2012.PubMed/NCBI
|
|
29
|
Sánchez-Valle J, Tejero H, Ibáñez K,
Portero JL, Krallinger M, Al-Shahrour F, Tabarés-Seisdedos R,
Baudot A and Valencia A: A molecular hypothesis to explain direct
and inverse co-morbidities between Alzheimer's disease,
glioblastoma and lung cancer. Sci Rep. 7:44742017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu T, Ren D, Zhu X, Yin Z, Jin G, Zhao Z,
Robinson D, Li X, Wong K, Cui K, et al: Transcriptional signaling
pathways inversely regulated in Alzheimer's disease and
glioblastoma multiform. Sci Rep. 3:34672013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lehrer S: Glioma and Alzheimer's disease.
J Alzheimers Dis Rep. 2:213–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Roe CM, Behrens MI, Xiong C, Miller JP and
Morris JC: Alzheimer disease and cancer. Neurology. 64:895–898.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Parsons CG, Danysz W, Dekundy A and Pulte
I: Memantine and cholinesterase inhibitors: Complementary
mechanisms in the treatment of Alzheimer's disease. Neurotox Res.
24:358–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hanif F, Muzaffar K, Perveen K, Malhi SM
and Simjee ShU: Glioblastoma multiforme: A review of its
epidemiology and pathogenesis through clinical presentation and
treatment. Asian Pac J Cancer Prev. 18:3–9. 2017.PubMed/NCBI
|
|
35
|
Davis ME: Glioblastoma: Overview of
Disease and Treatment. Clin J Oncol Nurs. 20 (Suppl 5):S2–S8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alphandéry E: Glioblastoma Treatments: An
Account of Recent Industrial Developments. Front Pharmacol.
9:8792018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Falzone L, Salomone S and Libra M:
Evolution of cancer pharmacological treatments at the turn of the
third millennium. Front Pharmacol. 9:13002018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Paskaš S, Krajnović T, Basile MS,
Dunđerović D, Cavalli E, Mangano K, Mammana S, Al-Abed Y, Nicoletti
F, Mijatović S, et al: Senescence as a main mechanism of Ritonavir
and Ritonavir-NO action against melanoma. Mol Carcinog. Apr
17–2019.(Epub ahead of print). doi: 10.1002/mc.23020. View Article : Google Scholar
|
|
39
|
Paskas S, Mazzon E, Basile MS, Cavalli E,
Al-Abed Y, He M, Rakocevic S, Nicoletti F, Mijatovic S and
Maksimovic-Ivanic D: Lopinavir-NO, a nitric oxide-releasing HIV
protease inhibitor, suppresses the growth of melanoma cells in
vitro and in vivo. Invest New Drugs. Feb 1–2019.(Epub ahead of
print). doi: 10.1007/s10637-019-00733-3. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Basile MS, Mazzon E, Krajnovic T, Draca D,
Cavalli E, Al-Abed Y, Bramanti P, Nicoletti F, Mijatovic S and
Maksimovic-Ivanic D: Anticancer and differentiation properties of
the nitric oxide derivative of lopinavir in human glioblastoma
cells. Molecules. 23:E24632018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lazarević M, Mazzon E, Momčilović M,
Basile MS, Colletti G, Petralia MC, Bramanti P, Nicoletti F and
Miljković Đ: The H2S donor GYY4137 stimulates reactive
oxygen species generation in BV2 cells while suppressing the
secretion of TNF and nitric oxide. Molecules. 23:E29662018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vinciguerra L, Lanza G, Puglisi V, Pennisi
M, Cantone M, Bramanti A, Pennisi G and Bella R: Transcranial
Doppler ultrasound in vascular cognitive impairment-no dementia.
PLoS One. 14:e02161622019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Puglisi V, Bramanti A, Lanza G, Cantone M,
Vinciguerra L, Pennisi M, Bonanno L, Pennisi G and Bella R:
Impaired cerebral haemodynamics in vascular depression: Insights
from transcranial doppler ultrasonography. Front Psychiatry.
9:3162018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lanza G, Bramanti P, Cantone M, Pennisi M,
Pennisi G and Bella R: Vascular cognitive impairment through the
looking glass of transcranial magnetic stimulation. Behav Neurol.
2017:14213262017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pennisi M, Lanza G, Cantone M, Ricceri R,
Spampinato C, Pennisi G, Di Lazzaro V and Bella R: Correlation
between motor cortex excitability changes and cognitive impairment
in vascular depression: Pathophysiological insights from a
longitudinal TMS study. Neural Plast. 2016:81549692016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Concerto C, Lanza G, Cantone M, Pennisi M,
Giordano D, Spampinato C, Ricceri R, Pennisi G, Aguglia E and Bella
R: Different patterns of cortical excitability in major depression
and vascular depression: A transcranial magnetic stimulation study.
BMC Psychiatry. 13:3002013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bella R, Ferri R, Lanza G, Cantone M,
Pennisi M, Puglisi V, Vinciguerra L, Spampinato C, Mazza T,
Malaguarnera G, et al: TMS follow-up study in patients with
vascular cognitive impairment-no dementia. Neurosci Lett.
534:155–159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Pennisi G, Ferri R, Lanza G, Cantone M,
Pennisi M, Puglisi V, Malaguarnera G and Bella R: Transcranial
magnetic stimulation in Alzheimer's disease: A neurophysiological
marker of cortical hyperexcitability. J Neural Transm (Vienna).
118:587–598. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lanza G, Lanuzza B, Aricò D, Cantone M,
Cosentino FII, Bella R, Pennisi G, Ferri R and Pennisi M: Impaired
short-term plasticity in restless legs syndrome: A pilot rTMS
study. Sleep Med. 46:1–4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lanza G, Cantone M, Aricò D, Lanuzza B,
Cosentino FII, Paci D, Papotto M, Pennisi M, Bella R, Pennisi G, et
al: Clinical and electrophysiological impact of repetitive
low-frequency transcranial magnetic stimulation on the
sensory-motor network in patients with restless legs syndrome. Ther
Adv Neurol Disorder. 11:17562864187599732018.
|
|
51
|
Lanza G, Cantone M, Lanuzza B, Pennisi M,
Bella R, Pennisi G and Ferri R: Distinctive patterns of cortical
excitability to transcranial magnetic stimulation in obstructive
sleep apnea syndrome, restless legs syndrome, insomnia, and sleep
deprivation. Sleep Med Rev. 19:39–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lanza G, Lanuzza B, Aricò D, Cantone M,
Cosentino FI, Pennisi M, Bella R, Pennisi G and Ferri R: Direct
comparison of cortical excitability to transcranial magnetic
stimulation in obstructive sleep apnea syndrome and restless legs
syndrome. Sleep Med. 16:138–142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cantone M, Bramanti A, Lanza G, Pennisi M,
Bramanti P, Pennisi G and Bella R: Cortical Plasticity in
Depression. ASN Neuro. 9:17590914177115122017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pennisi M, Lanza G, Cantone M, Ricceri R,
Ferri R, D'Agate CC, Pennisi G, Di Lazzaro V and Bella R: Cortical
involvement in celiac disease before and after long-term
gluten-free diet: A Transcranial Magnetic Stimulation study. PLoS
One. 12:e01775602017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lanza G, Bella R, Giuffrida S, Cantone M,
Pennisi G, Spampinato C, Giordano D, Malaguarnera G, Raggi A and
Pennisi M: Preserved transcallosal inhibition to transcranial
magnetic stimulation in nondemented elderly patients with
leukoaraiosis. BioMed Res Int. 2013:3516802013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Spampinato C, Aguglia E, Concerto C,
Pennisi M, Lanza G, Bella R, Cantone M, Pennisi G, Kavasidis I and
Giordano D: Transcranial magnetic stimulation in the assessment of
motor cortex excitability and treatment of drug-resistant major
depression. IEEE Trans Neural Syst Rehabil Eng. 21:391–403. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Presti M, Mazzon E, Basile MS, Petralia
MC, Bramanti A, Colletti G, Bramanti P, Nicoletti F and Fagone P:
Overexpression of macrophage migration inhibitory factor and
functionally-related genes, D-DT, CD74, CD44, CXCR2 and CXCR4, in
glioblastoma. Oncol Lett. 16:2881–2886. 2018.PubMed/NCBI
|
|
58
|
Mammana S, Fagone P, Cavalli E, Basile MS,
Petralia MC, Nicoletti F, Bramanti P and Mazzon E: The role of
macrophages in neuroinflammatory and neurodegenerative pathways of
Alzheimer's disease, amyotrophic lateral sclerosis, and multiple
sclerosis: Pathogenetic cellular effectors and potential
therapeutic targets. Int J Mol Sci. 19:E8312018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mangano K, Mazzon E, Basile MS, Di Marco
R, Bramanti P, Mammana S, Petralia MC, Fagone P and Nicoletti F:
Pathogenic role for macrophage migration inhibitory factor in
glioblastoma and its targeting with specific inhibitors as novel
tailored therapeutic approach. Oncotarget. 9:17951–17970. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Nicoletti F, Mazzon E, Fagone P, Mangano
K, Mammana S, Cavalli E, Basile MS, Bramanti P, Scalabrino G, Lange
A, et al: Prevention of clinical and histological signs of
MOG-induced experimental allergic encephalomyelitis by prolonged
treatment with recombinant human EGF. J Neuroimmunol. 332:224–232.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ludwig K and Kornblum HI: Molecular
markers in glioma. J Neurooncol. 134:505–512. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Maniati MS, Maniati M, Yousefi T,
Ahmadi-Ahangar A and Tehrani SS: New insights into the role of
microRNAs and long noncoding RNAs in most common neurodegenerative
diseases. J Cell Biochem. 120:8908–8918. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Battaglia R, Palini S, Vento ME, La
Ferlita A, Lo Faro MJ, Caroppo E, Borzì P, Falzone L, Barbagallo D,
Ragusa M, et al: Identification of extracellular vesicles and
characterization of miRNA expression profiles in human blastocoel
fluid. Sci Rep. 9:842019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Falzone L, Lupo G, Rosa GRM, Crimi S,
Anfuso CD, Salemi R, Rapisarda E, Libra M and Candido S:
Identification of novel MicroRNAs and their diagnostic and
prognostic significance in oral cancer. Cancers (Basel).
11:E6102019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hafsi S, Candido S, Maestro R, Falzone L,
Soua Z, Bonavida B, Spandidos DA and Libra M: Correlation between
the overexpression of Yin Yang 1 and the expression levels of
miRNAs in Burkitt's lymphoma: A computational study. Oncol Lett.
11:1021–1025. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Falzone L, Scola L, Zanghì A, Biondi A, Di
Cataldo A, Libra M and Candido S: Integrated analysis of colorectal
cancer microRNA datasets: Identification of microRNAs associated
with tumor development. Aging (Albany NY). 10:1000–1014. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Falzone L, Candido S, Salemi R, Basile MS,
Scalisi A, McCubrey JA, Torino F, Signorelli SS, Montella M and
Libra M: Computational identification of microRNAs associated to
both epithelial to mesenchymal transition and NGAL/MMP-9 pathways
in bladder cancer. Oncotarget. 7:72758–72766. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kozomara A, Birgaoanu M and
Griffiths-Jones S: miRBase: From microRNA sequences to function.
Nucleic Acids Res. 47:D155–D162. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Vlachos IS, Zagganas K, Paraskevopoulou
MD, Georgakilas G, Karagkouni D, Vergoulis T, Dalamagas T and
Hatzigeorgiou AG: DIANA-miRPath v3.0: Deciphering microRNA function
with experimental support. Nucleic Acids Res. 43:W460–W466. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Falzone L, Romano GL, Salemi R, Bucolo C,
Tomasello B, Lupo G, Anfuso CD, Spandidos DA, Libra M and Candido
S: Prognostic significance of deregulated microRNAs in uveal
melanomas. Mol Med Rep. 19:2599–2610. 2019.PubMed/NCBI
|
|
71
|
Szklarczyk D, Gable AL, Lyon D, Junge A,
Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork
P, et al: STRING v11: Protein-protein association networks with
increased coverage, supporting functional discovery in genome-wide
experimental datasets. Nucleic Acids Res. 47:D607–D613. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Nixon DW: The inverse relationship between
cancer and Alzheimer's disease: A possible mechanism. Curr
Alzheimer Res. 14:883–893. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Shafi O: Inverse relationship between
Alzheimer's disease and cancer, and other factors contributing to
Alzheimer's disease: A systematic review. BMC Neurol. 16:2362016.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Park S, Yu SJ, Cho Y, Balch C, Lee J, Kim
YH and Nam S: Network comparison of inflammation in colorectal
cancer and Alzheimer's disease. BioMed Res Int. 2015:2052472015.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Godlewski J, Lenart J and Salinska E:
MicroRNA in brain pathology: Neurodegeneration the other side of
the brain cancer. Noncoding RNA. 5:E202019.PubMed/NCBI
|
|
76
|
Ham S, Kim TK, Ryu J, Kim YS, Tang YP and
Im HI: Comprehensive MicroRNAome analysis of the relationship
between Alzheimer disease and cancer in PSEN double-knockout mice.
Int Neurourol J. 22:237–245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Polo A, Crispo A, Cerino P, Falzone L,
Candido S, Giudice A, De Petro G, Ciliberto G, Montella M, Budillon
A, et al: Environment and bladder cancer: Molecular analysis by
interaction networks. Oncotarget. 8:65240–65252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
McCubrey JA, Fitzgerald TL, Yang LV,
Lertpiriyapong K, Steelman LS, Abrams SL, Montalto G, Cervello M,
Neri LM, Cocco L, et al: Roles of GSK-3 and microRNAs on epithelial
mesenchymal transition and cancer stem cells. Oncotarget.
8:14221–14250. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Jesionek-Kupnicka D, Braun M,
Trąbska-Kluch B, Czech J, Szybka M, Szymańska B, Kulczycka-Wojdala
D, Bieńkowski M, Kordek R and Zawlik I: MiR-21, miR-34a, miR-125b,
miR-181d and miR-648 levels inversely correlate with MGMT and TP53
expression in primary glioblastoma patients. Arch Med Sci.
15:504–512. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
ParvizHamidi M, Haddad G, Ostadrahimi S,
Ostadrahimi N, Sadeghi S, Fayaz S and Fard-Esfahani P: Circulating
miR-26a and miR-21 as biomarkers for glioblastoma multiform.
Biotechnol Appl Biochem. 66:261–265. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wu W, Yu T, Wu Y, Tian W, Zhang J and Wang
Y: The miR155HG/miR-185/ANXA2 loop contributes to glioblastoma
growth and progression. J Exp Clin Cancer Res. 38:1332019.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Yin CY, Kong W, Jiang J, Xu H and Zhao W:
miR-7-5p inhibits cell migration and invasion in glioblastoma
through targeting SATB1. Oncol Lett. 17:1819–1825. 2019.PubMed/NCBI
|
|
83
|
Choi JY, Shin HJ and Bae IH: miR-93-5p
suppresses cellular senescence by directly targeting Bcl-w and p21.
Biochem Biophys Res Commun. 505:1134–1140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Shi L, Yuan Y and Li HY: MicroRNA-139-3p
suppresses growth and metastasis of glioblastoma via inhibition of
NIN1/RPNI2 binding protein 1 homolog. Eur Rev Med Pharmacol Sci.
23:4264–4274. 2019.PubMed/NCBI
|
|
85
|
Zeng A, Yin J, Li Y, Li R, Wang Z, Zhou X,
Jin X, Shen F, Yan W and You Y: miR-129-5p targets Wnt5a to block
PKC/ERK/NF-κB and JNK pathways in glioblastoma. Cell Death Dis.
9:3942018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Xu H, Hu Y and Qiu W: Potential mechanisms
of microRNA-129-5p in inhibiting cell processes including
viability, proliferation, migration and invasiveness of
glioblastoma cells U87 through targeting FNDC3B. Biomed
Pharmacother. 87:405–411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Shan ZN, Tian R, Zhang M, Gui ZH, Wu J,
Ding M, Zhou XF and He J: miR128-1 inhibits the growth of
glioblastoma multiforme and glioma stem-like cells via targeting
BMI1 and E2F3. Oncotarget. 7:78813–78826. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wu Y, Xu J, Xu J, Cheng J, Jiao D, Zhou C,
Dai Y and Chen Q: Lower serum levels of miR-29c-3p and miR-19b-3p
as biomarkers for Alzheimer's disease. Tohoku J Exp Med.
242:129–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yang G, Song Y, Zhou X, Deng Y, Liu T,
Weng G, Yu D and Pan S: MicroRNA-29c targets β-site amyloid
precursor protein-cleaving enzyme 1 and has a neuroprotective role
in vitro and in vivo. Mol Med Rep. 12:3081–3088.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Lei X, Lei L, Zhang Z, Zhang Z and Cheng
Y: Downregulated miR-29c correlates with increased BACE1 expression
in sporadic Alzheimer's disease. Int J Clin Exp Pathol.
8:1565–1574. 2015.PubMed/NCBI
|
|
91
|
Zong Y, Yu P, Cheng H, Wang H, Wang X,
Liang C, Zhu H, Qin Y and Qin C: miR-29c regulates NAV3 protein
expression in a transgenic mouse model of Alzheimer's disease.
Brain Res. 1624:95–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang Y, Li Y, Sun J, Wang Q, Sun C, Yan Y,
Yu L, Cheng D, An T, Shi C, et al: Tumor-suppressive effects of
miR-29c on gliomas. Neuroreport. 24:637–645. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Song K, Yuan Y, Lin Y, Wang YX, Zhou J,
Gai QJ, Zhang L, Mao M, Yao XX, Qin Y, et al: ERBB3, IGF1R, and
TGFBR2 expression correlate with PDGFR expression in glioblastoma
and participate in PDGFR inhibitor resistance of glioblastoma
cells. Am J Cancer Res. 8:792–809. 2018.PubMed/NCBI
|
|
94
|
Salemi R, Falzone L, Madonna G, Polesel J,
Cinà D, Mallardo D, Ascierto PA, Libra M and Candido S: MMP-9 as a
candidate marker of response to BRAF inhibitors in melanoma
patients with BRAFV600E mutation detected in circulating-free DNA.
Front Pharmacol. 9:8562018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Leonardi GC, Falzone L, Salemi R, Zanghì
A, Spandidos DA, Mccubrey JA, Candido S and Libra M: Cutaneous
melanoma: From pathogenesis to therapy (Review). Int J Oncol.
52:1071–1080. 2018.PubMed/NCBI
|
|
96
|
Guarneri C, Bevelacqua V, Polesel J,
Falzone L, Cannavò PS, Spandidos DA, Malaponte G and Libra M: NF-κB
inhibition is associated with OPN/MMP 9 downregulation in cutaneous
melanoma. Oncol Rep. 37:737–746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Weber GL, Parat MO, Binder ZA, Gallia GL
and Riggins GJ: Abrogation of PIK3CA or PIK3R1 reduces
proliferation, migration, and invasion in glioblastoma multiforme
cells. Oncotarget. 2:833–849. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hardy J: The discovery of
Alzheimer-causing mutations in the APP gene and the formulation of
the ‘amyloid cascade hypothesis’. FEBS J. 284:1040–1044. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ochalek A, Mihalik B, Avci HX,
Chandrasekaran A, Téglási A, Bock I, Giudice ML, Táncos Z, Molnár
K, László L, et al: Neurons derived from sporadic Alzheimer's
disease iPSCs reveal elevated TAU hyperphosphorylation, increased
amyloid levels, and GSK3B activation. Alzheimers Res Ther.
9:902017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Cadonic C, Sabbir MG and Albensi BC:
Mechanisms of mitochondrial dysfunction in Alzheimer's disease. Mol
Neurobiol. 53:6078–6090. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Shinohara M, Tachibana M, Kanekiyo T and
Bu G: Role of LRP1 in the pathogenesis of Alzheimer's disease:
Evidence from clinical and preclinical studies. J Lipid Res.
58:1267–1281. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Falzone L, Salemi R, Travali S, Scalisi A,
McCubrey JA, Candido S and Libra M: MMP-9 overexpression is
associated with intragenic hypermethylation of MMP9 gene in
melanoma. Aging (Albany NY). 8:933–944. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Mastroeni D, Grover A, Delvaux E,
Whiteside C, Coleman PD and Rogers J: Epigenetic mechanisms in
Alzheimer's disease. Neurobiol Aging. 32:1161–1180. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang Z, He C and Shi JS: Natural products
for the treatment of neurodegenerative diseases. Curr Med Chem. May
27–2019.(Epub ahead of print). doi:
10.2174/0929867326666190527120614. View Article : Google Scholar :
|
|
105
|
Lanza G, Centonze SS, Destro G, Vella V,
Bellomo M, Pennisi M, Bella R and Ciavardelli D: Comment on
‘Shiatsu as an adjuvant therapy for depression in patients with
Alzheimer's disease: A Pilot Study’. J Evid Based Integr Med.
24:2515690X188251052019. View Article : Google Scholar :
|
|
106
|
Lanza G, Centonze SS, Destro G, Vella V,
Bellomo M, Pennisi M, Bella R and Ciavardelli D: Shiatsu as an
adjuvant therapy for depression in patients with Alzheimer's
disease: A pilot study. Complement Ther Med. 38:74–78. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Calabrese V, Dattilo S, Petralia A,
Parenti R, Pennisi M, Koverech G, Calabrese V, Graziano A, Monte I,
Maiolino L, et al: Analytical approaches to the diagnosis and
treatment of aging and aging-related disease: Redox status and
proteomics. Free Radic Res. 49:511–524. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Giordano D, Kavasidis I, Spampinato C,
Bella R, Pennisi G and Pennisi M: An integrated computer-controlled
system for assisting researchers in cortical excitability studies
by using transcranial magnetic stimulation. Comput Methods Programs
Biomed. 107:4–15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Pennisi G, Ferri R, Cantone M, Lanza G,
Pennisi M, Vinciguerra L, Malaguarnera G and Bella R: A review of
transcranial magnetic stimulation in vascular dementia. Dement
Geriatr Cogn Disord. 31:71–80. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Faro A, Giordano D, Kavasidis I, Pino C,
Spampinato C, Cantone GM, Lanza G and Pennisi M: An Interactive
Tool for Customizing Clinical Transacranial Magnetic Stimulation
(TMS) Experiments. XII Mediterranean Conference on Medical and
Biological Engineering and Computing. 200–203. 2010.
|
|
111
|
Faro A, Giordano D, Pennisi M, Scarciofalo
G, Spampinato C and Tramontana F: Transcranial Magnetic Stimulation
(TMS) to Evaluate and Classify Mental Diseases Using Neural
Networks. Conference on Artificial Intelligence in Medicine in
Europe AIME Artificial Intelligence in Medicine. 310–314. 2005.
|
|
112
|
Zhang W, Zhang J, Hoadley K, Kushwaha D,
Ramakrishnan V, Li S, Kang C, You Y, Jiang C, Song SW, et al:
miR-181d: A predictive glioblastoma biomarker that downregulates
MGMT expression. Neuro Oncol. 14:712–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Chen L, Zhang W, Yan W, Han L, Zhang K,
Shi Z, Zhang J, Wang Y, Li Y, Yu S, et al: The putative tumor
suppressor miR-524-5p directly targets Jagged-1 and Hes-1 in
glioma. Carcinogenesis. 33:2276–2282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Jones TA, Jeyapalan JN, Forshew T,
Tatevossian RG, Lawson AR, Patel SN, Doctor GT, Mumin MA, Picker
SR, Phipps KP, et al: Molecular analysis of pediatric brain tumors
identifies microRNAs in pilocytic astrocytomas that target the MAPK
and NF-κB pathways. Acta Neuropathol Commun. 3:862015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Shigemizu D, Akiyama S, Asanomi Y,
Boroevich KA, Sharma A, Tsunoda T, Matsukuma K, Ichikawa M, Sudo H,
Takizawa S, et al: Risk prediction models for dementia constructed
by supervised principal component analysis using miRNA expression
data. Commun Biol. 2:772019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Nunez-Iglesias J, Liu CC, Morgan TE, Finch
CE and Zhou XJ: Joint genome-wide profiling of miRNA and mRNA
expression in Alzheimer's disease cortex reveals altered miRNA
regulation. PLoS One. 5:e88982010. View Article : Google Scholar : PubMed/NCBI
|