Introduction
Oxaliplatin (OXA) is a widely used third-generation
platinum anticancer drug that is becoming the standard-of-care in
the management of colorectal cancer (CRC). Conversely, other
platinum-based drugs lack anticancer efficacy in CRC (1). However, many patients do not benefit
from OXA treatment (2). Buss et
al (3) indicated that a reduction
in the influx rate of OXA has been observed in resistant CRC cell
lines; however, no difference in efflux rate exists between
sensitive and resistant CRC cell lines. In addition, Oguri et
al (4) demonstrated that the
intracellular accumulation of OXA in PC-14 cells resistant to OXA
is reduced compared with in parental cells; however, no difference
has been reported in the expression of efflux transporters SLC47A1
and SLC47A2. Organic cation transporter 3 (OCT3), which belongs to
the solute carrier 22 member family and is encoded by solute
carrier family 22 member 3 (SLC22A3), is critical for drug
transportation and cellular detoxification (5,6). The
expression of OCT3 in CRC cell lines is higher than that of other
organic cation transporters, and the higher the expression of OCT3
in CRC cell lines, the higher the concentration of OXA in the cells
(7), suggesting that OCT3 may have a
role in the uptake of OXA, although the results are contradictory
(8). Whether the expression and
regulation of OCT3 influences the effect of OXA on CRC and the
possible mechanisms remain to be investigated. A recent study
suggested that SLC22A3 may be a tumor suppressor gene
(9–12). Fu et al demonstrated that
SLC22A3 suppresses esophageal squamous cell carcinoma
metastasis by inhibiting epithelial-mesenchymal transition (EMT)
(12). Guo et al reported that
OXA-resistant HCT116 CRC cells exhibit an EMT phenotype
characterized by upregulated expression of matrix metalloproteinase
(MMP)2 and MMP9, and downregulated E-cadherin expression (13). Further investigation is required as to
whether OCT3 can increase the effects of OXA on CRC by inhibiting
the EMT of CRC cells. CRC is the third most common cancer worldwide
(14). At diagnosis, ~20% of patients
with CRC have distant metastases, with 30–40% having developed
vascular and lymph node metastasis (15). Therefore, the discovery of targets
that can affect the malignant behavior of CRC is of great
significance in developing novel preventative and treatment
strategies.
Aberrant DNA methylation is a potential mechanism
underlying the development of CRC and reflects the chemosensitivity
of patients receiving postoperative adjuvant chemotherapy (16–19). DNA
methylation is an epigenetic process that can lead to silencing of
gene expression and can be reversed by DNA-demethylating agents.
Analysis of SLC22A3 using MethPrimer (20) revealed that CpG islands exist in its
promoter region, suggesting that regulation of this gene by
methylation may have a role in its expression. The DNA
methyltransferase inhibitor decitabine (DAC) is a Food and Drug
Administration-approved drug used clinically to treat acute
myelodysplastic leukemia, and has been reported to inhibit cell
invasiveness and proliferation of CRC lines (21).
The present study aimed to explore the effect and
possible mechanisms of OCT3 in OXA treatment of CRC in vitro
and in vivo. Specifically, OCT3 expression was induced in
HCT116 cells with low OCT3 expression using DAC, and SLC22A3
expression was knocked down in HT29 cells with high OCT3 expression
via stable lentiviral interference, in order to investigate the
effects of OCT3 expression on OXA transport and CRC cell viability.
The direct effect of OCT3 on malignant biological behaviors, such
as migration and invasion of CRC cells, was investigated using a
wound healing assay and reverse transcription-quantitative
polymerase chain reaction (RT-qPCR). In addition, the curative
effect of DAC combined with OXA in a nude mice xenograft model of
CRC was explored and the association between OCT3 expression and
OXA concentration was investigated. The concentration of OXA was
detected by inductively coupled plasma mass spectrometry
(ICP-MS).
Materials and methods
Materials
OXA was purchased from Dalian Meilun Biotechnology
Co., Ltd. DAC (5-Aza-2′-deoxycytidine) was purchased from Selleck
Chemicals and MTT was purchased from Sigma-Aldrich (Merck KGaA).
Solutions of OXA (7 mmol/l) and DAC (40 mmol/l) were freshly
prepared in DMEM cell culture media (Gibco; Thermo Fisher
Scientific, Inc.).
Cell culture
CRC cell lines HCT116, HT29, SW620 and 293T were
obtained from the Shanghai Institute of Cell Biology, Chinese
Academy of Sciences. HCT116, HT29 and 293T cells were cultured in
high glucose DMEM (Gibco; Thermo Fisher Scientific, Inc.), and
SW620 cells were cultured in Leibovitz's L-15 (Gibco; Thermo Fisher
Scientific, Inc.). Cells were cultured in medium supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.). HCT116, HT29 and 293T cell lines
were grown at 37°C in a humidified atmosphere containing 5%
CO2/95% air, whereas SW620 cells were grown at 37°C in a
humidified atmosphere containing 100% air. HT29 STR profiles
matched the standards recommended for HT29 cell line
authentication, with an EV value of 94%.
Construction of stable transfected
cell lines
A lentiviral vector encoding SLC22A3 short
hairpin RNA (shRNA) was designed and synthesized by Shanghai Yile
Biotechnology Company, Ltd. The shRNA negative control sequence,
which had no significant homology to human gene sequences, was
5′-CAACAAGATGAAGAGCACCAA-3′. The shRNA sequence targeting
SLC22A3 (sh-OCT3) was 5′-GAGGAAATGCACACTTATTCT-3′ for HCT116
cells and 5′-GAATTGTACCCAACAACATTA-3′ for HT29 cells. These shRNA
fragments were cloned into pLv-shRNA-GP vector-Puro (Shanghai Yile
Biotechnology Company, Ltd.) to construct lentiviral vectors. A
total of 3×106 293T cells were inoculated in a 6 cm
culture dish 1 day prior to transfection. The cells were cultured
overnight in a 37°C and 5% CO2 to ensure 80–90% cell
confluence at the time of transfection. Subsequently, the
lentiviral vector (0.6 µg/ml) and auxiliary plasmids (0.6 µg/ml;
Gag-Pol:Rev:VSVG ratio of 5:2:3; Shanghai Yile Biotechnology
Company, Ltd.) were transfected into 293T cells using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol at a
ratio of 1:1 to produce the lentivirus. 293T cells were incubated
at 37°C and 5% CO2 for 48 h. The supernatant was
collected 48 h post-transfection, centrifuged at 4°C and 1,500 × g
for 10 min, and filtered through a 0.45-µm microporous membrane to
remove cell debris, and temporarily stored at 4°C. A total of
4×105 human CRC cell lines (HCT116 and HT29) were
inoculated in a 12-well plate and infected with the viral
suspension (100 µl crude viral liquid per well) alongside 6 µg/ml
polybrene [Yeasen Biotechnology (Shanghai) Co., Ltd.]; the optimal
lentivirus volume was determined through preliminary experiments. A
total of 72 h post-infection, the positive stably transduced cell
lines were screened using puromycin (Thermo Fisher Scientific,
Inc.) at 1 µg/ml for HCT116 cells and 0.5 µg/ml for HT29 cells.
RT-qPCR was employed to detect interference efficiency.
Cell treatment
For RT-qPCR analysis, a total of 2×105
cells were seeded into 6-well plates, 6 h after which, DAC was
added at different density (0.6, 1.25, 2.5, 5 and 50 µM for HCT116
cells; 0.63, 1.25, 2.5, 5 and 50 µM for SW620 cells; 2.5, 5, 20, 50
and 100 µM for HT29 cells) for 72 h. In addition, cells of
different concentrations were inoculated into 6-well plates, 2.5 µM
DAC was added to HCT116 cells for 24 h (6×105 cells per
well), 48 h (4×105 cells per well) and 72 h
(2×105 cells per well), and to SW620 and HT29 cells for
48 h (4×105 cells per well), 72 h (2×105
cells per well) and 96 h (1×105 cells per well). Cells
were incubated at 37°C and 5% CO2.
RT-qPCR analysis
Total RNA was isolated from cultured cells (80%
confluent) using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
First-strand cDNA was synthesized using a reverse transcriptase kit
(cat. no. TAKARA047A; Takara Biotechnology Co., Ltd.) according to
the manufacturer's instructions. Subsequently, qPCR was conducted
according to the manufacturer's protocols under the following
conditions: Denaturation for 5 min at 95°C, followed by 40 cycles
of 15 sec at 95°C, 20 sec at 60°C and 30 sec at 72 °C, and a final
extension step of 5 min at 72°C on a Linegene 9620 real-time PCR
system (Hangzhou Bioer Technology Co., Ltd.) using SYBR-Green
master mix [Yeasen Biotechnology (Shanghai) Co., Ltd.].
Gene-specific primers were synthesized by the Beijing Genomics
Institute and are listed in Table I.
All samples were normalized against ACTB expression.
Relative gene expression was analyzed using the 2−ΔΔCq
method (22).
 | Table I.Primers for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Primers for reverse
transcription-quantitative polymerase chain reaction analysis.
| Oligonucleotide
name | Source | Sequence
(5′-3′) | Amplified fragment
length (bp) |
|---|
|
SLC22A3-F | Human |
CCCTGGAATTGCCTACTTCA | 102 |
|
SLC22A3-R | Human |
GACTCAGGGACCACCCAGTA |
|
| β-actin-F | Human |
CATCGAGCACGGCATCGTCA | 211 |
| β-actin-R | Human |
TAGCACAGCCTGGATAGCAAC |
|
| MMP2-F | Human |
GATACCCCTTTGACGGTAAGGA | 112 |
| MMP2-R | Human |
CCTTCTCCCAAGGTCCATAGC |
|
Western blotting
Cells were lysed in ice-cold RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing PMSF, and protein
concentrations were detected using a bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology). A total of 40 µg
protein was separated by 10% SDS-PAGE and transferred onto
polyvinylidene fluoride membranes (EMD Millipore), which were
blocked by soaking in 5% non-fat milk for 1.5 h at 37°C.
Subsequently, the membranes were incubated with polyclonal rabbit
anti-human OCT3 antibody (1:500; cat. no. OM291394; OminimAbs) or
polyclonal rabbit anti-human GAPDH antibody (1:1,000; cat. no.
WL01114; Wanleibio Co., Ltd.) overnight at 4°C. After washing with
1X TBS- 0.1% Tween three times (10 min/wash), the membranes were
incubated with goat anti-rabbit immunoglobulin G/horseradish
peroxidase (1:10,000; cat. no. bs-0295G-HRP; BIOSS) for 1 h at
37°C. Finally, the bands were visualized using an enhanced
chemiluminescence kit (EMD Millipore). Signals were semi-quantified
by ImageJ software (version 1.8.0; National Institutes of Health)
and normalized to GAPDH.
Cell viability assay
The cytotoxicity of compounds was examined using the
MTT assay (Invitrogen; Thermo Fisher Scientific, Inc.). Cells were
seeded into 96-well plates at ~5,000 cells per well, incubated
overnight, and treated with 2.5 µM DAC for the following durations:
HCT116 for 24, 48 and 72 h; SW620 cells for 48 and 72 h; and HT29
cells for 72 h. OXA was added at different concentrations (0.625,
1.25, 2.5, 10, 20, 50, 200, 800, 3,200 and 6,400 µM for HCT116
cells; 1.25, 2.5, 10, 50, 75, 100, 200, 600, 1,200 and 2,400 µM for
SW620 cells; 0.625, 2.5, 5, 10, 25, 75, 150, 300, 350, 400 and 650
µM for HT29 cells) for a further 24 h, and 10 µl MTT reagent was
then added to each well and incubated at 37°C for 4 h. After gently
removing the culture medium from the 96-well plates, 100 µl
dimethyl sulfoxide was added to the 96-well plates, and the
crystals were dissolved in a shaker at low speed at 37°C for 10
min. An automatic microplate spectrophotometer (Multiskan MK3;
Thermo Fisher Scientific, Inc.) was used to measure absorbance at
490 nm. All experiments were repeated at least three times.
Cell migration assay
The bottoms of 9.5 cm2 dishes were marked
with a pen and ruler prior to cell seeding. Subsequently, control,
negative control and sh-OCT3 HT29 cells (1.6×105 cells)
were seeded into 9.5 cm2 dishes in 1% serum-containing
medium, and allowed to reach 90–100% confluence overnight, forming
a monolayer. Subsequently, the cells were scratched with a 10-µl
pipette tip perpendicularly to the surface to cross the marker
lines, forming fixed detection points. Cells were washed with PBS
twice followed by the addition of fresh media with 1% FBS. Images
were captured at 0, 24, 36 and 48 h using an inverted light
microscope.
Methylation-specific PCR (MSP)
assays
The promoter region of SLC22A3 was searched
for in Genecopoeia (www.genecopoeia.com). MSP primers, as detailed in
Table II, were designed using
MethPrimer (The Li Lab, www.urogene.org/methprimer) for the promoter region of
SLC22A3. Genomic DNA extraction and purification was
performed using the Genomic DNA Small Purification kit (Wanleibio
Co., Ltd.) according to the manufacturer's protocol. DNA bisulfite
transformation, purification, and detection of DNA concentration
and purity were conducted using a DNA bisulfite transformation kit
(centrifugal column type) [cat. no. DP215; Tiangen Biotech
(Beijing) Co., Ltd.] according to the manufacturer's protocol.
Subsequently, MSP was performed using a MSP kit [cat. no. EM101;
Tiangen Biotech (Beijing) Co., Ltd.], according to manufacturer's
protocol. After the reaction, 10 µl product was obtained for
agarose gel electrophoresis (12% agarose) and the products of gel
electrophoresis were retrieved according to E.Z.N.A.®
Gel Extraction kit (cat. no. D2500-01; Omega Bio-Tek, Inc.). The
PCR product was linked to the pClone007 vector using the pClone007
vector kit (cat. no. TSV-007; Beijing TsingKe Biotech Co., Ltd.)
and identified through sequencing by TSINGKE Biological
Technology.
| Table II.Primers for methylation-specific
polymerase chain reaction analysis. |
Table II.
Primers for methylation-specific
polymerase chain reaction analysis.
| Oligonucleotide
name | Source | Sequence
(5′-3′) |
|---|
| SLC22A3
M-F | Human |
TATATGGGCGTAGGAGGTTTC |
| SLC22A3
M-R | Human |
AAACCCGATCTCTCAACGAC |
| SLC22A3
U-F | Human |
TTTTATATGGGTGTAGGAGGTTTTT |
| SLC22A3
U-R | Human |
ACTTCTAAAACCCAATCTCTCAACA |
Xenograft tumor formation assay in
nude mice
Animal experiments were approved by the Ethical
Committee for Animal Research of Sichuan University. HCT116 cells
(5×106 cells; 100 µl) were subcutaneously inoculated
into the upper right flank of 6-week-old male nude mice (total
n=40; weight, ~15 g). BALB/c-nu/nu mice were purchased from Chengdu
Dossy Experimental Animals Co., Ltd. The mice were housed under a
12-h light/dark cycle at 25°C with 50–60% humidity and free access
to food and water. The length and width of tumors were measured
using calipers and the tumor volume was calculated as follows:
[(lengthxwidth2)/2] twice per week. The relative tumor
volume (RTV) was calculated as follows:
RTV=Vt/V0. Where V0 is the tumor
volume at grouping and Vt is the tumor volume at each measurement.
Approximately 10 days following inoculation, nude mice with tumor
volumes of 100–200 mm3 were selected and divided into
groups as followings: Control group, DAC group, OXA group and
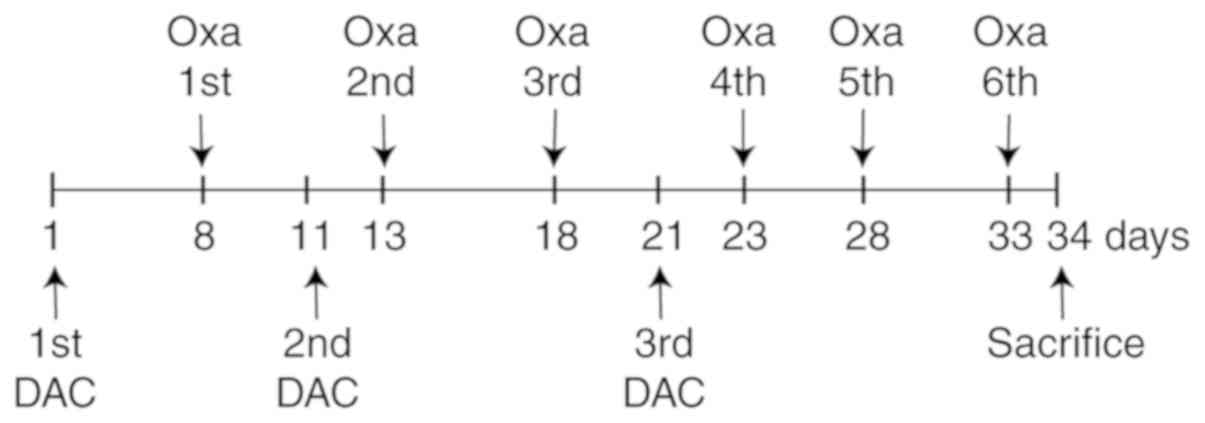
combination group. DAC (2.5 mg/kg) was intraperitoneally
administered into mice in the DAC and combination groups every 3 h
on days 1, 11 and 21, three times a day. OXA (10 mg/kg) was
intraperitoneally administered once into mice in the OXA and
combination groups on days 8, 13, 18, 23, 28 and 33. Mice in the
control group were intraperitoneally administered an equal volume
of solvent (5% glucose). The experimental procedure is shown in
Fig. 1. On day 34 mice were
sacrificed by cervical dislocation and tumors were collected for
the measurement of tumor weights, detection of SLC22A3 mRNA
and protein expression by qPCR and western blotting, respectively,
and measurement of OXA concentration by ICP-MS (iCAP Q; Thermo
Fisher Scientific, Inc.), according to manufacturer's protocol.
Briefly, for ICP-MS, samples were weighed (accurate to 0.1 mg) or
HT29 cells were counted prior to microwave digestion using a
WX-8000 Microwave Digester (PreeKem Scientific Instruments Co.,
Ltd.). Following microwave digestion, samples were analyzed by
ICP-MS with set parameters for detection.
Statistical analysis
Statistical analysis was performed using SPSS
version 17.0 (IBM Corp.) and GraphPad Prism 5.0 (GraphPad Software,
Inc.). Quantitative variables are presented as the mean ± standard
error of the mean. For comparisons of two groups of quantitative
data, Student's t-test was used. For multiple comparisons, one-way
ANOVA was performed followed by Dunnett's two-tailed test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Induction of OCT3 expression by
DAC
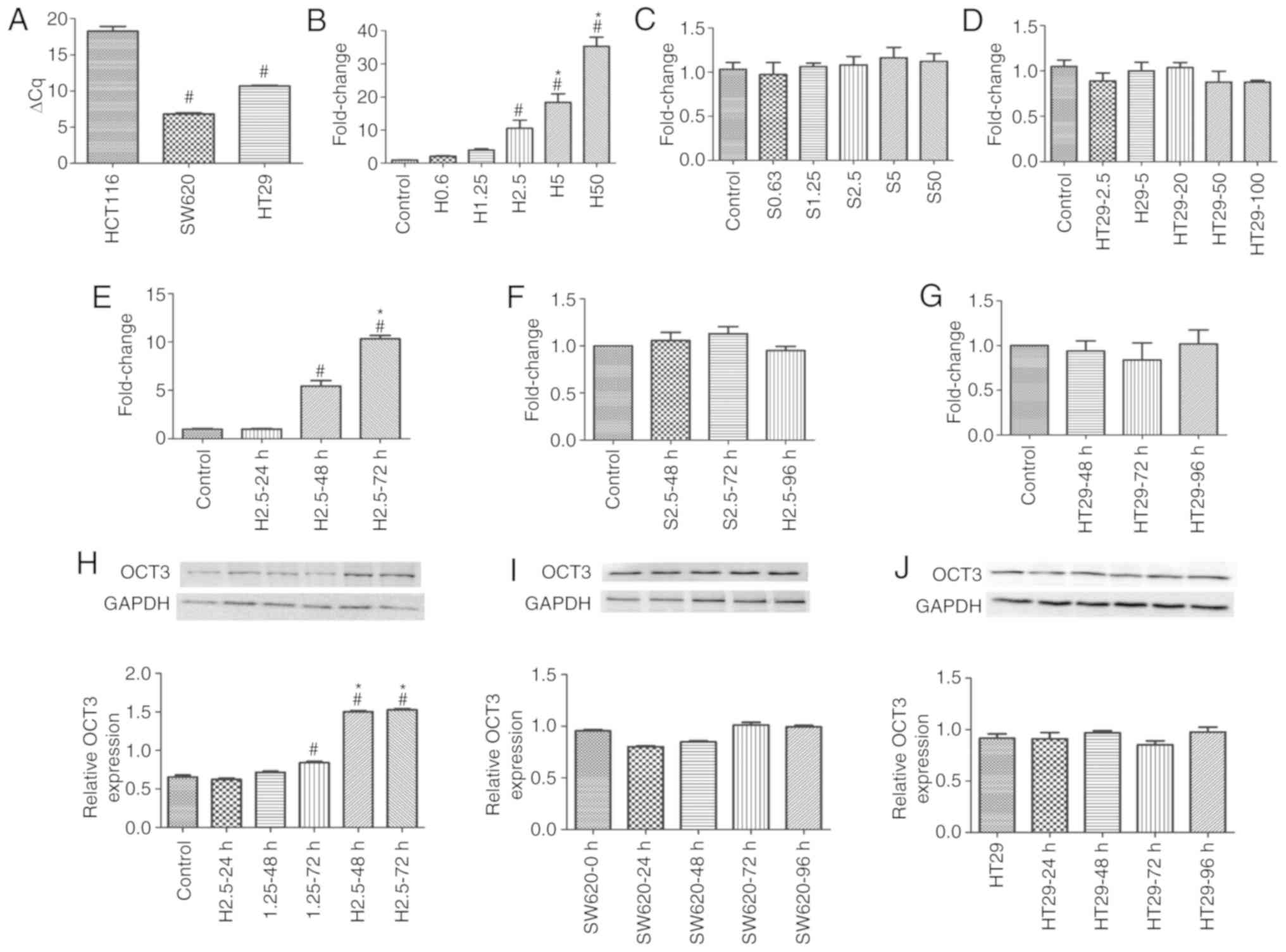
According to our preliminary experiments, HCT116 and
SW620 cells were treated with <50 µM DAC, and HT29 cells were
treated with <100 µM DAC, at which cell viability was >80%,
in order to detect the effects of DAC on OCT3 expression in the
three CRC cell lines. Baseline SLC22A3 mRNA expression was
lowest in HCT116 cells among the three CRC cell lines, and the mRNA
expression levels of SLC22A3 were significantly higher in
HT29 and SW620 cells (P<0.05, Fig.
2A). SLC22A3 mRNA expression was significantly induced
in HCT116 cells following treatment with DAC for 72 h in a
concentration-dependent manner (Fig.
2B), whereas SLC22A3 expression was not induced by DAC
in SW620 and HT29 cells (Fig. 2C and
D). The three CRC cell lines were then treated with 2.5 µM DAC;
the results revealed that the induction of SLC22A3 mRNA
expression was time-dependent in HCT116 cells, but not in HT29 or
SW620 cells (Fig. 2E-G). The
induction of OCT3 protein expression by DAC was similar to that of
its mRNA expression. DAC could upregulate OCT3 expression in HCT116
cells, in which the baseline expression of OCT3 was low, in a
concentration- and time-dependent manner, but not in SW620 or HT29
cells, in which the baseline expression levels of OCT3 were high
(Fig. 2H-J).
Methylation status of the SLC22A3 gene
promoter region as determined by MSP in CRC cells
To investigate the methylation status of the
SLC22A3 gene promoter region in CRC cells, MSP was
conducted. Methylated (−) and non-methylated products (+) in SW620
and HT29 cells, and methylated (+) and non-methylated products (−)
in HCT116 cells (Fig. 3A). As shown
in Fig. 3B, methylation products
decreased and non-methylation products increased in HCT116 cells
treated with 2.5 µM DAC for 48 and 72 h, respectively. Sequencing
revealed that the methylated PCR products were consistent with the
target sequence (Fig. 3C). These
findings confirmed that the methylation products were the desired
products.
Influence of DAC on the effects of OXA
and OCT3 on CRC cells
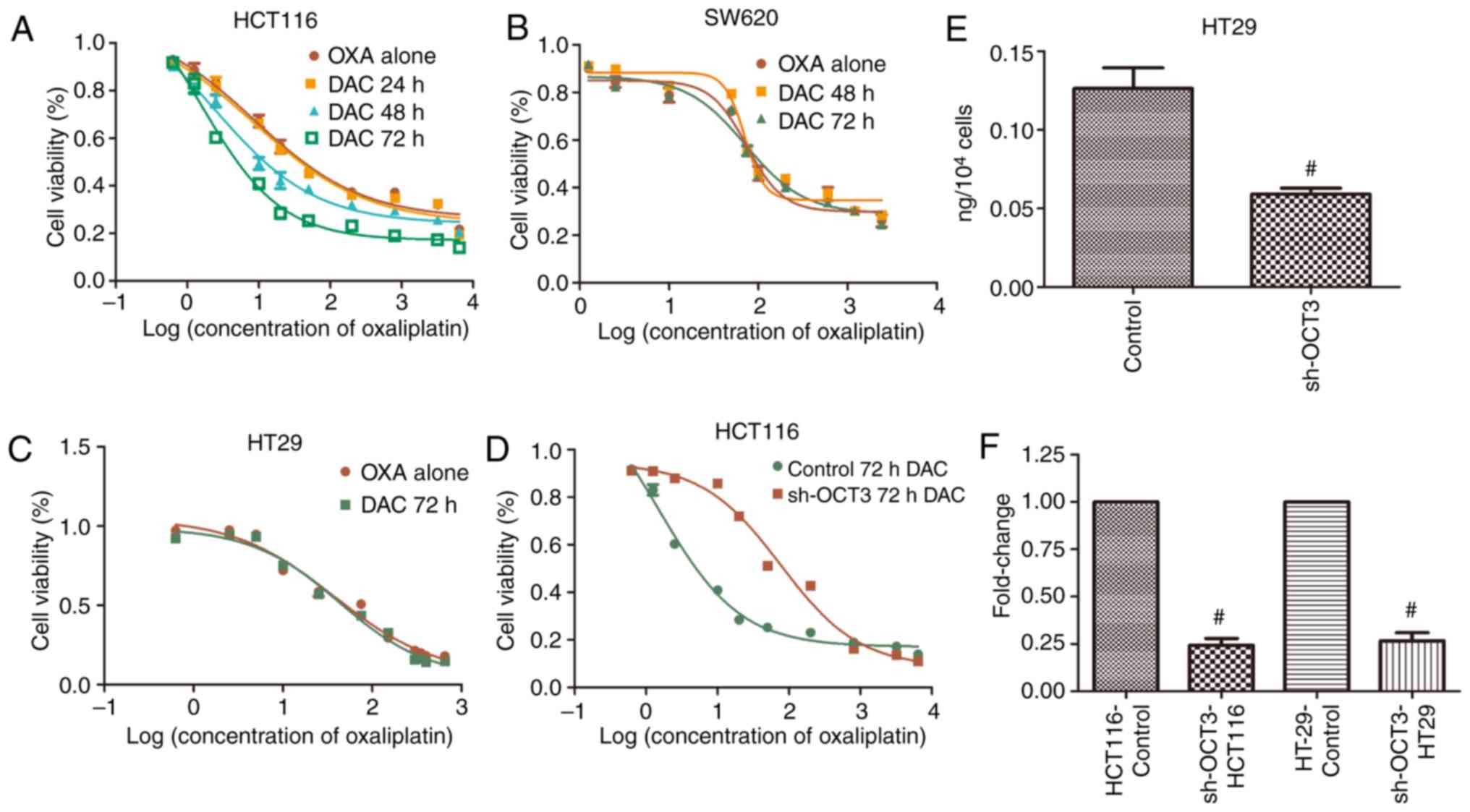
To explore the influence of DAC on the inhibitory
effects of OXA on CRC cells, CRC cell lines were treated with OXA
following treatment with 2.5 µM DAC for the indicated durations.
The results demonstrated that the concentration-activity curves of
OXA declined after HCT116 cells were treated with 2.5 µM DAC for 48
and 72 h, but not for 24 h (Fig. 4A),
and the IC50 values were also decreased (Table III). There was no significant effect
on SW620 cells treated with 2.5 µM DAC for 48 or 72 h, nor HT29
cells for 72 h (Fig. 4B and C).
Compared with wild-type HCT116 cells, the concentration-activity
curve of OXA in HCT116 cells treated with sh-OCT3 shifted to the
right after treatment with DAC (Fig.
4D). The IC50 value of OXA was markedly higher in
HCT116 cells expressing sh-OCT3 compared with in wild-type HCT116
cells following treatment with DAC (Table III). Furthermore, the effects of
OCT3 on the concentration of OXA in HT29 cells were determined; OXA
concentration in HT29 cells with SLC22A3 knockdown (sh-OCT3)
was significantly lower than that in the control group (Fig. 4E). The knockdown efficiency on SLC22A3
in HCT116 and HT29 was >75% (Fig.
4F).
| Table III.Effects of DAC on IC50 of
OXA in colorectal cancer cells. |
Table III.
Effects of DAC on IC50 of
OXA in colorectal cancer cells.
|
| 2.5 µM DAC |
|---|
|
|
|
|---|
| Cell line | 0 h | 24 h | 48 h | 72 h |
|---|
| HCT116 | 8.64
(3.68–20.29) | 8.37
(3.11–22.47) | 2.38
(0.70–8.1) | 1.50
(0.62–3.62) |
| SW620 | 71.94
(56.92–90.91) |
| 70.22
(63.78–77.33) | 74.41
(64.1–86.37) |
| HT29 | 37.58
(23.76–59.43) |
|
| 42.89
(31.10–59.16) |
| HCT116-sh-OCT3 |
|
|
| 76.61
(55.59–105.6) |
Direct effect of OCT3 expression on
the migration and invasion of CRC cells
To explore the role of OCT3 in the migration of CRC
cells, wound healing assays were conducted. The results revealed
that the percentage of migrating HT29 cells with SLC22A3
knockdown (sh-OCT3) was significantly higher compared with in the
control group (Fig. 5A-a and A-b). To
investigate the role of OCT3 in CRC cell invasion, the mRNA
expression levels of MMP2 were detected; MMP2 is an
indicator of cancer cell invasion, whereby elevated MMP2
expression indicates increased invasion (23,24). The
results demonstrated that the expression levels of MMP2 were
significantly higher in HT29 cells with SLC22A3 knockdown
compared with in the control group (Fig.
5B). Consistent with this, MMP2 mRNA expression in
HCT116 cells was reduced in a concentration-dependent manner once
OCT3 expression was induced by 1.25 and 2.5 µM DAC (Fig. 5C).
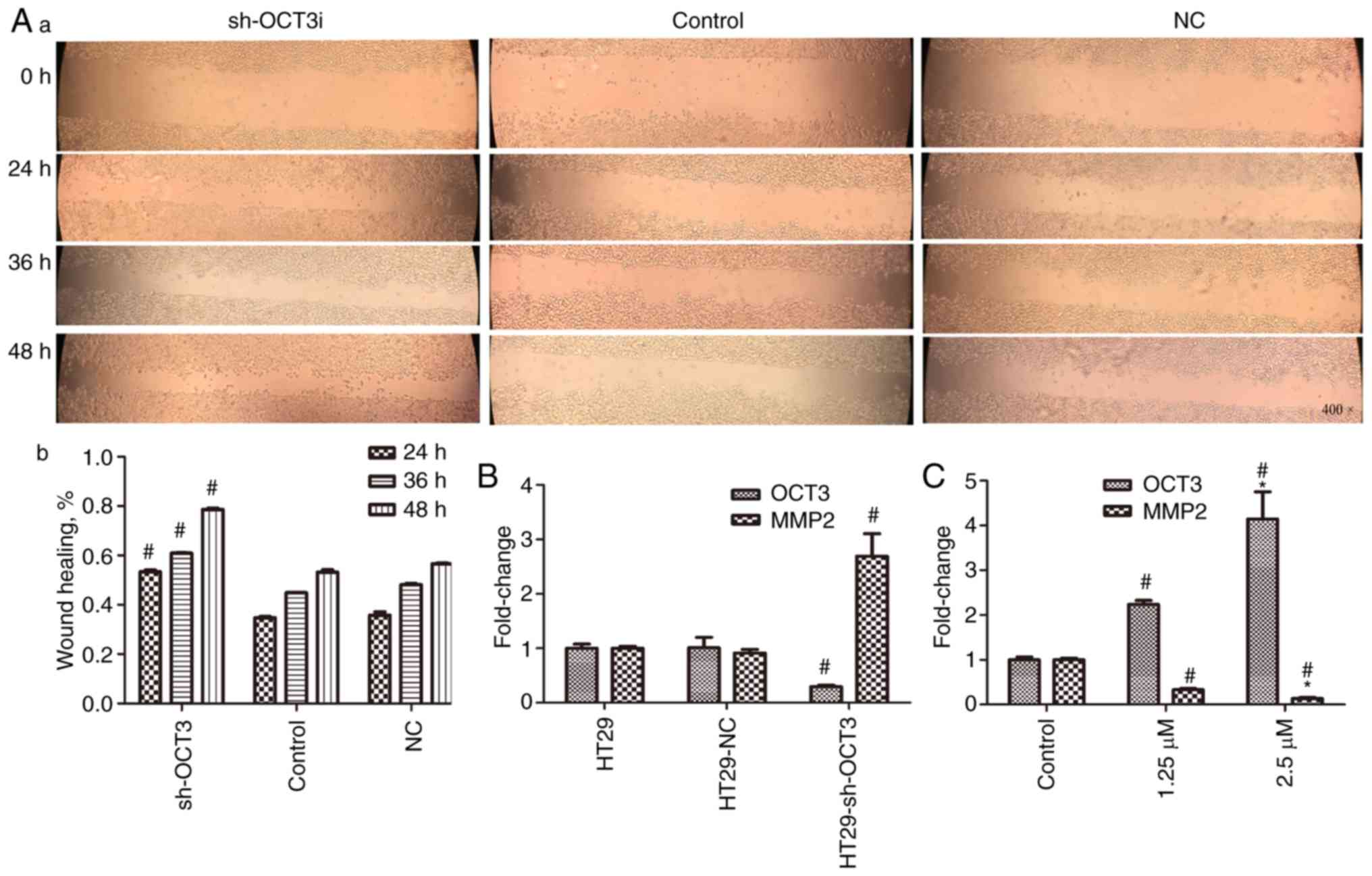 | Figure 5.Role of OCT3 expression in cell
migration, as determined by wound healing assay, and in cell
invasion, as determined by detecting MMP2 expression. (A-a)
Representative images of cell migration (magnification, ×400) and
(A-b) wound healing percentage in wild-type and sh-OCT3 HT29 cells
at different time points after scratching. (B) Impact of solute
carrier family 22 member 3 knockdown (sh-OCT3) on MMP2 mRNA
expression. (C) Impact of treatment with 1.25 and 2.5 µM DAC on
MMP2 mRNA expression. #P<0.05 compared with
Control group; *P<0.05 compared with 1.25 µM DAC group. DAC,
decitabine; MMP2, matrix metalloproteinase 2; NC, negative
control; OCT3, organic cation transporter 3; OXA, oxaliplatin; sh,
short hairpin RNA. |
Impact of DAC on the effects of OXA on
CRC xenografts and the association with OCT3 expression
Representative images of xenografts in nude mice are
shown in Fig. 6A. From alterations in
RTV curve of xenografts in nude mice, it was revealed that RTV was
lowest in the combined DAC and OXA group (Fig. 6B). The inhibitory effect was as
follows: Combination group >DAC group >OXA group (Fig. 6A and B). On day 7 following initiation
of DAC administration, OCT3 expression in xenografts of mice
administered DAC was higher than in xenografts of mice that did not
receive DAC (Fig. 7A-a and A-b). At
the end of the experiment, OCT3 expression in xenografts of mice
administered DAC remained higher than in xenografts of mice that
did not receive DAC (Fig. 7B-a and
7B-b). Furthermore, the concentration of OXA in xenografts of
mice treated with DAC was higher than in xenografts of mice not
treated with DAC (Fig. 7C).
Discussion
This study demonstrated that the DNA
methyltransferase inhibitor DAC could increase the effects of OXA
on CRC xenografts, along with an increase in OCT3 expression. DAC
upregulated OCT3 expression in CRC cells with OCT3 low expression,
which may be of great significance in reversing anticancer drug
resistance to OCT3 substrates (25,26),
including OXA in CRC. In addition, DAC may inhibit tumor growth by
inducing the expression of other tumor suppressor genes (27). Notably, long-term use of DAC may
promote resistance, rendering it unable to exert its role in
inhibiting DNA methyltransferase 1 (28,29). Tet
methylcytosine dioxygenase 2 (TET2) encodes a protein that
catalyzes the conversion of the modified DNA base methylcytosine to
5-hydroxymethylcytosine, and serves a key role in active DNA
demethylation. A recent study (30)
suggested that the addition of low-dose vitamin C to DAC may
improve complete remission and prolong overall survival compared
with DAC alone in elderly patients with AML. Activation of
TET2, due to the combination of DAC and vitamin C, may be
one of the underlying mechanisms. DAC is currently only approved
for use in hematological tumors, and clinical trials of DAC in
solid tumors are in progress. With the advances made in clinical
trials, indications for DAC may expand to solid tumors. The present
study provides evidence for the effectiveness of DAC combined with
OXA for the treatment of CRC through upregulation of OCT3
expression via DNA demethylation.
Tumor cell migration is one of the critical steps in
the invasion and metastasis of malignant tumors. Cell migration
assays suggested that OCT3 may inhibit the migration of CRC cells;
however, the underlying mechanism requires further investigation.
MMP2 belongs to the MMP family, and its expression is
associated with the invasion and metastasis of CRC (31,32).
Therefore, MMP2 was selected as a marker to indicate the
possible role of OCT3 in the biological behavior of malignant
tumors by detecting the impact of OCT3 expression on MMP2
expression. The results revealed that MMP2 expression was
upregulated in SLC22A3-knockdown CRC cells, which indicated
that OCT3 may inhibit the invasion and metastasis of CRC by
reducing MMP2 expression. However, the mechanism by which
OCT3 inhibits MMP2, directly or indirectly, is unclear.
Consistent with the present results, Fu et al (12) recently revealed that the expression of
SLC22A3 in non-tumorous tissues of patients with familial
esophageal cancer is significantly downregulated, and
adenosine-to-inosine RNA editing of this gene leads to
downregulation of its expression and is significantly associated
with lymph node metastasis. Further investigations have reported
that OCT3 can directly bind α-actinin-4 (ACTN4), and may
inhibit ACTN4-mediated actin cross-linking and cell
migration (12). A limitation of the
present study is that, for migration assays, 1% serum-containing
medium was added to the culture dish following wound generation,
instead of serum-free medium. Although 1% serum-containing medium
has little effect on cell proliferation, it may not completely
exclude the effect of cell proliferation on the experiment.
In conclusion, OCT3 may directly inhibit the
malignant biological behavior of cancer and may be considered a
novel target for intervention in CRC. OCT3-specific inhibitors have
recently been reported (33,34), and will be useful tools for further
investigating the function of OCT3.
Acknowledgements
The authors would like to thank Dr Nikki March for
editing a draft of this manuscript.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors contributions
JG, XJ and LW were involved in conception and
design. JG and WZ conducted cell biological experiments and
xenograft experiments. JG and TL analyzed and interpreted the data.
JG, ST and YW made substantial contributions to acquisition of
data, or analysis and interpretation of data, and drafted the
manuscript. All authors agreed to be accountable for all aspects of
the work in ensuring that questions related to the accuracy or
integrity of any part of the work are appropriately investigated
and resolved. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the Ethical
Committee for Animal Research of Sichuan University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
OCT3
|
organic cation transporter 3
|
|
SLC22A3
|
solute carrier family 22 member 3
|
|
DAC
|
decitabine
|
|
OXA
|
oxaliplatin
|
|
MMP2
|
matrix metalloproteinase-2
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
ICP-MS
|
inductively coupled plasma mass
spectrometry
|
|
FBS
|
fetal bovine serum
|
|
EMT
|
epithelial-mesenchymal transition
|
|
MSP
|
methylation-specific polymerase chain
reaction
|
|
CRC
|
colorectal cancer
|
References
|
1
|
Perego P and Robert J: Oxaliplatin in the
era of personalized medicine: From mechanistic studies to clinical
efficacy. Cancer Chemother Pharmacol. 77:5–18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martinez-Balibrea E, Martinez-Cardus A,
Gines A, Ruiz de Porras V, Moutinho C, Layos L, Manzano JL, Bugés
C, Bystrup S, Esteller M and Abad A: Tumor-Related molecular
mechanisms of oxaliplatin resistance. Mol Cancer Ther.
14:1767–1776. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Buss I, Garmann D, Galanski M, Weber G,
Kalayda GV, Keppler BK and Jaehde U: Enhancing lipophilicity as a
strategy to overcome resistance against platinum complexes. J Inorg
Biochem. 105:709–717. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oguri T, Kunii E, Fukuda S, Sone K, Uemura
T, Takakuwa O, Kanemitsu Y, Ohkubo H, Takemura M, Maeno K, et al:
Organic cation transporter 6 directly confers resistance to
anticancer platinum drugs. Biomed Rep. 5:639–643. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu X, Kekuda R, Huang W, Fei YJ, Leibach
FH, Chen J, Conway SJ and Ganapathy V: Identity of the organic
cation transporter OCT3 as the extraneuronal monoamine transporter
(uptake2) and evidence for the expression of the transporter in the
brain. J Biol Chem. 273:32776–32786. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koepsell H: Polyspecific organic cation
transporters: Their functions and interactions with drugs. Trends
Pharmacol Sci. 25:375–381. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yokoo S, Masuda S, Yonezawa A, Terada T,
Katsura T and Inui K: Significance of organic cation transporter 3
(SLC22A3) expression for the cytotoxic effect of oxaliplatin in
colorectal cancer. Drug Metab Dispos. 36:2299–2306. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang S, Lovejoy KS, Shima JE, Lagpacan
LL, Shu Y, Lapuk A, Chen Y, Komori T, Gray JW, Chen X, et al:
Organic cation transporters are determinants of oxaliplatin
cytotoxicity. Cancer Res. 66:8847–8857. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grisanzio C, Werner L, Takeda D, Awoyemi
BC Pomerantz MM, Yamada H, Sooriakumaran P, Robinson BD Leung R,
Schinzel AC, et al: Genetic and functional analyses implicate the
NUDT11, HNF1B, and SLC22A3 genes in prostate cancer pathogenesis.
Proc Natl Acad Sci USA. 109:11252–11257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vollmar J, Lautem A, Closs E, Schuppan D,
Kim YO, Grimm D, Marquardt JU, Fuchs P, Straub BK, Schad A, et al:
Loss of organic cation transporter 3 (Oct3) leads to enhanced
proliferation and hepatocarcinogenesis. Oncotarget.
8:115667–115680. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xiong JX, Wang YS, Sheng J, Xiang D, Huang
TX, Tan BB, Zeng CM, Li HH, Yang J, Meltzer SJ, et al: Epigenetic
alterations of a novel antioxidant gene SLC22A3 predispose
susceptible individuals to increased risk of esophageal cancer. Int
J Biol Sci. 14:1658–1668. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fu L, Qin YR, Ming XY, Zuo XB, Diao YW,
Zhang LY, Ai J, Liu BL, Huang TX, Cao TT, et al: RNA editing of
SLC22A3 drives early tumor invasion and metastasis in familial
esophageal cancer. Proc Natl Acad Sci USA. 114:E4631–E4640. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo C, Ma J, Deng G, Qu Y, Yin L, Li Y,
Han Y, Cai C, Shen H and Zeng S: ZEB1 promotes oxaliplatin
resistance through the induction of epithelial-mesenchymal
transition in colon cancer cells. J Cancer. 8:3555–3566. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaiser C, Meurice N, Gonzales IM, Arora S,
Beaudry C, Bisanz KM, Robeson AC, Petit J and Azorsa DO:
Chemogenomic analysis identifies Macbecin II as a compound specific
for SMAD4-negative colon cancer cells. Chem Biol Drug Des.
75:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luebeck GE, Hazelton WD, Curtius K, Maden
SK, Yu M, Carter KT, Burke W, Lampe PD, Li CI, Ulrich CM, et al:
Implications of epigenetic drift in colorectal neoplasia. Cancer
Res. 79:495–504. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou C, Pan R, Hu H, Li B, Dai J, Ying X,
Yu H, Zhong J, Mao Y, Zhang Y, et al: TNFRSF10C methylation is a
new epigenetic biomarker for colorectal cancer. PeerJ. 6:e53362018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chu CH, Chang SC, Wang HH, Yang SH, Lai KC
and Lee TC: Prognostic values of EPDR1 hypermethylation and its
inhibitory function on tumor invasion in colorectal cancer. Cancers
(Basel). 10:E3932018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yokoi K, Harada H, Yokota K, Ishii S,
Tanaka T, Nishizawa N, Shimazu M, Kojo K, Miura H, Yamanashi T, et
al: Epigenetic status of CDO1 gene may reflect chemosensitivity in
colon cancer with postoperative adjuvant chemotherapy. Ann Surg
Oncol. 26:406–414. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li LC and Dahiya R: MethPrimer: Designing
primers for methylation PCRs. Bioinformatics. 18:1427–1431. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tanaka S, Hosokawa M, Ueda K and Iwakawa
S: Effects of decitabine on invasion and exosomal expression of
miR-200c and miR-141 in oxaliplatin-resistant colorectal cancer
cells. Biol Pharm Bull. 38:1272–1279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cao LL, Pei XF, Qiao X, Yu J, Ye H, Xi CL,
Wang PY and Gong ZL: SERPINA3 silencing inhibits the migration,
invasion, and liver metastasis of colon cancer cells. Dig Dis Sci.
63:2309–2319. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Z, Ji N, Wang Z, Wu C, Sun Z, Li Y,
Hu F, Wang Z, Huang M and Zhang M: Fine particulate matter
(PM25) promoted the invasion of lung cancer cells via an
ARNT2/PP2A/STAT3/MMP2 pathway. J Biomed Nanotechnol. 15:4162019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shnitsar V, Eckardt R, Gupta S, Grottker
J, Müller GA, Koepsell H, Burckhardt G and Hagos Y: Expression of
human organic cation transporter 3 in kidney carcinoma cell lines
increases chemosensitivity to melphalan, irinotecan, and
vincristine. Cancer Res. 69:1494–1501. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsu CM, Lin PM, Chang JG, Lin HC, Li SH,
Lin SF and Yang MY: Upregulated SLC22A3 has a potential for
improving survival of patients with head and neck squamous cell
carcinoma receiving cisplatin treatment. Oncotarget. 8:74348–74358.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hagiwara H, Sato H, Ohde Y, Takano Y, Seki
T, Ariga T, Hokaiwado N, Asamoto M, Shirai T, Nagashima Y and Yano
T: 5-Aza-2′-deoxycytidine suppresses human renal carcinoma cell
growth in a xenograft model via up-regulation of the connexin 32
gene. Br J Pharmacol. 153:1373–1381. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qin T, Jelinek J, Si J, Shu J and Issa JP:
Mechanisms of resistance to 5-aza-2′-deoxycytidine in human cancer
cell lines. Blood. 113:659–667. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hosokawa M, Saito M, Nakano A, Iwashita S,
Ishizaka A, Ueda K and Iwakawa S: Acquired resistance to decitabine
and cross-resistance to gemcitabine during the long-term treatment
of human HCT116 colorectal cancer cells with decitabine. Oncol
Lett. 10:761–767. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao H, Zhu H, Huang J, Zhu Y, Hong M, Zhu
H, Zhang J, Li S, Yang L, Lian Y, et al: The synergy of vitamin C
with decitabine activates TET2 in leukemic cells and significantly
improves overall survival in elderly patients with acute myeloid
leukemia. Leuk Res. 66:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu XH, Wang JM, Yang SS, Wang FF, Hu JL,
Xin SN, Men H, Lu GF, Lan XL, Zhang D, et al: Down-regulation of
DAB2IP promotes colorectal cancer invasion and metastasis by
translocating hnRNPK into nucleus to enhance the transcription of
MMP2. Int J Cancer. 141:172–183. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu T, Wang L, Pan XL and Qi HL: Novel
compound, organic cation transporter 3 detection agent and organic
cation transporter 3 activity inhibitor, WO2015002150 A1: A patent
evaluation. Expert Opin Ther Pat. 26:857–860. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan X, Iyer KA, Liu H, Sweet DH and Dukat
M: A new chemotype inhibitor for the human organic cation
transporter 3 (hOCT3). Bioorg Med Chem Lett. 27:4440–4445. 2017.
View Article : Google Scholar : PubMed/NCBI
|