Introduction
Olive tree cultivation has been a common practice
for humans since 6,000 BC. Its products are widely used by
residents of the countries in the Mediterranean basin for food,
religious and medicinal applications (1). The olive tree (Olea europaea. L)
is one of the most extensively cultivated species worldwide, spread
over an area of 10 million hectares (2). Except for olives and olive oil, other
parts of the olive tree, such as blossoms and leaves have also
attracted attention. Previous findings have highlighted the health
effects of leaves, which comprise 10% of the total olive weight
(3). Specifically, olive leaves have
been used against various diseases, since it has been reported that
they possess antioxidant and antimicrobial activities, vasodilator
and hypoglycaemic effects (3,4). Additionally, extracts derived from olive
leaf have been used as foodstuff, food additives and functional
food constituents as they are rich in polyphenols (4).
Polyphenolic compounds are secondary metabolites of
plants that contribute to bitterness, astringency, color, flavor,
odor, flower pollination and oxidative stability, and also protect
against various pathogens and UV radiation (5). Furthermore, polyphenols and foods
enriched in them belong to the main scope of research activity
worldwide due to their putative advantageous effects on human
health, as regards their anticancer, antidiabetic and
antiatherogenic properties (5). The
flower is a part of the plant that contains a great variety of
natural antioxidants, such as phenolic acids, anthocyanin,
flavonoids and many other phenolic compounds. It has been
previously shown that, oleuropein aglycon (1.158–3.746 g/kg),
hydroxytyrosol (HT) (0.168–1.581 g/kg) and oleoside (0.143–1.325
g/kg) are the predominant phenolics in extracts originating from
the olive flower during several developmental stages (e.g., green
bud stage, white bud stage, recently opened flower stage, dehiscent
anther stage and at the stage where the anthers and petals are
abscised) (6). Olive blossoms have
not yet been fully investigated for their potential medicinal uses.
It has to be mentioned that a mature olive (Olea europaea.
L) tree produces approximately 500,000 flowers but only 1–2% of
them set fruits that reach maturity (7). Each inflorescence contains 15–30
flowers, depending on the cultivar (2). Notably, fruit growth that occurs from 1%
of the entire flower population is sufficient for the production of
a good commercial crop (7,8).
Nutrition, growth regulators, and environmental
factors, including temperature and light, have an impact on floral
bud induction and differentiation in olive. Specifically, the
optimum temperature for best flowering seems to be 10–13°C for a
period of 9–10 weeks. However, sensitivity to light seems to be
cultivar-dependent, thus some cultivars require less light for the
transformation of buds to flowers (9). Additionally, nutrients are dependent on
fruit load, thus sodium and potassium concentrations are decreased
when the fruit load is high, whereas calcium levels are increased.
Finally, the high levels of chlorogenic acids induce the
transformation of buds to flower (9).
The olive blossom phenotype depicts an annual cycle,
characterized by the bud formation during the preceding summer,
dormancy during winter, budburst in late winter, and structural
development of flowers, from budburst to spring. The sexual
reproduction of the olive tree is underlined by earnest phenomena
such as alternate bearing, pistil abortion, and the reproductive
self and cross-incompatibility (10).
The olive floral buds differentiate into inflorescences during
winter and floral bud differentiation during February.
Differentiation occurs in late February and bloom in May when the
formation of each flower part responds to the inflorescence. There
are also olive (Olea europaea. L) cultivars that are almost
completely self-incompatible, where the flowers are not fertilized
by pollen of the same cultivar (9).
The level of the fruit set seems to be independent form the amount
of flowers and the number of inflorescence, when a tree enters an
‘ON’ year where the flowering depicts the maximum percentage
(7).
Olive oil production is a physiological process,
nevertheless it is clear that it is accompanied by an extensive
enhancement in the amount of olive oil by-products, which are
serious waste agents causing environmental problems (11). Thus, it is imperative to develop new
ways to utilize such by-products in order to protect the
environment (11). Olive oil, fruit
and leaves have a well-known chemical composition and have been
extensively studied for their biological activities. However, less
attention has been given to olive tree flowers and especially those
derived from wild olive varieties for which higher phenolic content
is expected. The olive flower production occurs on a large scale
although a small amount of blossoms produce a mature crop.
Specifically, after petal drop 25% of the ovaries remain, but only
2% of the floral entities become a mature fruit (7). Therefore, our aim was to evaluate the
biological effects of four polyphenolic olive floral extracts (3
from the olive varieties Lianolia and Koroneiki and one from a wild
olive variety), with an holistic in vitro approach using
both chemical-based and cell culture-based tests. Owing to the
difficulties in the production of purified phenolic compounds and
given that extracts from mixtures usually exhibit stronger
antioxidant activities compared with individual molecules, our
interest was focused on the use of mixture plant extracts rather
than single compounds. Our ultimate aim was to shed light on the
antioxidant, antigenotoxic and antimutagenic potential of the
floral extracts in order to develop new products derived from the
extracts with pharmaceutical, nutritional and cosmetic
applications.
Materials and methods
Chemicals and reagents
Dulbecco's modified Eagle's medium (DMEM),
L-glutamine, penicillin, streptomycin, fetal bovine serum (FBS),
phosphate-buffered saline (PBS), 2′,7′-dichlorofluorescein
diacetate (DCF-DA), mercury orange, and trypsin were purchased from
Gibco. Cell proliferation kit II (XTT assay; Roche) was purchased
from Roche Diagnostics. Ethanol (EtOH) was purchased from Carlo
Erba Reactifs SDS. Methanol (MeOH) was obtained from Fisher
Scientific UK. All the solvents were of analytical grade. Distilled
water was used to prepare all aqueous solutions. H2O was
purchased from Macron Fine Chemicals [high-performance liquid
chromatography (HPLC) grade] and 2,2′-azobis
(2-methyl-propionamide) dihydrochloride (AAPH) was purchased from
Sigma-Aldrich.
Plant material and extraction
procedure
Four different samples of olive flowers belonging to
Greek varieties were collected. More specifically, two of these
samples (KTKT and ANKT) belong to the variety Lianolia from Corfu
Island with collection dates 5–6/5/17 and 14–15/5/17, respectively.
The third sample (AGRI) consists of flowers of the olive wild tree,
collected on 13–14/5/17 from the island of Corfu. The final sample
of olive flowers (EKPA) belongs to the variety Koroneiki and was
collected on 13/5/17 from the area of the National and Kapodistrian
University of Athens. The samples were collected under conducive
conditions (hot and dry weather). Drying of the samples was carried
out for 20 days in a dry, dark, and well-ventilated room. At the
end of 20 days, the plant material was stored in the herbarium.
Subsequently, 20 g of each dry sample were extracted
with 150 ml of EtOH/H2O (50:50 v/v) for 30 min in an
ultrasonic bath (Branson 2510). The above procedure was repeated
twice for each sample. Subsequently, vacuum filtration was carried
out and evaporation at 40°C in RotaVapor until EtOH removal. The
extracts were initially stored in the freezer at −80°C for 24 h and
then lyophilized on a Christ Alpha 1–5 lyophilizer (Martin Christ
GmbH and CoHG).
HPLC analysis of the extracts
HPLC device (Thermo Finnigan) was used for the
qualitative and related quantitative analysis of the extracts and
comprised a SpectraSystem P4000 pump, a SpectraSystem 1000
Degasser, a SpectraSystem AS3000 automatic sampling probe, and
SpectraSystem UV6000LP detection probes (PDAs). Subdivision of the
substances was performed on a Supelcosil RP-18 C18 chromatographic
layer 25 cm × 4.6 mm i.d., 5.0 μm (Discovery). The mobile
phases used were water with acetic acid (0.1%) (phase A) and
acetonitrile with MeOH (2%) (phase B). The solvent gradient changed
according to the following conditions: from 0 to 15 min, 95% (A):
5% (B) to 85% (A): 15% (B); from 15 to 40 min, 85% (A): 15% (B) to
55% (A): 45% (B); from 40 to 50 min, 55% (A): 45% (B) to 5% (A):
95% (B); from 50 to 55 min, 5% (A): 95% (B) to 5% (A): 95% (B);
from 55 to 56 min, 5% (A): 95% (B) to 95% (A): 5% (B); from 56 to
60 min, 95% (A): 5% (B) to 95% (A): 5% (B). The flow of the mobile
phase was set at 1 ml/min, and the injection volume of the samples
was set to 10 μl. The detection of the eluted metabolites
was performed using a PDA detector (254, 280 and 355 nm). For the
related quantitative analysis 10 mg of each extract was diluted in
1 ml of MeOH and the samples were analyzed in triplicate. The
chromatogram analysis and the peak area calculation were performed
at 254 nm.
Assays of in vitro redox biomarkers
Determination of the total phenolic
content (TPC)
Total phenolic content was determined using
Folin-Ciocalteu colorimetric method as presented previously by
Blainski et al (12),
according to which: 10 mg of gallic acid (97% purity) was dissolved
in 1 ml of DMSO and serial solutions of decreasing concentration
were prepared (1, 0.8, 0.7, 0.5, 0.4, 0.3, 0.2 and 0.1 mg/ml).
After the addition of the Folin-Ciocalteu reagent, absorbance of
the samples was measured at 765 nm using a spectrophotometer (TECAN
Infinite M200 Pro UV/Vis Reader). For the construction of the
reference curve, the absorbances corresponding to the linear region
of the curve were selected (y=0.0479× +0.2653, R2=
0.9996). Then, 10 mg of each extract was dissolved in 1 ml of DMSO
and serial solutions of decreasing concentration were prepared (5,
2.5 and 1.25 mg/ml). In each cell of the 96-well plate was
transferred 25 µl of each sample dissolved in DMSO, 125 µl of
Folin-Ciocalteu solution (2.5 ml of distilled H2O
contains 0.25 ml of Folin-Ciocalteu solution reagent) and 100 µl of
7.5% (w/v) aqueous sodium carbonate solution that acts as a
promoter of the reaction. After appropriate agitation the samples
remained in the dark for 30 min at 25°C and the absorption was
measured by a spectrophotometer set at 765 nm (TECAN Infinite M200
Pro UV/Vis Reader). The total phenolic content was expressed as
milligrams of Gallic Acid (GA) equivalent per gram of the olive
flower extract (y=0.0479× +0.2653, R2= 0.9996) (15).
2,2-Diphenyl-1-picrylhydrazyl (DPPH)
radical scavenging assay
The radical scavenging capacity (RSC) of the blossom
extracts was evaluated using the DPPH• assay (13) with slight modifications, as previously
described (14,15). Briefly, 1 ml of freshly prepared
methanolic solution of DPPH• (100 μΜ) was mixed with the tested
extracts at various concentrations, ranging between 2.5 and 100 μg
per extract, (ODsample). After 20 min of incubation in the dark the
absorbance was monitored at 517 nm on a Hitachi U-1900 radio beam
spectrophotometer (serial no. 2023–029; Hitachi). MeOH was used as
a blank and DPPH alone in MeOH was used as the control (ODcontrol).
The percentage RSC of the tested extracts was calculated using the
equation: RCS% = (ODcontrol-ODsampleODcontrol) ×100.
ABTS•+ radical scavenging
assay
The ABTS•+ RSC of the tested extracts was
determined as previously described by Cano et al (16), with minor modifications (14). Briefly, 1 ml of the reaction mixture
containing ABTS•+ (1 mM), H2O2 (30
μM) and horseradish peroxidase (6 μM) in 50 mM PBS
(pH=7.5) was prepared in distilled water (dH2O).
Following incubation for 45 min in the dark, 10 μl of the
tested extracts, at various concentrations, ranging from 2.5 to 50
μg per extract, was added (ODsample) and the absorbance at
730 nm was read on a Hitachi U-1900 radio beam spectrophotometer
(serial no. 2023–029; Hitachi). In each experiment, a blank without
the peroxidase was used, while the ABTS•+ radical
solution without the extract was used as the control (ODcontrol).
The RSC percentage was determined using the same equation as that
described for the DPPH assay.
Evaluation of the antimutagenic
capacity of the extracts using the Ames test
To evaluate the antimutagenic capacity of the tested
extracts we applied the Ames test using the bacterium strain
Salmonella typhimurium TA102 (MolTox) according to Maron and Ames
(17). In brief, 700 μl of the
bacterium culture were used to inoculate 30 ml of autoclaved Oxoid
nutrient broth no. 2. The cultures were placed on a vibrator (100
rpm) and incubated in the dark at 37°C until the cells reached a
density of 1–2×109 colony forming units (CFU/ml,
OD540 between 0.1 and 0.2). The following substances
were then added in the sterile tubes: Plates with oxidant + the
tested compound; 2 ml of top agar, 100 μl of the bacterial
culture, 50 μl of tert-butyl hydroperoxide (0.4 mM) and 50
μl of each extract at various concentrations, ranging from 2
to 32 μg per extract/plate. In addition, a plate with the
oxidizing agent alone and a plate without the oxidizing agent or
the tested compound were used as positive and negative controls.
Moreover, each extract was examined at the two highest
concentrations used in the assay for putative induction of
mutations. The aforementioned tubes were poured onto plates covered
by glucose minimal agar and incubated at 37±2°C for 48 h. Then, the
histidine revertant colonies (His+) were counted. The number of
induced revertants was obtained by subtracting the number of
spontaneous revertants from the number of revertants on the plates
with the mutagen and/or antioxidant. The percentage inhibition of
mutagenicity was calculated as: inhibition = no. of colonies per
plate with oxidant + tested compound number of colonies per plate
with oxidant alone ×100.
Evaluation of the antigenotoxicity of
the extracts using the DNA relaxation assay
The DNA relaxation assay has been previously
described (18). The principle of
this assay is dependent on the conformational changes of the
plasmid (pBluescript-SK+, Fermentas) DNA, which natively exists in
the supercoiled conformation but after a single-strand break is
converted to an open circular one. Based on this, the protective
activity of the olive blossom extracts against DNA single-strand
breaks by 2,2′-azobis AAPH (2.5 mM) were assessed. Specifically, in
a total reaction volume of 10 μl, 2 µl of DNA (4 µg/ml) was
mixed with PBS, AAPH and different concentrations of the tested
extracts ranging between 1 and 300 μg/ml and the mixture was
incubated at 37°C for 45 min. For each assay, a negative control
(DNA without the tested compounds and AAPH) and a positive control
(DNA with AAPH and without the tested compounds) were used.
Moreover, the maximum tested concentrations were mixed alone with
DNA for possible induction of DNA strand breaks. However, none of
the tested concentrations were found to induce DNA breaks.
Subsequently, 3 µl of loading buffer (bromophenol blue 0.25%+30%
glycerol) was added and the samples were loaded on a 0.8% agarose
gel, following electrophoresis at 70 V for 60 min. Eventually, the
gel was stained with 12.5 µl of ethidium bromide (10 mg/ml) in 250
ml of dH2O for 30 min, and then washed with 250 ml of
dH2O for another 30 min. Finally, the gel was exposed to
UV, the MultiImage Light Cabinet (Alpha Innotech) was used to
capture the gel images and the results were analyzed with the Alpha
View suite.
Cell culture experiment
According to the international guidelines on good
cell culture practice (19), the cell
lines used in this research were checked for mycoplasma, using PCR.
According to PCR results the tested cell lines, were mycoplasma
free. Furthermore, a morphology check, both at high and low culture
densities via microscope were conducted to authenticate the state
of cells, through their phenotypic characteristics. Finally, the
passage number for each cell line did not exceed the 30 population
doublings.
Cell culture conditions
The cervical cancer (HeLa), murine myoblasts (C2C12)
and the liver cancer (HepG2) cells were cultured at 37°C in 5%
CO2 in Dulbecco's modified Eagle's medium (DMEM)
containing fetal bovine serum (FBS) (10% v/v), L-glutamine (2 mM),
penicillin (100 U/ml) and streptomycin (100 U/ml). The endothelial
cells (EA.hy926) were cultured at 37°C in 5% CO2 in DMEM
containing FBS (10% v/v), HEPES (25 mM), L-glutamine (2 mM),
penicillin (100 U/ml) and streptomycin (100 U/ml).
HeLa and HepG2 cell lines were donated by Assistant
Professor Kalliopi Liadaki (Department of Biochemistry and
Biotechnology, University of Thessaly, Larissa, Greece). The C2C12
myoblasts were donated by Professor Koutsilieris (National and
Kapodistrian University of Athens, Athens, Greece). Finally, the
EA.hy926 cells were donated by Professor Koukoulis (University
Hospital of Thessaly, Larissa, Greece).
Cell viability assay
The working concentrations of the tested extracts
did not induce cytotoxicity in any cell line. In order to check
which concentrations of the extracts were cytotoxic (i.e., which of
them compromised cell viability) the XTT assay kit was used,
according to the manufacturer's protocol.
Treatment of the cell lines with the
extracts
The cells of each cell line were incubated in
culture medium in flasks (25 m2) for 24 h. The medium
was then removed and serum-free medium containing the tested
extracts at non-cytotoxic concentrations was re-added in the
flasks. The treatment of the cells with the extracts (or with the
serum-free medium only for the control cells) lasted 24 h.
Subsequently, they were trypsinised, collected, centrifuged (300 ×
g, 10 min, 5°C) and the supernatant fluid was discarded. The
cellular pellet was re-suspended in PBS.
Measurement of endogenous GSH and ROS
levels in cell lines using flow cytometry
The intracellular GSH and ROS levels were assessed
using the fluorescent dyes mercury orange and DCF-DA, respectively
(20). A 400 μM stock solution
of mercury orange was prepared in acetone and a 400 μM stock
solution of DCF-DA was prepared in MeOH. The cell pellet was
resuspended in PBS at the concentration of 1×106
cells/ml and incubated with mercury orange (40 μΜ) or DCF-DA
(10 μΜ) at 37°C for 30 min. The cells were then washed and
re-suspended in PBS and subjected to analysis using a FACSCalibur
flow cytometer (BD Biosciences) with excitation and emission
wavelengths at 488 and 530 nm for ROS and at 488 and 580 nm for
GSH. The cells were analyzed at a flow rate of 1,000 events/sec.
Analyses were performed on 10,000 cells per sample and the
fluorescence intensities were measured on a logarithmic scale. Data
were analyzed using BD Cell Quest software (BD Biosciences).
Statistical analysis
SPSS version 21.0 was used (SPSS Inc., Chicago, IL,
USA) for data analysis. All data were analyzed using one-way ANOVA
followed by Tukey's test for multiple pair wise comparisons. Each
experiment was repeated at least 3 times. Data are presented as
mean ± standard error of the mean (SEM). The significance level was
set at P<0.05, and the subset of alpha level was at 0.05. A
bivariate Spearman's correlation was conducted to correlate the
total polyphenolic content (TPC) of the extracts with the four
assays tested, DPPH, ABTS, plasmid relaxation assay and Ames
test.
Results
HPLC analysis
The qualitative HPLC analysis of the olive flower
hydroalcoholic extracts revealed that the major compounds belong in
two chemical categories, secoiridoid derivatives and flavonoid
derivatives (Fig. 1). Of these,
quercetin-3-O-sophoroside (peak 2, Fig.1A) and oleuropein (peak 6, Fig.1A) are the main representatives of each
category. The comparison study of the extracts showed a high
similarity of the chemical composition of AGRI, EKPA and ANKT
samples and only small differences on the quantities of the major
compounds were observed. On the other hand, KTKT sample appeared to
be poorer in terms of chemical composition while the main
compounds, quercetin-3-O-sophoroside and oleuropein were present in
small quantities. In more detail, the related quantification
analysis revealed that all extracts contain similar concentration
of the compound 1 (peak 1) in contrast to other compounds where
significant variations were observed between the different extracts
(Fig. 1B). Regarding the
quercetin-3-O-sophoroside, AGRI extract contains the highest amount
similar to EKPA and ANKT extracts while KTKT contains significantly
lower levels than the other three extracts. By contrast, oleuropein
was found in greater quantities in the EKPA extract followed by
AGRI extract (similar concentration) and ANKT while KTKT contains
low amount of oleuropein (Fig.
1B).
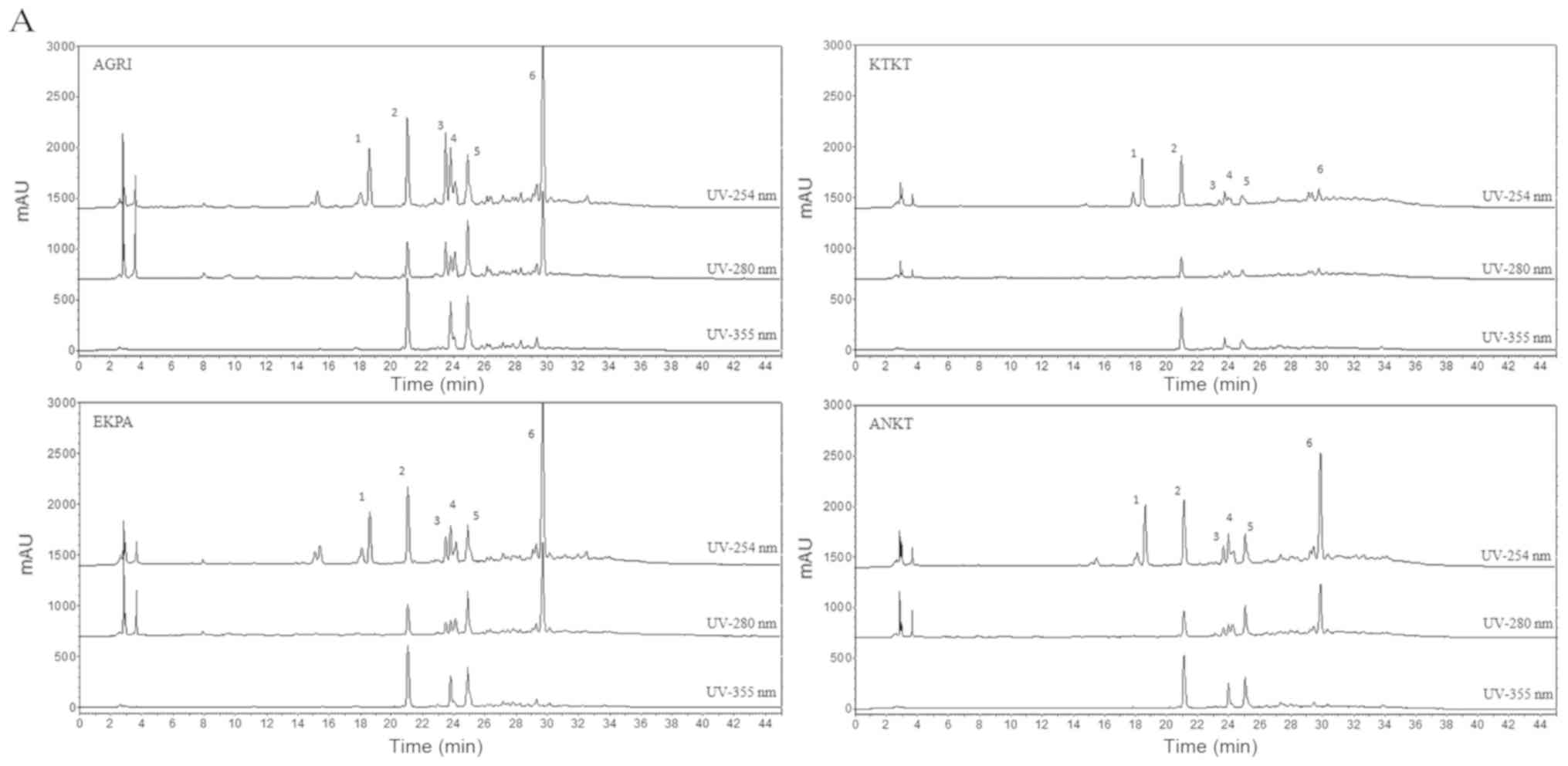 | Figure 1.(A) HPLC chromatograms of the tested
hydroalcoholic extracts (AGRI, KTKT, EKPA and ANKT) at 254 nm
(upper line), 280 nm (middle line) and 365 nm (lower line).
Compounds: 1, secoiridoid derivative; 2, quercetin-3-O-sophoroside;
3, secoiridoid derivative; 4, rutin; 5, flavonoid derivative; 6,
oleuropein. (B) Related quantification analysis of the major
components of olive flower extracts using HPLC at 254 nm. Peak 1,
secoiridoid derivative; Peak 2, quercetin-3-O-sophoroside; Peak 3,
secoiridoid derivative; Peak 4, rutin; Peak 5, flavonoid
derivative; Peak 6, oleuropein HPLC, high-performance liquid
chromatography. |
IC50 values of the extracts
in the DPPH• and ABTS•+ assays and total
phenolic content (TPC) measurement
As indicated by the results regarding the
DPPH• and ABTS•+ assays, all the extracts
exhibited antioxidant activity. Specifically, regarding the
DPPH• assay the IC50 values of AGRI, KTKT,
EKPA and ANKT were equal to 40.50, 73.25, 50.75 and 73.25 μg
of extract, respectively (Table I).
In detail, statistical analysis revealed that AGRI exerted a
stronger antioxidant activity compared with KTKT (P=0.002) and ANKT
(P=0.002). By contrast, EKPA was more potent than KTKT (P=0.003)
and ANKT (P=0.003). Moreover, as assessed by the ABTS•+
assay the AGRI, KTKT, EKPA and ANKT extracts exhibited
IC50 values equal to 9.25, 15.77, 32.88 and 23.06
μg of extract, respectively (Table
I). From the obtained results it seems that all extracts
exhibited a statistically significant difference (P=0.0001).
IC50 values represent the amount of tested compounds
required for 50% reduction of the two radicals. Given that the
lower the IC50 value the more powerful antioxidant
activity, our results revealed that in both assays AGRI exerted the
strongest antioxidant activity, compared with the remaining three
(i.e., KTKT, EKPA and ANKT). Total phenolic content was estimated
in four olive flower extracts, expressed in mg of gallic acid/g of
extract. Estimated values varied from 50.92 to 81.03 mg of gallic
acid/g extract (Table I), showing a
significant difference in the phenolic content among the extracts
tested. AGRI extract exhibited the highest phenolic content (81.03)
followed by EKPA (76.15) and ANKT (66.06) extracts while KTKT
exhibited the lowest value (50.92). It is noteworthy that the
calculated TPC values of the extracts are in accordance with the
corresponding antioxidant capacity expressed by DPPH values.
 | Table I.IC50 values of the
extracts in the DPPH• and ABTS•+ assays and
their total phenolic content (TPC) expressed as mg of Gallic Acid
(GA) equivalent per g of extract. |
Table I.
IC50 values of the
extracts in the DPPH• and ABTS•+ assays and
their total phenolic content (TPC) expressed as mg of Gallic Acid
(GA) equivalent per g of extract.
|
|
DPPH• |
ABTS•+ | TPC |
|---|
|
|
|
|
|
|---|
| Extracts | IC50
(μg/ml) | IC50
(μg/ml) | mgGA/g extract |
|---|
| AGRI |
40.50±2.66a |
9.25±0.78a | 81.03 |
| KTKT |
73.25±1.29b |
15.77±0.23b | 50.92 |
| EKPA |
50.75±2.16a |
32.88±0.19c | 76.15 |
| ANKT |
73.25±1.49b |
23.06±0.19d | 66.06 |
Antigenotoxic and antimutagenic
activity of the tested extracts as assessed by the plasmid
relaxation assay and the Ames test
The obtained results from the plasmid relaxation
assay revealed that AGRI (P=0.003), EKPA (P=0.019) and ANKT
(P=0.005) possess equal antigenotoxic activity and were more prone
to protect the plasmid DNA from lesions compared with KTKT
(Table II). Specifically, the
IC50 values of AGRI, KTKT, EKPA and ANKT were calculated
at 1.717, 8.233, 2.85 and 2.17 µg/µl, respectively. The same
pattern was also observed in the Ames test. More specifically, the
IC50 values of AGRI, KTKT, EKPA and ANKT were calculated
at 3.33, 4.11, 3.09 and 2.79 μg of extract, respectively
(Table II). Thus, AGRI (P=0.043),
EKPA (P=0.011) and ANKT (P=0.002) possess equal antimutagenic
activity compared with KTKT.
| Table II.Antigenotoxic and antimutagenic
activity of the tested extracts.a |
Table II.
Antigenotoxic and antimutagenic
activity of the tested extracts.a
|
| Plasmid relaxation
assay | Ames test |
|---|
|
|
|
|
|---|
| Extracts | IC50
(μg/μl) | IC50
μg extract/plate |
|---|
| AGRI |
1.717±0.27b |
3.33±0.30b |
| KTKT |
8.233±9.62c |
4.11±0.09c |
| EKPA |
2.850±9.62b |
3.09±0.2b |
| ANKT |
2.117±2.24b |
2.79±0.17b |
Amounts where the tested extracts
exhibited cytotoxicity as assessed by the XTT assay
The antioxidant capacity of the tested extracts was
measured in four cell lines: EA.hy926, HeLa, and HepG2 cells, as
well as C2C12 myoblasts. Prior to examining the potential
antioxidant activity of the olive blossom extracts in cell culture,
the concentration threshold above which the tested compounds
exhibited cytotoxic effects in the cell lines was investigated. A
range of amounts for each extract between 1.0 and 100.0 μg
of extract was administered to the cells. The results from the XTT
assay indicated that AGRI was more cytotoxic in EA.hy926 and HepG2
cells, exhibiting cytotoxicity at 10 μg of extract.
Additionally, ANKT exhibited cytotoxicity at 25.0 μg in
C2C12 myoblasts. Finally, the cytotoxicity level was observed at
25.0 μg for EKPA in HeLa cells (Table III). The obtained results revealed a
tissue-specific activity of the extracts. Moreover, EA.hy926 seems
to be the most sensitive cell line compared to the remaining cell
lines, since the majority of the extracts induced cytotoxicity at
low concentrations.
| Table III.Amounts where the tested extracts
exhibited cytotoxicity, as assessed by the XTT assay. |
Table III.
Amounts where the tested extracts
exhibited cytotoxicity, as assessed by the XTT assay.
| Cell lines | Extracts | Cytotoxic amount
(μg/ml) |
|---|
| EA.hy926 | AGRI | 10 |
|
| KTKT | 25 |
|
| EKPA | 50 |
|
| ANKT | 100 |
| C2C12 | AGRI | 70 |
|
| KTKT | 60 |
|
| EKPA | 100 |
|
| ANKT | 25 |
| HeLa | AGRI | 50 |
|
| KTKT | 100 |
|
| EKPA | 25 |
|
| ANKT | 100 |
| HepG2 | AGRI | 10 |
|
| KTKT | 100 |
|
| EKPA | 25 |
|
| ANKT | 100 |
GHS and ROS levels in EA.hy926
cells
The results obtained from flow cytometry revealed
that all tested extracts significantly increased GSH levels
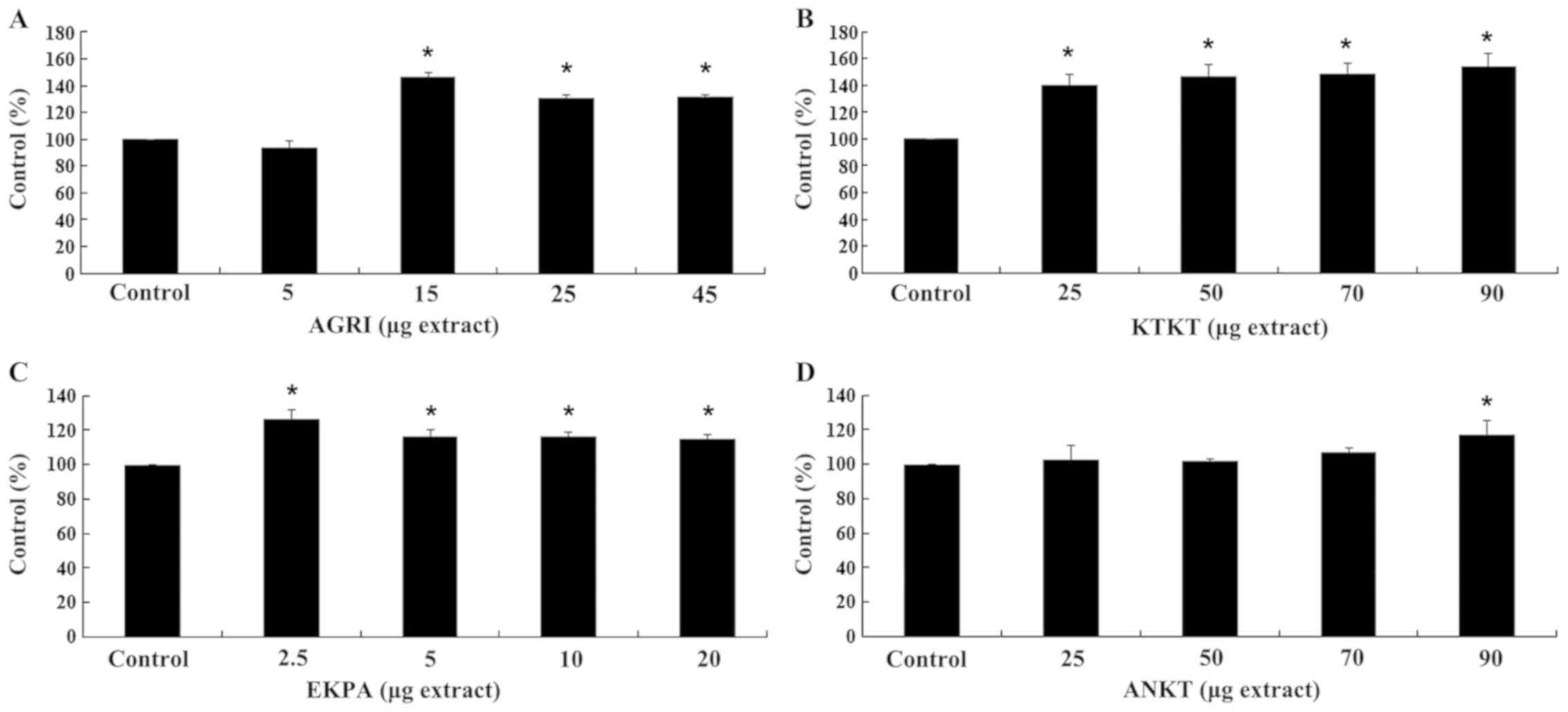
compared with the control in the four cell lines (Figs. 2, 4,
6 and 8). However, the corresponding ROS levels
were not uniformly accompanied by statistical alterations (Figs. 3, 5,
7 and 9). Specifically, the AGRI increased the GSH
levels of the EA.hy926 cells by 24 and 11% at 2.5 and 5 μg
of extract, respectively, compared with the control (Fig. 2A). The ROS levels were decreased by
17% at 2.5 μg AGRI, compared with control (Fig. 3A). The KTKT extract also increased GSH
levels by 18, 20 and 23% at 5, 10 and 20 μg of extract
(Fig. 2B), respectively, while there
was no alteration at ROS levels (Fig.
3B). Additionally, the GSH levels due to the EKPA extract were
elevated by 12, 24, 22 and 25% at 5, 10, 20 and 40 μg of
extract (Fig. 2C), with no
alterations at ROS levels (Fig. 3C).
Finally, ANKT increased GSH levels by 11 and 16% at 50 and 70
μg of extract, respectively (Fig.
2D) with a concomitant decrease at ROS levels by 21% at 70 and
90 μg (Fig. 3D).
GHS and ROS levels in C2C12 cells
According to the obtained results from flow
cytometry in C2C12, GSH levels were elevated after 24 h incubation
with AGRI by 40, 50, 18 and 13% at 10, 25, 50 and 60 μg of
extract, respectively, compared with the control (Fig. 4A), while, ROS levels remained
unaffected (Fig. 5A). Moreover, KTKT
increased GSH levels by 70, 99 and 22% at 10, 25 and 50 μg
of extract, respectively, compared with control (Fig. 4B) with no effect on ROS levels
(Fig. 5B). Additionally, EKPA
increased GSH by 21, 46 and 37% at 50, 60 and 80 μg of
extract, respectively (Fig. 4C) with
a concomitant reduction at ROS levels by 51 and 24% at 60 and 80
μg of EKPA, respectively, compared with control (Fig. 5C). Finally, ANKT also increased GSH
levels by 14, 38, 39 and 44% at 2.5, 5, 10 and 20 μg of
extract, respectively, compared with control (Fig. 4D) and decreased ROS levels by 10 and
12% at 10 and 20 μg of extract (Fig. 5D).
GHS and ROS levels in HeLa cells
Furthermore, after AGRI administration of HeLa
cells, GSH levels were increased by 46, 31 and 32% at 15, 25 and 45
μg of extract compared with control, respectively (Fig. 6A), while ROS levels remained
relatively unaffected (Fig. 7A).
Additionally, KTKT increased GSH by 41, 47, 49 and 55% at 25, 50,
70 and 90 μg of extract, respectively (Fig. 6B). By contrast, ROS levels were not
altered (Fig. 7B). A mild increase at
GSH levels was also observed after EKPA administration for 24 h, by
27, 17, 17 and 15% at 2.5, 5, 10 and 20 μg of extract,
respectively (Fig. 6C) with no
effects on ROS levels (Fig. 7C).
Additionally, ANKT administration increased GSH by 17% at 90
μg of extract compared with control (Fig. 6D). By contrast, ROS levels were
decreased by 32, 28 and 20% at 50, 70 and 90 μg of ANKT,
respectively (Fig. 7D).
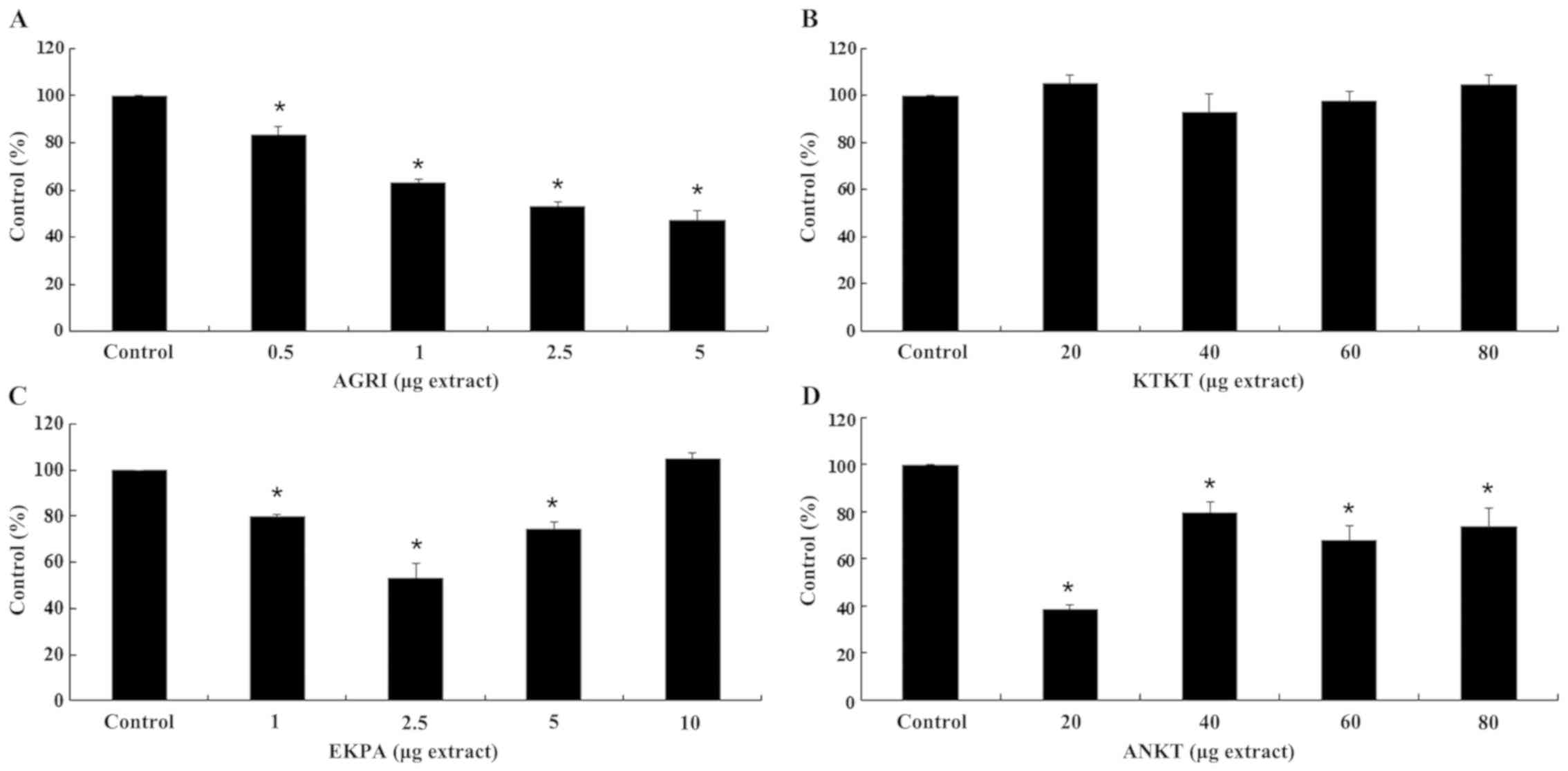
GHS and ROS levels in HepG2 cells
Finally, with respect to the effects on HepG2 cells
(Fig. 8), AGRI, increased GSH levels
by 10, 31 and 19% at 0.5, 1 and 2.5 μg of extract compared
with control, respectively. Nevertheless, at 5 μg of AGRI,
GSH levels were decreased by 12% indicating a pro-oxidant effect
(Fig. 8A). Additionally, ROS levels
were decreased by 16, 37, 47 and 53% after administration of 0.5,
1, 2.5 and 5 μg of AGRI, respectively (Fig. 9A). Moreover, GSH levels were increased
by 31 and 17% at 40 and 60 μg of KTKT, respectively.
However, at 80 μg of KTKT GSH levels were decreased by 32%,
also indicating a pro-oxidant effect (Fig. 8B). Unlike GSH, ROS levels were not
significantly affected (Fig. 9B).
Furthermore, EKPA increased GSH levels at all tested
concentrations. In detail, after administration of 1, 2.5, 5 and 10
μg of EKPA GSH levels were increased by 32, 65, 57 and 46%
compared with control, respectively (Fig.
8C). EKPA administration was accompanied by ROS decrease by 20,
47 and 26% at 1, 2.5 and 5 μg of extract, respectively
(Fig. 9C). Additionally, ANKT
increased GSH levels by 11 and 20% at 60 and 80 μg of
extract, respectively (Fig. 8D),
whereas ROS levels were decreased by 62, 20, 32 and 26% at 20, 40,
60 and 80 μg of ANKT compared with control, respectively
(Fig. 9D).
Discussion
In the present study, we evaluated the antioxidant,
antimutagenic and antigenotoxic effects of four polyphenolic olive
blossom extracts in vitro and in cell culture. Our results
show that all the tested extracts exert a great antioxidant
capacity as assessed by scavenging free radicals (DPPH•,
ABTS•+). In addition, they showed antimutagenic and
antigenotoxic activity and they also have the ability to increase
the endogenous GSH levels with a concomitant decrease in the
endogenous ROS concentration in general. It is worth mentioning
that 20% of olive tree flowers set a mature fruit, whereas the
remaining 80% fall onto the ground without any benefit. It is known
from the literature, and verified from our qualitative HPLC
analysis (Fig. 1) that, olive flowers
contain a large number of bioactive compounds that may be of
benefit to human health (21). Thus,
the potential exploitation of olive blossoms due to their bioactive
role could offer great financial support to producers. Furthermore,
they are considered as sources for natural pharmaceutical products
minimizing the need for industrial production of chemical
compounds.
All extracts exhibited antioxidant activity, which
depicted a correlation between phenol content and DPPH and
ABTS•+ radicals scavenging activity (Table I). The IC50 values of the
olive flower extracts ranged from 40.50 to 73.25 μg of
extract for the DPPH assay and between 9.25 and 32.88 μg of
extract for the ABTS•+ assay. AGRI possessed the lower
IC50 value in the two methods indicating that it is the
most potent between the tested extracts. This is probably due to
the high concentration of phenols and mainly Oleuropein and
flavonoid glucosides (Quercetin-3-O-sophoroside). In previous
studies, olive flower extracts exhibited a strong antioxidant
potential as assessed by the DPPH• and ABTS•+
assays (6,11). The lower IC50 values were
observed in the samples harvested in the last developmental stage
of the flower (August), thus, it seems that the antioxidant
activity was increased according to the developmental stage
(6). Furthermore, DPPH•
scavenging capacity of different olive tree parts was found to be
higher than that of synthetic antioxidant butylated hydroxytoluene
(BHT) [89.16% inhibition of DPPH radical (22)], suggesting the existence of specific
bioactive compounds. Specifically, the percentage of inhibition of
the free radical DPPH• was between 93.75–95.22% for
leaves and 95.43–96.06% for fruits (23,44). In
addition, the percentage of inhibition of the free radical
ABTS•+ ranged between 91.97–92.42% for stems,
58.38–74.55% for fruits, while the same scavenging ability was
observed for the leaves. However, the extracts from olive leaves
possessed the highest antioxidant capacity compared to stems and
fruits (23). Moreover, our previous
studies indicated that the antioxidant capacity of biophenolic
extracts derived from different olive oils depicted IC50
values of 9.25–49.45 μg/ml indicating the significance of the
different biophenolic composition on free radical scavenging
activity of the extracts (25,26). The
aforementioned data indicate the heterogeneous presence of
bioactive compounds between olive tree parts and, therefore, the
variability of their antioxidant capacities (23). The antioxidant activity and the amount
of total phenols present in the extracts suggest that their RSC can
be attributed to the hydroxylated phenolic compounds and in
particular, to the number of hydroxyl substituents in the aromatic
ring, the nature of the substituents at the para or ortho position
(27), as well as to the availability
of phenolic hydrogens (28). These
compounds react with free radicals formed through the autoxidation
process, giving rise to a newborn radical, which is stabilized by
the resonance effect of the aromatic core. Moreover, the synergism
between the antioxidants in the mixture render the antioxidant
capacity dependent both on the concentration and the interaction
between the antioxidants and the structure (6).
The results obtained from the plasmid relaxation
assay, which assessed the protective effect of extracts against the
ROO•-induced single-stranded DNA fragments showed that
AGRI, ANKT and EKPA extracts had statistically significant greater
ability to protect DNA fragmentation, compared to KTKT extract,
exhibiting an IC50 at 1.717, 2.117 and 2.850 μg,
respectively, while KTKT depicted an IC50 at 82.33
μg. AGRI also showed the lowest IC50 value in
this test. In comparison with other studies, biophenolic extracts
derived from olive oils had the ability to protect DNA from lesions
at 1.4–82.3 µg extract per assay (or 0.14–8.23 µg/µl) as assessed
with the plasmid relaxation assay (26). Moreover, the antimutagenic activity of
coffee polyphenols exhibited IC50 values of 51.03–132.29
µg/ml (29). According to the
literature, olive and wild olive polyphenolic extracts possess
potent anticancer properties. Specifically, wild olive extracts
lead to the reduction of liver carcinoma biomarkers (30). By contrast, olive leaf extracts
inhibit the growth and differentiation of leukemia cancer cells
(31). It has also been shown that
olive polyphenols reduce cell proliferation, invasiveness and tumor
growth in cell models of breast cancer (32). Furthermore, olive oil polyphenolic
extracts with different polyphenolic composition, in terms of HT
and tyrosol (T) had the ability to protect DNA damages induced by
ROO•, where the HT-rich ones had greater antigenotoxic
activity (i.e., lower IC50 values) (26). In line with our findings, numerous
studies have reported that olive oil and the by-products derived
from its generation possess both antioxidant and anticancer
properties (4,11,20,25,33–37).
For example, wild leaf extracts promoted the apoptosis of colon
cancer cells (36). In addition,
daily consumption of olive oil appeared to alleviate the
detrimental effects of oxidative stress in DNA stability (37). Extracts from different parts of olive
trees, including flowers, had great antioxidant, antibacterial and
antiallergic abilities (11). It is
noteworthy that the composition of olive leaf extracts is similar
to that of olive oil (34) and of
flower extracts containing polyphenols such as HT, T and oleuropein
(6,31).
The results from the test of mutagenicity (i.e.,
the Ames test) showed that AGRI, EKRA and ANKT extracts have a
similar capacity to protect plasmid DNA from mutations with an
IC50 at 3 μg/well while KTKT depicted
IC50 at 4 μg/well. The results may be explained
due to the time of harvesting of the olive blossoms. Although olive
flowers KTKT and ANKT belong to the same variety (Lianolia) from
the same region, the difference in collection time (5-6/5 and
14–15/5, respectively) resulted in different chemical content of
the hydroalcoholic extracts (Fig. 1A)
and thus in different biological activity of the extracts. Our
results are in agreement with other relevant studies, according to
which polyphenols from natural plant extracts, such as coffee
(29), pomegranate (38) and olives (39) possess strong antimutagenic activities.
According to Spearman's correlation, a statistically significant
(p<0.05) correlation between Folin-Ciocalteu and DPPH was found.
Furthermore, the differences in the extract activities may be due
to their chemical composition, as assessed by the related
quantification analysis (Fig. 1B). It
seems that KTKT possesses less amount of querqetin 3-O-sophoroside,
oleuropein, secoiridoid derivatives, rutin and flavonoid
derivatives. Additionally, Folin-Ciocalteu indicated that it
contains the lowest polyphenolic content, thus being the less
powerful extract. Specifically, KTKT exhibits the lowest
IC50 values at DPPH, plasmid relaxation assay and Ames
Test compared with other extracts, a fact that indicates the
importance of chemical polyphenolic composition to the activity of
plant extracts. Such significant differences in the concentrations
of polyphenols probably reflect the metabolic behavior of olive
flower during the development stage on the basis of the genotype
and the environmental conditions (40).
Regarding the effects of the tested extracts on
myotube redox status, the extracts all increased GSH levels in all
the tested cell lines compared with control. It is worth noting
that in some cases (e.g., AGRI administration at HepG2 cells) when
the concentration of the extract exhibited a threshold value, the
endogenous levels of GSH decreased indicating a pro-oxidative
phenomenon. The pro-oxidant effect caused by polyphenols depends on
several factors, such as their chemical nature, concentration, and
the micro-environmental conditions (e.g., the cell type, the redox
state and the pH value) (25,41). Olive oil polyphenolic extracts also
had the ability to increase GSH levels through the Nrf2 pathway
(25,26,42). In
addition, coffee polyphenols increased GSH both in vitro and
in vivo as assessed in C2C12 and EA.hy296 cell lines
(43) and in different tissues of
Wistar rats (44), respectively,
demonstrating an active role for the transcription factor Nrf2.
With respect to the chemical composition, HPLC
analysis revealed the presence of common secondary metabolites in
all the tested extracts. The AGRI, EKPA and ANKT extracts had
similar absorbance in all tested wavelengths (i.e., 254, 280 and
355 nm) with maximum absorbance at 254 nm and retention time, 15–35
min. The tree main compounds found in these extracts are
Quercetin-3-O-sophoroside (retention time 21.5 min), Rutin
(retention time 23.75 min) and oleuropein (retention time 30.0
min). By contrast, for KTKT extract, which was collected earlier in
comparison to other samples, low absorbance values were depicted at
254 nm while only a trace of oleuropein was found. In a study
focused on olive blossoms from the Tunisia cultivar ‘Chemlali’ it
appeared that the oleuropein levels were increased with a
concomitant increase of flower maturation period (21).
On the basis of our results, all tested olive
flower extracts exhibited potent antioxidant, antimutagenic and
antigenotoxic activities. Furthermore, they improved redox status
at the cellular level as indicated by the enhancement of the GSH
values and the reduced ROS levels. Olive tree flowers are
considered as by-products of olive oil production. Taking into
account that during olive tree cultivation a considerable amount of
olive flowers is generated, the burden of environmental pollution
is high. Given the optimistic findings we present in this study, we
believe that although further relevant in vivo studies are
required, the flower-derived extracts could have high added since
they could be used as antioxidants or as foodstuffs, food additives
and functional food constituents.
Acknowledgements
Not applicable
Funding
The study was funded by the Hellenic General
Secretariat for Research and Technology (GSRT) and the Hellenic
Foundation for Research and Innovation (HFRI) (grant no. 5547).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable request
Authors' contributions
PK, DK and ALS conceived the study. The research
methodology was designed by PK, DK, ALS and AA. Formal analysis of
the data was conducted by PK. Software analysis of data and figures
was conducted by PK, and supervision of the research was conducted
by DK and ALS. Writing of the original draft was undertaken by PK
and AA, and review and editing of the manuscript were carried out
by ASV, ALS and DK. The experimental procedures were conducted by
PK, FT, KV, PM and AA. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable
Competing interests
All the authors declare that they have no competing
interests.
References
|
1
|
Riley FR: Olive oil production on bronze
age crete: Nutritional properties, processing methods and storage
life of Minoan olive oil. Oxford J Archaeol. 21:63–75. 2002.
View Article : Google Scholar
|
|
2
|
Bouknana D, Hammouti B, Jodeh S, Sbaa M
and Lgaz H: Extracts of olive inflorescence flower pre-anthesis, at
anthesis and grain pollen as eco-friendly corrosion inhibitor for
steel in 1M HCl medium. Anal Bioanal Electrochem. 10:751–777.
2018.
|
|
3
|
De Leonardis A, Aretini A, Alfano G,
MacCiola V and Ranalli G: Isolation of a hydroxytyrosol-rich
extract from olive leaves (Olea Europaea L.) and evaluation
of its antioxidant properties and bioactivity. Eur Food Res
Technol. 226:653–659. 2008. View Article : Google Scholar
|
|
4
|
Lafka T-I, Lazou A, Sinanoglou V and Lazos
E: Phenolic Extracts from Wild Olive Leaves and Their Potential as
Edible Oils Antioxidants. Foods. 2:18–31. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pandey KB and Rizvi SI: Plant polyphenols
as dietary antioxidants in human health and disease. Oxid Med Cell
Longev. 2:270–278. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rekik O, Ben Mansour A and Bouaziz M:
Evaluation of phenolic composition and antioxidant activity changes
in olive flowers during development using HPLC/DAD and LC-MS/MS.
Electrophoresis. 39:1663–1672. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lavee S, Rallo L, Rapoport HF and Troncoso
A: The floral biology of the olive: Effect of flower number, type
and distribution on fruitset. Sci Hortic (Amsterdam). 66:149–158.
1996. View Article : Google Scholar
|
|
8
|
Griggs W.H..Hartman H.T..Bradley
M.V..Iwakiri B.T..Whistler JE: Olive polination in Claifornia.
Calif Agric Exp Sta Bull. 869:501975.
|
|
9
|
Fabbri A and Benelli C: Flower bud
induction and differentiation in olive. J Hortic Sci Biotechnol.
75:131–141. 2000. View Article : Google Scholar
|
|
10
|
Cuevas J and Polito VS: The role of
staminate flowers in the breeding system of Olea europaea
(Oleaceae): an andromonoecious, wind-pollinated taxon. Ann Bot.
93:547–553. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kishikawa A, Ashour A, Zhu Q, Yasuda M,
Ishikawa H and Shimizu K: Multiple biological effects of olive oil
by-products such as leaves, stems, flowers, olive milled waste,
fruit pulp, and seeds of the olive plant on skin. Phytother Res.
29:877–886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blainski A, Lopes GC and De Mello JCP:
Application and analysis of the folin ciocalteu method for the
determination of the total phenolic content from limonium
brasiliense L. Molecules. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brand-Williams W, Cuvelier ME and Berset
C: Use of a free radical method to evaluate antioxidant activity.
LWT - Food Sci Technol. 28:25–30. 1995. View Article : Google Scholar
|
|
14
|
Kouka P, Priftis A, Stagos D, et al:
Assessment of the antioxidant activity of an olive oil total
polyphenolic fraction and hydroxytyrosol from a Greek Olea europea
variety in endothelial cells and myoblasts. Int J Mol Med.
40:703–712. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Veskoukis A, Kerasioti E, Priftis A, Kouka
P, Spanidis Y, Makri S and Kouretas D: A battery of translational
biomarkers for the assessment of the in vitro and in vivo
antioxidant action of plant polyphenolic compounds: The biomarker
issue. Curr Opin Toxicol. 13:99–109. 2018. View Article : Google Scholar
|
|
16
|
Cano A, Hernández-Ruíz J, García-Cánovas
F, Acosta M and Arnao MB: An end-point method for estimation of the
total antioxidant activity in plant material. Phytochem Anal.
9:196–202. 1998. View Article : Google Scholar
|
|
17
|
Maron DM and Ames BN: Revised methods for
the Salmonella mutagenicity test. Mutat Res. 113:173–215. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Priftis A, Papikinos K, Koukoulanaki M, et
al: Development of an assay to assess genotoxicity by particulate
matter extract. Mol Med Rep. 15:1738–1746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bal-Price A and Coecke S: Guidance on Good
Cell Culture Practice (GCCP). Neuromethods. 56:1–25. 2011.
View Article : Google Scholar
|
|
20
|
Kouka P, Priftis A, Stagos D, et al:
Assessment of the antioxidant activity of an olive oil total
polyphenolic fraction and hydroxytyrosol from a Greek Olea europea
variety in endothelial cells and myoblasts. Int J Mol Med.
40:703–712. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abaza L, Taamalli A, Arraez-Roman D,
Segura-Carretero A, Fernandez-Gutierrerez A, Zarrouk M and Youssef
N Ben: Changes in phenolic composition in olive tree parts
according to development stage. Food Res Int. 100:454–461. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ceylan Y, Usta K, Usta A, Maltas E and
Yildiz S: Evaluation of antioxidant activity, phytochemicals and
ESR Analysis of Lavandula Stoechas. Acta Physica Polonica A.
128:B-483–B-487. 2015. View Article : Google Scholar
|
|
23
|
Brahmi F, Mechri B, Dhibi M and Hammami M:
Variation in antioxidant activity and phenolic content in different
organs of two Tunisian cultivars of Olea europaea L. Acta
Physiol Plant. 36:169–178. 2014. View Article : Google Scholar
|
|
24
|
Moudache M, Colon M, Nerin C and Zaidi F:
Phenolic content and antioxidant activity of olive by-products and
antioxidant film containing olive leaf extract. Food Chem.
212:521–527. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kouka P, Priftis A, Stagos D, et al:
Assessment of the antioxidant activity of an olive oil total
polyphenolic fraction and hydroxytyrosol from a Greek Olea europea
variety in endothelial cells and myoblasts. Int J Mol Med.
40:703–712. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kouka P, Tsakiri G, Tzortzi D, Dimopoulou
S, Sarikaki G, Stathopoulos P, Veskoukis AS and Halabalaki MSA-L:
The poly-phenolic composition of extracts derived from different
greek extra virgin olive oils is correlated with their antioxidant
potency. Oxid Med Cell Longev. 2019:132019. View Article : Google Scholar
|
|
27
|
Cheng Z, Ren J, Li Y, Chang W and Chen Z:
Study on the multiple mechanisms underlying the reaction between
hydroxyl radical and phenolic compounds by qualitative structure
and activity relationship. Bioorg Med Chem. 10:4067–4073. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wright JS, Johnson ER and DiLabio GA:
Predicting the activity of phenolic antioxidants: theoretical
method, analysis of substituent effects, and application to major
families of antioxidants. J Am Chem Soc. 123:1173–1183. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Priftis A, Mitsiou D, Halabalaki M, Ntasi
G, Stagos D, Skaltsounis LA and Kouretas D: Roasting has a distinct
effect on the antimutagenic activity of coffee varieties. Mutat
Res. 829-830:33–42. 2018. View Article : Google Scholar
|
|
30
|
Amereh Z, Hatami N, Shirazi FH, et al:
Cancer chemoprevention by oleaster (Elaeagnus angustifoli L.) fruit
extract in a model of hepatocellular carcinoma induced by
diethylnitrosamine in rats. EXCLI J. 16:1046–1056. 2017.PubMed/NCBI
|
|
31
|
Abaza L, Talorete TP, Yamada P, Kurita Y,
Zarrouk M and Isoda H: Induction of growth inhibition and
differentiation of human leukemia HL-60 cells by a Tunisian gerboui
olive leaf extract. Biosci Biotechnol Biochem. 71:1306–1312. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Akl MR, Ayoub NM, Mohyeldin MM, Busnena
BA, Foudah AI, Liu Y-Y and Sayed KAE: Olive phenolics as c-Met
inhibitors: (−)-Oleocanthal attenuates cell proliferation,
invasiveness, and tumor growth in breast cancer models. PLoS One.
9:e976222014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pampaloni B, Mavilia C, Fabbri S, et al:
In Vitro Effects of Extracts of Extra Virgin Olive Oil on Human
Colon Cancer Cells. Nutr Cancer. 667:1228–1236. 2014. View Article : Google Scholar
|
|
34
|
De Leonardis A, Aretini A, Alfano G,
MacCiola V and Ranalli G: Isolation of a hydroxytyrosol-rich
extract from olive leaves (Olea Europaea L.) and evaluation
of its antioxidant properties and bioactivity. Eur Food Res
Technol. 226:653–659. 2008. View Article : Google Scholar
|
|
35
|
El-Kholy TA, Hilal MA, Al-Abbadi HA,
Serafi AS, Al-Ghamdi AK, Sobhy HM and Richardson JRC: The effect of
extra virgin olive oil and soybean on dna, cytogenicity and some
antioxidant enzymes in rats. Nutrients. 6:2376–2386. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zeriouh W, Nani A, Belarbi M, et al:
Phenolic extract from oleaster (Olea europaea var. Sylvestris)
leaves reduces colon cancer growth and induces caspase-dependent
apoptosis in colon cancer cells via the mitochondrial apoptotic
pathway. PLoS One. 12:1–19. 2017. View Article : Google Scholar
|
|
37
|
Salvini S, Sera F, Caruso D, et al: Daily
consumption of a high-phenol extra-virgin olive oil reduces
oxidative DNA damage in postmenopausal women. Br J Nutr.
95:742–751. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cano-Lamadrid M, Marhuenda-Egea FC,
Hernandez F, Rosas-Burgos EC, Burgos-Hernandez A and
Carbonell-Barrachina AA: Biological Activity of Conventional and
Organic Pomegranate Juices: Antioxidant and Antimutagenic
Potential. Plant Foods Hum Nutr. 71:375–380. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kirkland D, Edwards J, Woehrle T and
Beilstein P: Investigations into the genotoxic potential of olive
extracts. Mutat Res Genet Toxicol Environ Mutagen. 777:17–28. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Leopoldini M, Russo N and Toscano M: The
molecular basis of working mechanism of natural polyphenolic
antioxidants. Food Chem. 2011. View Article : Google Scholar
|
|
41
|
Leon-Gonzalez AJ, Auger C and Schini-Kerth
VB: Pro-oxidant activity of polyphenols and its implication on
cancer chemoprevention and chemotherapy. Biochem Pharmacol.
98:371–380. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kouka P, Chatzieffraimidi G-A, Raftis G,
et al: Antioxidant effects of an olive oil total polyphenolic
fraction from a Greek Olea Europaea variety in different
cell cultures. Phytomedicine. 47:135–142. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Priftis A, Goutzourelas N, Halabalaki M,
et al: Effect of polyphenols from coffee and grape on gene
expression in myoblasts. Mech Ageing Dev. 172:115–122. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Priftis A, Soursou V, Makiou A-S, et al: A
lightly roasted coffee extract improves blood and tissue redox
status in rats through enhancement of GSH biosynthesis. Food Chem
Toxicol. 125:305–312. 2019. View Article : Google Scholar : PubMed/NCBI
|