Introduction
The tumor-specific promoter may be used to study
gene targeting treatment, but the simple or natural promoter does
not always meet the requirements of cancer gene therapy. One reason
for this was that the activity of the partial tumor-specific
promoter was weak, meaning it was difficult to induce a high
expression of therapeutic genes in certain tumor cells. Modifying
or transforming the structure of the promoter is crucial to
accurately locate therapeutic genes and improve its expression in
specific cells. Gene transcription of eukaryotic cells may be
regulated by enhancers. Findings of previous studies revealed that
the transcription activity of certain tumor-specific promoters was
increased by enhancers. Activity of the gancyclovir (GCV) suicide
gene driven by the cytokeratin 19 (CK19) promoter was markedly
improved when the CK19 enhancer was added upstream of the promoter
(1), and the human
prostate-specific antigen (PSA) enhancer increased the expression
of green fluorescent protein (GFP) regulated by the PSA promoter in
prostate cancer cells (2).
Therefore, the use of specific enhancers to modify the
tumor-specific promoter in order to improve its transcription
activity and the expression of therapeutic genes is crucial. Simian
virus 40 (SV40) enhancer has been extensively applied to improve
the activity of promoter, rendering it the most common molecular
tool (3,4).
Heparanase (HPSE) acts as an extracellular matrix
catabolic enzyme that is closely correlated with tumor metastasis,
angiogenesis and other diverse pathological and physiological
processes (5–8). Results of previous studies have
suggested that high levels of HPSE mRNA or its protein were
expressed in most malignant tumors, although this expression was
not found in normal somatic cells (9–12).
Previously, we cloned a 561-bp-long human HPSE gene core promoter
and constructed the eukaryotic expression plasmid pEGFP-Hp using
the promoter (13). The vector was
found to be expressed only in tumor cells following cell
transfection. Additionally, the HPSE promoter was found to exhibit
tumor specificity, and could be used for gene therapy research,
although its activity was insufficient or weak (13). Therefore, modification of the HPSE
promoter in order to improve its transcription activity is
essential. In this study, the SV40 enhancer sequence was amplified,
and inserted into the multiple clone site (MCS) downstream of the
HPSE promoter in the vector pEGFP-Hp in order to construct
recombinant plasmid pEGFP-Hp-SV40e. The transcription activity
prior to and following modification was detected and compared, and
the effect of the SV40 enhancer on the activity and tumor
specificity of HPSE promoter was evaluated. The results obtained
may provide an experimental basis for further research into the
role played by the SV40 enhancer.
Materials and methods
Cell lines, main reagents and
instruments
Normal human umbilical vein endothelial cells
ECV304, human hepatocellular carcinoma cells HepG2, laryngeal
epithelial cells Hep2 and positive control plasmid pEGFP-N1 were
obtained from Yili Company (Hefei, China). The plasmid pEGFP-Hp was
constructed and identified in our previous study (13). The SV40 enhancer-containing vector
pGL3-control was purchased from Shanghai Sangon Biotech Co., Ltd.
(Shanghai, China). PFX, T4 DNA ligase, restriction endonuclease
EcoRI, SalI, XhoI and HindIII were all
purchased from MBI Fermentas (Burlington, Ontario, Canada). The PCR
purification kit, DNA gel extraction kit and a small amount of
plasmid extraction kit were obtained from Daopu Company, Beijing,
China. The cloning vector plasmid pMD18-T simple, DL2,000, DL5,000
and DL15,000 DNA markers, as well as dNTP and PCR buffers were
purchased from Takara, Japan.
Agarose gel, tryptone and yeast extract were
obtained from Oxoid, UK. Cell cultures were performed using 10%
fetal bovine serum, Lipofectamine™ 2000, DMEM and RPMI-1640 media
(Invitrogen, Grand Island, NY, USA). The SV40 enhancer upstream and
downstream primers were synthesized by Shanghai Sangon Biotech Co.,
Ltd. The E. coli strain DH5α was provided by Professor
Xuelong Wang (Department of Parasitology, Anhui Medical University,
Hefei, China). Fluorescence microscopy (Nikon, Japan) and flow
cytometry (Beckman Coulter, Inc., Brea, CA, USA) were used for the
subsequent analysis.
Amplification and purification of SV40
enhancer PCR
According to the GenBank DNA sequence of
pGL3-control (accession no. U47296) and the principles of primer
design, two primers of SV40 enhancer (forward,
5′-ATATGCgaattc CGATGGAGCGGAGAATGGG-3′ and reverse,
5′-GCATGT gtcgacGCTGTGGAATGTGTGTCAGTTAGG-3′) were designed
using primer design software 5.0. Besides protection bases (italic
letters), restriction enzyme cutting site EcoRI and
SalI (lower case letters) were introduced into the 5′ ends
of primers, respectively. The sequence of the SV40 enhancer was
amplified by PCR using 1 μl pGL3-enhancer DNA as a template. The
reaction system was 50 μl. The PCR conditions included initial
denaturation at 94°C for 5 min, 35 cycles of amplification with
subsequent denaturation at 94°C for 30 sec, annealing at 57°C for
50 sec, and extension for 50 sec at 72°C, followed by extension for
10 min at 68°C. Electrophoresis was performed using 1% agarose gel.
The PCR product was purified according to the manufacturer’s
instructions and two-way sequencing was completed by Shanghai
Sangon Biotech Co., Ltd.
Identification of the SV40 enhancer
DH5α competent cells were prepared by the calcium
chloride method. PCR products of SV40 enhancer and pMD18-T simple
were incubated overnight for ligation, and then transformed into
competent cells. Positive colonies were selected and amplified,
plasmid DNA was extracted by the alkaline lysis method and
identified by double digestion with EcoRI and SalI.
After 1% agarose gel electrophoresis, the positive recombinant
plasmid was sent to Shanghai Sangon Biotech Co., Ltd. for
sequencing.
Construction and identification of the
recombinant plasmid pEGFP-Hp-SV40e
The PCR products of SV40 enhancer were digested by
double digestion with EcoRI and SalI. The digested
PCR product was analysed using agarose gel electrophoresis and the
large fragment was retrieved. The purified product was the DNA
fragment of SV40 enhancer. Plasmid pEGFP-Hp was also digested by
double digestion and the large fragment was retrieved. During a
ligation reaction, the two recycled products were incubated
overnight at 4°C to insert the DNA fragment of the SV40 enhancer
into MCS of the pEGFP-Hp vector. After competent cell suspension
was thawed at room temperature, ligation products were added for
the transformation test. The recombinant plasmid was screened, and
the plasmid DNA was extracted by the alkaline lysis method, then
identified by single digestion with BamHI, PCR amplification
and DNA sequencing, respectively. The positive recombinant plasmid
was designated as pEGFP-Hp-SV40e.
Functional analysis of the recombinant
plasmid vector pEGFP-Hp-SV40e
ECV, HepG2, Hep2 and K562 cells were cultured in
96-well plates in RPMI-1640 cell culture medium with 10% fetal calf
serum (FCS) and incubated at 37°C in a humidified atmosphere
containing 5% CO2. For each well, at 70–80% confluence,
cells were inoculated into 3 wells at a density of
5×105–1×106 cells/well and transfected with
the plasmid of pEGFP-Hp, positive control plasmid pEGFP-N1 and
recombinant plasmid pEGFP-Hp-SV40e, respectively. The procedure was
undertaken according to the instructions for Lipofectamine™ 2000.
The cells were subsequently cultured for 48 h, and the suspension
cells were collected. The adherent cells were digested by trypsin
(0.2%). The digestion was terminated by adding blood serum culture
medium. Fluorescence intensity was observed using fluorescence
microscopy, and the percentage of light-emitting cell and
transfection efficiency were determined by flow cytometry.
Experiments were repeated at least 3 times.
Results
PCR amplification of SV40 enhancer
Agarose gel electrophoresis of PCR products of the
SV40 enhancer showed a specific band of 250–500 bp, which was
consistent with the theoretical length of 261 bp, including 24-bp
enzyme cutting sites and protection bases (Fig. 1).
Identification of SV40 enhancer
After PCR amplification of positive colonies, the
electrophoretogram also showed a 261-bp-long band (Fig. 2). The sequencing results of the
identified positive plasmid showed that the 237 bp sequence from 46
to 282 bp was consistent with the GenBank database of pGL3-control
(accession no. U47296). The sequences from 40 to 45 bp and from 283
to 288 bp were restriction enzyme cutting sites of EcoRI and
SalI, respectively. No base transition, transversion,
insertion and deletion were found (Fig.
3).
Construction and identification of the
recombinant plasmid pEGFP-Hp-SV40e
The lengths of previous plasmid pEGFPHp and SV40
enhancer were 4,673 and 261 bp, respectively. Following digestion
with EcoRI and SalI, two fragments were generated,
however, the electrophoretogram only showed the longer fragments,
of 4,659 and 247 bp, respectively. Since the cut 14-bp nucleotides
moved out of the gel, it could not be found in the
electrophoretogram (Fig. 4). By
using the two long fragments, recombinant plasmid pEGFP-Hp-SV40e
was constructed.
Recombinant plasmid pEGFP-Hp-SV40e was identified by
double digestion with EcoRI and/or SalI. Two
fragments were detected in the electrophoresis when the recombinant
plasmid pEGFP-Hp-SV40e was digested by double digestion with
EcoRI and SalI. Only a single fragment was found if
the plasmid was digested with EcoRI or SalI. This
finding proved that the recombinant plasmid was correctly
constructed. When plasmid pEGFP-Hp-SV40e was used as a template,
the SV40 enhancer fragment was amplified, and the sequencing
results were consistent with those of the GenBank data. Following
reverse sequencing, complementary sequences of the recombinant
plasmid pEGFP-Hp-SV40e were obtained using Chromas Lite software.
The plasmid contained a complete 237-bp SV40 enhancer sequence
corresponding with the bases between 65 and 301 bp in the
sequencing graph (Fig. 5). The
sequences of 59–64 and 302–307 bp were restriction enzyme cutting
sites of SalI and EcoRI, respectively. The downstream
sequence of the HindIII site was the DNA sequence of HPSE
promoter in the vector. These findings showed that the SV40
enhancer sequence was properly inserted downstream of the HPSE
promoter in plasmid pEGFP-Hp and that recombinant plasmid
pEGFP-Hp-SV40e was successfully constructed.
Functional identification of the
recombinant plasmid pEGFP-Hp-SV40e
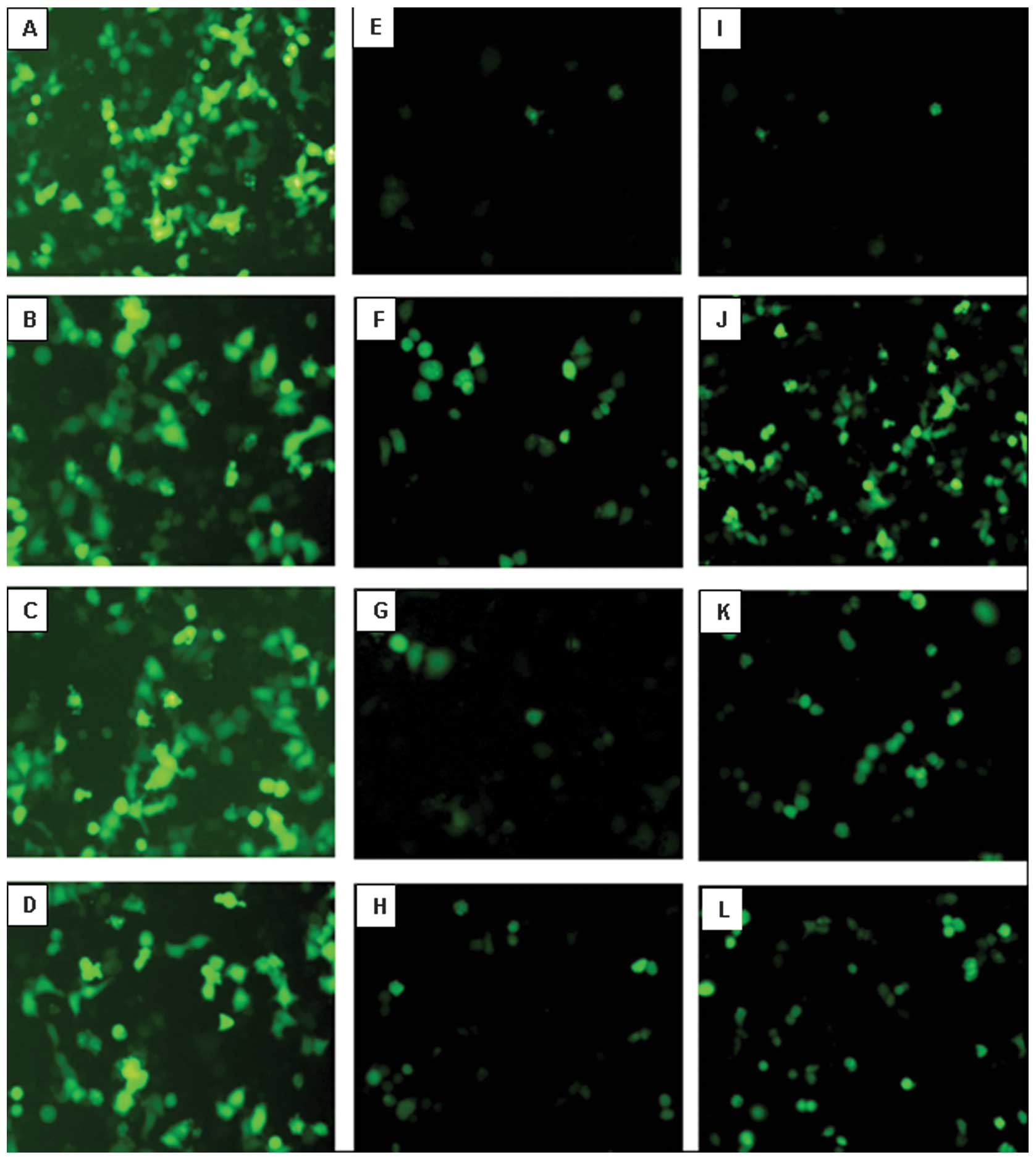
Fluorescence microscopy showed that the fluorescence
intensity of plasmid pEGFP-Hp was the weakest in normal ECV cells
and enhanced in human tumor cell lines HepG2, Hep2 and K562.
Positive control of plasmid pEGFP-N1 showed a strong fluorescence
expression in all of the cells. The fluorescence intensity of
recombinant plasmid pEGFP-Hp-SV40e in normal ECV cells was weak
while the fluorescence intensity in the 3 types of tumor cells was
stronger than that of pEGFP-Hp and similar to pEGFP-N1 (Fig. 6). Flow cytometric analysis detected
the percentage of light-emitting cells of plasmid pEGFP-Hp to be
only 4.1±0.3% in ECV cells; and 17.2±1.9, 8.8±1.0 and 6.4±0.5% in
HepG2, Hep2 and K562 cells, respectively. The positive control
plasmid pEGFP-Hp was found to have higher transfection rates in all
cells and the percentages of light-emitting cells were 18.3±1.9,
29.3±3.0, 17.0±1.8 and 13.0±1.2%, in ECV, HepG2, Hep2 and K562
cells, respectively, suggesting that it also had a high activity in
normal ECV cells. The light-emitting cell percentages of
recombinant plasmid pEGFP-Hp-SV40e were 4.3±0.4, 28.8±2.9, 16.4±1.5
and 11.7±1.2%, in ECV, HepG2, Hep2 and K562 cells, respectively.
The ratios of transfection rates of pEGFP-Hp-SV40e vs. pEGFP-Hp
were 1.05, 1.67, 1.86 and 1.83, in the four cell lines,
respectively. These findings suggest that inserting SV40 enhancer
did not affect the transfection efficiency of pEGFP-Hp in normal
ECV cells, but that it is capable of increasing HPSE promoter
activity in tumor cells.
Discussion
Human HPSE gene was cloned and identified almost at
the same time in 1999 by several research teams (5–8). The
gene, located on chromosome 4q22, includes 14 exons separated by 13
introns. The complete cDNA of HPSE is 1,758-bp long, and contains
an open reading frame encoding a polypeptide of 543 amino acids
with a calculated molecular weight of 61,192 Da. A number of
studies have suggested that high levels of HPSE mRNA or protein
were expressed in most malignant tumors, but no expression was
detected in normal somatic cells (5–11). By
degrading glycosaminoglycans, whose main component is heparan
sulfate proteoglycans in extracellular matrix, HPSE plays an
important role in tumor metastasis and vascularization. In recent
years, HPSE has become a new therapeutic target for cancer
biotherapy (14,15).
The core sequence of the HPSE promoter was cloned in
our previous experiment (13).
Results of the sequence analysis showed that it contained the
reported transcription factor binding site (TFBS) (16,17) as
well as the 6 E-box sites. By inserting the HPSE core promoter, we
constructed a plasmid pEGFP-Hp driven by the HPSE promoter and
transfected it into normal and tumor cell lines. Plasmid pEGFP-Hp
was found to be expressed only in tumor cell lines and not in
normal cell lines. This finding suggested that the HPSE promoter
was a broad-spectrum tumor peculiar promoter and could be used as a
new available tool for oncogene-targeted therapy (13). However, the detection result of
microscopy and flow cytometry indicated that the GFP expression
levels of pEGFP-Hp in tumor cells were lower than those of
pEGFP-N1, which contains a cytomegalovirus promoter (13). The phenomenon was also observed and
verified in this study. Therefore, improving the activity of HPSE
promoter without changing its tumor specificity is essential.
Enhancers are usually cis-acting elements that could
significantly improve the transcription efficiency of homologous or
heterologous genes. Enhancer may be located upstream, downstream or
within the target gene that it regulates, even at a distance from
the transcription starting point. The SV40 enhancer was one of the
first enhancers to exhibit strong activity. It is composed of a 72
bp repeated sequence without obvious partiality in race and can
exert a 2- to 20-fold enhancement of gene transcription in
different host cells (18,19). Previous results have shown that the
transcription activity of human telomerase reverse transcriptase
(hTERT) promoter was significantly enhanced after it was modified
using SV40 enhancer (20). The
mechanism of SV40 enhancer in gene regulation was associated with
its Z-DNA structure. Alternating purine and pyrimidine were also
detected in its repeated sequence allowing the formation of a
rotating DNA structure that binds with a specific protein for
transcription (21). Furthermore,
SV40 enhancer promoted the ingression of plasmid vector from the
cytoplasm to the nucleus to potentialize the nucleic localization
of DNA that was sequence specific. For these reasons, SV40 enhancer
could be applied to promote the transfection efficiency and
expression of therapeutic genes.
In this study, the SV40 enhancer sequence was
amplified by using the plasmid pGL3 as a template, which contained
the SV40 enhancer. The sequencing result was in accordance with the
GenBank data (accession no. U47296), although no insertion,
deletion or substitution was evident. The result verified that the
amplified SV40 enhancer sequence was correct. The plasmid
pEGFP-Hp-SV40e was constructed using the amplified SV40 enhancer
sequence and plasmid pEGFP-Hp, and then identified using enzyme
digestion. The 237-bp SV40 enhancer was found to be successfully
inserted into the appointed MCS of pEGFP-Hp by sequencing. It
further proved the construction of recombinant plasmid
pEGFP-Hp-SV40e was successful and consistent with the principle of
molecular biology.
Following the transfection of recombinant plasmid
pEGFPHp-SV40e into diverse cells, stronger fluorescence was found
in 3 types of human tumor cells, using fluorescent microscopy,
which was similar to the positive control plasmid pEGFP-N1. The
fluorescence intensity of plasmid pEGFP-Hp was obviously weaker
than the other two plasmids. Results of flow cytometry showed that
the transfection efficiency of recombinant plasmid pEGFP-Hp-SV40e
in the abovementioned tumor cells was similar to the control
plasmid pEGFP-N1, but significantly stronger than that of pEGFP-Hp.
The ratios of transfection efficiency (pEGFP-Hp-SV40e/pEGFP-Hp) in
these tumor cells exceeded 1.65. These findings suggest that the
inserted SV40 enhancer markedly improved the expression of HPSE
promoter-driven downstream genes. The core element of the enhancer
was able to combine with the endonuclear reactivator and indirectly
transmit the signal from activator to transcription factor (TF),
resulting in a marked increase of promoter transcription activity.
Therefore, the enhancer was able to increase transcription
efficiency and become an important component of ‘gene switching’
(22). We presume the inserted SV40
enhancer was capable of specifically combining with the endonuclear
reactivator, thereby enhancing the transcription activity of HPSE
promoter and the expression of GFP.
Findings of the present study showed that the
fluorescence was extremely weak in the pEGFP-Hp-SV40e and pEGFPHp
groups but remained strong in the pEGFP-N1 group following
transfection into the ECV cells. The transfection efficiency of
pEGFP-Hp-SV40e was also low and similar to that of pEGFP-Hp. These
findings suggest that the inserted SV40 enhancer does not affect
the tumor specificity of HPSE promoter, although it is capable of
improving HPSE promoter activity. Enhancer is a type of specific
sequence in gene. Its structure is similar to that of the promoter.
Following activation by the endonuclear reactivator, enhancer must
specifically combine with endonuclear TF in order to improve
promoter activity as well as maintain its specificity. In the
plasmid pEGFP-Hp-SV40e, the function of the inserted SV40 enhancer
depends on the expression of the sequence downstream of the single
HPSE promoter. Therefore, the SV40 enhancer does not affect the
specificity of the HPSE promoter when it enhances its activity,
i.e., the recombinant plasmid pEGFP-Hp-SV40e is expressed only in
tumor cells but not in normal cells. This finding is crucial to
future research and application.
In conclusion, we successfully amplified a complete
SV40 enhancer sequence and constructed the recombinant plasmid
pEGFP-Hp-SV40e by inserting it into pEGFP-Hp. Results of the
fluorescent and flow cytometric analyses found that the
transfection efficiency of recombinant plasmid pEGFP-Hp-SV40e in
tumor cells was significantly higher than the plasmid pEGFP-Hp,
while the transfection efficiency in normal cells remained
unchanged. These results demonstrate that SV40 enhancer can be used
to improve HPSE promoter activity without damaging its tumor
specificity. However, more studies should be carried out to assess
whether or not SV40 enhancer affects the tumor specificity of the
HPSE promoter when it increases transcription activity.
Abbreviations:
|
HPSE
|
heparanase
|
|
SV40
|
simian virus 40
|
|
GCV suicide gene
|
gancyclovir suicide gene
|
|
CK19
|
cytokerain 19
|
|
PSA
|
prostate specific antigen
|
|
GFP
|
green fluorescent protein
|
|
MCS
|
multiple clone site
|
|
TFBS
|
transcription factor binding site
|
|
TF
|
transcription factor
|
Acknowledgements
This study was supported by the
Natural Science Foundation of Anhui, China (grant no. 11040606M181)
and the Provincial Natural Science Foundation of College in Anhui,
China (grant no. kj2010B239). We thank Dr Kong Lv from the Wannan
Medical College (Anhui, China) for technical assistance in flow
cytometry.
References
|
1.
|
Ishiwata N, Inase N, Fujie T, et al:
Suicide gene therapy using keratin 19 enhancer and promoter in
malignant mesothelioma cells. Anticancer Res. 23:1405–1409.
2003.PubMed/NCBI
|
|
2.
|
Chapel-Fernandes S, Jordier F, Lauro F, et
al: Use of the PSA enhancer core element to modulate the expression
of prostate-and non-prostate-specific basal promoters in a
lentiviral vector context. Cancer Gene Ther. 13:919–929. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Li S, MacLaughlin FC, Fewell JG, et al:
Muscle-specific enhancement of gene expression by incorporation of
SV40 enhancer in the expression plasmid. Gene Ther. 8:494–497.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Li HS, Liu Y, Li DF, et al: Enhancement of
DNA vaccine-induced immune responses by a 72-bp element from SV40
enhancer. Chin Med J. 120:496–502. 2007.PubMed/NCBI
|
|
5.
|
Vlodavsky I, Friedmann Y, Elkin M, et al:
Mammalian heparanase: gene cloning; expression and function in
tumor progression and metastasis. Nat Med. 5:793–802. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Hulett MD, Freeman C, Hamdorf BJ, et al:
Cloning of mammalian heparanase; an important enzyme in tumor
invasion and metastasis. Nat Med. 5:803–809. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Toyoshima M and Nakajima M: Human
heparanase. Purification, characterization, cloning and expression.
J Biol Chem. 274:24153–24160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Kussie PH, Hulmes JD, Ludwig DL, et al:
Cloning and functional expression of a human heparanase gene.
Biochem Biophys Res Commun. 261:183–187. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
El-Assal ON, Yamanoi A, Ono T, et al: The
clinicopathological significance of heparanase and basic fibroblast
growth factor expressions in hepatocellular carcinoma. Clin Cancer
Res. 7:1299–1305. 2001.PubMed/NCBI
|
|
10.
|
Chen XP, Liu YB, Rui J, et al: Heparanase
mRNA expression and point mutation in hepatocellular carcinoma.
World J Gastroenterol. 10:2795–2799. 2004.PubMed/NCBI
|
|
11.
|
Komatsu N, Waki M, Sue M, et al:
Heparanase expression in B16 melanoma cells and peripheral blood
neutrophils before and after extravasation detected by novel
anti-mouse heparanase monoclonal antibodies. J Immunol Methods.
331:82–93. 2008. View Article : Google Scholar
|
|
12.
|
Ilan N, Elkin M and Vlodavsky I:
Regulation, function and clinical significance of heparanase in
cancer metastasis and angiogenesis. Int J Biochem Cell Biol.
38:2018–2039. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Chen XP, Hu LH, Cui W, et al: Activity and
tumor specificity of of human heparanase gene core promoter. Mol
Med Rep. (In press).
|
|
14.
|
Boyd DD and Nakajima M: Involvement of
heparanase in tumor metastases: a new target in cancer therapy? J
Natl Cancer Inst. 96:1194–1195. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Yang JM, Wang HJ, Du L, et al: Screening
and identification of novel B cell epitopes in human heparanase and
their anti-invasion property for hepatocellular carcinoma. Cancer
Immunol Immunother. 58:1387–1396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Jiang P, Kumar A, Parrillo JE, et al:
Cloning and characterization of the human heparanase-1 (HPR1) gene
promoter. Role of GA-binding protein and Sp1 in regulating HPR1
BASAL promoter activity. J Biol Chem. 277:8989–8998. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
de Mestre AM, Rao S, Hornby JR, et al:
Early growth response gene 1 (EGR1) regulates heparanase gene
transcription in tumor cells. J Biol Chem. 280:35136–35147.
2005.
|
|
18.
|
Wasylyk B, Wasylyk C, Augereau P, et al:
The SV40 72 bp repeat preferentially potentiates transcription
starting from proximal natural or substitute promoter elements.
Cell. 32:503–514. 1983. View Article : Google Scholar
|
|
19.
|
Lee-Chen GJ and Woodworth-Gutai M: Simian
virus 40 DNA replication: functional organization of regulatory
elements. Mol Cell Biol. 6:3086–3093. 1986.PubMed/NCBI
|
|
20.
|
Song JS: Adenovirus-mediated suicide SCLC
gene therapy using the increased activity of the hTERT promoter by
the MMRE and SV40 enhancer. Biosci Biotechnol Biochem. 69:56–62.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Hagen FK, Zarling DA and Jovin TM:
Electron microscopy of SV40 DNA cross-linked by anti-Z DNA IgG.
EMBO J. 4:837–844. 1985.PubMed/NCBI
|
|
22.
|
Pennisi E: Searching for the genome’s
second code. Science. 306:632–635. 2004.
|