Introduction
DNA-damaging agents, or genotoxic agents, are those
chemicals that can produce alterations in the genetic material of
the host. Such agents can be further subdivided into direct- and
indirect-acting agents. Direct-acting agents are intrinsically
reactive and do not require metabolic activation by cellular
enzymes to interact with DNA. By contrast, indirect-acting agents
require metabolic activation by cellular enzymes to form the
DNA-reactive metabolite. DNA-damaging agents exist widely in our
natural and social environment, with examples including some of the
chemotherapeutic agents and environmental pollutants (1).
Cisplatin, a direct-acting agent, is one of the most
widely used chemotherapeutic agents in the treatment of a wide
variety of malignancies (2).
Although its biochemical mechanism of action has yet to be
elucidated, cisplastin is believed to exert cytotoxic effects
through the interaction and formation of adducts with DNA, which
then leads to apoptosis and necrosis (3,4).
Despite its clinical efficacy in treating malignancies,
cisplatin-based chemotherapy regimens have been reported to be
associated with vascular toxicity and serious vascular
complications (e.g., myocardial infarction and stroke) (5–7). Such
vascular toxicity has been manifested by increased von Willebrand
factor plasma levels as well as an enhanced intima-media thickness
of the carotid artery (8). In
addition, cisplatin has the potential to induce ototoxicity and
toxicity towards renal, peripheral sensory and autonomic nervous
systems, potentially attributed to cisplatin-caused microvascular
damage (9–11).
Ca2+ plays an important role in the
regulation of vascular tone, which is generally relatively
constant. Initiation of contraction in vascular smooth muscle is
due to an increase in the free cytosolic Ca2+
concentration, which is caused by Ca2+ entry via
adenosine triphosphate (ATP)-dependent ion channels (12). Aortic tension also plays an
important role in regulating the entire cardiovascular system.
Large arteries are involved in the regulation of hydrostatic
capillary pressure (13,14) and blood pressure through changing
pulsatile pressure (15). Arteries
have a prominent cushioning effect in addition to their conducting
function and this effect is associated with the viscoelastic
properties of the arterial wall (16). In particular, the vasomotor
reactions have been reported to be related to plasma volume and to
help maintaining cardiovascular homeostasis (13). Although in previous studies it was
reported that cisplatin is associated with a higher incidence of
cardiovascular mortality and morbility (17,18),
the effect of cisplatin on vascular tone remains to be elucidated.
Therefore, the aim of the present study was to evaluate the
cardiovascular toxicity of cisplatin on the contractile function of
rat thoracic aortas and to elucidate the possible underlying
mechanisms.
Materials and methods
Reagents
Phenylephrine (PE), acetylcholine (ACh), cisplatin,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
Fluo-4/AM and ATP were purchased from Sigma (St. Louis, MO, USA).
All other chemicals were of analytical grade and were purchased
from Sinopharm Chemical Reagent (Shanghai, China). Cisplatin was
dissolved in phosphate-buffered saline (PBS) for in vitro
and in vivo experiments. Current guidelines for use of
biohazardous materials in Zhejiang University were followed when
using cisplatin.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
grown in RPMI-1640 medium supplemented with 10% (v/v) fetal bovine
serum (Gibco, Carlsbad, CA, USA) at 37°C under humidified air (5%
CO2).
Cytotoxicity assay
The toxicity of cisplatin was examined using the MTT
assay. Briefly, cells were seeded into a 96-well culture plate at
density of 1×104 cells/well. After 24 h, the medium was
discarded and replaced with fresh medium containing the indicated
concentrations of chemicals. The cells were treated for 6 or 12 h
and by the end of each time point, 20 μl of MTT (5 mg/ml in PBS)
was added to each well and further incubated for 3 h. The solution
was then discarded and 150 μl of isopropanol was added. Following
agitation for 10 min to dissolve the formazan, the absorbance at
570 nm was read on a microtiter plate reader (BioTek, Winooski, VT,
USA). Relative survival was represented as the absorbance of the
treated sample/absorbance of the control group.
Animals and treatment of cisplatin
Male Sprague-Dawley (SD) rats weighing 230–260 g
were obtained from the Laboratory Animal Center of Zhejiang
University (Hangzhou, China). All the procedures were approved by
the Ethics Committee for the Use of Experimental Animals in
Zhejiang University. Rats were housed in an air-conditioned colony
room at 20±2°C and diet and water were provided ad libitum.
To detect the effect of cisplatin on thoracic aortas, the rats were
injected intraperitoneally with cisplatin at a dose of 5 mg/kg body
weight and an equivalent volume of saline was used as control
(11).
Preparation of thoracic aortic rings
Rats were anaesthetized with 10% chloral hydrate
(0.4 g/kg, intraperitoneally) and sacrificed by cervical
dislocation. The chest was opened and the thoracic aorta was
rapidly removed and placed in a 4°C Krebs-Henseleit (K-H) solution
[mM: NaCl, 120.0; KCl, 4.5; CaCl2, 1.25;
KH2PO4, 1.2; MgSO4, 1.2;
NaHCO3, 25.0; and glucose, 10.0 (pH 7.4)]. Following
removal of the superficial connective tissue, the aorta was cut
into rings of 3–4 mm in length and subjected to contractile
functional studies and electron microscopic measurements.
Electron microscopy
Three aortic rings cut from different segments of
the thoracic aorta (superior, middle and inferior segment) were
fixed with 2.5% glutaraldehyde at 4°C for >24 h and then washed
twice in 0.1 M PBS at 4°C for 15 min. The tissue was incubated in
1% osmic acid at 4°C for 1 h and then rinsed twice in the same
buffer for 15 min. After staining in 2% uranyl acetate (en bloc) at
4°C for 30 min, the tissues were processed by graded dehydration
with different concentrations of ethanol (50, 70, 90 and 100%) and
100% acetone, respectively, each for 15 min. The dehydrated
thoracic aortic rings were then agitated with a mixture of
embedding medium (Epon 812) to acetone (1:1) for 1 h and then
transferred into pure embedding medium (Epon 812) for another 1.5–2
h. Subsequently, the aortas were embedded with the same medium at
37, 45 and 60°C, each for 24, 24 and 48 h, respectively. Sections
of ~120 nm were taken from the prepared rings using an
ultramicrotome (Leica Microsystems, Milton Keynes, UK). The
sections were stained with 4% uranyl acetate for 20 min and lead
citrate for 5 min and then observed under transmission electron
microscopy (Tecnai 10, 80 kV). The number of smooth muscle cells
was counted depending on the image (x970) captured from electron
microscopy.
Measurement of contractile function of
thoracic aortic rings
Fresh isolated rings were mounted in 5.0 ml organ
baths containing the K-H solution. The bath solution was maintained
at 37°C and bubbled continuously with a gas mixture of 95%
O2 and 5% CO2. Aortic rings were equilibrated
for 60 min at resting tensions of 2 g. Isometric tension was
measured with force isometric transducers connected to a data
acquisition system (MedLab; Nanjing Medease Co., Ltd., Nanjing,
China). In some aortic rings, the endothelium was mechanically
removed by gentle rubbing with moistened cotton. The removal of
endothelial cells was confirmed by the loss of ACh-induced
relaxation (19–21). After 6 h pre-incubation with
cisplatin (200 μM), contraction of the endothelium-intact and
endothelium-denuded aortic rings was induced by KCl (60 mM) or PE
(10−6 M). The vascular response was expressed as a
percentage of the ‘plateau’ contraction evoked by KCl or PE
(22).
Measurement of intracellular
Ca2+ concentration ([Ca2+]i)
HUVECs grown on coverslips were treated with
cisplatin (200 μM) for 6 h. The cells were loaded with the
Ca2+ sensitive dye Fluo-4/AM (1 nM) for 30 min at 37°C
in serum-free RPMI-1640 medium. After loading, the cells were
washed three times with serum-free RPMI-1640 medium. The loaded
cells were maintained at 37°C for 15 min before measurement of
[Ca2+]i in order to allow the Fluo-4/AM in
the cytosol to deesterify. The cells were subsequently placed on
the stage of a Nikon inverted microscope equipped with a SFX-1
microfluorimeter (Solamere Technology Group, Salt Lake City, UT,
USA). During the experiments, the cells were superfused in a 0.3-ml
bath chamber at 35°C under a constant flow (1 ml/min) of
HEPES-buffered saline (135 mM NaCl, 5 mM KCl, 1.5 mM
CaCl2, 1 mM MgCl2, 10 mM glucose and 10 mM
HEPES; pH 7.4) (23). To examine
the action of cisplatin on ATP-induced increased
[Ca2+]i, the cells were stimulated with ATP
(10 μM) for 1 min. ATP was then washed away twice by HEPES-buffered
saline. Fluo-4 fluorescence was excited at 470 nm. Emitted Fluo-4
fluorescence was measured at wavelengths of 520 nm. Changes in
[Ca2+]i were reported as the Fluo-4/AM ratio
ΔF/ΔF0, where ΔF is the difference between baseline
fluorescence intensity and maximal fluorescence intensity after ATP
stimulation, while ΔF0 is the difference between
baseline fluorescence intensity and the minimal fluorescence
intensity prior to ATP stimulation.
Statistical analysis
Data were expressed as mean ± standard error of the
mean (SEM). Statistical analysis was carried out using Student’s
paired t-test or one-way analysis of variance followed by Dunnett’s
t test. Differences were considered statistically significant at
P<0.05.
Results
Effects of cisplatin on the contractile
function of thoracic aortas of SD rats
First the cytotoxicity of cisplatin was examined by
MTT using HUVECs to determine the appropriate dose for subsequent
experimentation. As shown in Fig.
1, cisplatin at doses ranging 5–500 μM induced cytotoxicity at
6 and 12 h in a dose-dependent manner, with maximal decreases in
cell survival (20–30%) observed at the higher concentrations.
Using the ex vivo perfusion system, the
effects of cisplatin on the contraction of aortic rings were
examined. At concentrations ranging 10–500 μM, cisplatin itself had
no direct effect on the baseline tension of endothelium-intact
aortic rings (Table I). Combined
with the cytotoxicity data, 200 μM was selected as the
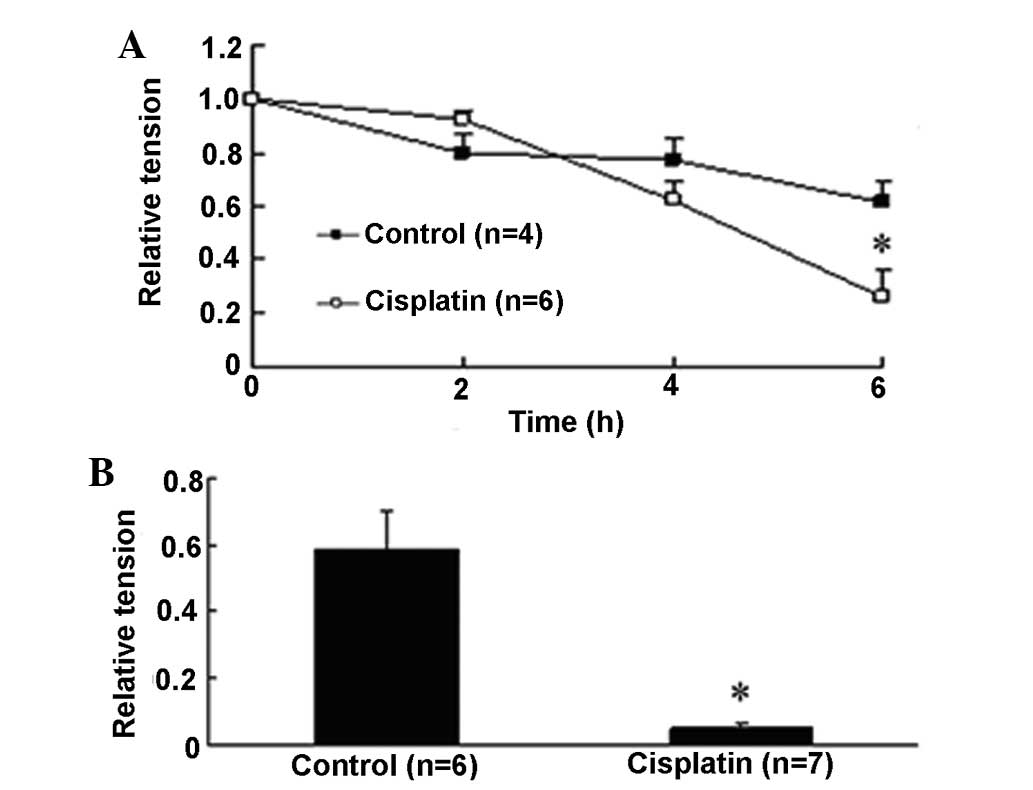
concentration for subsequent experimentation. KCl (60 mM) induced
significant contraction with the relative tension as 1 at time 0
and gradually decreased to 0.61±0.08 after 6 h. It was found that
cisplatin decreased the contraction tension provoked by KCl in a
time-dependent manner, with a maximum inhibition of 57.6%
(0.26±0.10, P<0.05 versus 0.61±0.08) at 6 h (Fig. 2A). PE (10−6 M) increased
the contraction tension to a similar level as KCl (0.59±0.12) at 6
h and it was almost abolished by cisplatin, as shown in Fig. 2B, in which cisplatin reduced
PE-induced contraction tension by 91.8% (0.049±0.01, P<0.05
versus 0.59±0.12).
 | Table IThe relative baseline tension of
aortic rings treated with cisplatin. |
Table I
The relative baseline tension of
aortic rings treated with cisplatin.
| Thoracic aortic
rings | Control (n) | Cisplatin (n) |
|---|
|
|---|
| 10 μM | 100 μM | 200 μM | 500 μM |
|---|
| +E | 0.018±0.036
(6) | 0.033±0.045
(5) | 0.073±0.054
(6) | 0.091±0.063
(6) | 0.059±0.034
(5) |
| −E | 0.033±0.052
(9) | 0.037±0.050
(5) | 0.056±0.058
(6) | 0.021±0.045
(7) | 0.063±0.069
(5) |
Effects of cisplatin on the contractile
function of endothelium-denuded thoracic aortas of SD rats
Endothelium is known to play an important role in
regulating the vascular tone by releasing factors involved in
relaxation and contraction (24).
Therefore, we also assessed the effects of cisplatin on
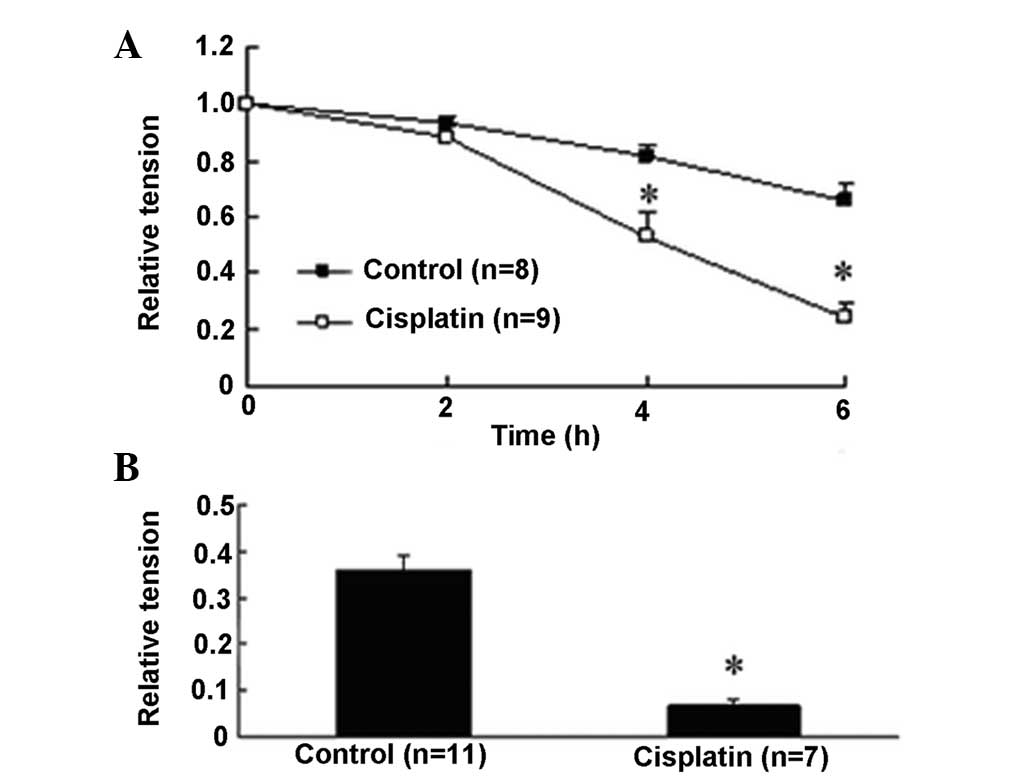
endothelium-denuded aortic rings. In the aortic rings of
endothelium denudation, KCl and PE also increased vascular tension
to 0.65±0.07 and 0.36±0.03 at 6 h, respectively (Fig. 3). Similar to the endothelium-intact
situation, cisplatin decreased KCl-induced contraction tension by
63.0% (0.24±0.05, P<0.05 versus 0.65±0.07) and PE-induced
contraction tension by 81.3% (0.067±0.01, P<0.05 versus
0.36±0.03).
Effect of cisplatin on
[Ca2+]i in HUVECs
As stated before, since
[Ca2+]i is also an important factor in
regulating vascular tone, the effect of cisplatin on
[Ca2+]i was examined in HUVECs. As shown in
Fig. 4, cisplatin (200 μM)
significantly decreased the [Ca2+]i in HUVECs
increased by ATP (2.79±0.58 versus 4.42±0.33).
Effects of cisplatin on aortic vessel
wall components in vivo
To examine the effects of cisplatin on the aortic
walls, aortas from the control- and cisplatin-treated rats were
isolated and processed for electron microscopic observation. As
shown in Fig. 5, cisplatin caused
severe damage to the smooth muscle layer. Compared to the control,
the boundary of smooth muscle layers was less clearly defined and
almost touching each other in the aortas from cispatin-treated
rats. Cisplatin also caused significant loss of smooth muscle
cells, for example, the average smooth muscle cell count was
decreased by 34.7% (from 26,383±2,561 to 17,217±1,452
mm−2 with cisplatin treatment, P<0.05).
Discussion
In the present study, by measuring the contractile
function of thoracic aortas in an ex vivo perfusion system,
we evaluated the vascular effects of the DNA-damaging agent
cisplatin. It has been reported that there is a high incidence of
cardiovascular events in survivors of cisplatin-based chemotherapy
cancer patients (25), either in
acute or long-term chemotherapy (26,27).
Cisplatin was reported to lead to aortic occlusion in some cases
and this impairment was usually irreversible (28,29).
In one case, a 36-year-old man who was treated with
bleomycin-etoposide-cisplatin therapy for testicular cancer
received chronic coronary artery dissection (30). A long-term follow-up in a large
cohort of testicular cancer survivors showed that compared with
healthy controls, cisplatin-treated patients had, at follow-up,
increased systolic blood pressure, increased diastolic blood
pressure and a higher prevalence of hypertension (18). In that cohort, patients in the
high-dose cisplatin-treated group had increased odds for having
intermediate/high risk in cardiovascular mortality, compared with
the surgery group (OR=3.4, 95% CI: 1.3–8.7) (17). All of the above case reports led to
the hypothesis that cisplatin may cause deleterious effects on the
cardiovascular system. Thus, we undertook a first step to
investigate whether cisplatin has such an effect and in particular,
we focused on the contractile function of aortic rings as an
indicator.
First, we demonstrated that the baseline tensions of
thoracic aortas were not altered by cisplatin treatment, which
means that cisplatin had no direct deleterious effect on the
contractile function of aortas. However, cisplatin inhibited KCl-
or PE-induced contraction in the endothelium-intact and
endothelium-denuded aortas, suggesting an endothelium-independent
mechanism (Figs. 2 and 3). To investigate the reason for cisplatin
decreasing the tension of aortic rings, we examined the structure
of the aortic walls. As shown in Fig.
5, cisplatin severely damaged the smooth muscle layer, and
significantly decreased smooth muscle cells in the thoracic aortic
wall. The fundamental structural and functional unit of the aortic
wall is the medial lamellar unit. The media layer comprises elastic
membrane layers between which are the smooth muscle layer and a
small amount of collagen and elastic fibers. The integrity of the
vascular smooth muscle layer is crucial in the maintenance of
normal vascular morphology and tone. Changes in arterial wall
composition and function underlie all forms of vascular disease
(31). Therefore, the finding that
cisplatin damaged the arterial wall may partially explain its
inhibitory effect on the contraction of aortic rings.
In addition, we have also shown that cisplatin
inhibited ATP-induced Ca2+ influx in HUVECs (Fig. 4). However, whether cisplatin
similarly affects Ca2+ in aortic tissues remains to be
determined.
The effects of another envionmentally prevalent DNA
damaging agent, benzo(a)pyrene [B(a)P] on the contractile function
of thoracic aortas have also been investigated. Unlike cisplatin,
B(a)P is an indrect-acting agent and has to be metabolically
processed by aryl hydrocarbon receptor-induced enzymes, such as
cytochrome P4501A1 (CYP1A1) and epoxide hydrolase, to form
carcinogenic active form B(a)P-diol epoxides (32). Of note, B(a)P only affected the
contractile function of thoracic aortas following long-term in
vivo exposure (4 weeks) (data not shown).
In conclusion, in the present study, we have shown
that the DNA-damaging agent cisplatin: i) decreased the contractile
function of thoracic aortas; and ii) caused direct vessel well
damage and cytotoxicity towards smooth muscle cells. Therefore, it
seems that DNA-damaging agents may also have cardiovascular
toxicity and thus, the potential cardiovascular complications
associated with DNA-damaging agents should be investigated in
future studies.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81373036, 81172692
and 81202241) and Zhejiang Provincial of Science and Technology
Department (grant no. 2013C14016). J. Yang is a recipient of the
Zhejiang Provincial Program for the Cultivation of High-Level
Innovative Health Talents.
References
|
1
|
Williams GM and Weisburger JH: Chemical
carcinogenesis. Casarett and Doull’s Toxicology: The Basic Science
of Poisons. Amdur MO, Doull J and Klaassen CD: 4th edition.
Pergamon Press; New York, NY: pp. 127–132. 1991
|
|
2
|
Abu-Surrah AS and Kettunen M: Platinum
group antitumor chemistry: design and development of new anticancer
drugs complementary to cisplatin. Curr Med Chem. 13:1337–1357.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fuertes MA, Castilla J, Alonso C and Perez
JM: Cisplatin biochemical mechanism of action: from cytotoxicity to
induction of cell death through interconnections between apoptotic
and necrotic pathways. Curr Med Chem. 10:257–266. 2003. View Article : Google Scholar
|
|
4
|
Shinohara K, Bando T, Sasaki S, Sakakibara
Y, Minoshima M and Sugiyama H: Antitumor activity of
sequence-specific alkylating agents: pyrolle-imidazole CBI
conjugates with indole linker. Cancer Sci. 97:219–225. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Doll DC, List AF, Greco FA, Hainsworth JD,
Hande KR and Johnson DH: Acute vascular ischemic events after
cisplatin-based combination chemotherapy for germ-cell tumors of
the testis. Ann Intern Med. 105:48–51. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huddart RA, Norman A, Shahidi M, et al:
Cardiovascular disease as a long-term complication of treatment for
testicular cancer. J Clin Oncol. 21:1513–1523. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vaughn DJ, Palmer SC, Carver JR, Jacobs LA
and Mohler ER: Cardiovascular risk in long-term survivors of
testicular cancer. Cancer. 112:1949–1953. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nuver J, Smit AJ, van der Meer J, et al:
Acute chemotherapy-induced cardiovascular changes in patients with
testicular cancer. J Clin Oncol. 23:9130–9137. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kirchmair R, Walter DH, Ii M, et al:
Antiangiogenesis mediates cisplatin-induced peripheral neuropathy:
attenuation or reversal by local vascular endothelial growth factor
gene therapy without augmenting tumor growth. Circulation.
111:2662–2670. 2005. View Article : Google Scholar
|
|
10
|
Kohn S, Fradis M, Ben-David J, Zidan J and
Robinson E: Nephrotoxicity of combined treatment with cisplatin and
gentamicin in the guinea pig: glomerular injury findings.
Ultrastruct Pathol. 26:371–382. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu M, Han J, Cui P, et al: Cisplatin
up-regulates ICAM-1 expression in endothelial cell via a NF-kappaB
dependent pathway. Cancer Sci. 99:391–397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aromolaran AS, Zima AV and Blatter LA:
Role of glycolytically generated ATP for CaMKII-mediated regulation
of intracellular Ca2+ signaling in bovine vascular endothelial
cells. Am J Physiol Cell Physiol. 293:C106–C118. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mellander S: Contribution of small vessel
tone to the regulation of blood volume and formation of oedema.
Proc R Soc Med. 61:55–61. 1968.PubMed/NCBI
|
|
14
|
Suga H: Physiological interpretation of
negative circumferential tension in vascular walls. Jpn Heart J.
32:473–480. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nichols WV and O’Rourke MF: McDonald’s
Blood Flow in Arteries: Theoretical, Experimental, and Clinical
Principles. 5th edition. Hodder Arnold; London: pp. 344–347.
2005
|
|
16
|
Struijker Boudier HAJ: Vascular growth and
hypertension. Textbook of Hypertension. Swales JD: Blackwell
Scientific; Oxford: pp. 200–213. 1994
|
|
17
|
Haugnes HS, Aass N, Fossa SD, et al:
Predicted cardiovascular mortality and reported cardiovascular
morbidity in testicular cancer survivors. J Cancer Surviv.
2:128–137. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sagstuen H, Aass N, Fossa SD, et al: Blood
pressure and body mass index in long-term survivors of testicular
cancer. J Clin Oncol. 23:4980–4990. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gong Z, Yang J, Yang M, et al:
Benzo(a)pyrene inhibits expression of inducible heat shock
protein 70 in vascular endothelial cells. Toxicol Lett.
166:229–236. 2006.
|
|
20
|
Kamata K and Kojima S: Characteristics of
contractile responses of aorta to norepinephrine in db/db mice. Res
Commun Mol Pathol Pharmacol. 96:319–328. 1997.PubMed/NCBI
|
|
21
|
Qian LB, Wang HP, Qiu WL, Huang H, Bruce
IC and Xia Q: Interleukin-2 protects against endothelial
dysfunction induced by high glucose levels in rats. Vascul
Pharmacol. 45:374–382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ru XC, Qian LB, Gao Q, Li YF, Bruce IC and
Xia Q: Alcohol induces relaxation of rat thoracic aorta and
mesenteric arterial bed. Alcohol Alcohol. 43:537–543. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sheng JZ, Wang D and Braun AP: DAF-FM
(4-amino-5-methylamino-2′,7′-difluorofluorescein) diacetate detects
impairment of agonist-stimulated nitric oxide synthesis by elevated
glucose in human vascular endothelial cells: reversal by vitamin C
and L-sepiapterin. J Pharmacol Exp Ther. 315:931–940. 2005.
|
|
24
|
Luscher TF and Tanner FC: Endothelial
regulation of vascular tone and growth. Am J Hypertens.
6:283S–293S. 1993.PubMed/NCBI
|
|
25
|
Ishioka J, Fujii Y, Kageyama Y, Fukuda H,
Higashi Y and Kihara K: Cardiovascular events in survivors of
high-dose chemotherapy for germ cell tumors. Int J Urol.
15:642–645. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fukunaga T, Soejima H, Sugamura K, et al:
Acute myocardial infarction induced by cisplatin-based combination
chemotherapy for malignant melanoma: a case report. J Cardiol.
47:191–195. 2006.PubMed/NCBI
|
|
27
|
Oh JH, Baum DD, Pham S, et al: Long-term
complications of platinum-based chemotherapy in testicular cancer
survivors. Med Oncol. 24:175–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Etgen T, Weidenhofer G and Kubin T:
Cisplatin-associated occlusion of the internal carotid artery.
Onkologie. 32:754–757. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Grenader T, Shavit L, Ospovat I, Gutfeld O
and Peretz T: Aortic occlusion in patients treated with
cisplatin-based chemotherapy. Mt Sinai J Med. 73:810–812.
2006.PubMed/NCBI
|
|
30
|
Ghosh N, Chow CM, Korley V and Chisholm R:
An unusual case of chronic coronary artery dissection: did
cisplatin play a role? Can J Cardiol. 24:795–797. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dabagh M, Jalali P, Konttinen YT and
Sarkomaa P: Distribution of shear stress over smooth muscle cells
in deformable arterial wall. Med Biol Eng Comput. 46:649–657. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Oesterling E, Toborek M and Hennig B:
Benzo[a]pyrene induces intercellular adhesion molecule-1 through a
caveolae and aryl hydrocarbon receptor mediated pathway. Toxicol
Appl Pharmacol. 232:309–316. 2008.
|