Introduction
Diabetes mellitus is a disease affecting ~5% of
people worldwide. Type I diabetes is mainly due to autoimmune
damage of pancreatic β-cells and patients are dependent on
exogenous insulin, and this accounts for 5–10% of all the diabetic
cases. Type II diabetes develops more slowly and the intensity of
metabolic disturbances is much lower and accounts for 90–95% of all
diabetic cases (1,2). In type II diabetes, the reduction in
β-cell mass and insulin secretion was observed (3–5).
Metformin is the first choice for the treatment of
type II diabetes. In type II diabetes, metformin lowers blood
glucose concentrations without causing overt hypoglycemia, and
therefore is an antihyperglycemic agent. Metformin is also
described as an insulin-sensitizer causing reduction in insulin
resistance and the plasma fasting insulin level. The positive
effects of metformin on insulin receptor expression and tyrosine
kinase activity could be improving the insulin sensitivity
(6,7).
However, the primary function of metformin is to decrease hepatic
glucose production (8), mainly by
inhibiting gluconeogenesis (9,10).
Studies on the rat pancreas (11) and isolated human islets (12) have shown that metformin improved
insulin release in response to glucose. A previous study has
demonstrated that metformin can restore the normal secretory
pattern in isolated rat islets that were incubated in the presence
of elevated glucose or free fatty acid concentrations (13).
In the adult pancreas, pancreatic and duodenal
homeobox 1 (Pdx-1) regulates the genes associated with pancreatic
cell differentiation and maturation, which includes the glucose
transporter 2, insulin, glucokinase and amyloid precursor protein
genes (14). The decision to test
metformin was based on these considerations and the results showed
that the drug can reverse the majority of the alterations found in
type II diabetes islets.
The aim of the present study was to investigate the
direct effects of metformin on pancreatic β-cells by incubation of
pancreatic islets cells undergoing glucose stress to assess the
expression of Pdx-1 and insulin genes, β-cell function and
viability.
Materials and methods
Animals
A total of 56 male NMRI mice (25–30 g) (9 mice in
each group) were obtained from the Animal House of Ahvaz
Jundishapur University of Medical Sciences (Ahvaz, Iran), The mice
were housed in cages (22±2°C, under a standard 12-h light:12-h dark
cycle) and allowed ad libitum feed access. All the
experimental protocols were performed according to the Standards
for Animal Care, and they were approved by the Ethics Committee of
Ahvaz Jundishapur University of Medical Sciences.
Isolation of mice pancreatic
islets
Pancreatic islets were isolated from
overnight-fasted male NMRI mice by the Lacy and
Kostianovsky-modified collagenase digestion method (13). In brief, following cervical
dislocation, the abdomen of the animals was opened. The common bile
duct was occluded at the distal end, close to the duodenum. Hanks'
balanced salt solution (HBSS; 5 ml) [115 mmol/l NaCl, 10 mmol/l
NaHCO3, 5 mmol/l KCl, 1.1 mmol/l MgCl2, 1.2
mmol/l NaH2PO4, 2.5 mmol/l CaCl2,
25 mmol/l HEPES and 5 mmol/l D-glucose (pH 7.4), as well as 1%
bovine serum albumin (Merck KGaA, Darmstadt, Germany)] containing
1.4 mg/ml of collagenase IV (Sigma, St. Louis, MO, USA) was
injected into the duct (14,15).
After removal of the pancreas, it was placed into a
50 ml conical tube and incubated for 15 min in a 37°C water bath.
Subsequently, 15 ml of cold HBSS was added to the tube to dilute
the collagenase concentration and stop a further digestion process.
For washing the collagenase from the islet tissues, the tube was
centrifuged for 2 min at 1,200 rpm and the supernatant has
discarded. The washing procedure of islets was repeated again and
the remainding sample was transferred to a petri dish (16). The islets were separated by handpicking
under a stereomicroscope (Euromex Microscopen BV, Arnhem, The
Netherlands) and were cultured overnight in RPMI-1640 medium
(Gibco, Invitrogen Life Technologies Carlsbad, CA, USA),
supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100
U/ml streptomycin, 5 mmol/l D-glucose and was gassed with 95%
O2-5% CO2 atmosphere. Subsequently, the
pancreatic islets were divided into 6 groups, including 3 control
and 3 treatment groups: Control group 1: Incubated for 24 h in
RPMI-1640 containing 5 mmol/l glucose (Sigma); control group 2:
Incubated for 48 h in RPMI-1640 containing 5 mmol/l glucose; and
control group 3: Incubated for 24 h in RPMI-1640 containing 25
mmol/l glucose; and treatment group 1: Incubated for 24 h in
RPMI-1640 containing 5 mmol/l glucose and 15 µmol/l metformin;
treatment group 2: Incubated for 48 h in RPMI-1640 containing 5
mmol/l glucose and 15 µmol/l metformin; and treatment group 3:
Incubated for 24 h in RPMI-1640 containing 25 mmol/l glucose and 15
µmol/l metformin.
Insulin secretion measurement
Islet function was determined by monitoring insulin
release following a static incubation glucose challenge assay.
Function was determined by exposing the islets to glucose and
metformin for different concentrations of glucose and for different
periods, and determining how much insulin was secreted in
response.
After in vitro treatment, 5 sets of 4 islets
were incubated in HBSS containing 5 or 25 mmol/l glucose at 37°C
for 1 h. At the end of the incubation period, the supernatant was
collected and frozen at −20°C for later insulin radioimmunoassay
(Bio-Rad, Hercules, CA, USA) using a mouse insulin standard
(17).
Quantitative polymerase chain reaction
(qPCR)
Total RNA was extracted from mouse pancreatic islets
using the RNeasy plus mini kit (Qiagen, Hilden, Germany). RNA
concentration was measured using a NanoDrop (Thermo Fisher
Scientific, Inc., Wilmington, DE, USA). RNA (200 ng) was used to
generate cDNA using the RevertAid First Strand cDNA Synthesis kit
(Fermentas, Thermo Fisher Scientific) and amplified by qPCR using a
QuantiTect SYBR-Green kit (Qiagen), the ABI StepOnePlus qPCR
instrument and software (Applied Biosystems, Foster City, CA, USA).
Primer sequences used are provided in Table I. All the quantifications were
performed with mouse β-actin as an internal standard. The PCR was
performed for 40 cycles at 95°C for 15 sec, 60°C for 30 sec and
72°C for 30 sec (16,17).
 | Table I.Sequence information on the primers
used for qPCR. |
Table I.
Sequence information on the primers
used for qPCR.
| Gene | Sequences
(5′→3′) | Product size, bp | Gene ID |
|---|
| Pdx-1 | F:
CCGAGAGACACATCAAAATCTGG | 80 | NM_008814.3 |
|
| R:
CCCGCTACTACGTTTCTTATCTTCC |
|
|
| Insulin | F:
AGGACCCACAAGTGGCACA | 184 | NM_001185084.1 |
|
| R:
GAGGGGTAGGCTGGGTAGTG |
|
|
| Glut-2 | F:
TTGACTGGAGCCCTCTTGATG | 73 | NM_031197.2 |
|
| R:
CACTTCGTCCAGCAATGATGA |
|
|
| β-actin | F:
GGCCAACCGTGAAAAGATGA | 79 | NM_007393.3 |
|
| R:
CACAGCCTGGATGGCTACGT |
|
|
Islet viability
Islet cell viability was tested by a colorimetric
assay that detected the conversion of MTT (Sigma) into the formazan
by the mitochondrial enzyme succinate dehydrogenase in viable cells
(18,19). After in vitro treatment, islets
dissociated into single cells by incubation in HBSS containing 0.25
mg/ml trypsin for 10 min at 37°C with gentle agitation and were
resuspended in RPMI-1640. Islet cells were cultured in a 96-well
plate supplemented with 0.5 mg/ml MTT. After 4 h incubation, the
insoluble formazan crystals within islet cells were extracted by
dimethyl sulfoxide and absorbance was measured by a microplate
reader (Bio-Rad) at a wavelength of λtest = 570 nm and
λreference = 650 nm.
Statistical analysis
Data were expressed by SPSS software (SPSS, Inc.,
Chicago, IL, USA) as mean ± standard error of the mean. One-way
analysis of variance was used for comparison of the data from
different groups, followed by the Tukey's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
In vitro islet function
The rate of insulin secretion (ng/ml/60 min/islet)
from isolated islets in 5 mmol/l glucose in the 48 h treatment
group (glucose plus metformin) increased significantly compared
with that of the 24 h treatment group (P<0.05). The differences
between the control groups compared with the treatment groups were
not significant (Fig. 1).
To determine the effect of different concentrations
of glucose on insulin secretion in the glucose alone and glucose
plus metformin groups, insulin secretion was examined at different
extracellular glucose concentrations. Treated mice islets were
incubated for 60 min in the presence of 5 or 25 mmol/l glucose, and
the results demonstrated that in the glucose compared with the
glucose and metformin group, insulin secretion increased but this
difference was not significant. Data demonstrated that insulin
secretion increased with the increase of glucose concentration
(Fig. 2).
qPCR
The Pdx-1 and insulin gene expression was
analyzed by qPCR to demonstrate whether metformin has a significant
effect on β-cell function during different time exposures to
glucose or glucose plus metformin.
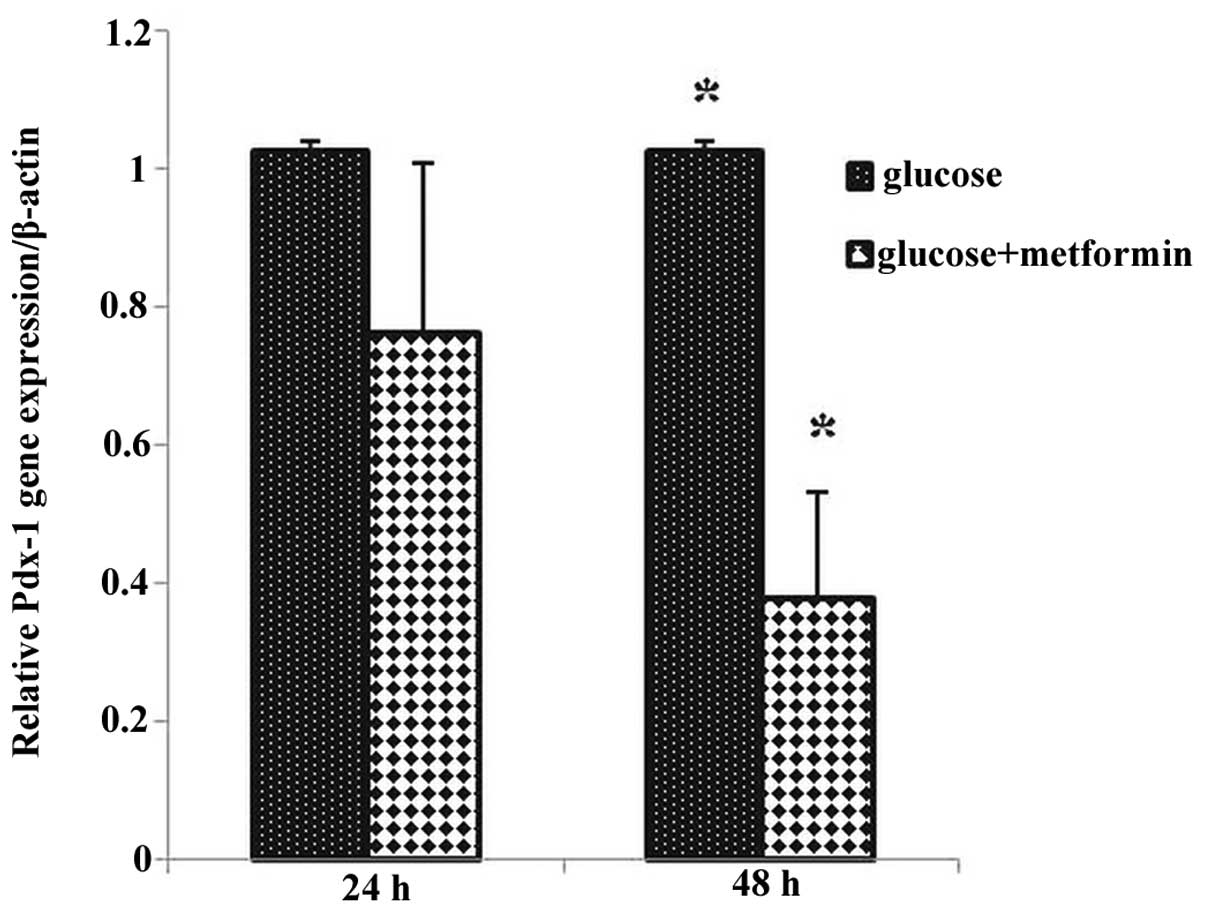
The Pdx-1 gene expression in the glucose plus
metformin group 1 decreased compared with that of the control group
1, and Pdx-1 gene expression in glucose plus metformin group
2 decreased significantly compared with the control group 2
(P<0.05), as well as during different time exposures to glucose
plus metformin group 2 (48 h) compared with the glucose plus
metformin group 1 (24 h) (Fig. 3).
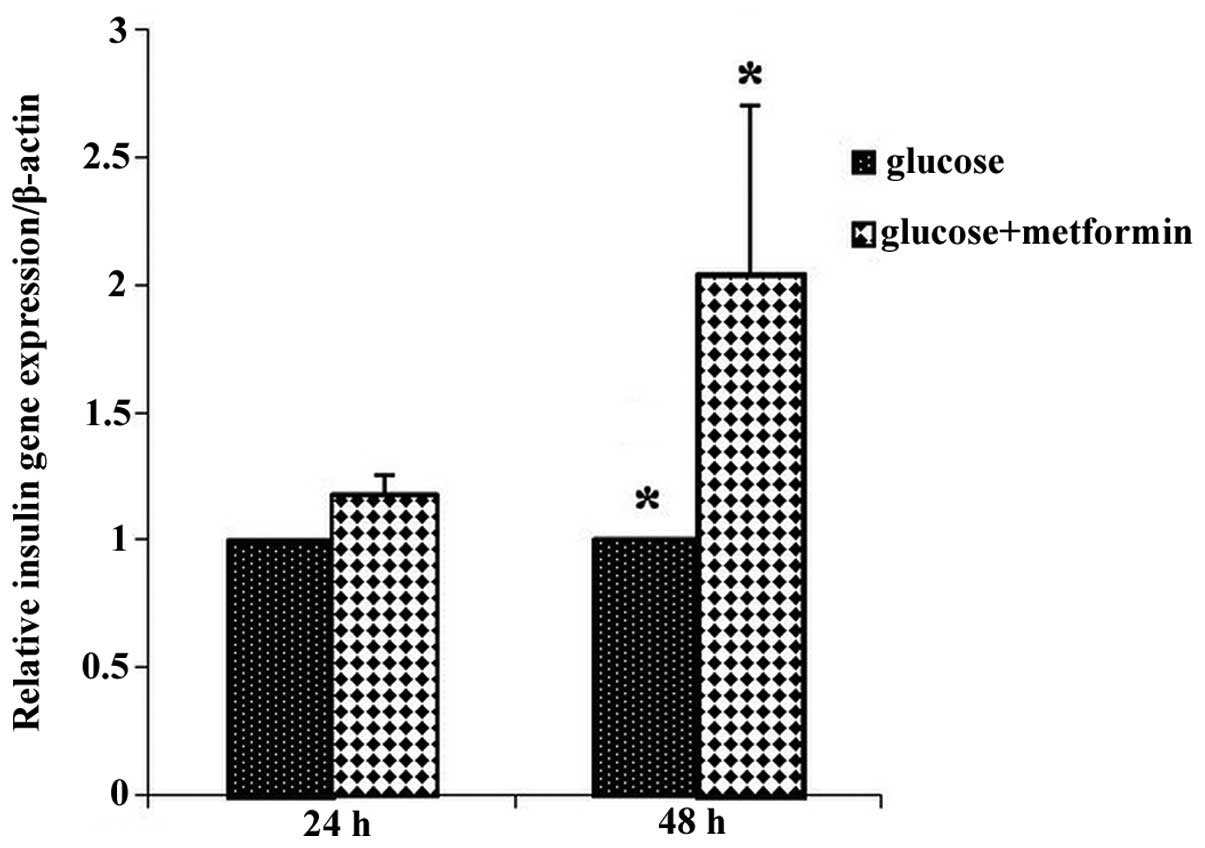
To investigate the effect of metformin on insulin
gene expression during different time exposures of metformin and
glucose, the same experiment was performed. The expression of the
insulin gene in treatment group 1 increased compared with that of
control group 1, and in glucose plus metformin group 2 there was a
2-fold increase in insulin gene expression compared with that of
control group 2. The expression of the insulin gene in the glucose
plus metformin group 2 increased compared with that of glucose plus
metformin group 1 (Fig. 4).
Islet viability
Incubations of pancreatic islet cells with MTT
demonstrated that reduction of this compound to formazan differed
in islet cells of different treatment groups. Formazan formation
was increased in treatment group 3 (25 mmol/l glucose and
metformin) by ~40% compared with the islets of treatment groups 1
and 2 (P<0.05) (Fig. 5).
Discussion
In the present study, metformin and glucose had
evident effects on β-cell genes expression, islet function and
viability in mouse pancreas.
Previous studies indicated that in type II diabetes
islets secrete less insulin in response to glucose compared wtih
that of the control group. This deficiency is characterized by loss
of first-phase insulin secretion (20,21), and the
present study showed that insulin release in the metformin plus
glucose group increased in a time-dependent manner. The data
demonstrated that insulin secretion increased with the increase of
glucose concentration, consistent with the study by Marchetti et
al (22), which showed that
metformin incubation restored insulin release. This effect was
associated with replenishment of islet insulin storage (as
indicated by the total insulin content and mature insulin
granules). Leclerc et al (23)
demonstrated that in the control group, increases in glucose
concentration activate insulin release, which is consistent with
the present study. However, we demonstrated that in the metformin
plus glucose group the increases in glucose concentration activate
insulin release, but Leclerc et al (23) showed that increases in glucose
concentration in the metformin-treated group inhibits insulin
secretion.
The increase in insulin release could be due to a
more efficient insulin synthesis. The present study demonstrated
that exposure of the islets in the metformin and glucose group
caused a 2-fold increase in insulin mRNA in a time-dependent
manner. A previous study by Marchetti et al (22) indicated that in type II diabetes islets
insulin mRNA was markedly lower than that of the control islets and
increased significantly following exposure to metformin (24). The present findings showed a
significant decrease in Pdx-1 mRNA during time exposure to
metformin plus glucose. This data is consistent with a previous
study by Richardson et al (25), which showed that metformin can
stimulate Pdx-1 post-transcriptional modification, leading to
immobilization of the Pdx-1 protein into the nucleus >24 h and
there was no Pdx-1 upregulation in the level of Pdx-1
promoter activity in the electrophoresis mobility shift assay. They
demonstrated that metformin stimulates Pdx-1 at the protein
production levels by western blot analysis and not at the
Pdx-1 gene expression level. Therefore, a significant
decrease in mRNA of Pdx-1 in the metformin plus glucose
group in a time-dependent manner may be due to the increase in
translation of Pdx-1 mRNA, and therefore, the increase in
the Pdx-1 transcription factor causes transcription of the
insulin gene, and therefore the insulin mRNA level increases.
In type II diabetes, a deficiency of β-cell survival
exists. Previously, Butler et al (4) examined autoptic pancreatic tissue,
reported a several-fold increase in the frequency of β-cell
apoptosis. Our present study showed that metformin plus 25 mmol/l
glucose caused an ~40% increase in islet cell viability. This is
consistent with the study by Marchetti et al (22), which found that metformin had an
antiapoptotic effect, paralleled by a reduction of caspase-3 and −8
activities.
By contrast, Kefas et al (26) showed that metformin impairs glucose
responsivity and enhances susceptibility to apoptosis.
The differences between the results may be due to
variation in culture processes and experimental models, the dose of
metformin, as differences are species specific, and the duration of
metformin and glucose presence.
In conclusion, the present study indicates that
metformin has a profound effect on insulin release, transcriptional
regulation in pancreatic islets and islet cell viability, and these
effects are dependent on the presence of glucose. These data
indicate that metformin has direct effects on islet function and
suggests that this widely prescribed antidiabetic drug may play a
previously unrecognized role in the direct regulation of pancreatic
β-cell function. Further studies are required to fully delineate
the cell signaling mechanisms regulating these events.
Acknowledgements
The present study was financially supported by grant
no. CMRC-6 from the Vice-Chancellor for Research Affairs of Ahvaz
Jundishapur University of Medical Sciences.
References
|
1
|
American Diabetes Association, . Diagnosis
and classification of diabetes mellitus. Diabetes Care. 29:(Suppl
1). S43–S48. 2006.PubMed/NCBI
|
|
2
|
Szkudelski T, Zywert A and Szkudelska K:
Metabolic disturbances and defects in insulin secretion in rats
with streptozotocin-nicotinamide-induced diabetes. Physiol Res.
62:663–670. 2013.PubMed/NCBI
|
|
3
|
Wang H, Kouri G and Wollheim CB: ER stress
and SREBP-1 activation are implicated in β-cell glucolipotoxicity.
J Cell Sci. 118:3905–3915. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Butler AE, Janson J, Bonner-Weir S, Ritzel
R, Rizza RA and Butler PC: β-cell deficit and increased β-cell
apoptosis in humans with type 2 diabetes. Diabetes. 52:102–110.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weir GC, Laybutt DR, Kaneto H, Bonner-Weir
S and Sharma A: Beta-cell adaptation and decompensation during the
progression of diabetes. Diabetes. 50:(Suppl 1). S154–S159. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Viollet B, Guigas B, Sanz Garcia N,
Leclerc J, Foretz M and Andreelli F: Cellular and molecular
mechanisms of metformin: An overview. Clin Sci (Lond). 122:253–270.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gunton JE, Delhanty PJ, Takahashi S and
Baxter RC: Metformin rapidly increases insulin receptor activation
in human liver and signals preferentially through insulin-receptor
substrate-2. J Clin Endocrinol Metab. 88:1323–1332. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cusi K, Consoli A and DeFronzo RA:
Metabolic effects of metformin on glucose and lactate metabolism in
noninsulin-dependent diabetes mellitus. J Clin Endocrinol Metab.
81:4059–4067. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hundal RS, Krssak M, Dufour S, Laurent D,
Lebon V, Chandramouli V, Inzucchi SE, Schumann WC, Petersen KF,
Landau BR, et al: Mechanism by which metformin reduces glucose
production in type 2 diabetes. Diabetes. 49:2063–2069. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Natali A and Ferrannini E: Effects of
metformin and thiazolidinediones on suppression of hepatic glucose
production and stimulation of glucose uptake in type 2 diabetes: A
systematic review. Diabetologia. 49:434–441. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gregorio F, Filipponi P, Ambrosi F,
Cristallini S, Marchetti P, Calafiore R, Navalesi R and Brunetti P:
Metformin potentiates B-cell response to high glucose: An in vitro
study on isolated perfused pancreas from normal rats. Diabete
Metab. 15:111–117. 1989.PubMed/NCBI
|
|
12
|
Marchetti P, Scharp DW, Giannarelli R,
Benzi L, Cicchetti P, Ciccarone AM, Lacy PE and Navalesi R:
Metformin potentiates glucose-stimulated insulin secretion.
Diabetes Care. 19:781–782. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lacy PE, Kostianovsky M and Louis S:
Method for the isolation of intact islets of Langerhans from the
rat pancreas. Diabetes. 16:35–39. 1967. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perfetti R, Zhou J, Doyle ME and Egan JM:
Glucagon-like peptide-1 induces cell proliferation and
pancreatic-duodenum homeobox-1 expression and increases endocrine
cell mass in the pancreas of old, glucose-intolerant rats.
Endocrinology. 141:4600–4605. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Panza JL, Wagner WR, Rilo HL, Rao RH,
Beckman EJ and Russell AJ: Treatment of rat pancreatic islets with
reactive PEG. Biomaterials. 21:1155–1164. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marselli L, Dotta F, Piro S, Santangelo C,
Masini M, Lupi R, Realacci M, del Guerra S, Mosca F, Boggi U, et
al: Th2 cytokines have a partial, direct protective effect on the
function and survival of isolated human islets exposed to combined
proinflammatory and Th1 cytokines. J Clin Endocrinol Metab.
86:4974–4978. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lupi R, Del Guerra S, Marselli L, Bugliani
M, Boggi U, Mosca F, Marchetti P and Del Prato S: Rosiglitazone
prevents the impairment of human islet function induced by fatty
acids: Evidence for a role of PPARgamma2 in the modulation of
insulin secretion. Am J Physiol Endocrinol Metab. 286:E560–E567.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Janjic D and Wollheim CB: Islet cell
metabolism is reflected by the MTT (tetrazolium) colorimetric
assay. Diabetologia. 35:482–485. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang ZQ, Lu FE, Leng SH, Fang XS, Chen G,
Wang ZS, Dong LP and Yan ZQ: Facilitating effects of berberine on
rat pancreatic islets through modulating hepatic nuclear factor 4
alpha expression and glucokinase activity. World J Gastroenterol.
14:6004–6011. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fernandez-Alvarez J, Conget I, Rasschaert
J, Sener A, Gomis R and Malaisse WJ: Enzymatic, metabolic and
secretory patterns in human islets of type 2
(non-insulin-dependent) diabetic patients. Diabetologia.
37:177–181. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deng S, Vatamaniuk M, Huang X, Doliba N,
Lian M-M, Frank A, Velidedeoglu E, Desai NM, Koeberlein B, Wolf B,
et al: Structural and functional abnormalities in the islets
isolated from type 2 diabetic subjects. Diabetes. 53:624–632. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Marchetti P, Del Guerra S, Marselli L,
Lupi R, Masini M, Pollera M, Bugliani M, Boggi U, Vistoli F, Mosca
F, et al: Pancreatic islets from type 2 diabetic patients have
functional defects and increased apoptosis that are ameliorated by
metformin. J Clin Endocrinol Metab. 89:5535–5541. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Leclerc I, Woltersdorf WW, da Silva Xavier
G, Rowe RL, Cross SE, Korbutt GS, Rajotte RV, Smith R and Rutter
GA: Metformin, but not leptin, regulates AMP-activated protein
kinase in pancreatic islets: Impact on glucose-stimulated insulin
secretion. Am J Physiol Endocrinol Metab. 286:E1023–E1031. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin J-M, Fabregat ME, Gomis R and Bergsten
P: Pulsatile insulin release from islets isolated from three
subjects with type 2 diabetes. Diabetes. 51:988–993. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Richardson H, Campbell SC, Smith SA and
Macfarlane WM: Effects of rosiglitazone and metformin on pancreatic
beta cell gene expression. Diabetologia. 49:685–696. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kefas BA, Cai Y, Kerckhofs K, Ling Z,
Martens G, Heimberg H, Pipeleers D and Van de Casteele M:
Metformin-induced stimulation of AMP-activated protein kinase in
β-cells impairs their glucose responsiveness and can lead to
apoptosis. Biochem Pharmacol. 68:409–416. 2004. View Article : Google Scholar : PubMed/NCBI
|