Introduction
Cancer is a complex genomic disease with
multi-system disorders. In recent years, lung cancer has been one
of the highest incidences of cancer, with 1.59 million people
succumbing to lung cancer in 2012 worldwide (1,2). Although a
great deal of progressive treatment has been applied clinically,
including radiotherapy, chemotherapy and biotherapy, locally
advanced unresectable lung cancer accounts for ~35% of lung cancer
diagnoses, for which treatment remains a challenge. It has been
observed that ~90% of patients succumb from metastasis rather than
primary tumors (3,4). Therefore, identification of a sensitively
predictive tumor marker and understanding of the underlying
molecular mechanisms during tumor stage and metastasis have a major
importance and may propose an effective therapeutic strategy
against non-small cell lung cancer (NSCLC).
Non-coding RNA (ncRNA) regions, which were once
thought to be non-functional, account for 98% of the transcriptome
of the human genome (5,6); however, it has been found that the ncRNA
act as a regulator in flora and fauna biology. Recently, the
association of non-coding small regulatory RNAs and tumorigenesis
has attracted increasing attention. An increasing number of studies
have confirmed that the non-coding small RNA, including the
microRNAs (miRNAs), short interfering RNAs (siRNAs) or
PIWI-associated RNAs (piRNAs or piRs), have an important role in
carcinogenesis (7–9). The research of miRNA and carcinogenesis
is the most sufficient among these non-coding small regulatory
RNAs, and has identified that the miRNAs may be oncogenes or tumor
suppressors in carcinogenesis by regulating their target mRNA
(10–12). However, the study regarding piRNA and
carcinogenesis remains to be elucidated.
piRNAs are a type of novel non-coding small RNA with
a length of 25–33 nucleotides. They have biological roles through
the specific combination with the PIWI protein and are produced by
the Dicer-independent manner (13–15). The
crucial roles of piRNA were involved in maintaining DNA integrity,
regulation of epigenetics and germline stem cells, the
differentiation of embryonic development, and the occurrence and
development of diseases. Several previous studies have identified
that analogous with the miRNAs regulating tumorigenesis, the
expression levels of piRNAs were also up and downregulated in
certain cancer tissue and cell lines, revealing that they may have
oncogene or antioncogenic roles in carcinogenesis. For example, the
expression levels of piR-49322 (16),
piR-823 (17), piR-651 (18), piR-932 (19), piR-4987, piR-20365, piR-20485,
piR-20582 (20), piR-DQ594040
(21) and piR-Hep1 (22) were up and downregulated in breast
cancer, cervical cancer, gastric cancer, hepatocellular carcinoma,
multiple myeloma and others. piR-651, upregulated in several cancer
tissues and cell lines including NSCLC, may be an oncogene in
carcinogenesis (18). However, the
regulatory mechanism of piR-651 and NSCLC carcinogenesis remains to
be elucidated. Therefore, the present study focused on the
mechanism of piR-651 in regulating carcinogenesis in lung
cancer.
Materials and methods
Cell lines and culture conditions
The human high metastasis lung cancer cell line,
95-D, was purchased from the Shanghai Institute of Biochemistry and
Cell Biology, Chinese Academy of Science (Shanghai, China). 95-D
cells were cultured with Dulbecco's modified Eagle's medium (DMEM)
(Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 100 U/ml penicillin, 100 µg/ml streptomycin and
10% fetal bovine serum (FBS) (PAA Laboratories, Pasching, Austria).
Cells were growth at 37°C in a humidified atmosphere of 5%
CO2.
Cell transfection
The piR-651 inhibitor (Shanghai GenePharma Co.,
Shanghai, China) was transfected into 95-D cells to decrease the
expression of endogenous piR-651 and detect the effect of piR-651
on carcinogenesis. The specific 2′-O-methyl oligo RNA
oligonucleotides were synthesized and purified by Shanghai
GenePharma Co. Their sequences were as follows:
5′-GACGCUUUCCAAGGCACGGGCCCCUCUCU-3′ (piR-651 inhibitor) and
5′-CAGUACUUUUGUGUAGUACAA-3′ [non-specific sequences for the
negative control (NC)]. In brief, the cells were seeded in 6-well
plates 1 day before transfection. When the cells reached 50–60%
confluence, 100 pmol of the piR-651 inhibitor or 100 pmol of NC was
transfected using the Lipofectamine 2000 reagent (Invitrogen). The
transfecting rate was determined by fluorescence microscope and
flow cytometry (BD LSR II; Becton-Dickinson, Franklin Lakes, NJ,
USA).
Transwell invasion and migration
assays
A Transwell chamber with 8-mm membrane pores
(Corning Costar, Inc., Corning, NY, USA) was used to test the
effect of piR-651 on the migration and invasion in 95-D cells. For
migration, 3–4×104 transfected cells per well in
serum-free medium were placed into the upper chamber with the
non-coated membrane. For invasion, 5–6×104 cells in
serum-free medium were seeded in the upper chamber, which was
pre-coated with Matrigel (BD Biosciences, Bedford, MA, USA). The
assays were conducted three independent times.
Wound-healing assays
The 95-D cells were seeded in 6-well plates to near
confluence. A linear wound was carefully generated by moving a
10-µl sterile pipette tip across the confluent cell monolayer, and
the cell fragments were removed by washing with phosphate-buffered
saline (PBS) and incubated with DMEM without FBS. Subsequently,
images of the wounded monolayers were captured 48 h after
wounding.
Proliferation assay
To observe the effect of piR-651 on cell
proliferation, the MTT assay was used. Each group was performed in
six repeated wells in this assay. 95-D cell line was cultured in
96-well plates for 24 h and transfected with the piR-651 inhibitor
and NC, respectively. After transfection at 12, 24, 48 and 72 h,
the optical density (OD) values of each dose were determined. In
total, 20 µl MTT (Sigma-Aldrich, St. Louis, MO, USA) solution was
added into each of the 96-wells. After the cells were incubated at
37°C for 4 h in the dark, the supernatant was removed and 150 µl of
dimethylsulphoxide (Thermo Fisher Scientific, Loughborough, UK) was
added to dissolve the crystalization. The 96-well plate was mildly
agitated on the shaking table for ≥10 min. The OD values were
measured at 490 nm using a Wellscan reader (Labsystems, Santa Fe,
NM, USA). Growth proliferation was calculated as a percentage as
follows: (OD experiment/OD control) × 100%. Three independent
experiments were performed.
Cell apoptosis analysis
In the cell apoptosis assays, cells were collected
and washed with cold PBS twice after 24 h of transfection.
Subsequently, 5×105 cells/ml were stained with Annexin
V-propidium iodide in the dark, according to the manufacturer's
protocol (Biosea Biotechnology, Co., Beijing, China). The cells
were suspended in 300 µl binding buffer and assessed using flow
cytometry (BD Biosciences, San Jose, CA, USA) in 1 h. All the
assays were performed in triplicate.
Western blotting
To quantify the expression levels of
apoptosis-related proteins following downregulation of piR-651,
western blotting was used. In summary, the cells were collected 48
h after transfection and were lysed with radioimmunoprecipitation
assay buffer (Beyotime Biotechnology, Jiangsu, China), and
subsequently the lysates underwent centrifugation at 12,000 × g for
10 min at 4°C. The supernatants were collected, boiled and
subsequently separated by sodium dodecyl
sulfate-polyacrylamidegels, blotted onto polyvinylidene fluoride
membrane (Millipore Corporation, Bedford, MA, USA) and finally
detected with appropriate antibodies for bax (mouse anti-human
monoclonal antibody; cat. no. sc-7480)/B-cell lymphoma-2 (bcl-2;
mouse anti-human monoclonal antibody; cat. no. sc-7382), β-actin
(mouse anti-human monoclonal antibody; cat. no. sc-58679; all from
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) or from for
caspase-p17 (rabbit anti-human polyclonal antibody; cat. no.
25546-1-AP; Proteintech Group, Inc., Wuhan, China). Two or three
independent experiments were performed for this analysis.
Statistical analysis
Statistical analysis was performed using the
Statistical Program for Social Sciences (SPSS) version 17.0 (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Decrease in the expression of piR-651
decreases the cell proliferation rate in 95-D cells
Numerous studies have identified that piR-651 was
upregulated in lung cancer tissues and cell lines by deep
sequencing analysis. Therefore, the expression of endogenous
piR-651 was decreased by transfecting with its inhibitor. The
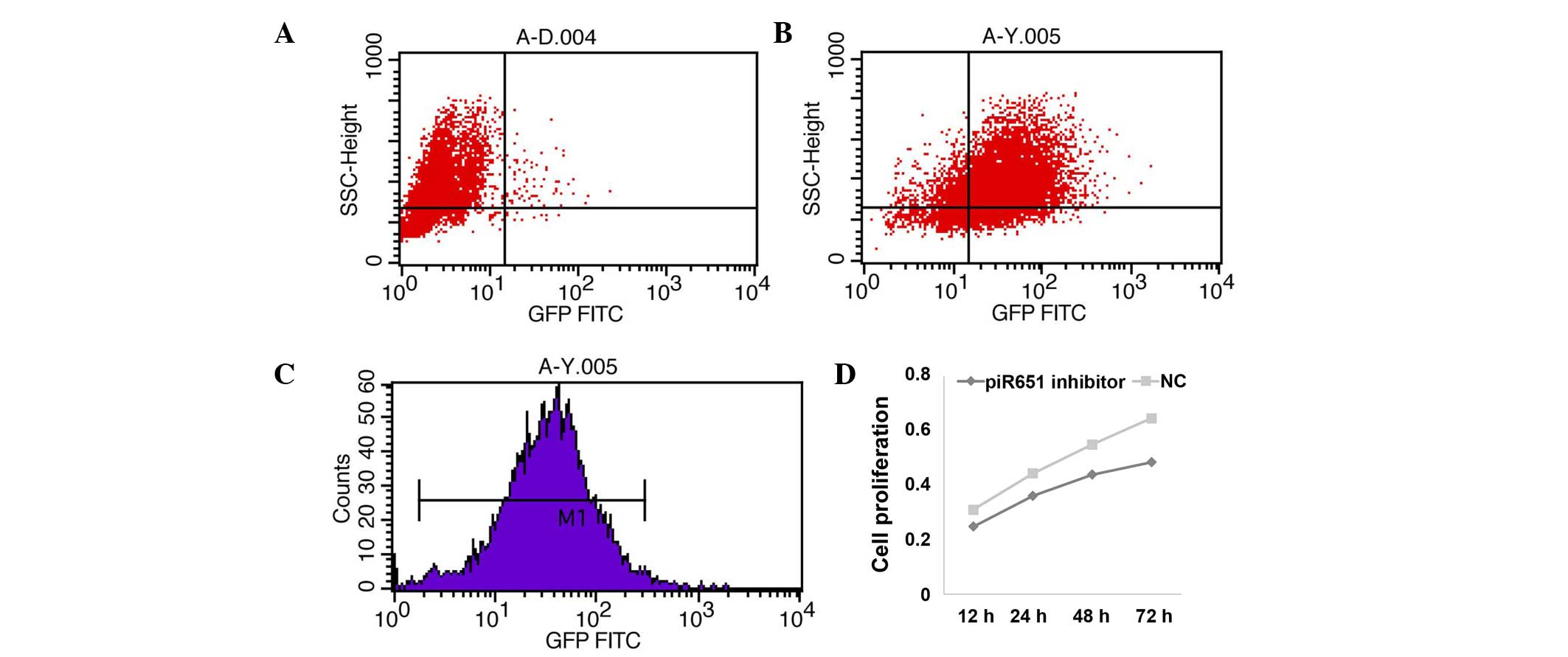
transfecting rate was detected using the FAM genes. The results
showed that 82.5% of all cells had fluorescent tags, and the mean
fluorescence intensity was 31.87% (Fig.
1A–C). Cell proliferation was suppressed following transfection
of the piR-651 inhibitor and the proliferation rate was decreased
in a time-dependent manner in the 95-D cells (Fig. 1D) (P<0.001).
Decrease in the expression of piR-651
induces cell apoptosis and regulates the apoptosis-related protein
expression level in 95-D cells
Numerous previous studies have confirmed that
apoptosis has an important role in the maintenance of the internal
environment and this balance is lost in a number of tumor patients.
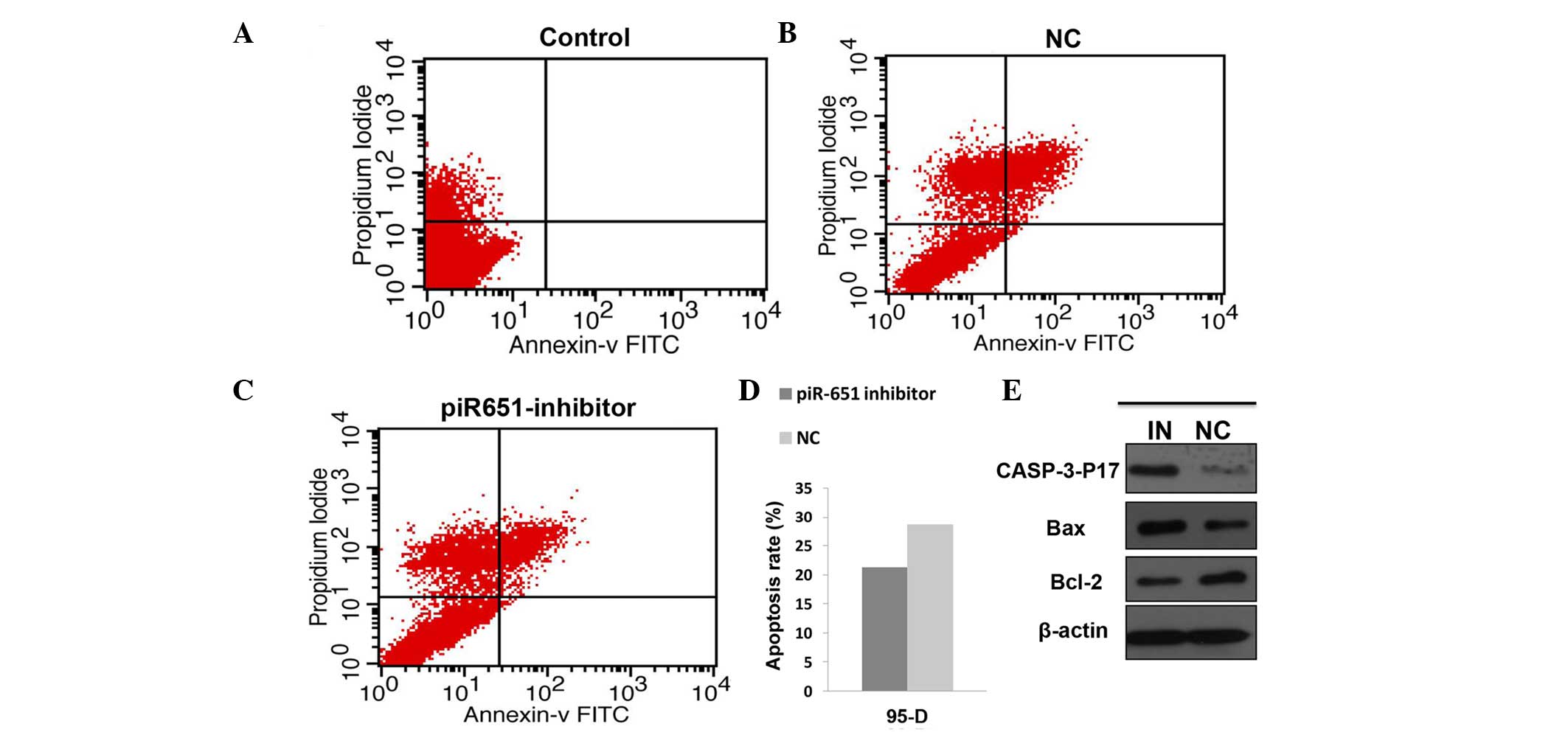
In the present study, the cell apoptosis rate in the piR-651
inhibitor cells was evidently higher compared to the NC in the 95-D
cell line (P<0.001). The expression level of the
apoptosis-related protein was assessed by western blotting. The
expression of piR-651 was reduced by transfecting the piR-651
inhibitor and the transfection rate was assessed by observing the
fluorescence. The expression levels of caspase-p17 (P=0.022) and
bax (P<0.001) were upregulated and bcl-2 (P<0.001) was
downregulated (Fig. 2).
Decrease in the expression level of
piR-651 restrains the cell ability of migration and invasion
Numerous previous studies have shown that the
majority of patients succumb from metastasis rather than primary
tumors. Metastasis and invasion are some of the most important
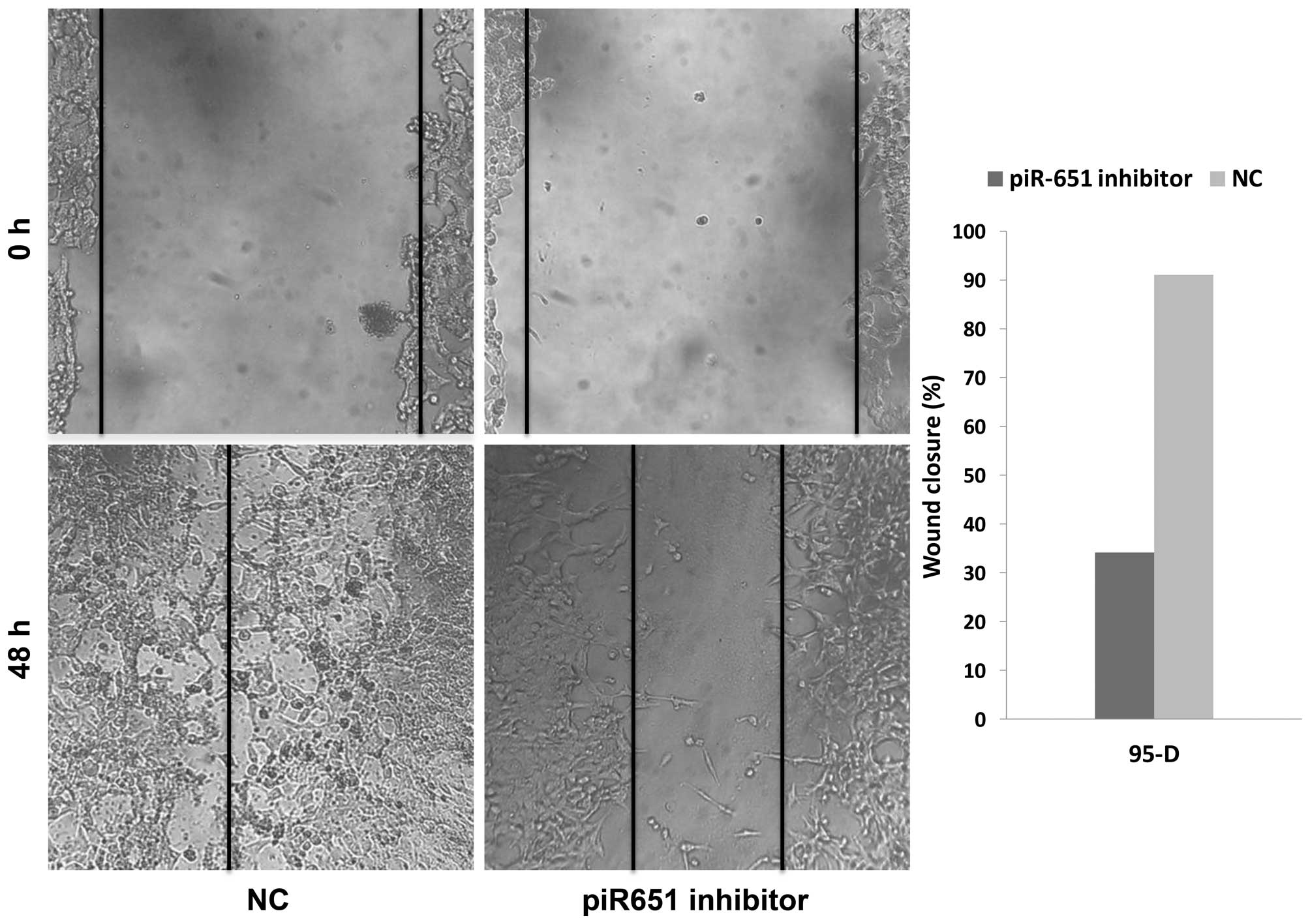
features of tumor cells. In the present study, the number of
migrating cells in the piR-651 inhibitor cells was evidently less
compared to the NC in 95-D (Fig. 3)
(P<0.001). As shown in Fig. 3,
downregulation of piR-651 impeded the invasion of 95-D cells
compared with that in the control cells (P<0.001). In the
wound-healing assays, the wound closure (%) in the piR-651
inhibitor was larger compared to the NC, as shown in Fig. 4 (P=0.001). These experiments
illustrated that piR-651 may have an important role in regulating
the ability of migration and invasion.
Discussion
In recent years, the research of non-coding small
RNA, including miRNA, siRNA and piRNA has become a significant area
of interest in the field of biological gene research. Increasing
evidence has demonstrated that the abnormal non-coding small RNA
expression was detected in numerous human cancers and cell lines,
including lung cancer, demonstrating their various functions by
regulating several biological characteristics (23). Different from the miRNAs, by
collectively regulating a substantial fraction of the
transcriptome, the piRNAs mainly protect the genome from
transposons (24). A previous study
confirmed that the piRNAs are mainly expressed in germline cells,
but several studies also demonstrated that piRNAs are expressed in
certain somatic cells and cancer cells (25,26).
Therefore, with the development of deep sequencing, several studies
have demonstrated that the piRNAs were dysregulated in a number of
cancer tissue and cell lines (17–22).
A previous study confirmed that piR-651 was
upregulated in gastric, colon, lung and breast cancer tissues and
cell lines compared with the paired non-cancerous tissues (18). Therefore, we speculated that piR-651
had a role in tumorigenesis as an oncogene. However, the mechanism
of piR-651 in regulating carcinogenesis remains to be elucidated.
In the present study, the cell proliferation rate was evidently
decreased in a time-dependent manner following transfection with
the piR-651 inhibitor in 95-D cells. The flow cytometry assays
confirmed that the piR-651 inhibitor induced cell apoptosis, and
therefore we deduced that the biological function of piR-651 was
closely associated with proliferation and apoptosis of tumor cells.
Subsequently, up and downregulation of the apoptosis-related
protein following a change in the expression of endogenous piR-651
confirmed this deduction. A previous study identified that the
caspase family had a vital role in regulating cell apoptosis, and
caspase-3 was the main executor of this process (27). Bax is a member of the bcl-2 family,
which is involved in the process of cell apoptosis. When cell
apoptosis was induced, bcl-2 was downregulated and bax was
overexpressed with the removal from cytochylema to mitochondria and
karyotheca (28). In the present
study, the level of expression of cleaved caspase-3 and bax was
upregulated and bcl-2 was downregulated following a decrease in the
expression of endogenous piR-651. Therefore, we can infer that
piR-651 regulated tumorigenesis by inhibiting apoptosis and
altering the expression level of the apoptosis-related protein.
Invasion and migration, which are usually recognized
as the main reason for the high recurrence and fatality rates of
lung cancer, restrict the efficacy of surgery and other therapies
(29). In the present study, the
ability of invasion and migration was reduced following a decrease
in the expression of endogenous piR-651 in 95-D cells. Therefore,
we can infer that piR-651 has an important role in regulating
invasion and migration.
In conclusion, the present study demonstrated that
piR-651 has an important role in human NSCLC pathogenesis. Reduced
endogenous piR-651 expression may restrain the tumor progression by
inhibiting cell proliferation, migration and invasion, and inducing
cell apoptosis. These results indicate that targeting endogenous
piR-651 may constitute a potential treatment modality for lung
cancer.
References
|
1
|
WHO International Agency for Research on
Cancer (IARC): Globocan 2012: Estimated cancer incidence, mortality
and prevalence worldwide in 2012. WHO. Geneva: 2012.
|
|
2
|
Chen B, Zhang B, Xia L, Zhang J, Chen Y,
Hu Q and Zhu C: Knockdown of eukaryotic translation initiation
factor 4E suppresses cell growth and invasion, and induces
apoptosis and cell cycle arrest in a human lung adenocarcinoma cell
line. Mol Med Rep. 12:7971–7978. 2015.PubMed/NCBI
|
|
3
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun Y, Ai X, Shen S and Lu S:
NF-κB-mediated miR-124 suppresses metastasis of non-small-cell lung
cancer by targeting MYO10. Oncotarget. 2:1–11. 2015.
|
|
5
|
Lander ES, Linton LM, Birren B, Nusbaum C,
Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, et al:
International Human Genome Sequencing Consortium: Initial
sequencing and analysis of the human genome. Nature. 409:860–921.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohno S: So much ‘junk’ DNA in our genome.
Brookhaven Symp Biol. 23:366–370. 1972.PubMed/NCBI
|
|
7
|
Peters L and Meister G: Argonaute
proteins: Mediators of RNA silencing. Mol Cell. 26:611–623. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hutvagner G and Simard MJ: Argonaute
proteins: Key players in RNA silencing. Nat Rev Mol Cell Biol.
9:22–32. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siomi MC, Sato K, Pezic D and Aravin AA:
PIWI-interacting small RNAs: The vanguard of genome defence. Nat
Rev Mol Cell Biol. 12:246–258. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Pan X, Cobb GP and Anderson TA:
MicroRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dieckmann KP, Spiekermann M, Balks T, Flor
I, Löning T, Bullerdiek J and Belge G: MicroRNAs miR-371-3 in serum
as diagnostic tools in the management of testicular germ cell
tumours. Br J Cancer. 107:1754–1760. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takahashi M, Cuatrecasas M, Balaguer F,
Hur K, Toiyama Y, Castells A, Boland CR and Goel A: The clinical
significance of MiR-148a as a predictive biomarker in patients with
advanced colorectal cancer. PLoS One. 7:e466842012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aravin A, Gaidatzis D, Pfeffer S,
Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ,
Kuramochi-Miyagawa S, Nakano T, et al: A novel class of small RNAs
bind to MILI protein in mouse testes. Nature. 442:203–207.
2006.PubMed/NCBI
|
|
14
|
Girard A, Sachidanandam R, Hannon GJ and
Carmell MA: A germline-specific class of small RNAs binds mammalian
Piwi proteins. Nature. 442:199–202. 2006.PubMed/NCBI
|
|
15
|
Grivna ST, Beyret E, Wang Z and Lin H: A
novel class of small RNAs in mouse spermatogenic cells. Genes Dev.
20:1709–1714. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu Y, Li C, Zhang K, Sun H, Tao D, Liu Y,
Zhang S and Ma Y: Identification of piRNAs in Hela cells by massive
parallel sequencing. BMB Rep. 43:635–641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng J, Deng H, Xiao B, Zhou H, Zhou F,
Shen Z and Guo J: piR-823, a novel non-coding small RNA,
demonstrates in vitro and in vivo tumor suppressive activity in
human gastric cancer cells. Cancer Lett. 315:12–17. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng J, Guo JM, Xiao BX, Miao Y, Jiang Z,
Zhou H and Li QN: piRNA, the new non-coding RNA, is aberrantly
expressed in human cancer cells. Clin Chim Acta. 412:1621–1625.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Ren Y, Xu H, Pang D, Duan C and
Liu C: The expression of stem cell protein Piwil2 and piR-932 in
breast cancer. Surg Oncol. 22:217–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang G, Hu H, Xue X, Shen S, Gao E, Guo
G, Shen X and Zhang X: Altered expression of piRNAs and their
relation with clinicopathologic features of breast cancer. Clin
Transl Oncol. 15:563–568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chu H, Hui G, Yuan L, Shi D, Wang Y, Du M,
Zhong D, Ma L, Tong N, Qin C, et al: Identification of novel piRNAs
in bladder cancer. Cancer Lett. 356(2 Pt B): 561–567. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Law PT, Qin H, Ching AK, Lai KP, Co NN, He
M, Lung RW, Chan AW, Chan TF and Wong N: Deep sequencing of small
RNA transcriptome reveals novel non-coding RNAs in hepatocellular
carcinoma. J Hepatol. 58:1165–1173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Yan S, Pei C and Cui Y: Decreased
microRNA-132 and its function in human non-small cell lung cancer.
Mol Med Rep. 11:3601–3608. 2015.PubMed/NCBI
|
|
24
|
Grimson A, Srivastava M, Fahey B,
Woodcroft BJ, Chiang HR, King N, Degnan BM, Rokhsar DS and Bartel
DP: Early origins and evolution of microRNAs and Piwi-interacting
RNAs in animals. Nature. 455:1193–1197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meister G: Argonaute proteins: Functional
insights and emerging roles. Nat Rev Genet. 14:447–459. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Siddiqi S and Matushansky I: Piwis and
piwi-interacting RNAs in the epigenetics of cancer. J Cell Biochem.
113:373–380. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cryns V and Yuan J: Proteases to die for.
Genes Dev. 12:1551–1570. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang F, Huang M, Zhang H, Zhang C, Zhang
D and Zhou G: Changes in apoptotic factors and caspase activation
pathways during the postmortem aging of beef muscle. Food Chem.
190:110–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Y, Mayo MW, Xiao A, Hall EH, Amin EB,
Kadota K, Adusumilli PS and Jones DR: Loss of BRMS1 promotes a
mesenchymal phenotype through NF-κB-dependent regulation of Twist1.
Mol Cell Biol. 35:303–317. 2015. View Article : Google Scholar : PubMed/NCBI
|