Introduction
Genetic factors that may alter the repair of damaged
gastric and colonic epithelial cell DNA, resulting in their
transformation into cancer cells remain poorly understood.
Accumulation of permanently generated reactive oxygen species
during cellular metabolism and extracellular processes may lead to
carcinogenesis caused by oxidative DNA damage. p53, a tumor
suppressor protein encoded in humans by the p53 gene,
regulates the cell cycle and preserves the stability of the human
genome to prevent cancer initiation (1,2). Gastric
cancer (GC) is the fourth most common malignancy and the second
primary cause of cancer mortality, resulting in >800,000
mortalities worldwide annually (3,4). In addition
to environmental factors, genetic variants are significant. As a
crucial tumor suppressor gene, p53 is critical in cell-cycle
regulation, DNA repair, cellular differentiation and apoptosis in
various types of cancer. Furthermore, variants of p53 gene
exert an important effect on tumor development and cancer risk
(5–7).
Recently, numerous studies have evaluated the
interaction between p53 and its regulators in
carcinogenesis. Of these regulators, the murine double minute clone
2 (MDM2) proto-oncogene, an important negative regulator of the
p53 gene, has become a point of interest (8). MDM2 is a major regulator of p53
function. p53 and MDM2 act in a feedback cycle
whereby p53 activates MDM2 at the transcriptional level,
while MDM2 binds, obstructs and degrades the p53
protein through E3 ligase activity (9–11). The human
MDM2 gene is located on chromosome 12q13-14 (genomic size,
34 kb) and contains two promoters, the constituent promoter and the
p53-responsive intronic promoter (12). The inherently occurring sequence
variation in the MDM2 promoter results in a changed
MDM2 protein expression leading to the alteration in
p53 mediated tumor suppression activity. A single nucleotide
polymorphism (SNP; T309G) located in the first intron of the core
promoter region of the MDM2 gene influences binding of the
transcription factor, Sp1. Sp1 binds with higher affinity to the G
allele than to the T allele, which results in increased
transcription of the MDM2 gene and higher expression levels
of MDM2 protein, thereby obstructing the tumor suppressor function
of p53. This SNP leads to increased MDM2 mRNA and
protein expression levels and, as a result, p53 inhibition.
Additionally, there is a disposition, which demonstrated earlier
cancer inception in certain groups of patients harboring the GG or
TG genotype for MDM2 T309G (13). The MDM2 T309G polymorphism has
been reported to be associated with certain tumors, such as colon,
gastric and hepatocellular cancer (14–16).
In the present study, the aim was to investigate the
association between genetic polymorphisms of the MDM2 gene,
a major regulator of p53 function, and primary GC risk in a
Turkish population.
Materials and methods
Study population
The study was approved by the local ethic committee
and informed consent was obtained from all patients. All subjects
agreed to participate in the study and completed a short
questionnaire, which included questions about their occupation,
tobacco use, alcohol consumption and family history of cancer. In
the present study, a total of 132 individuals (65 GC patients and
67 control subjects) were investigated. Blood samples were
collected from the 65 patients who were diagnosed with GC at the
Department of General Surgery, Cumhuriyet University, Faculty of
Medicine (Sivas, Turkey). The diagnosis of GC was histologically
confirmed and the tumor types were classified according to WHO
guidelines, (https://www.iarc.fr/en/publications/pdfs-online/pat-gen/bb2/bb2-chap3.pdf)
No age and sex restrictions were applied for the selection of
healthy volunteers, who were free of any chronic diseases, lived in
the same geographic area, and had no history of cancer. All cases
and controls were born and lived in Turkey.
DNA isolation
Peripheral blood samples (2 ml) were obtained and
collected into citrate-containing tubes from all subjects. The DNA
was extracted from whole blood using the salting out procedure as
soon as the samples reached the laboratory (17).
MDM2 genotyping
The MDM2 T309G polymorphism was analyzed
using the polymerase chain reaction (PCR)-restriction fragment
length polymorphism method. The following primers were used:
Forward, 5′-CGCGGGAGTTCAGGGTAAAG-3′ and reverse,
5′-CTGAGTCAACCTGCCCACTG-3′ to amplify the MDM2 polymorphism.
Amplification was performed using the following: 25 pmol each
primer, 200 mM total dNTP, 1.5 mM MgCI2, 1X PCR buffer
and 2.5 U Taq DNA polymerase and 50–100 ng DNA in a total volume of
50 µl. The PCR program was initiated with denaturation at 95°C for
5 min, followed by 30 cycles of 94°C for 60 sec, 55°C for 60 sec
(annealing) and 72°C for 60 sec (extension). The PCR was completed
with a final extension cycle at 72°C for 5 min. Following
confirmation of PCR amplification by 1.5% agarose gel
electrophoresis, the amplified product was digested overnight with
MspA1I restriction enzyme at 37°C and electrophoresed on 3%
agarose gel stained with ethidium bromide and visualized under UV
light. Genotypes were identified for the polymorphism as TT (157
bp), TG (157, 110 and 47 bp), or GG (110 and 47 bp) (18).
Statistical analysis
All statistical analyses were performed using SPSS
version 11 (SPSS, Inc., Chicago, IL, USA). Genotype-associated odds
ratios (ORs), their corresponding 95% confidence intervals (CIs),
and associated P-values were estimated via unconditional logistic
regression. Differences in the distributions of demographic
characteristics between the cases and control subjects were
evaluated using Student's t-test. χ2 or Fischer's exact
test (two-sided) were used to compare the sex distribution, to test
the association between the genotypes and alleles in relation to
the controls, and to test for deviation of the genotype
distribution from Hardy-Weinberg equilibrium. Pearson's
χ2 test was used to determine whether there were any
significant differences in allele and genotype frequencies between
patients and control subjects. Logistic regression procedures were
performed to assess the interaction between age, sex and all
genotypes. In addition, the recessive or dominant effect of the
MDM2 genotype on risk was estimated, and statistical
analysis was performed on the relative risk of the GG genotype
against the TG + TT genotype or the TG + GG genotype against the TT
genotype.
Results
In the present study, the association between the
MDM2 polymorphism and GC was investigated in a Turkish population.
The polymorphism in the MDM2 gene, T309G has been
determined in 65 patients with GC and in 67 healthy control
subjects using the PCR-RFLP method.
The demographic characteristics of the study
population are presented in Table I.
The mean age of the GC patients and control subjects were
62.25±10.95 years (males, 62.32±10.55; females, 61.92±13.08) and
62.34±10.54 years (males, 62.58±9.86; females, 61.43±13.19),
respectively. The percentage of males and females in the cases were
81.5 and 18.5%, respectively (Table
I). No statistically significant differences were identified
between the cases and control subjects by age and sex (P>0.05).
Nor were there statistically significant associations between the
cases and control subjects for smoking history (OR=0.99; 95% CI,
0.50–1.97; P=0.989; Table II).
Furthermore, no statistically significant differences were
identified regarding alcohol consumption among the cases and
control subjects (OR=2.35; 95% CI, 0.88–6.27; P=0.082) (Table II). Comparison between GC cases and
control subjects indicated statistically significant differences in
family history of cancer (OR=0.17; 95% CI, 0.05–0.56; P=0.01;
Table II). Individuals with the GG
genotype were associated with a higher risk for GC and the GG
polymorphism was significantly associated with this disease
(OR=4.58; 95% CI, 1.18–17.79; P=0.022; Table III). Analysis of the MDM2 gene
T309G polymorphisms is presented in Fig. 1.
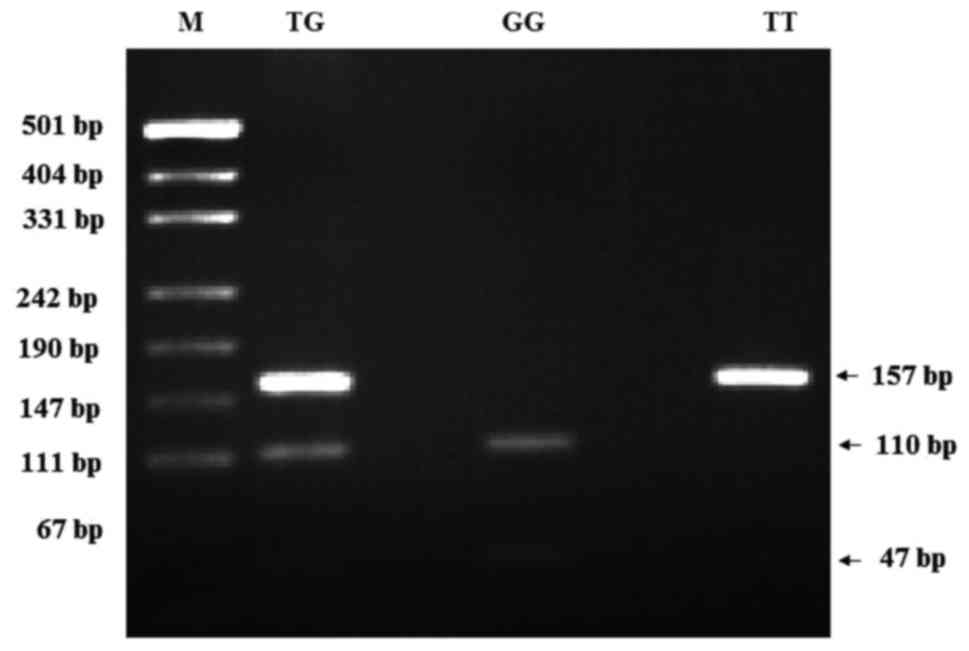 | Figure 1.PCR-restriction fragment length
polymorphism patterns of polymorphisms of MDM2 T309G. M:
pUCI19/Msp I DNA ladder (501, 489, 404, 331, 242, 190, 147, 111 and
110 bp); MDM2 PCR product, 157 bp, TG (157, 110 and 47 bp); GG (110
and 47 bp); TT (157 bp). PCR, polymerase chain reaction; MDM2,
murine double minute-2. |
 | Table I.Demographics of control subjects
(n=67) and gastric cancer cases (n=65) recruited in the present
study. |
Table I.
Demographics of control subjects
(n=67) and gastric cancer cases (n=65) recruited in the present
study.
| Variable | Control, n (%) | Gastric cancer, n
(%) |
|---|
| Sex |
|
|
|
Males | 53 (79.1) | 53 (81.5) |
|
Females | 14 (20.9) | 12 (18.5) |
| Age (years) |
|
|
|
Range | 40–85 | 40–85 |
| Means ±
SD | 62.34±10.54 | 62.25±10.95 |
|
Males | 62.58±9.86 | 62.32±10.55 |
|
Females | 61.43±13.19 | 61.92±13.08 |
| Smoking
history |
|
|
|
Smoker | 31(46.3) | 30 (46.2) |
|
Males | 30 (56.6) | 28 (52.8) |
|
Females | 1 (7.1) | 2 (16.7) |
| Alcoholic drink
consumption |
|
|
|
Smoker | 7 (10.4) | 14 (21.5) |
|
Males | 7 (13.2) | 11 (20.8) |
|
Females | 0 | 3 (25.0) |
| Family history of
cancer | 18 (26.9) | 4 (6.2) |
| Table II.Distribution of selected variables in
gastric cancer cases and control subjects. |
Table II.
Distribution of selected variables in
gastric cancer cases and control subjects.
|
| Gastric cancer
(n=65) | Control (n=67) |
|
|
|---|
|
|
|
|
|
|
|---|
| Variable | n | % | n | % | P-value | Odds ratio |
|---|
| Smoking status |
|
|
|
|
|
|
| No | 35 | 53.8 | 36 | 53.7 | 0.989 | 0.99
(0.50–1.97) |
|
Yes | 30 | 46.2 | 31 | 46.3 |
|
|
| Drinking
status |
|
|
|
|
|
|
| No | 51 | 78.5 | 60 | 89.6 | 0.082 | 2.35
(0.88–6.27) |
|
Yes | 14 | 21.5 | 7 | 10.4 |
|
|
| Family history of
cancer |
|
|
|
|
|
|
| No | 61 | 93.8 | 41 | 73.1 | 0.010a | 0.17
(0.05–0.56) |
|
Yes | 4 |
6.2 | 18 | 26.9 |
|
|
| Table III.Stratification analyses between
murine double minute-2 T309G genotypes and gastric cancer
risk. |
Table III.
Stratification analyses between
murine double minute-2 T309G genotypes and gastric cancer
risk.
| MDM2
T309G | Controls
(n=67) | Gastric cancer
(n=65) | χ2 | P-value | Crude odds
ratio |
|---|
| Allele
frequency |
|
|
|
|
|
| T
allele | 65 | 47 | Ref. | 0.042 | 1.66
(0.99–2.81) |
| G
allele | 69 | 83 | 4.12 |
| Genotype
frequency |
|
|
|
|
|
| TT | 10 (14.9) | 4 (6.2) | Ref. | – | – |
| TG | 45 (67.2) | 39 (60.0) | 1.55 | 0.213a | 2.16
(0.62–7.45) |
| GG | 12 (7.9) | 22 (33.8) | 5.21 | 0.022 | 4.58
(1.18–17.79) |
| TG +
GG | 57 (85.1) | 61 (93.8) | 2.67 | 0.102 | 2.65
(0.79–9.01) |
| TT +
TG | 45 (67.2) | 39 (60.0) | 0.73 | 0.392 | 0.73
(0.36–1.49) |
Discussion
GC is the fourth most common cancer and the second
leading cause of cancer-associated mortality worldwide (4). GC is a complex trait caused by genetic
and environmental factors (19).
Genetic polymorphisms in the carcinogen detoxification, antioxidant
protection, DNA repair and cell proliferation processes are also
crucial in the development of GC (20).
In the current study, of 65 GC cases and 67 control
subjects, the crude OR of GC patients for family history of cancer
was 0.17, indicating an association between GC incidence and family
history of cancer (P=0.01; Table II).
To the best of our knowledge, GC and family history of cancer
association has not been proposed in previous studies. In addition,
GC patients and control subjects were evaluated by logistic
regression analysis for smoking habits. No statistically
significant difference was identified between GC patients and
smoking habits in the Turkish population investigated in the
current study (P=0.989; Table
II).
The MDM2 T309G polymorphism in GC patients
has been investigated in various different regions of the world;
however, to the best of our knowledge, no research has been
performed in Turkey. This study was the first study to evaluate the
polymorphic variants of MDM2 T309G polymorphism and risk of GC. The
MDM2 protein negatively regulates p53 function via multiple
independent mechanisms, such as by binding to transcription
activation domains, inhibiting acetylation, promoting nuclear
export and, most importantly, promoting proteasomal degradation via
ubiquitination (9,11,21,22). The MDM2 T309G polymorphism is a
single nucleotide T to G polymorphism located at the 309th
nucleotide in the MDM2 gene promoter. Over the last two
decades, it has been confirmed that the MDM2 T309G
polymorphism is capable of significantly altering the expression
levels of MDM2, thereby suppressing the p53 signaling
pathway. Furthermore, a number of epidemiological studies have
investigated the association between the MDM2 T309G
polymorphism and the risk of various types of cancer (23–26),
including renal, hepatocellular, endometrial, bladder and stomach
cancers. However, the results of previous studies regarding the
association between the MDM2 T309G polymorphism and GC risk
were different or even contradictory (27,28).
Recently, numerous studies demonstrated that significant
associations between the MDM2 T309G polymorphism and GC risk
were identified for the G and T alleles, and the GG and TT
genotypes. The G allele was revealed to act as a key factor for
increasing the risk of GC, which is consistent with the study by
Pan et al (29). In addition,
in a meta-analysis, Chen et al (30) proposed that the G allele of the MDM2
T309G polymorphism is a high risk factor for developing GC. In
the present study, a statistically significant difference was
identified in the GG genotype distribution between GC patients and
control subjects in the investigated Turkish population (OR=4.58;
95% CI, 1.18–17.79; P=0.022; Table
III). MDM2 T309G GG genotype was associated with an
increased risk of gastric carcinoma when compared with the TT
genotype or T carriers (P<0.01), and a joint effect between
MDM2 T309G GG and the Helicobacter pylori infection
was observed to intensify the GC risk (31). Tian et al (32) proposed that the GG genotype is
associated with a significantly increased risk of GC (OR=1.43; 95%
CI, 1.08–1.91; P=0.013). Consistent with these studies, the current
study identified that the overall GC risk for T309G GG was
significantly increased when it was compared with T carriers
(OR=4.58; 95% CI, 1.18–17.79; P=0.022), as presented in Table III. The distribution of MDM2
genotypes in different populations is provided in Table IV. While the genotype distribution of
the control group of the present study was similar to those of the
other studies (15,26–29,31,33) the case
groups differed.
| Table IV.Distribution of polymorphisms of
murine double minute-2 T309G genotype frequencies in
different populations. |
Table IV.
Distribution of polymorphisms of
murine double minute-2 T309G genotype frequencies in
different populations.
|
| Gastric cancer
(%) | Control (%) |
|
|
|---|
|
|
|
|
|
|
|---|
| Country | TT | TG | GG | TT | TG | GG | Year | (Refs.) |
|---|
| Japan | 23.9 | 45.9 | 30.2 | 22.6 | 55.0 | 22.4 | 2006 | (15) |
| China | 21.4 | 50.0 | 28.6 | 29.8 | 49.8 | 20.4 | 2007 | (33) |
| Korea | 26.8 | 46.0 | 27.2 | 20.4 | 50.8 | 28.8 | 2008 | (28) |
| China | 21.7 | 48.1 | 30.2 | 36.8 | 43.4 | 19.8 | 2009 | (31) |
| China | 28.5 | 46.2 | 25.4 | 31.5 | 54.3 | 14.2 | 2009 | (27) |
| China | 20.9 | 54.5 | 24.6 | 20.5 | 51.6 | 27.9 | 2011 | (26) |
| China | 30.1 | 45.3 | 24.6 | 34.7 | 51.6 | 13.8 | 2013 | (29) |
| Turkey | 6.2 | 60.0 | 33.8 | 14.9 | 67.2 | 17.9 | 2017 | Current study |
In conclusion, the present results indicate a
significant association between the MDM2 GG genotype and GC;
although, it is not possible to conclude that a single polymorphism
determines an individual's susceptibility to GC development.
Furthermore, the MDM2 GG genotype may be defined as an
independent marker for GC.
Acknowledgements
This study was funded by Cumhuriyet University
(grant no. CÜBAP T-547).
References
|
1
|
McBride OW, Merry D and Givol D: The gene
for human p53 cellular tumor antigen is located on chromosome 17
short arm (17p13). Proc Natl Acad Sci USA. 83:130–134. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Strachan T and Read AP: Human molecular
genetics. 2nd. Wiley-Liss; New York, NY: 1999, PubMed/NCBI
|
|
3
|
Nakao M, Matsuo K, Ito H, Shitara K,
Hosono S, Watanabe M, Ito S, Sawaki A, Iida S, Sato S, et al: ABO
genotype and the risk of gastric cancer, atrophic gastritis, and
Helicobacter pylori infection. Cancer Epidemiol Biomarkers
Prev. 20:1665–1672. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marte B: Cancer: Super p53. Nature.
420:2792002. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ventura A, Kirsch DG, McLaughlin ME,
Tuveson DA, Grimm J, Lintault L, Newman J, Reczek EE, Weissleder R
and Jacks T: Restoration of p53 function leads to tumour regression
in vivo. Nature. 445:661–665. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hong H, Takahashi K, Ichisaka T, Aoi T,
Kanagawa O, Nakagawa M, Okita K and Yamanaka S: Suppression of
induced pluripotent stem cell generation by the p53-p21 pathway.
Nature. 460:1132–1135. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Misra C, Majumder M, Bajaj S, Ghosh S, Roy
B and Roychoudhury S: Polymorphisms at p53, p73, and MDM2 loci
modulate the risk of tobacco associated leukoplakia and oral
cancer. Mol Carcinog. 48:790–800. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Haupt Y, Maya R, Kazaz A and Oren M: Mdm2
promotes the rapid degradation of p53. Nature. 387:296–299. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Honda R, Tanaka H and Yasuda H:
Oncoprotein MDM2 is a ubiquitin ligase E3 for tumor suppressor p53.
FEBS Lett. 420:25–27. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kubbutat MH, Jones SN and Vousden KH:
Regulation of p53 stability by Mdm2. Nature. 387:299–303. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Momand J and Zambetti GP: Mdm-2: ‘big
brother’ of p53. J Cell Biochem. 64:343–352. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bond GL, Hu W, Bond EE, Robins H, Lutzker
SG, Arva NC, Bargonetti J, Bartel F, Taubert H, Wuerl P, et al: A
single nucleotide polymorphism in the MDM2 promoter attenuates the
p53 tumor suppressor pathway and accelerates tumor formation in
humans. Cell. 119:591–602. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Menin C, Scaini MC, De Salvo GL, Biscuola
M, Quaggio M, Esposito G, Belluco C, Montagna M, Agata S, D'Andrea
E, et al: Association between MDM2-SNP309 and age at colorectal
cancer diagnosis according to p53 mutation status. J Natl Cancer
Inst. 98:285–288. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ohmiya N, Taguchi A, Mabuchi N, Itoh A,
Hirooka Y, Niwa Y and Goto H: MDM2 promoter polymorphism is
associated with both an increased susceptibility to gastric
carcinoma and poor prognosis. J Clin Oncol. 24:4434–4440. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoon YJ, Chang HY, Ahn SH, Kim JK, Park
YK, Kang DR, Park JY, Myoung SM, Kim DY, Chon CY and Han KH: MDM2
and p53 polymorphisms are associated with the development of
hepatocellular carcinoma in patients with chronic hepatitis B virus
infection. Carcinogenesis. 29:1192–1196. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miller SA, Dykes DD and Polesky HF: A
simple salting out procedure for extracting DNA from human
nucleated cells. Nucleic Acids Res. 16:12151988. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yilmaz M, Tas A, Kacan T, Sari M and Silig
Y: Is there a relation between murine double minute 2 T309G
polymorphism and lung cancer risk in the Turkish population? Turk J
Biochem. 42:123–129. 2017.
|
|
19
|
Crew KD and Neugut AI: Epidemiology of
upper gastrointestinal malignancies. Semin Oncol. 31:450–464. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Geng JS, Song HT and Wang WR: Diversity of
invasiveness and matrix metalloproteinases expression profile of
human gastric carcinoma xenografted in different tissue
environments. Zhonghua Bing Li Xue Za Zhi. 33:53–56. 2004.(In
Chinese). PubMed/NCBI
|
|
21
|
Brady M, Vlatkovic N and Boyd MT:
Regulation of p53 and MDM2 activity by MTBP. Mol Cell Biol.
25:545–553. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Freedman DA, Wu L and Levine AJ: Functions
of the MDM2 oncoprotein. Cell Mol Life Sci. 55:96–107. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hirata H, Hinoda Y, Kikuno N, Kawamoto K,
Suehiro Y, Tanaka Y and Dahiya R: MDM2 SNP309 polymorphism as risk
factor for susceptibility and poor prognosis in renal cell
carcinoma. Clin Cancer Res. 13:4123–4129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Terry K, McGrath M, Lee IM, Buring J and
De Vivo I: MDM2 SNP309 is associated with endometrial cancer risk.
Cancer Epidemiol Biomarkers Prev. 17:983–986. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olsson H, Hultman P, Rosell J, Söderkvist
P and Jahnson S: MDM2 SNP309 promoter polymorphism and p53
mutations in urinary bladder carcinoma stage T1. BMC Urol.
13:52013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang GX, Li YQ and Pan XL: Polymorphism
of MDM2 promoter, regulated by Helicobacter pylori
lipopolysaccharide, is associated with both an increased
susceptibility to gastric carcinoma and poor prognosis in Chinese
patients. J Gastroenterol Hepatol. 26:2802011.
|
|
27
|
Wang X, Yang J, Ho B, Yang Y, Huang Z,
Zhang Z and Zhang G: Interaction of Helicobacter pylori with
genetic variants in the MDM2 promoter, is associated with gastric
cancer susceptibility in Chinese patients. Helicobacter.
14:114–119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cho YG, Choi BJ, Song JH, Kim CJ, Cao Z,
Nam SW, Lee JY and Park WS: No association of MDM2 T309G
polymorphism with susceptibility to Korean gastric cancer patients.
Neoplasma. 55:256–260. 2008.PubMed/NCBI
|
|
29
|
Pan X, Li Y, Feng J, Wang X, Hao B, Shi R
and Zhang G: A functional polymorphism T309G in MDM2 gene promoter,
intensified by Helicobacter pylori lipopolysaccharide, is
associated with both an increased susceptibility and poor prognosis
of gastric carcinoma in Chinese patients. BMC Cancer. 13:1262013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen W, Wu Q and Ren H: Meta-analysis of
associations between MDM2 SNP309 polymorphism and gastric cancer
risk. Biomed Rep. 2:105–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Er LM, Zhang LW, Niu WW, Xu ZB, Du CQ, Gao
Y, Wang SP and Yuan L: Relation of MDM2 gene polymorphism and
Helicobacter pylori infection to gastric cardiac carcinoma.
Lin Chuang Hui Cui. 24:1594–1597. 2009.(In Chinese).
|
|
32
|
Tian X, Tian Y, Ma P, Sui C-G, Meng F-D,
Li Y, Fu L-Y, Jiang T, Wang Y, Ji FJ, et al: Association between
MDM2 SNP309 T>G and risk of gastric cancer: A meta-analysis.
Asian Pac J Cancer Prev. 14:1925–1929. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang M, Guo Y, Zhang X, Miao X, Tan W, Sun
T, Zhao D, Yu D, Liu J and Lin D: Interaction of P53 Arg72Pro and
MDM2 T309G polymorphisms and their associations with risk of
gastric cardia cancer. Carcinogenesis. 28:1996–2001. 2007.
View Article : Google Scholar : PubMed/NCBI
|














