Introduction
Cisplatin remains the primary and most frequently
used chemotherapeutic option for the management of solid tumors
(1). However, cisplatin, even at
commonly used doses, may induce acute renal injury to varying
degrees, and this may affect the continuous administration of
chemotherapy and thus, the prognosis of patients. If left
untreated, acute kidney damage can lead to renal interstitial
fibrosis and, in severe cases, renal failure (2).
The renal tubular area is the key area affected by
acute kidney injury (AKI) caused by cisplatin that eventually
develops into interstitial fibrosis (3), and renal interstitial fibrosis is
considered to be a key factor underlying chronic renal failure
(4). However, the mechanism
underlying renal interstitial fibrosis following acute renal injury
induced by cisplatin has not been fully elucidated, and there is
still a lack of effective preventative and curative measures.
Inflammatory injury, oxidative stress injury and
apoptosis are the primary mechanisms of cisplatin-induced kidney
injury (5,6), and hypoxia is hypothesized to be a
common feature of all types of AKI (7). Cisplatin has been reported to induce
renal cell damage by upregulating the phosphorylation of p38MAPK
(8,9). Additionally, hypoxia-inducible factor
(HIF)-1α levels are significantly increased in rats with
cisplatin-induced acute kidney damage, which, in-turn protects
against renal injury (10,11). However, the roles of p38MAPK and
HIF-1α, and their association with apoptosis in renal interstitial
fibrosis following cisplatin-induced acute renal injury have not
been previously reported, to the best of our knowledge.
Ginaton injection is derived from ginkgo biloba
leaves (GBE) and has been used for several decades for the
treatment of cardiovascular and cerebrovascular diseases (12,13).
Previous reports have shown that GBE can prevent testicular injury
via its anti-apoptotic and anti-inflammatory effects (14), reducing
H2O2-induced cell cytotoxicity via
downregulation of p38 MAPK (15),
and can also inhibit the hepatic fibrosis and attenuate brain
death-induced renal injury by reducing the activity of p38MAPK
(16,17). Additionally, GBE can inhibit the
growth of transplanted solid tumors in mice, and dose-dependently
reduce the protein and mRNA expression levels of HIF-1α (18). Our previous study showed that GBE can
protect against cisplatin-induced acute renal injury, and thus the
subsequent renal interstitial fibrosis (19). However, whether GBE can reduce renal
interstitial fibrosis following cisplatin-induced acute renal
injury, and the mechanisms underlying its effects, remain to be
determined. In the present study, the effects of GBE were assessed
on renal interstitial fibrosis following cisplatin-induced acute
renal injury via detection of apoptosis, and based on the
expression of p38MAPK, TGF-β1 and HIF-1α during this process.
Materials and methods
Drugs and antibodies
GBE injection (batch number IB122) was purchased
from Dr Willmar Schwabe Pharmaceuticals. Each 17.5 mg ampule of GBE
consisted of 24% Ginkgo flavonol glycosides and 6% terpene lactones.
Cisplatin power injection (batch no. 5050272DB) was provided by
Qilu Pharmaceutical Co., Ltd. The anti-α-smooth muscle actin (SMA)
(cat. no. BM0002), anti-collagen 1 (Col I) (cat. no. BA0325),
anti-p38MAPK (cat. no. 9215S), anti phospho-(p-)p38MAPK (cat. no.
8690S), anti-TGF-β1 (cat. no. ab179695), anti-HIF-1α (cat. no
BS-3514) and anti-β-actin (cat. no. 3700S) antibodies were
purchased from Cell Signaling Technology, Inc. Antibodies against
Bcl-2 (cat. no. BM4985), caspase-3 (cat. no. BM4620) and Bax (cat.
no. BM3964) were obtained from Wuhan Boster Biological Technology,
Ltd.
Animals
Male Sprague-Dawley rats (body weight, 200±20 g)
were purchased from the Experimental Animal Center of Guangxi
Medical University, and were bred in an air-conditioned room where
the relative humidity was 60±10% at 25±2˚C, with a 12-h light/dark
cycle and ad libitum access to food and water. The present
study was approved by the Ethics Committee of Guangxi Medical
University (approval no. 201310009).
Experimental design
Rats were randomly divided into 5 groups following a
1-week acclimation period (n=9 per group). The rats were grouped as
follows: i) Control group, on day 1, rats received saline equal to
the volume of cisplatin, and equal to the volume of GBE from days
22 to 40; ii) CDDP group, rats received saline equivalent to the
volume of GBE, once a day from days 22 to 40; iii) CDDP + GBE
group, rats received GBE (3.17 mg/kg) once a day from days 22 to
40; iv) CDDP + SB203580 (SB; a specific p38 MAPK inhibitor) group,
rats received SB (1 mg/kg) once a week from days 22 to 40, and
saline equivalent to the volume of GBE once a day (on days where no
SB was administered); and v) CDDP + Enalapril group, rats received
enalapril (10 mg/kg) once a day from days 22 to 40.
Additionally, the rats in groups ii) to v) were
treated with a single dose of cisplatin (5 mg/kg) on day 1 to
induce AKI, which developed into renal interstitial fibrosis, as
described previously (10,11). Cisplatin, GBE and SB were
administered via intraperitoneal injection, whereas enalapril was
administered by gavage.
According to the drug dose conversion formulas of
different species of animals in China's pharmacological
experimental methodology (20), the
dose of EGB in this study was converted from the clinically
commonly used adult dose to the rat dose, and was also based on the
results of the preliminary experiments (data not shown). Enalapril
exhibits antioxidant effects and alleviates renal fibrosis
(21,22), and these effects are similar to those
of GBE (23,24), and is has been used as a positive
control drug in an anti-fibrotic study previously (25), thus enalapril was used as a positive
control in the present study as well.
Blood, urine and renal tissue
collection
On the 40th day, 12 h after administration of the
final dose, urine, blood and kidneys were collected and stored at
-80˚C. First, urine from each rat was gathered separately. Next,
the rats were anesthetized by intraperitoneal injection of sodium
pentobarbital (30 mg/kg, ip). When rats were under deep anesthesia
(the limbs and abdominal muscles became relaxed, breathing became
slow, and the corneal reflex was absent), blood samples were
collected from the abdominal aorta and centrifuged at 4˚C for 15
min at 1,409 x g. Finally, exsanguination was performed to
euthanize the animals, and when no reflexes and no breathing was
observed, the renal samples were obtained and washed using ice-cold
saline. A portion of these renal specimens were fixed at room
temperature using buffered formalin (10%) for 24 h, and then TUNEL
staining, Masson's trichrome staining, hematoxylin and eosin
(H&E) staining and immunohistochemistry were performed as soon
as possible. The remainder of the renal specimens were immediately
refrigerated at -80˚C for western blotting and reverse
transcription-quantitative (RT-q)PCR.
Determination of blood urea nitrogen
(BUN), serum creatinine (Scr) and urinary
N-acetyl-β-D-glucosaminidase (NAG) levels
The levels of NAG (cat no. A031) in the urine were
determined using the nitrophenol colorimetric method, and the
levels of BUN (cat. no. C013-2) and Scr (cat. no. C011-1) in the
peripheral blood were detected using a 7100 automatic biochemical
analyzer (Hitachi, Ltd.) using specific kits purchased from Nanjing
Jiancheng Bioengineering Research Institute. All procedures were
performed strictly in accordance with the manufacturer's protocol
(Nanjing Jiancheng Bioengineering Institute).
Hematoxylin and eosin (H&E)
staining
Kidney tissues were fixed (as described above) and
routinely embedded in paraffin. Next, the kidney tissues were
sectioned into 3-4 µm thick slices and then stained with H&E
according to the manufacturer's protocol (Lot no. 0904A18, Beijing
Leagene Biotech. Co., Ltd.). Slices were dewaxed with xylene,
hydrated using a decreasing gradient of ethanol solutions, and then
stained with hematoxylin staining solution for 15 min at room
temperature, differentiated using the differentiation solution for
30 sec, stained with the eosin staining solution for 2 min at room
temperature, and finally sealed with neutral gum after dehydration
using an increasing gradient of ethanol solutions, and clearing
using xylene. The stained slices were examined using a light
microscope (IX51; Olympus Corporation; magnification, x200). In
each sample evaluated, five randomly selected fields of view were
assessed and the average score of tubule interstitial injury was
calculated. The scoring criteria were based on a previous study
(26): 0, normal; 1, cortical damage
≤25%; 2, cortical damage 25-50%; 3, cortical damage 50-75%; and 4,
cortical damage >75%. Histopathological changes were scored by
an experienced pathologist who was blinded to the conditions of the
study.
Masson's trichrome staining
To evaluate renal interstitial fibrosis, the fixed
renal sections were stained using a Masson's trichrome staining kit
(Lot no. 0215A16, Beijing Leagene Biotech. Co., Ltd). Sections were
dewaxed and hydrated as above, then stained at room temperature
using a Weigert ferryhematoxylin staining solution for 8 min,
Lichunred fuchsin staining solution for 5 min, washed with
phosphomolybdate solution for 2 min, and finally, stained with
aniline blue staining solution for 2 min. Each renal section was
evaluated in five randomly selected non-overlapping visual fields
(magnification, x400). The relative area of interstitial fibrosis
was measured using the Colour Image Analyser function of Image-Pro
Plus version 6.0 (Media Cybernetics, Inc.). The areas overlaying
the tubular basement membrane and interstitial space were
evaluated, whereas the glomeruli and large vessels were not
included in the analysis.
Immunohistochemical detection of renal
α-SMA, Col I, TGF-β1 and HIF-1α protein expression
The protein expression levels of α-SMA, Col I,
TGF-β1 and HIF-1α in renal tissues were detected using
immunohistochemical staining. Briefly, paraffin-embedded renal
specimens were cut into slices. After dewaxing with xylene and
dehydrating with a gradient of increasing ethanol solutions (95%
ethanol followed by anhydrous ethanol 3 times, 5-10 sec each time),
the renal slices were incubated in hydrogen peroxide solution
(0.3%, 37˚C, 10 min) to quench the endogenous peroxidase activity,
followed by incubation at room temperature for 20 min with 10% goat
serum for blocking non-specific binding. Next, the corresponding
primary antibody was added and incubated with the renal sections
(4˚C, overnight). The primary antibodies used were: α-SMA (1:6,000;
cat. no. BM0002; Cell Signaling Technology, Inc.), Col I (1:1,400;
cat. no. BA0325; Cell Signaling Technology, Inc.), TGF-β1 (1:6,000;
cat. no. ab179695; Cell Signaling Technology, Inc.) and HIF-1α
(1:1,000; cat. no. BS-3514; Novus Biologicals, LLC). After
incubation at 37˚C for 1 h with the secondary antibody diluted in
PBS (1:100; cat. no. WP151228; Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd), the renal sections were treated with DAB
(Wuhan Boster Biological Technology, Ltd.) to develop the signal.
Finally, renal slices were visualized using Olympus Soft Imaging
Solutions. The sections were observed using a light microscope
(IX51; Olympus Corporation). The integrated optical density (IOD)
was used to quantify the positively stained area in Image Pro-Plus.
The average value of 5 randomly selected fields of view
(magnification, x400) for each renal sample was calculated. The
protein levels of α-SMA, Col I, TGF-β1 and HIF-1α in renal tissues
are expressed as the ratio of IOD to the positive staining
area.
TUNEL staining
The paraffin-embedded renal tissues were cut into
slices (4 µm). Next, the slices were stained using a TUNEL
Apoptosis Detection kit I, POD, according to the manufacturer's
protocol (Wuhan Boster Biological Technology, Ltd.). Finally, 5
non-overlapping fields of view of the tubules per renal section
were randomly selected using a light microscope (magnification,
x400), and the number of positive cells were counted.
TUNEL-positive cells showed brown staining in the nucleus,
indicative of apoptotic cells. The apoptotic rate (%) was
calculated by counting the proportion of positive cells in the
total number of cells using Image Pro-Plus.
Western blotting
Kidney tissues of rats were lysed into homogenate
with an appropriate amount of tissue Lysis Buffer and liquid
nitrogen, and the protein concentration of the lysates was measured
using a BCA protein assay kit. Next, equal amounts of protein
lysates (100 µg) were separated by 12% SDS-PAGE at 100 V for 2 h,
and transferred to PVDF membranes (EMD Millipore) at 4˚C (100 V,
1.5 h). After blocking using Quick Block™ Buffer (Beyotime
Institute of Biotechnology) for 15 min at room temperature, the
membranes were incubated overnight at 4˚C with the primary
antibodies. The primary antibodies included the following: Anti-Bax
(1:200; cat. no. BM3964; Wuhan Boster Biological Technology, Ltd.),
anti-Bcl-2 (1:200; cat. no. BM4985; Wuhan Boster Biological
Technology, Ltd.), anti-caspase-3 (1:200; cat. no. BM4620; Wuhan
Boster Biological Technology, Ltd.), anti-p38MAPK (1:500; cat. no.
9215S; Cell Signaling Technology, Inc.) and anti-p-p38MAPK
(1:1,000; cat. no. 8690S; Cell Signaling Technology, Inc.). β-actin
(1:10,000; cat. no. 3700S, Cell Signaling Technology, Inc.) was
used as the internal control. Finally, the membranes were incubated
for 1 h with secondary antibody (fluorescent goat anti-rabbit
antibody; 1:10,000; cat. no. 5151S; Cell Signaling Technology,
Inc.) and signals were visualized using a near-infrared bicolor
fluorescence imaging system (Odyssey CLx; Li-COR Biosciences).
Densitometry analysis was performed using ImageJ2x (National
Institutes of Health).
RT-qPCR
The samples of renal tissues stored at -80˚C were
grounded in homogenate with liquid nitrogen to extract RNA.
AxyPrep™ Multisource Total RNA Miniprep kit (Axygen; Corning, Inc.)
was used to extract total RNA, and then the gDNA Eraser (Takara
Bio, Inc.) and the Prime Script™ RT reagent kit were used to
reverse transcribe 1 µg total RNA, according to the manufacturer's
protocol. qPCR was used to detect the expression levels of target
genes using a SYBR® Premix Ex Taq™ II kit (Takara Bio,
Inc.) and an Applied Biosystems 7500 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR reaction mix
(10 µl) consisted of SYBR Premix Ex Taq II, forward primer (0.5
µl), reverse primer (0.5 µl), ROX Reference DyeII (50x), cDNA (2
µl) and RNase free dH2O (2 µl). The thermocycling
conditions were: Denaturation at 95˚C for 30 sec; followed by 40
cycles of 5 sec at 95˚C and 60˚C for 34 sec. The sequences of the
primers were: p38MAPK forward, 5'-TTACCGATGACCACGTTCAGTTTC-3' and
reverse, 5'-AGCGAGGTTGCTGGGCTTTA-3'; and GAPDH forward,
5'-GGCACAGTCAAGGCTGAGAATG-3' and reverse,
5'-ATGGTGGTGAAGACGCCAGTA-3'. The levels of p38MAPK gene were
normalized to that of GAPDH using the 2-ΔΔCq method:
ΔΔCq=(Cq control-Cq treatment) reference-(Cq
control-Cq treatment) target (27).
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS 24.0 (IBM Corp.) was used for statistical analysis. The
differences between multiple groups were assessed using a one-way
ANOVA with a post-hoc Tukey's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of GBE on kidney function in
rats treated with cisplatin
As shown in Fig. 1,
cisplatin induced damage to renal function and increased the levels
of BUN, Scr and urinary NAG significantly (P<0.05). GBE
treatment significantly decreased the cisplatin-induced increase in
the levels of BUN and Scr (P<0.05), and there was no significant
difference in the NAG levels (P>0.05). SB treatment
significantly lowered Scr levels (P<0.05), but had no
significant effect on BUN and NAG levels (P>0.05).
Enalapril treatment significantly reduced Scr levels, but increased
NAG levels (P<0.05). No significant differences were observed in
Scr, BUN and NAG levels between the GBE treatment and SB treatment
(P>0.05; Fig. 1B and
C).
Effect of GBE on renal tissue damage
induced by cisplatin
H&E staining showed that there were no
histopathological changes in the control rats. Cisplatin treatment
resulted in renal tubular lumen stenosis, and a portion of the
tubular epithelial cells exhibited signs of oedema, degeneration or
necrosis. Conversely, the GBE treatment alleviated renal injury
induced by cisplatin, as did the enalapril and SB treatments
(P<0.05; Fig. 2A).
The tubular injury score was significantly higher in
the CDDP group compared with the Control group (P<0.05). The
increased tubular injury score induced by cisplatin was
significantly reduced following GBE, SB or enalapril treatment
(P<0.05). Additionally, GBE treated rats exhibited significantly
lower tubule injury scores compared with the rats treated with SB
or enalapril (P<0.05; Fig.
2B).
Masson's trichrome staining showed there was no
apparent renal fibrogenesis in the control rats, whereas cisplatin
treatment resulted in severe renal interstitial fibrosis (Fig. 3). Cisplatin treatment resulted in a
higher relative area of renal interstitial fibrosis compared with
the control rats (P<0.05). The area of renal interstitial
fibrosis induced by cisplatin was significantly reduced following
treatment with GBE or SB (P<0.05), whereas enalapril had no
notable effect (P>0.05). Additionally, The rats treated with GBE
exhibited minimal areas of renal interstitial fibrosis compared
with the rats treated with SB or enalapril (P<0.05; Fig. 3).
Effect of GBE on the protein
expression levels of renal α-SMA and Col I in rats treated with
cisplatin
As shown in Fig. 4,
immunohistochemical staining showed that the expression of α-SMA
and Col I in renal tubular tissues was very low in the control
rats. Cisplatin increased the levels of α-SMA and Col I
significantly (P<0.05). Additionally, GBE, SB or Enalapril
treatment reduced the levels of α-SMA and Col I induced by
cisplatin (P<0.05). The levels of α-SMA and Col I were higher in
the SB treated rats compared with the GBE treated rats
(P<0.05).
Effect of GBE on renal apoptosis in
rats treated with cisplatin
TUNEL staining showed the proportion of apoptotic
cells in the control rats was very low, and cisplatin significantly
increased apoptosis (P<0.05). GBE and SB notably reduced the
increase in apoptosis induced by cisplatin (P<0.05), and the
effects of GBE were greater than that of SB (P<0.05; Fig. 5).
Effect of GBE on the protein
expression levels of apoptosis associated proteins
Western blotting analysis indicated that compared
with the control rats, Bax and cleaved caspase-3 protein expression
levels, as well as the cleaved caspase-3/caspase-3 ratio in rats
treated with cisplatin were all markedly elevated (P<0.05). GBE
and SB lowered the levels of Bax and cleaved caspase-3 levels, as
well as the cleaved caspase-3/caspase-3 ratio induced by cisplatin
significantly (P<0.05), and the effects of GBE were greater than
that of SB (P<0.05; Fig. 6).
Conversely, Bcl-2 levels in the rats treated with
cisplatin were reduced significantly compared with the control rats
(P<0.05). GBE and SB increased Bcl-2 protein expression levels
significantly (P<0.05; Fig.
6).
Effect of GBE on the protein
expression levels of renal p38MAPK, p-p38MAPK and the
phosphorylation ratio in rats treated with cisplatin
The expression levels of p38MAPK and p-p38MAPK were
detected using western blotting. The results showed that the
endogenously low expression of p-p38MAPK, as well as the low
p-p38MAPK/p38MAPK ratio in control rats was increased following
cisplatin treatment (P<0.05). Additionally, GBE and SB
significantly decreased the p-p38MAPK levels and the
p-p38MAPK/p38MAPK ratio compared with cisplatin treated rats
(P<0.05). The effects of GBE were greater than that of SB in
reducing p38MAPK levels (P<0.05; Fig.
7).
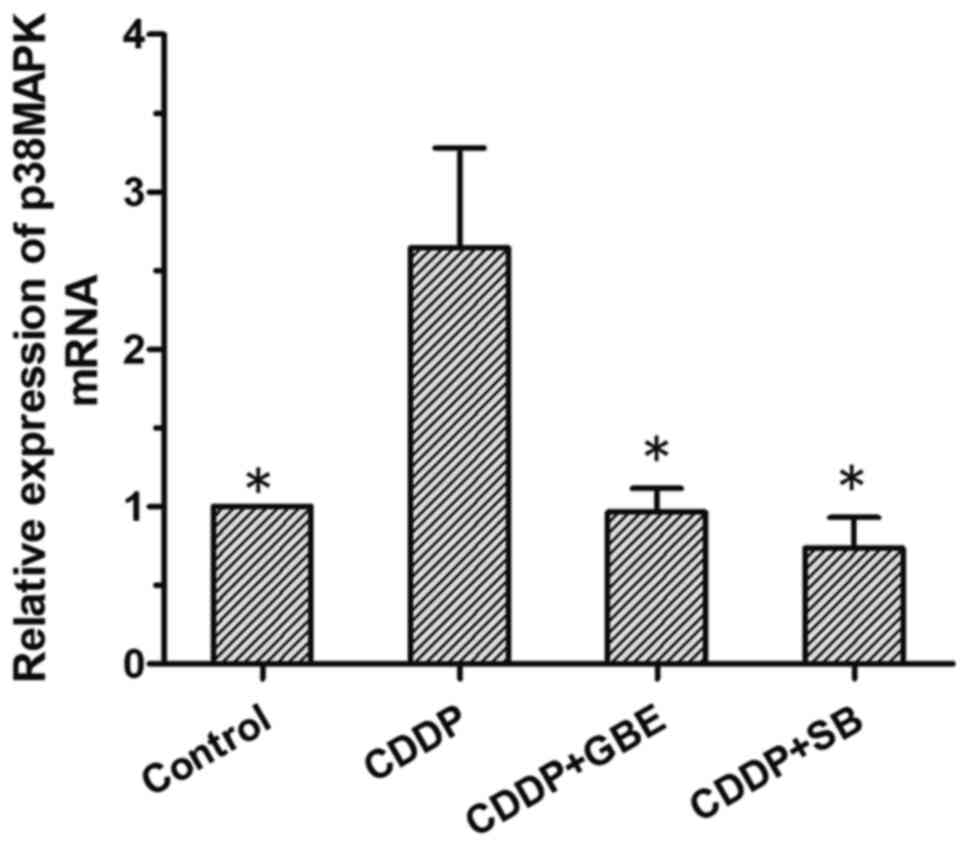
Effect of GBE on the levels of p38MAPK
mRNA in rats treated with cisplatin
The results of RT-qPCR analysis showed that the
p38MAPK mRNA expression levels were low in the control rats, and
that cisplatin treatment alone significantly increased its levels
(P<0.05). The levels of p38MAPK mRNA were significantly reduced
following GBE or SB treatment. (P<0.05; Fig. 8).
Effect of GBE on the protein
expression levels of renal TGF-β1 and HIF-1α in rats treated with
cisplatin
As shown in Fig. 9,
immunohistochemical analysis was used to measure TGF-β1 and HIF-1α
levels. It could be seen that TGF-β1 and HIF-1α protein expression
was concentrated in the epithelial cells of renal tubules. The
TGF-β1 and HIF-1α levels in the cisplatin treated rats were
significantly increased compared with the normal rats.
Additionally, GBE and SB all significantly reduced TGF-β1 levels,
as well as the HIF-1α levels induced by cisplatin (P<0.05).
Moreover, the effect of GBE on lowering HIF-1α levels was greater
than that of SB (P<0.05).
Discussion
Cisplatin is a frequently-used chemotherapeutic
option, and nephrotoxicity is a common adverse reaction (28). Our previous studies have shown that a
single dose of cisplatin (converted from a single common clinical
dose used in adults) can induce AKI in rats (10,11,29).
Without intervention, AKI may develop into renal interstitial
fibrosis (30,31). In the present study, the Scr, BUN and
NAG levels were significantly increased in rats treated with a
single dose of cisplatin. Additionally, the tubular injury score,
renal interstitial fibrosis, and protein expression levels of α-SMA
and Col I were all significantly increased, showing that cisplatin
resulted in renal injury and interstitial fibrosis in rats. These
results confirmed the successful establishment of a rat model of
renal interstitial fibrosis induced by cisplatin, and suggested
that cisplatin-induced renal interstitial fibrosis may be common in
clinical applications.
The mechanism by which cisplatin induces renal
interstitial fibrosis remains unclear, and there is a lack of
effective treatment measures. According to previous studies, cell
apoptosis and inflammatory damage are the primary mechanisms by
which cisplatin induces AKI (32-34).
Some traditional Chinese medicines, which possess anti-inflammatory
or anti-apoptotic effects have been shown to protect against kidney
injury induced by cisplatin (35,36). GBE
is extracted from ginkgo biloba leaves, which is a traditional
Chinese natural herb and has a long history of clinical use for
treatment of cardiovascular and cerebrovascular diseases (37). Previously, GBE has been reported to
ameliorate AKI in animal models induced by ischemia reperfusion
(38,39) or by cisplatin (19,40).
However, whether GBE can ameliorate renal interstitial fibrosis
following cisplatin-induced AKI has not been assessed. The results
of the present study showed that GBE significantly reduced the
deterioration in renal function induced by cisplatin, reversed the
increase in fibrosis related indicators, and reduced the degree of
renal interstitial injury and fibrosis in rats. These findings
suggest that GBE protected against renal function in rats and
improved renal interstitial fibrosis induced by cisplatin. However,
the molecular mechanisms underlying its anti-renal interstitial
fibrosis effects have not been determined.
Several studies have shown that renal interstitial
fibrosis is related to apoptosis of renal tubular epithelial cells
and tubular damage (41,42). Apoptosis of renal tubular epithelial
cells is a major mechanism underlying renal interstitial fibrosis
and cisplatin-induced AKI (9,43).
Cisplatin may induce apoptosis of renal tubular epithelial cells
both in vitro and in vivo (44). Previously, it has been shown GBE
inhibits hepatocyte apoptosis and improves liver fibrosis by
regulating the p38MAPK and Bcl-2/Bax pathways (45). In the present study, GBE
significantly decreased cisplatin-induced apoptosis in rat renal
tissues, decreased the levels of Bax and caspase-3, as well as the
cleaved caspase-3/caspase-3 ratio, and increased Bcl-2 protein
expression. These results suggest that GBE attenuated
cisplatin-induced renal interstitial fibrosis via inhibition of
apoptosis.
p38 MAPK is closely related to apoptosis and serves
a key role in this process (46).
The activation of p38 MAPK (p-p38 MAPK) mediated chemotherapeutic
drug-induced apoptosis (47), and
administration of a p38 MAPK inhibitor completely abolished TGF-β1
levels and inhibited fibrosis in human mesangial cells (48). The p38 MAPK/TGF-β1 pathway was shown
to mediate oxidative damage and promoted fibrosis of the liver
(49). Additionally, HIF-1α
inhibition attenuated hypoxia-induced apoptosis of renal tubular
cells (50), and elevated levels of
HIF-1α may aggravate tissue fibrosis (51). Our previous study found that HIF-1α
was elevated in rat kidneys, which had a protective effect on
cisplatin-induced AKI (11).
Importantly, GBE improved liver fibrosis by inhibiting the
apoptosis of hepatic stellate cells via the p38 MAPK pathway
(16), and prevented rats from
CCl4-induced liver fibrosis by inhibiting TGF-β1(52). GBE also regulated the expression of
HIF-1α mRNA and protein expression induced by hypoxia or ischemic
stroke (53,54). A previous study demonstrated that GBE
prevented renal fibrosis in rats with diabetic nephropathy
(24), which likely result3ed in
reduced formation of glomerular lesions. This suggests that GBE
improved tissue fibrosis via inhibition of apoptosis, by regulation
of p38 MAPK, TGF-β1 and HIF-1α. In the present study, p38 MAPK mRNA
expression levels, and the protein expression levels of p-p38 MAPK,
TGF-β1 and HIF-1α were significantly increased, and this was
accompanied by an increase in the rate of apoptosis and
apoptosis-related proteins, and decreased expression of Bcl-2 in
renal tissues exposed to cisplatin. Furthermore, GBE significantly
reversed the effects of cisplatin on these indicators of apoptosis,
and this was accompanied by a decrease in the apoptotic rate and in
the expression of apoptosis-related proteins. These findings
suggest that GBE improved cisplatin-induced renal interstitial
fibrosis via reduction of renal apoptosis and inhibition of p38
MAPK, TGF-β1 and HIF-1α function.
It is worth noting that the specific p38MAPK
inhibitor SB significantly reversed all cisplatin-induced effects
on indicators of renal function, renal tissue injury and fibrosis.
These results confirm that p38MAPK is involved in promoting renal
interstitial fibrosis in cisplatin-induced AKI. Additionally, SB
also significantly reversed the rate of renal tissue apoptosis and
the changes in the expression of apoptosis-associated proteins
induced by cisplatin, and these results confirm that p38MAPK
promoted renal interstitial fibrosis following cisplatin-induced
AKI by increasing renal tissue apoptosis. Moreover, SB also
significantly reversed cisplatin-induced changes in expression of
p38 MAPK, TGF-β1 and HIF-1α, and these results confirm that p38MAPK
promoted the expression of TGF-β1 and HIF-1α. The above results
indicate that GBE improved renal interstitial fibrosis following
cisplatin-induced AKI by inhibiting apoptosis via the p38
MAPK/TGF-β1 and p38 MAPK/HIF-1α pathways.
Although SB functions in a similar manner to GBE,
GBE was significantly more effective than SB in reducing p-P38
MAPK, TGF-β1 and HIF-1α protein expression levels, as well as renal
apoptosis, injury and fibrosis. This may be due to the fact that
GBE contains multiple active ingredients (55), and several of these may exhibit
anti-fibrotic effects via different pathways, including quercetin
and kaempferol (56,57). Kaempferoll exhibits an inhibitory
effect on TGF-β1-induced fibrosis related genes in renal tubular
epithelial cells, and improves renal function in rats (58). Therefore, in addition to p38MAPK, GBE
may inhibit TGF-β1 and HIF-1α through other pathways, thereby
inhibiting apoptosis and fibrosis, and this may explain the
improved effectiveness of GBE compared with SB, and highlights a
key advantage of Traditional Chinese Medicines.
In conclusion, the present study is the first to
show that GBE could effectively ameliorate renal interstitial
fibrosis following cisplatin-induced AKI by inhibiting renal
apoptosis, and this was mediated by downregulation of the
p38MAPK/TGF-β1 and p38MAPK/HIF-1α signaling pathways.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by funding from China's
National Natural Science Foundation (grant nos. 82060801, 81560729
and 81260598), the Natural Science Foundation of Guangxi (grant
nos. 2017GXNSFAA198262 and 2018GXNSFAA294043), and a self-funded
project from the Health Commission of Guangxi Zhuang Autonomous
Region (grant no. Z20200180).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
TL, CW and SL performed the experiments. MQ, GQ, YZ
and XZhong performed the statistical analysis. XZou assisted in the
design of the study. YY conceived and designed the study, and wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Guangxi Medical University (approval no. 201310009)
(Nanning, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Parr SK and Siew ED: Delayed consequences
of acute kidney injury. Adv Chronic Kidney Dis. 23:186–194.
2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yu CC, Chien CT and Chang TC: M2
macrophage polarization modulates epithelial-mesenchymal transition
in cisplatin-induced tubulointerstitial fibrosis. Biomedicine
(Taipei). 6(5)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Qi W, Chen X, Poronnik P and Pollock CA:
The renal cortical fibroblast in renal tubulointerstitial fibrosis.
Int J Biochem Cell Biol. 38:1–5. 2006.PubMed/NCBI View Article : Google Scholar
|
|
5
|
El-Naga RN: Pre-treatment with cardamonin
protects against cisplatin-induced nephrotoxicity in rats. Impact
on NOX-1, inflammation and apoptosis. Toxicol Appl Pharmacol.
274:87–95. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Qi ZL, Wang Z, Li W, Hou JG, Liu Y, Li XD,
Li HP and Wang YP: Nephroprotective effects of anthocyanin from the
fruits of Panax ginseng (GFA) on Cisplatin-induced acute kidney
injury in mice. Phytother Res. 31:1400–1409. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Nangaku M, Rosenberger C, Heyman SN and
Eckardt KU: Regulation of hypoxia-inducible factor in kidney
disease. Clin Exp Pharmacol Physiol. 40:148–157. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kim IH, Kwon MJ, Jung JH and Nam TJ:
Protein extracted from Porphyra yezoensis prevents
cisplatin-induced nephrotoxicity by downregulating the MAPK and
NF-κB pathways. Int J Mol Med. 41:511–520. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Thongnuanjan P and Soodvilai S,
Chatsudthipong V and Soodvilai S: Fenofibrate reduces
cisplatin-induced apoptosis of renal proximal tubular cells via
inhibition of JNK and p38 pathways. J Toxicol Sci. 41:339–349.
2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liang X, Yang Y, Huang Z, Zhou J, Li Y and
Zhong X: Panax notoginseng saponins mitigate cisplatin induced
nephrotoxicity by inducing mitophagy via HIF-1α. Oncotarget.
8:102989–103003. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Liu X, Huang Z, Zou X, Yang Y, Qiu Y and
Wen Y: Possible mechanism of PNS protection against
cisplatin-induced nephrotoxicity in rat models. Toxicol Mech
Methods. 25:347–354. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Li X, Lu L, Chen J, Zhang C, Chen H and
Huang H: New Insight into the mechanisms of ginkgo Biloba extract
in vascular aging prevention. Curr Vasc Pharmacol. 18:334–345.
2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
EGb 761. Ginkgo biloba extract, Ginkor.
Drugs R D. 4:188–193. 2003.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gevrek F, Biçer Ç, Kara M and Erdemir F:
The ameliorative effects of Ginkgo biloba on apoptosis, LH-R
expression and sperm morphology anomaly in testicular torsion and
detorsion. Andrologia: Feb 7, 2018 (Epub ahead of print).
|
|
15
|
Wang A, Yang Q, Li Q, Wang X, Hao S, Wang
J and Ren M: Ginkgo Biloba L. Extract reduces H2O2-induced bone
marrow mesenchymal stem cells cytotoxicity by regulating
mitogen-activated protein kinase (MAPK) signaling pathways and
oxidative stress. Med Sci Monit. 24:3159–3167. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wang R, Zhang H, Wang Y, Song F and Yuan
Y: Inhibitory effects of quercetin on the progression of liver
fibrosis through the regulation of NF-кB/IкBα, p38 MAPK, and
Bcl-2/Bax signaling. Int Immunopharmacol. 47:126–133.
2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li Y, Xiong Y, Zhang H, Li J, Wang D, Chen
W, Yuan X, Su Q, Li W, Huang H, et al: Ginkgo biloba extract GBE761
attenuates brain death-induced renal injury by inhibiting
pro-inflammatory cytokines and the SAPK and JAK-STAT signalings.
Sci Rep. 7(45192)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Cao CJ, Su Y, Sun J, Wang GY, Jia XQ, Chen
HS and Xu AH: Anti-tumor effect of ginkgo biloba exocarp extracts
on B16 melanoma bearing mice involving P I3K/Akt/HIF-1α/VEGF
signaling pathways. Iran J Pharm Res. 18:803–811. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yang YF, Lao S, Luo M and Zeng J: Dynamic
observation of the protective effect of ginkgo biloba extract on
cisplatin kidney damage in rabbits. Lishizhen Medicine and Materia
Medica. 22:2897–2898. 2011.
|
|
20
|
Wei W, Wu XM and Li YJ: Experimental
Methodology of Pharmacology. 4th edition. People's Medical
Publishing House, Beijing, pp1439-1442, 2010.
|
|
21
|
Asaad GF, Hassan A and Mostafa RE:
Anti-oxidant impact of Lisinopril and Enalapril against acute
kidney injury induced by doxorubicin in male Wistar rats:
Involvement of kidney injury molecule-1. Heliyon.
7(e05985)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sun N, Zhai L, Li H, Shi LH, Yao Z and
Zhang B: Angiotensin-converting enzyme inhibitor (ACEI)-mediated
amelioration in renal fibrosis involves suppression of mast cell
degranulation. Kidney Blood Press Res. 41:108–118. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ražná K, Sawinska Z, Ivanišová E, Vukovic
N, Terentjeva M, Stričík M, Kowalczewski PŁ, Hlavačková L, Rovná K,
Žiarovská J and Kačániová M: Properties of Ginkgo biloba L.:
Antioxidant characterization, antimicrobial activities, and genomic
MicroRNA based marker fingerprints. Int J Mol Sci.
21(3087)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lu Q, Zuo WZ, Ji XJ, Zhou YX, Liu YQ, Yao
XQ, Zhou XY, Liu YW, Zhang F and Yin XX: Ethanolic Ginkgo biloba
leaf extract prevents renal fibrosis through Akt/mTOR signaling in
diabetic nephropathy. Phytomedicine. 22:1071–1078. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yang H, Zhang W, Xie T, Wang X and Ning W:
Fluorofenidone inhibits apoptosis of renal tubular epithelial cells
in rats with renal interstitial fibrosis. Braz J Med Biol Res.
52(e8772)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li A, Zhang X, Shu M, Wu M, Wang J, Zhang
J, Wang R, Li P and Wang Y: Arctigenin suppresses renal
interstitial fibrosis in a rat model of obstructive nephropathy.
Phytomedicine. 30:28–41. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yuan JS, Reed A, Chen F and Stewart CN Jr:
Statistical analysis of real-time PCR data. BMC Bioinformatics.
7(85)2006.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yamamoto Y, Watanabe K, Tsukiyama I,
Matsushita H, Yabushita H, Matsuura K and Wakatsuki A:
Nephroprotective effects of hydration with magnesium in patients
with cervical cancer receiving cisplatin. Anticancer Res.
35:2199–2204. 2015.PubMed/NCBI
|
|
29
|
Li Q, Liang X, Yang Y, Zeng X, Zhong X and
Huang C: Panax notoginseng saponins ameliorate cisplatin-induced
mitochondrial injury via the HIF-1α/mitochondria/ROS pathway. FEBS
Open Bio. 10:118–126. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lu S, Zhong X, Yang Y, Zou X, Liang X and
Cai G: Effects of single dose of cisplatin on renal interstitial
fibrosis indicators in rats. China Pharmacy. 29:298–302. 2018.
|
|
31
|
Johnson FL, Patel NSA, Purvis GSD, Chiazza
F, Chen J, Sordi R, Hache G, Merezhko VV, Collino M, Yaqoob MM and
Thiemermann C: Inhibition of IκB kinase at 24 hours after acute
kidney injury improves recovery of renal function and attenuates
fibrosis. J Am Heart Assoc. 6(e005092)2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ozkok A and Edelstein CL: Pathophysiology
of cisplatin-induced acute kidney injury. Biomed Res Int.
2014(967826)2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Pabla N and Dong Z: Cisplatin
nephrotoxicity: Mechanisms and renoprotective strategies. Kidney
Int. 73:994–1007. 2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Pabla N and Dong Z: Curtailing side
effects in chemotherapy: A tale of PKCδ in cisplatin treatment.
Oncotarget. 3:107–111. 2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Huang SJ, Huang J, Yan YB, Qiu J, Tan RQ,
Liu Y, Tian Q, Guan L, Niu SS, Zhang Y, et al: The renoprotective
effect of curcumin against cisplatin-induced acute kidney injury in
mice: Involvement of miR-181a/PTEN axis. Ren Fail. 42:350–357.
2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Lee IC, Ko JW, Park SH, Shin NR, Shin IS,
Kim YB and Kim JC: Ameliorative effects of pine bark extract on
cisplatin-induced acute kidney injury in rats. Ren Fail.
39:363–371. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Tian J, Liu Y and Chen K: Ginkgo biloba
extract in vascular protection: Molecular mechanisms and clinical
applications. Curr Vasc Pharmacol. 15:532–548. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wang Y, Pei DS, Ji HX and Xing SH:
Protective effect of a standardized Ginkgo extract (ginaton) on
renal ischemia/reperfusion injury via suppressing the activation of
JNK signal pathway. Phytomedicine. 15:923–931. 2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sener G, Sener E, Sehirli O, Oğünç AV,
Cetinel S, Gedik N and Sakarcan A: Ginkgo biloba extract
ameliorates ischemia reperfusion-induced renal injury in rats.
Pharmacol Res. 52:216–222. 2005.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Gulec M, Iraz M, Yilmaz HR, Ozyurt H and
Temel I: The effects of ginkgo biloba extract on tissue adenosine
deaminase, xanthine oxidase, myeloperoxidase, malondialdehyde, and
nitric oxide in cisplatin-induced nephrotoxicity. Toxicol Ind
Health. 22:125–130. 2006.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Younis NN, Elsherbiny NM, Shaheen MA and
Elseweidy MM: Modulation of NADPH oxidase and Nrf2/HO-1 pathway by
vanillin in cisplatin-induced nephrotoxicity in rats. J Pharm
Pharmacol. 72:1546–1555. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Jing Z, Hu L, Su Y, Ying G, Ma C and Wei
J: Potential signaling pathway through which Notch regulates
oxidative damage and apoptosis in renal tubular epithelial cells
induced by high glucose. J Recept Signal Transduct Res: Sep 16,
2020 (Epub ahead of print).
|
|
43
|
Zhang XF, Yang Y, Zhang J and Cao W:
Microvesicle-containing miRNA-153-3p induces the apoptosis of
proximal tubular epithelial cells and participates in renal
interstitial fibrosis. Eur Rev Med Pharmacol Sci. 23:10065–10071.
2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Du B, Dai XM, Li S, Qi GL, Cao GX, Zhong
Y, Yin PD and Yang XS: MiR-30c regulates cisplatin-induced
apoptosis of renal tubular epithelial cells by targeting Bnip3L and
Hspa5. Cell Death Dis. 8(e2987)2017.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wang Y, Wang R, Wang Y, Peng R, Wu Y and
Yuan Y: Ginkgo biloba extract mitigates liver fibrosis and
apoptosis by regulating p38 MAPK, NF-κB/IκBα, and Bcl-2/Bax
signaling. Drug Des Devel Ther. 9:6303–6317. 2015.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417.
2010.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Deacon K, Mistry P, Chernoff J, Blank JL
and Patel R: p38 Mitogen-activated protein kinase mediates cell
death and p21-activated kinase mediates cell survival during
chemotherapeutic drug-induced mitotic arrest. Mol Biol Cell.
14:2071–2087. 2003.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Liu Z, Xue L, Liu Z, Huang J, Wen J, Hu J,
Bo L and Yang R: Tumor necrosis factor-like weak inducer of
apoptosis accelerates the progression of renal fibrosis in lupus
nephritis by activating SMAD and p38 MAPK in TGF-β1 signaling
pathway. Mediators Inflamm. 2016(8986451)2016.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Li ZL, Shi Y, Le G, Ding Y and Zhao Q:
24-week exposure to oxidized tyrosine induces hepatic fibrosis
involving activation of the MAPK/TGF-β1 signaling pathway in
Sprague-Dawley Rats model. Oxid Med Cell Longev.
2016(3123294)2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Liu LL, Li D, He YL, Zhou YZ, Gong SH, Wu
LY, Zhao YQ, Huang X, Zhao T, Xu L, et al: MiR-210 protects renal
cell against hypoxia-induced apoptosis by targeting HIF-1alpha. Mol
Med. 23:258–271. 2017.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Liu L, Zhang P, Bai M, He L, Zhang L, Liu
T, Yang Z, Duan M, Liu M, Liu B, et al: p53 upregulated by HIF-1α
promotes hypoxia-induced G2/M arrest and renal fibrosis in vitro
and in vivo. J Mol Cell Biol. 11:371–382. 2019.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Liu SQ, Yu JP, Chen HL, Luo HS, Chen SM
and Yu HG: Therapeutic effects and molecular mechanisms of ginkgo
biloba extract on liver fibrosis in rats. Am J Chin Med. 34:99–114.
2006.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Oh JH, Oh J, Togloom A, Kim SW and Huh K:
Effects of ginkgo biloba extract on cultured human retinal pigment
epithelial cells under chemical hypoxia. Curr Eye Res.
38:1072–1082. 2013.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Chen M, Zou W, Chen M, Cao L, Ding J, Xiao
W and Hu G: Ginkgolide K promotes angiogenesis in a middle cerebral
artery occlusion mouse model via activating JAK2/STAT3 pathway. Eur
J Pharmacol. 833:221–229. 2018.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Singh B, Kaur P, Gopichand Singh RD and
Ahuja PS: Biology and chemistry of Ginkgo biloba. Fitoterapia.
79:401–418. 2008.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wang T, Xiao J, Hou H, Li P, Yuan Z, Xu H,
Liu R, Li Q and Bi K: Development of an ultra-fast liquid
chromatography-tandem mass spectrometry method for simultaneous
determination of seven flavonoids in rat plasma: Application to a
comparative pharmacokinetic investigation of Ginkgo biloba extract
and single pure ginkgo flavonoids after oral administration. J
Chromatogr B Analyt Technol Biomed Life Sci. 1060:173–181.
2017.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Ji X, Cao J, Zhang L, Zhang Z, Shuai W and
Yin W: Kaempferol protects renal fibrosis through activating the
BMP-7-Smad1/5 signaling pathway. Biol Pharm Bull. 43:533–539.
2020.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Liu X, Sun N, Mo N, Lu S, Song E, Ren C
and Li Z: Quercetin inhibits kidney fifibrosis and the epithelial
to mesenchymal transition of the renal tubular system involving
suppression of the Sonic Hedgehog signaling pathway. Food Funct.
10:3782–3797. 2019.PubMed/NCBI View Article : Google Scholar
|