Introduction
Human choriocarcinomas, which are a rare form of
cancer that develops in the uterus from tissue that would normally
become the placenta, are a trophoblastic gestational disease. These
carcinomas have been studied largely to investigate conditions
associated with pregnancy, such as preeclampsia (1). However, the mechanism underlying
choriocarcinoma development remains to be elucidated.
microRNAs (miRNAs) are small RNA molecules (21–23
nt) that act as negative regulators of gene expression, either by
blocking mRNA translation into protein or by RNA interference
(2,3). Previous studies have revealed that
dysregulation of specific miRNAs is associated with certain types
of cancer and are considered to act either as oncogenes or tumor
suppressors, depending on the target gene (2,4–10).
Examples include, miR-15a, which has been associated with chronic
lymphocytic leukemia (2,5,6), and
also miR-21 and miR-17, which are upregulated, while miR-143 and
miR-145 are down-regulated in colorectal cancer (11,12).
The presence of the let-7 family of miRNAs is a prognostic factor
in lung cancer (5,6) and miR-182 inhibits the proliferation
and invasion ability of the human lung adenocarcinoma cells via its
effect on human cortical actin-associated protein (13). To date, investigation of the
decreased expression of miR-199b in human choriocarcinoma by Chao
et al indicated that epigenetic mechanisms play an important
role in increasing the expression levels of protein phosphatase 2A
inhibitor and contribute to the pathogenesis of human
choriocarcinoma (14). However, no
studies have linked miR-145 expression with the proliferation and
invasion capacity of human choriocarcinoma cells (2).
The transcription activator Sox2 was originally
studied in the context of sexual determination during the
development of Drosophila embryos and thus, its name is an
acronym for ‘sex determination Y-box2’ (15–17).
Numerous studies have indicated a primary role for Sox2 factor in
the maintenance of embryonic stem cell pluripotency, and in later
stages of development, in the repression of trophectoderm and
epiblast genes. In addition, Sox2 appears to have a significant
role in the differentiation of the nervous system (16). Extensive studies have indicated
that Oct4, Sox2 and Nanog are required for self-renewal and
pluripotency of embryonic stem cells (17,18).
Investigation of the expression and methylation profiles of Sox2 in
placentas and gestational trophoblastic disease by Li et al
indicated that epigenetic mechanisms play an important role in the
transcriptional regulation of Sox2 and contribute to the
pathogenesis of gestational trophoblastic disease (19). By contrast, Xu et al
reported that endogenous miR-145 represses the 3′-untranslated
regions (3′-UTRs) of Oct4, Sox2 and Klf4, and that increased
miR-145 expression inhibits human embryonic stem cell self-renewal,
represses expression of pluripotency genes and induces
lineage-restricted differentiation (18). In addition, Sox2 was closely
associated with certain tumors, its inappropriate activation being
involved in the development processes of human tumors, including
the abnormal methylation modification of the promoter region of the
Sox2 gene. Nakatsugawa et al(20) analyzed the functions of Sox2 in
cancer stem-like cells/cancer-initiating cells derived from human
lung adenocarcinoma. Nakatsugawa et al revealed that the
Sox2 protein was detected in >80% of cancer stem-like
cells/cancer-initiating cells in primary lung carcinoma tissues.
However, Sox2 mRNA knockdown of the human lung cancer stem-like
cells/cancer-initiating cells by gene-specific siRNA eliminated
tumorigenicity in vitro and in vivo. These
observations indicate that Sox2 has a role in the maintenance of
stemness and tumorigenicity of human lung carcinoma and is a
potential target for treatment.
In view of this evidence, in the current study, the
miR-145 expression vector was transfected into the human
choriocarcinoma cell lines JAR and JEG-3 to determine its specific
role in Sox2 regulation and inhibition of cell proliferation, and
invasion. These results are of potential importance for the
identification of therapeutic targets in human choriocarcinoma.
Materials and methods
Cell lines and animals
The human choriocarcinoma cell lines JAR and JEG-3
were grown in DMEM (Hyclone, Logan, UT, USA) supplemented with 10%
fetal bovine serum (PAA Laboratories Inc., Queensland, Australia),
penicillin (100 U/ml), streptomycin (100 U/ml) and 2 mM L-glutamine
(all were purchased from Hyclone). The JAR cells were incubated at
37°C in a humidified atmosphere of air containing 5%
CO2. All experiments with BALB/c nude mice of 6–7 weeks
of age and 20–22 g of weight were carried out at the Laboratory
Animal Center of Tongji University with Institutional Animal Care
and Use Committee approval in accordance with institutional
guidelines.
Recombinant lentivirus generation and
vector construction
The method for creating the recombinant lentivirus
package was as previously described (18). An RNAi pLL3.7 (LentiLox 3.7)
lentiviral system was used to create lentiviral vectors (Clontech,
Beijing, China). For vector pLL3.7-mir145 (pre-miRNA of miR-145
expression element), oligonucleotide pairs for pre-miRNA of miR-145
and linker sequences with HpaI and XhoI sites were
chemically synthesized. The sequences of the oligonucleotides were:
top strand, 5′-CGg tta acC ACC TTG TCC TCA CGG TCC AGT
TTT CCC AGG AAT CCC TTA GAT GCT AAG ATG GGG ATT CCT GGA AAT ACT
GTT CTT GAG GTC ATG GTT ctc gag CG-3′; and bottom strand,
5′-CGc tcg agA ACC ATG ACC TCA AGA ACA GTA TTT CCA GGA ATC
CCC ATC TTA GCA TCT AAG GGA TTC CTG GGA AAA CTG GAC CGT GAG GAC AAG
GTG gtt aac CG-3′ (sequences corresponding to miR-145 seed
sequences are capitalized and bold, and restriction enzyme sites
are lower case and bold) (18). To
build the expression plasmid, the pairs of oligos were annealed and
inserted into the multiple cloning sites between the HpaI
and XhoI sites in the pLL3.7 vector. The negative control
plasmid pLL3.7-mir145-Mut was similarly constructed, with the
exception that 23 nucleotides in sequences corresponding to miR-145
were mutated (GTC CAG TTT TCC CAG GAA TCC CT to Gaa Ct Gaa TTa gCA
cGA AgC aCT, mutations shown in lower-case). The pLL3.7-mir145 or
pLL3.7-mir145-Mut was recombined in the package cell line 293T to
create lentiviruses. Recombinant viruses were propagated in 293T
cells, purified and titered by standard methods, as previously
described by our laboratory (21).
The corresponding viruses were designated Ldv-mir145 and
Ldv-mir145-Mut. Co-transfection of human iPS cells used
4×107 PFU/ml Ldv-mir145 or Ldv-mir145-Mut lentivirus
according to the manufacturer’s instructions. The cells were seeded
in a six-well plate in DMEM (Sigma-Aldrich, St. Louis, MO, USA)
supplemented with 10% fetal bovine serum, 100 U/ml penicillin and
100 μg/ml streptomycin at 37°C in a humidified atmosphere of
air containing 5% CO2, until cells were 80%
confluent.
RNA extraction and analysis by
quantitative real-time PCR (qRT-PCR)
Total RNA from each cell was isolated using TRIzol
reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer’s instructions. The RNA samples were treated with
DNase I (Sigma-Aldrich), quantified and reverse-transcribed into
cDNA using the ReverTra Ace-α First Strand cDNA Synthesis kit
(Toyobo, Osaka, Japan). qRT-PCR was conducted using a RealPlex4
real-time PCR detection system from Eppendorf Co., Ltd., (Hamburg,
Germany), with SYBR Green RealTime PCR Master mix (Toyobo) used as
the detection dye. qRT-PCR amplification was performed over 40
cycles with denaturation at 95°C for 15 sec and annealing at 58°C
for 45 sec. Target cDNA was quantified using the relative
quantification method. A comparative threshold cycle (Ct) was used
to determine gene expression relative to a control (calibrator) and
steady-state mRNA levels were reported as an n-fold difference
relative to the calibrator. For each sample, the marker genes Ct
values were normalized using the formula ΔCt=Ct_genes-Ct_18S rRNA.
To determine relative expression levels, the following formula was
used: ΔΔCt= ΔCt_all_groups-ΔCt_blankcontrol_group. The values used
to plot relative expressions of markers were calculated using the
expression 2−ΔΔCt. The mRNA levels were calibrated based
on levels of 18S rRNA. The cDNA of each gene was amplified using
primers as previously described (19).
Methyl-thiazolyl-tetrazolium (MTT) assay
for cell proliferation
Each group of JAR and JEG-3 cell lines was seeded at
2×103 per well in 96-well plates and cultured in DMEM
supplemented with 10% FBS at 37°C with 5% CO2, until
cells were 85% confluent. The MTT (Sigma Chemicals, St. Louis, MO,
USA) reagent (5 mg/ml) was added to the maintenance cell medium at
various time points and incubated at 37°C for an additional 4 h.
The reaction was terminated with 150 μl dimethylsulfoxide
(DMSO, Sigma Chemicals) per well and the cells were lysed for 15
min, and the plates were agitated every 5 min. Absorbance values
were determined using the enzyme linked immunosorbent assay (ELISA)
reader (Model 680; Bio-Rad, Hercules, CA, USA) at 490 nm.
Flow cytometric (FCM) analysis of cell
cycle by propidium iodide (PI) staining
Each group of JAR and JEG-3 cell lines was seeded at
3×105 per well in 6-well plates and cultured until 85%
confluent. Each group of cells was washed with PBS three times,
then collected by centrifuging (Allegra X-22R; Beckman Coulter,
Miami, FL, USA) at 1000 × g for 5 min. The cell pellets were
resuspended in 1 ml PBS, fixed in 70% ice-cold ethanol and kept in
a freezer for >48 h. Prior to FCM analysis, the fixed cells were
centrifuged, washed twice with PBS and resuspended in PI staining
solution (Sigma Chemicals) containing 50 μl/ml PI and 250
μg/ml RNase A (Sigma Chemicals). The cell suspension, which
was kept in the dark, was incubated for 30 min at 4°C and analyzed
by FACS (FCM-500, Beckman Coulter). A total of 20,000 events were
acquired for analysis using CellQuest software.
Luciferase report assay
All steps of the luciferase reporter assay were as
previously described (18,22,23).
NIH-3T3 cells were seeded at 3×104 per well in 48-well
plates and co-transfected with 400 ng pLL3.7-mir145, pLL3.7 or
pLL3.7-mir145-Mut, 20 ng pGL3cm-Sox2-3UTR-WT or
pGL3cm-Sox2-3UTR-Mut, and pRL-TK (Promega, Madison, WI, USA) using
Lipofectamine 2000 reagent according to the manufacturer’s
instructions. Luciferase activity was measured 48 h after
transfection using the dual-luciferase reporter assay system
(Promega).
RNA extraction and northern blot
analysis
Northern blotting was performed as previously
described (13,24). For all groups, 20 μg good
quality total RNA was analyzed on a 7.5 M urea 12% PAA denaturing
gel and transferred to a Hybond N+ nylon membrane
(Amersham, Freiburg, Germany). Membranes were crosslinked using UV
light for 30 sec at 1,200 mJ/cm2. Hybridization was
performed with the miR-145 antisense starfire probe, 5′-AGG GAT TCC
TGG GAA AAC TGG AC-3′ (IDT, Coralville, IA, USA), to detect the
22-nt miR-199a fragments according to the manufacturer’s
instructions. After washing, membranes were exposed for 20–40 h to
Kodak XAR-5 films (Sigma-Aldrich). As a positive control, all
membranes were hybridized with a human U6 snRNA probe, 5′-GCA GGG
GCC ATG CTA ATC TTC TCT GTA TCG-3′. Exposure times for the U6
control probe varied between 15 and 30 min.
Western blot analysis
Total protein extracts from each group of cells were
resolved by 12% SDS-PAGE and transferred on PVDF (Millipore,
Billerica, MA, USA) membranes. After blocking, the PVDF membranes
were washed 4 times for 15 min with TBST at room temperature and
incubated with the primary antibody [rabbit anti-human Sox2
polyclonal antibody (1:200; Chemicon, Temecula, CA, USA)]. After
extensive washing, membranes were incubated with secondary
peroxidase-linked goat anti-rabbit IgG (1:1000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) for 1 h. After washing 4
times for 15 min with TBST at room temperature, the
immunoreactivity was visualized by enhanced chemiluminescence (ECL
kit; Pierce Biotechnology, Inc., Rockford, IL, USA) and the
membranes were exposed to Kodak XAR-5 films.
Soft agar colony formation assay
The method used was as previously described
(25). Soft agar assays were
constructed in 6-well plates. The base layer of each well consisted
of 2 ml with final concentrations of 1X medium (DMEM+10% FBS) and
0.6% low melting point agarose. The plates were chilled at 4°C
until solid. Subsequently, a 1.0-ml agar growth layer, consisting
of 1×104 cells suspended in 1X media and 0.3% low
melting point agarose, was poured onto the base layer. The plates
were again chilled at 4°C until the growth layer congealed.
Additional 1X media without agarose (1.0 ml) was added to the top
of the growth layer on day 0 and again on day 15 of growth. The
cells were allowed to grow at 37°C for 1 month and total colonies
counted. The assays were repeated a total of 3 times. Results were
statistically analyzed by paired t-test using the PRISM Graphpad
program (Graphpad Software, La Jolla, CA, USA).
Transwell migration assay
All steps were as previously described (26). The cells (2×105) were
resuspended in 200 μl serum-free medium and seeded on the
top chamber of the 6.5 mm polycarbonate transwell filters (8.0
μm pores; Corning Inc., Corning, NY, USA). The full medium
(600 μl) containing 10% FBS was added to the bottom chamber.
The cells were allowed to migrate for 24 h at 37°C in a humidified
incubator with 5% CO2. The cells attached to the lower
surface of the membrane were fixed in 4% paraformaldehyde at room
temperature for 30 min and stained with
4,6-diamidino-2-phenylindole (DAPI; C1002; Beyotime Institute of
Biotechnology, Jiangsu, China), and the number of cells on the
lower surface of the filters was counted under the microscope. A
total of 5 fields were counted for each transwell filter.
In vivo xenograft experiments
Logarithmically growing ovarian cancer-initiating
cells (∼1×105) were inoculated into BALB/c nude/nude
mice. Each experimental group consisted of four mice. After 4 weeks
of observation, the mice were sacrificed and tumors were obtained
(27). The tumor weight was
measured and tumor volume was calculated according to the formula:
tumor volume (mm3) = length (mm) × width (mm) × height
(mm).
Statistical analysis
Each experiment was performed at least three times
and data were expressed as the mean±SE. The differences were
evaluated using Student’s t-tests. P<0.05 was considered to
indicate a statistically significant result.
Results
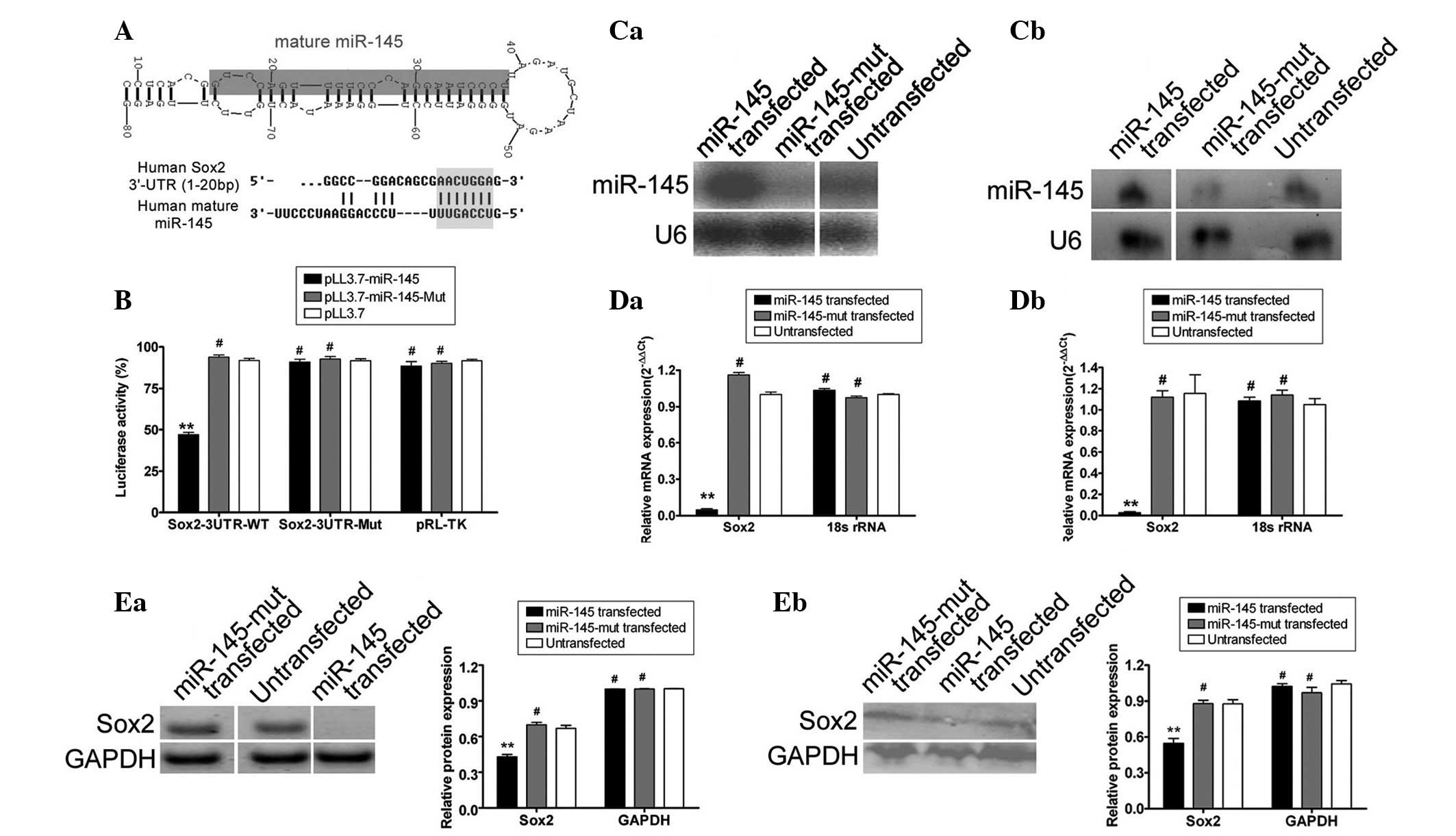
miR-145 binding with the 3′-UTR sites in
Sox2
Using an online research tool, the miRBase Target
database (http://www.mirbase.org) (28,29),
the precursor miRNA (pre-miRNA) sequences, mature miRNA sequences,
chromosomal locations and length of miR-145 and the target gene
Sox2 were analyzed. Seven putative miRNA target sites were
identified in the 3′-UTR of Sox2 mRNA, depending on species.
This study focused on human miR-145, which targets the human
Sox2 3′-UTR, although conservation in this sequence
indicates the possibility of binding to varying degrees, across
species (Fig. 1). Plasmid DNA
encoding each Sox2 mRNA 3′-UTR site [wild-type (wt)
Sox2, empty plasmid and mutant Sox2] was
co-transfected with the miR-145 expression lentivirus (wt miR-145,
empty lentivirus and mutant miR-145 lentivirus) into the mouse
embryonic fibroblast cell line NIH-3T3, to examine regulation of
Sox2 gene expression by mature miR-145. The luciferase
activity of the Sox2 3′-UTR sites was significantly
inhibited by wt miR-145 (Fig. 1),
while the luciferase activity of the mutated Sox2 3′-UTR
sites was not inhibited, suggesting that Sox2 was targeted
by miR-145.
miR-145 specifically influences
expression of Sox2 protein in human choriocarcinoma cell lines
Northern blot analysis demonstrated that the
hybridized signal of mutant miR-145 in the JAR and JEG-3
choriocarcinoma cell lines was weaker than in cells transfected
with wt miR-145. qRT-PCR and western blot analyses were used to
determine the effect of exogenous and endogenous miR-145 expression
on Sox2 expression. qRT-PCR analyses revealed decreased Sox2
mRNA expression in wt miR-145 lentivirus-transfected JAR and JEG-3
cells than in untransfected and mutant miR-145-transfected cells.
The relative mRNA expression after normalization to 18S ribosomal
RNA (rRNA), which served as an internal control, is shown in
Fig. 1. Notably, western blotting
revealed that Sox2 levels in untransfected cells (JAR or JEG-3 cell
lines) and mutant miR-145 transfected cells (JAR or JEG-3 cell
lines) were 0.667±0.026 or 0.876±0.036, and 0.669±0.020 or
0.879±0.028 relative to those of GAPDH, respectively (Fig. 1). These values were significantly
higher than those for the wt miR-145 transfected group (JAR:
0.429±0.019; JEG-3: 0.547±0.040 relative to GAPDH), which indicated
that exogenous miR-145 down-regulated Sox2 expression. Therefore,
miR-145 expression may influence endogenous Sox2 expression.
Proliferation and invasion of human
choriocarcinoma cell lines were inhibited by miR-145
The results of the proliferation assays performed
are shown in Fig. 2. Using an MTT
assay, the survival rate of wt miR-145 lentivirus-transfected cells
was demonstrated to be markedly lower than that of untransfected
cells and mutant miR-145-transfected cells at both 3 and 5 days
post-transfection. By contrast, no differences in viability were
observed in untransfected cells, mutant miR-145-transfected cells
and wt miR-145 transfected cells 1 and 2 days post-transfection.
The survival rates remained unchanged for untransfected cells and
mutant miR-145 transfected cells for the remainder of the time
course, which indicated that induced exogenous miR-145 expression
inhibited the growth of human choriocarcinoma JAR and JEG-3 cell
lines in vitro. In addition, migration and invasion ability
were shown to be reduced in JAR and JEG-3 cells with stably
repressed Sox2 mediated by miR-145 transfection using transwell
migration analysis and soft agar colony formation assays,
respectively (Fig. 2). Transwell
migration invasion assays showed that the number of invading wt
miR-145-transfected JAR cells was significantly lower than the
numbers of invading untransfected and mutant miR-145-transfected
JAR cells (invading cell numbers: miR-145 transfected group, 13±2;
untransfected cells, 24±2; mutant miR-145 transfected cells, 26±2).
The results of transwell migration invasion assays in JEG-3 cells
were similar to those of JAR cells. These results indicate that the
repression of Sox2 expression by miR-145 significantly attenuates
the invasion and migration ability of human choriocarcinoma cells.
Soft agar colony formation assays consistently indicated that
miR-145-transfected cells formed substantially fewer colonies
compared with controls or mutant miR-145-transfected cells when
plated at low density (Fig. 2). In
addition, miR-145 transfected, mutant miR-145-transfected and
untransfected JAR or JEG-3 cells were stained with PI, and analyzed
by flow cytometry to detect changes in cell cycle progression. As
shown in Fig. 3, the majority of
the wt miR-145-transfected JAR cells were arrested in the
G0/G1 phase of the cell cycle and the
percentage of cells in the S phase were markedly decreased. By
contrast, no significant differences were observed in the cell
cycle distribution of the mutant miR-145-transfected and
untransfected JAR cells. In addition, the majority of wt
miR-145-transfected JEG-3 cells were arrested in the
G0/G1 phase of the cell cycle and the
percentage of cells in the G2/M phase were markedly
decreased. However, no significant differences were observed in the
cell cycle distribution of the mutant miR-145-transfected and
untransfected cells. The results suggested that wt miR-145
expression affected cell cycle regulation in human choriocarcinoma
cells in vitro.
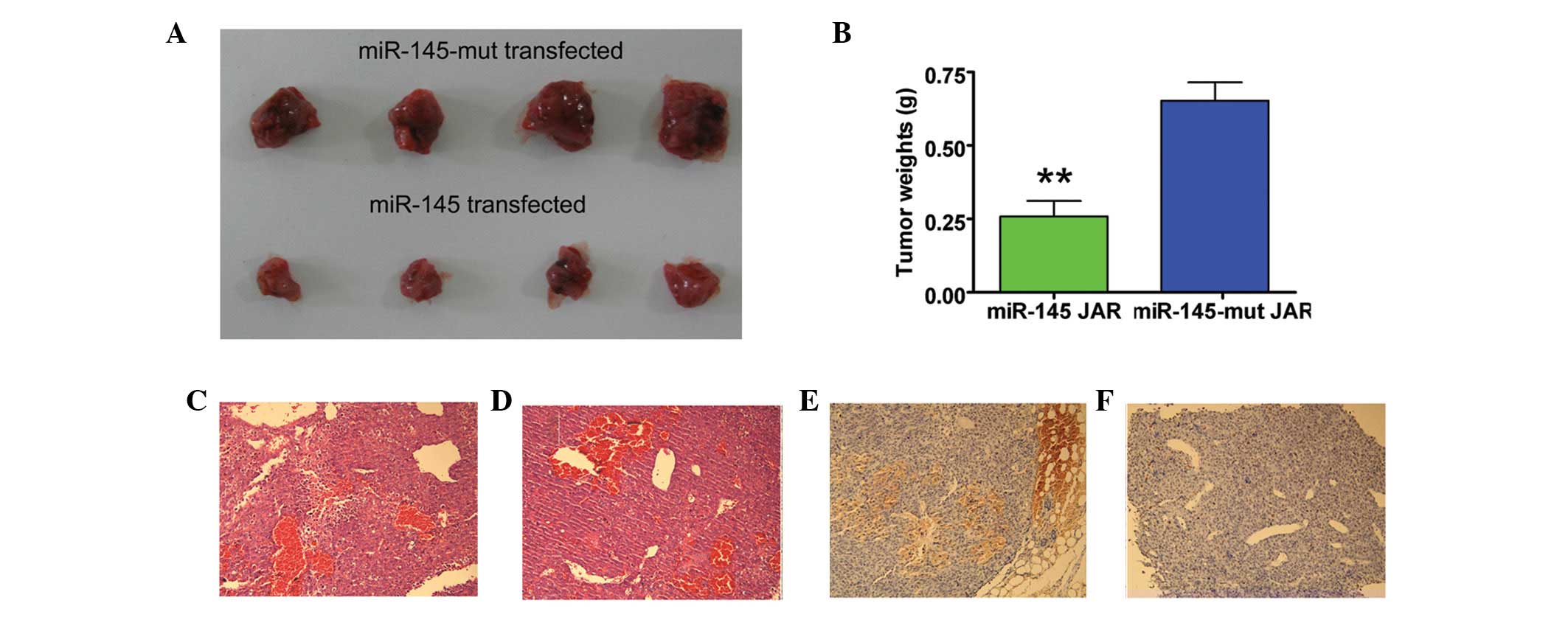
Expression of wt miR-145 in JAR cells
inhibited subcutaneous tumor growth in nude mice
The effect of miR-145 expression on tumor growth was
investigated in vivo by subcutaneous inoculation of the
miR-145 lentivirus-transfected JAR cells and mutant miR-145
lentivirus-transfected JAR cells into two groups of nude mice. All
the mice in the mutant miR-145 group developed tumors ∼37 days
after injection, whereas tumors were detected in only one in four
mice from the miR-145 transfected group at this time. Although both
groups developed tumors, the tumors formed by wt
miR-145-transfected cells grew more slowly than those in the mutant
miR-145-transfected group (Fig.
4). In addition, when the mice were sacrificed 62 days after
injection, tumor weights in the mutant miR-145 transfected group
were significantly heavier than those in the wt miR-145
lentivirus-transfected group. Furthermore, miR-145 expression in
the JAR cell line was associated with a significant decrease in
tumor volume (Fig. 4). These
results suggested that miR-145 expression in the human
choriocarcinoma cell line suppressed in vivo tumor
growth.
Discussion
Increasing evidence has shown that miRNA plays an
important role in the proliferation and invasion ability of
numerous types of cancer cells. However, the miRNAs that regulate
human choriocarcinoma cell growth and invasion have not yet been
reported. In the present study, it was observed that miR-145
interfered with Sox2 expression via putative sites located in the
3′-UTR region. Therefore, it was hypothesized that miR-145
suppresses the human choriocarcinoma cell line JAR by
downregulation of Sox2 expression. Putative miRNA target sites in
the 3′-UTR of Sox2 mRNA were used to construct a wt miR-145
expression lentivirus, which was then transfected into the JAR
human choriocarcinoma cell line. Luciferase activity assays
indicated that the activity of the Sox2 3′-UTR site was
significantly inhibited by wt miR-145, while that of the mutated
Sox2 3′-UTR site was unchanged, which suggested that miR-145
targeted Sox2. In addition, qRT-PCR and western blot
analysis demonstrated that Sox2 protein expression was reduced in
wt miR-145 lentivirus-transfected JAR cells compared with the
levels detected in mutant miR-145 lentivirus-transfected or
untransfected cells. FCM analysis revealed that the majority of wt
miR-145-transfected cells were arrested in the
G0/G1 phase of the cell cycle with reduced
percentages in the S and G2/M phases, which suggested
that miR-145 expression affected the cell cycle regulation of
choriocarcinoma cells in vitro. Similarly, exogenous miR-145
expression was shown to inhibit the growth of the JAR cell line
in vitro using MTT assays. Soft agar colony formation assay
and transwell migration invasion assays showed that the number of
invading wt miR-145 transfected cells was significantly lower than
the numbers of invading untransfected and mutant miR-145
transfected cells. Finally, xenograft experiments indicated that
miR-145 expressed in the JAR cell line also suppressed tumor growth
in vivo.
This is in contrast to a previous report which found
that the expression of high levels of Sox2 was associated with
malignancy in human lung cancer stem-like cells/cancer-initiating
cells (20). In studies of human
choriocarcinoma, it was observed that expressed levels of Sox2 in
human gestational trophoblastic neoplasia cells were higher than in
the normal trophoblast cells (19). Thus, we considered that there was
an association between Sox2 expression and malignancy in human
choriocarcinoma. By contrast, certain studies have also shown that
miR-145 may specifically regulate the target gene Sox2 expression
(21). Therefore, we investigated
whether the expression of endogenous Sox2 in human choriocarcinoma
was silenced when the miR-145 overexpression might weaken the
proliferation and invasion of carcinoma. In the current study,
exogenous miR-145 was transfected into the two human
choriocarcinoma cell lines, JAR and JEG-3 using a lentiviral
system. The effectiveness of miR-145 overexpression not only as an
inhibitor of endogenous Sox2 expression, but also as a suppressor
of proliferation and invasion in human choriocarcinoma cell lines,
was investigated in several ways, including cell proliferation,
invasion and infiltration assays, and tumorigenicity assays in nude
mice. The results suggest that not only proliferation, but also
invasion and infiltration were reduced following miR-145
overexpression in the human choriocarcinoma cell lines, JAR or
JEG-3. In comparison with the wt cell lines, the tumorigenicity in
nude mice of JAR cells transfected with exogenous miR-145 was
reduced. These results suggest that the proliferation and invasion
capacity of human choriocarcinoma cells is associated with Sox2
expression. It may be speculated that inhibition or loss of miR-145
expression results in excessive Sox2 expression, and therefore
influences tumor growth. Further studies are required to fully
elucidate the function of miR-145 in this process. However, it is
clear that miR-145 and Sox2 play potentially important roles in the
pathogenesis of human choriocarcinomas.
Acknowledgements
This study was supported by grants
from the Shanghai Committee Medical Science Foundation of China
(No.10411967100) to Te Liu and the National Natural Science
Foundation of China (31140037) to Zhixue Liu.
References
|
1
|
Bagley RG, Ren Y, Kurtzberg L, et al:
Human choriocarcinomas: placental growth factor-dependent
preclinical. Int J Oncol. 40:479–486. 2012.PubMed/NCBI
|
|
2
|
Monzo M, Navarro A, Bandres E, et al:
Overlapping expression of microRNAs in human embryonic colon and
colorectal cancer. Cell Res. 18:823–833. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim VN and Nam JW: Genomics of microRNA.
Trends Genet. 22:165–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hatfield SD, Shcherbata HR, Fischer KA,
Nakahara K, Carthew RW and Ruohola-Baker H: Stem cell division is
regulated by the microRNA pathway. Nature. 435:974–978. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eder M and Scherr M: MicroRNA and lung
cancer. N Engl J Med. 352:2446–2448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA, Ferracin M, Cimmino A, et al: A
MicroRNA signature associated with prognosis and progression in
chronic lymphocytic leukemia. N Engl J Med. 353:1793–1801. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Houbaviy HB, Murray MF and Sharp PA:
Embryonic stem cell-specific MicroRNAs. Dev Cell. 5:351–358. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lanza G, Ferracin M, Gafà R, et al:
mRNA/microRNA gene expression profile in microsatellite unstable
colorectal cancer. Mol Cancer. 6:542007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schetter AJ, Leung SY, Sohn JJ, et al:
MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. JAMA. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bandrés E, Cubedo E, Agirre X, et al:
Identification by Real-time PCR of 13 mature microRNAs
differentially expressed in colorectal cancer and non-tumoral
tissues. Mol Cancer. 5:292006.PubMed/NCBI
|
|
12
|
Volinia S, Calin GA, Liu CG, et al: A
microRNA expression signature of human solid tumors defines cancer
gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Liu T, Huang Y and Liu J:
microRNA-182 inhibits the proliferation and invasion of human lung
adenocarcinoma cells through its effect on human cortical
actin-associated protein. Int J Mol Med. 28:381–388.
2011.PubMed/NCBI
|
|
14
|
Chao A, Tsai CL, Wei PC, et al: Decreased
expression of microRNA-199b increases protein levels of SET
(protein phosphatase 2A inhibitor) in human choriocarcinoma. Cancer
Lett. 291:99–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tay Y, Zhang J, Thomson AM, Lim B and
Rigoutsos I: MicroRNAs to Nanog, Oct4 and Sox2 coding regions
modulate embryonic stem cell differentiation. Nature.
455:1124–1128. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chew JL, Loh YH, Zhang W, et al:
Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the
Oct4/Sox2 complex in embryonic stem cells. Mol Cell Biol.
25:6031–6046. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fong H, Hohenstein KA and Donovan PJ:
Regulation of self-renewal and pluripotency by Sox2 in human
embryonic stem cells. Stem Cells. 26:1931–1938. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu N, Papagiannakopoulos T, Pan G, Thomson
JA and Kosik KS: MicroRNA-145 regulates OCT4, SOX2, and KLF4 and
represses pluripotency in human embryonic stem cells. Cell.
137:647–658. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li AS, Siu MK, Zhang H, et al:
Hypermethylation of SOX2 gene in hydatidiform mole and
choriocarcinoma. Reprod Sci. 15:735–744. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nakatsugawa M, Takahashi A, Hirohashi Y,
et al: SOX2 is over-expressed in stem-like cells of human lung
adenocarcinoma and augments the tumorigenicity. Lab Invest.
91:1796–1804. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu T, Cheng W, Huang Y, Huang Q, Jiang L
and Guo L: Human amniotic epithelial cell feeder layers maintain
human iPS cell pluripotency via inhibited endogenous microRNA-145
and increased Sox2 expression. Exp Cell Res. 318:424–434. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eis PS, Tam W, Sun L, et al: Accumulation
of miR-155 and BIC RNA in human B cell lymphomas. Proc Natl Acad
Sci USA. 102:3627–3632. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Chao T, Li R, et al: MicroRNA-128
inhibits glioma cells proliferation by targeting transcription
factor E2F3a. J Mol Med (Berl). 87:43–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng W, Liu T, Jiang F, et al:
microRNA-155 regulates angiotensin II type 1 receptor expression in
umbilical vein endothelial cells from severely pre-eclamptic
pregnant women. Int J Mol Med. 27:393–399. 2011.PubMed/NCBI
|
|
25
|
Gupta RA, Shah N, Wang KC, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang F, Liu T, He Y, et al: MiR-125b
promotes proliferation and migration of type II endometrial
carcinoma cells through targeting TP53INP1 tumor suppressor in
vitro and in vivo. BMC Cancer. 11:4252011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carro MS, Lim WK, Alvarez MJ, et al: The
transcriptional network for mesenchymal transformation of brain
tumours. Nature. 463:318–325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kluiver J, Haralambieva E, de Jong D, et
al: Lack of BIC and microRNA miR-155 expression in primary cases of
Burkitt lymphoma. Genes Chromosomes Cancer. 45:147–153. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martin MM, Lee EJ, Buckenberger JA,
Schmittgen TD and Elton TS: MicroRNA-155 regulates human
angiotensin II type 1 receptor expression in fibroblasts. J Biol
Chem. 281:18277–18284. 2006. View Article : Google Scholar : PubMed/NCBI
|