Introduction
Reperfusion, the prompt restoration of the blood
supply to the ischemic tissue, is the most effective way to reduce
the process of ischemic injury, which ultimately leads to cell
death. However, reperfusion following even brief periods of
ischemia causes irreversible damage, known as ischemia-reperfusion
injury (1). Although the
mechanisms underlying ischemia-reperfusion injury are complicated,
oxidative stress is considered to play a pivotal role.
Hydrogen sulfide (H2S) was the third
gaseous signaling molecule to be discovered, following nitric oxide
and carbon monoxide (2).
H2S is produced endogenously from cysteine by the
pyridoxal-5′-phosphate-dependent enzymes, cystathionine β-synthase
(CBS) and/or cystathionine γ-lyase (CSE). Previously, a number of
studies have suggested that H2S has anti-inflammatory,
anti-oxidative and anti-apoptotic effects (3,4).
H2S inhibits lipid peroxidation during heart
ischemia-reperfusion and decreases the mortality of myocardial
cells induced by ischemia by reducing oxygen free radicals
(4). Also, in brain
ischemia-reperfusion injury, H2S exerts a protective
effect on neurons by eliminating oxygen free radicals (5). Studies have shown that H2S
has protective effects on the heart, brain, liver and lung in the
case of ischemia-reperfusion injury (6–9). In
the gastrointestinal tract, it has been reported that the systemic
administration of sodium hydrogen sulfide (NaHS), a H2S
donor, attenuates gastric mucosal injury by downregulating mRNA
expression and plasma release of proinflammatory cytokines in rats
(10). The protective effect of
exogenously administered H2S and its precursors NaHS and
L-cysteine (L-cys), has been shown against ischemia-reperfusion
injury. However, the potential of endogenous H2S has not
yet been investigated. The aim of this study was to investigate
whether endogenous H2S plays a role in gastric
ischemia-reperfusion (GI-R) injury.
We hypothesize that H2S plays an
important role in the gastric mucosa under ischemia-reperfusion.
Thus, we explored the effect of endogenous H2S on rat
GI-R injury by pretreatment with DL-propargylglycine (PAG) and
L-cys to block CSE and H2S synthetase and provide a
precursor of H2S synthesis. Furthermore, we determined
whether the effect of endogenous H2S is related to
oxidative enzymes involved in GI-R.
Materials and methods
Animals
Adult male Sprague-Dawley rats weighing 200–250 g
were provided by the Animal Department of Xuzhou Medical College.
All animal care and experimental protocols complied with the Animal
Management Rules of the Ministry of Health of the People’s Republic
of China and the guidelines for the Care and Use of Laboratory
Animals of Xuzhou Medical College.
Chemicals
L-cys and PAG were purchased from Sigma-Aldrich (St.
Louis, MO, USA). Malondialdehyde (MDA), glutathione (GSH),
superoxide dismutase (SOD) and superoxide anion
(O2−) assay kits were purchased from Nanjing
Jiancheng Bioengineering Institute (Nanjing, China). Rabbit
polyclonal antibodies to SOD-1 (sc-11407) and xanthine oxidase
(XOD, sc-20991) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Rabbit polyclonal antibodies to CSE (BA2198)
were purchased from Wuhan Boster Biological Technology Co. Ltd.,
(Wuhan, China). Goat anti-rabbit IgG was purchased from Beijing
Zhong Shan-Golden Bridge Biological Technology Co., Ltd. (Beijing,
China). All other chemicals and reagents were of analytical
grade.
GI-R injury model
The rats were fasted without water deprivation for
24 h before the experiments. After inducing anesthesia with an
intraperitoneal injection of 10% chloral hydrate (400 mg/kg body
weight), the rats were fixed on an operating table. Following
laparotomy, the celiac artery was carefully separated from
surrounding tissues, clamped with an artery clamp to induce
ischemia and later removed to allow reperfusion (11). Following surgery, all rats were
sacrificed under anesthesia and the stomach was carefully excised
for determination of gastric mucosal damage or stored at −80°C for
MDA and GSH assays, and the measurement of XOD and SOD expression
by western blot analysis.
Measurement of H2S
concentration in gastric mucosa and serum
Following sacrifice, a blood sample was rapidly
collected from each rat and centrifuged at 4,000 rpm for 4 min.
Then the supernatant was used for H2S measurement using
a commercially-available kit (12). The mucosa of the stomach was
scraped off, homogenized and centrifuged and then the supernatant
was collected for the measurement of H2S
concentration.
Measurement of gastric mucosal
damage
The stomach was cut open along the greater gastric
curvature, rinsed and flattened. The general injury area of the
gastric mucosa was calculated using Adobe Photoshop 6.0 (Adobe
Systems Inc., San Jose, CA, USA). The injury area was expressed as
the percentage of congestion, edema and erosion in the whole
gastric mucosa area. A 1-mm tissue sample was removed from between
the greater and lesser gastric curvatures and fixed in 10%
formaldehyde, then processed by routine paraffin embedding,
sectioning and hematoxylin and eosin (H&E) staining. The degree
of pathological injury was assessed under a light microscope (each
sample was blindly evaluated by a pathologist). The degree of
injury was scored according to the Masuda criteria (13) with slight modification: normal, 0;
injury in surface epithelium, 1; congestion and edema in the upper
mucosa, 2; congestion, hemorrhage and edema in the middle and lower
mucosa, 3; structural disorder or necrosis in the upper mucosal
glands, 4 and deep necrosis and ulceration, 5. The average injury
score for each section was calculated. The mean of 10 visual scores
from each slide was calculated as the score for one rat.
Assay of MDA and GSH content, SOD
activity and the inhibition of O2−
production
The gastric mucosa was made into a 10% tissue
homogenate and centrifuged at 3,000 rpm, 4°C for 10 min; then the
supernatant was collected. Following the manufacturer’s
instructions, the MDA and GSH contents in the supernatant were
assessed by the thiobarbituric acid and
3,3′-dithiobis(6-nitrobenzoic) acid methods, respectively. The
protein content was determined by the bicinchoninic acid (BCA)
method. SOD activity in the supernatant was evaluated by the
inhibition of XOD, according to the manufacturer’s instructions and
expressed as units per milligram tissue (U/mg). Inhibition of
O2− production was measured according to the
manufacturer’s instructions and expressed as units per gram tissue
(U/g).
Western blot analysis of CSE, XOD and
SOD
Gastric mucosal protein was extracted according to
the BCA method. The concentration of each sample was diluted to the
same level, then mixed with 1/3 volume protein denaturation
solution and boiled for 5 min to denature the protein. Samples were
separated on 10% sodium dodecyl sulphate (SDS)-polyacrylamide gels.
Membranes were probed with polyclonal antibodies to CSE, SOD and
XOD overnight at 4°C. The secondary antibody was conjugated to
horseradish peroxidase with a BCIP/NBT kit (Promega Corporation,
Madison, WI, USA). β-actin was used to normalize for loading
variations.
Experimental protocol
The rats were randomly divided into 4 groups with 10
per group: i) sham group, age-matched healthy rats were only
laparotomized without celiac artery clamping; ii) GI-R group,
age-matched healthy rats were intraperitoneally injected with
normal saline for 7 days, then the celiac arteries were clamped for
30 min ischemia and then reperfused for 60 min; iii) PAG group, the
rats were intraperitoneally injected with the H2S
synthetase blocker PAG (50 mg/kg/day) for 7 days, then the celiac
arteries were clamped for 30 min ischemia and then reperfused for
60 min and iv) L-cys group, the rats were intraperitoneally
injected with L-cys (50 mg/kg/day) for 7 consecutive days, then the
celiac arteries were clamped for 30 min ischemia and then
reperfused for 60 min.
Statistical analysis
All data are expressed as mean ± standard deviation
(SD). Statistical analysis was performed by SPSS 13.0 (SPSS Inc.,
Chicago, IL, USA) for Windows. Statistical significance was
calculated by one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
Gastric mucosal injury
The mucosal surface of the sham group was smooth and
no significant abnormality in the gastric mucosa was observed under
the light microscope. However, significant hemorrhage and edema and
several erosions of varying depths and sizes were observed on the
surface of the mucosa of the GI-R group. The epithelial cells and
gland ducts of the mucosa at the hemorrhage sites were shed and
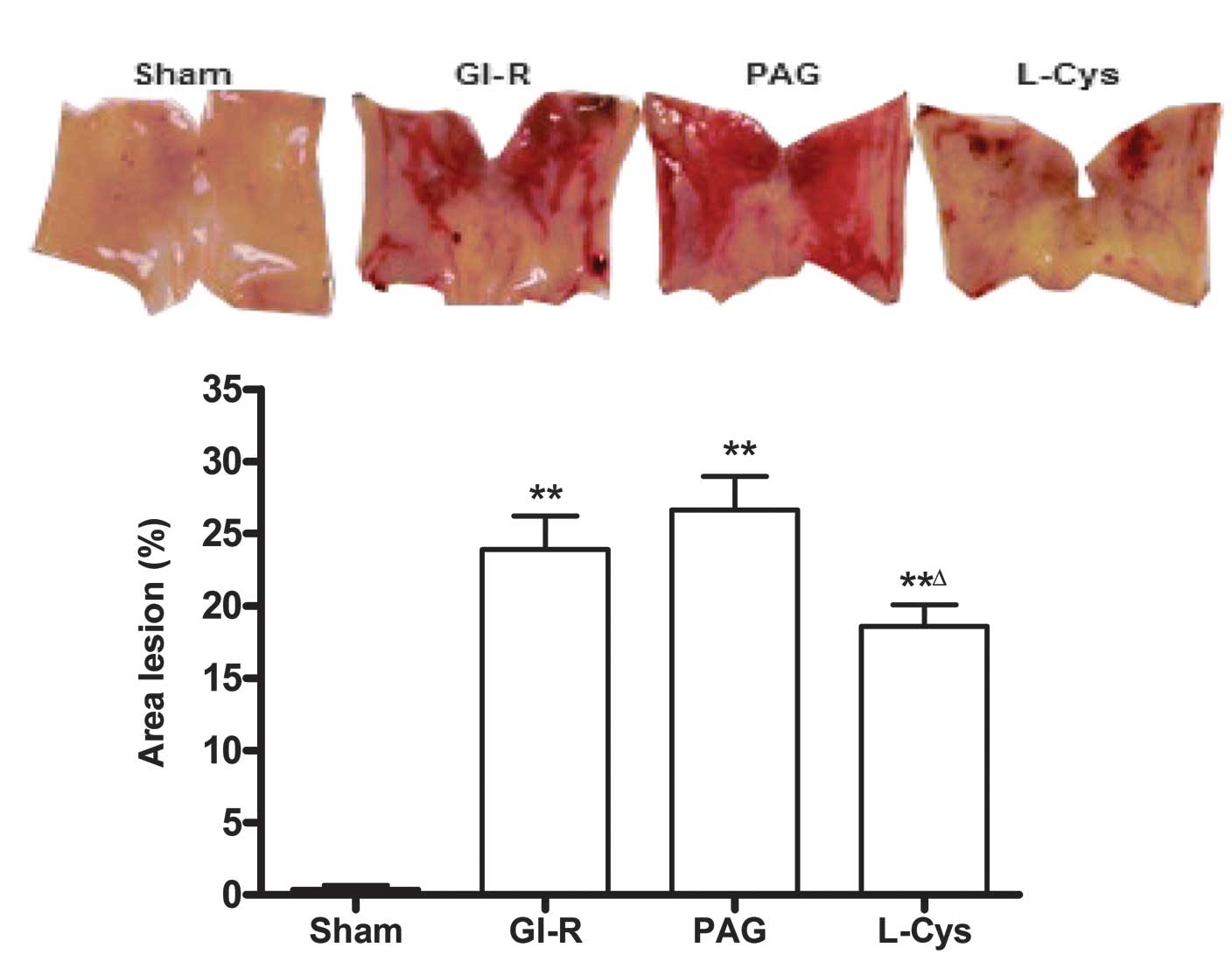
disorganized in the GI-R group. Compared with the GI-R group, the
injury area and the extent of mucosal damage significantly
increased in the PAG group and L-cys inhibited the damage induced
by GI-R (P<0.05; Fig. 1). Under
the light microscope, the gastric damage scores in the GI-R, PAG
and L-cys groups were much higher than those in the sham group
(P<0.01). However, the damage score in the L-cys group was lower
than those of the GI-R and PAG groups (P<0.05, Fig. 2).
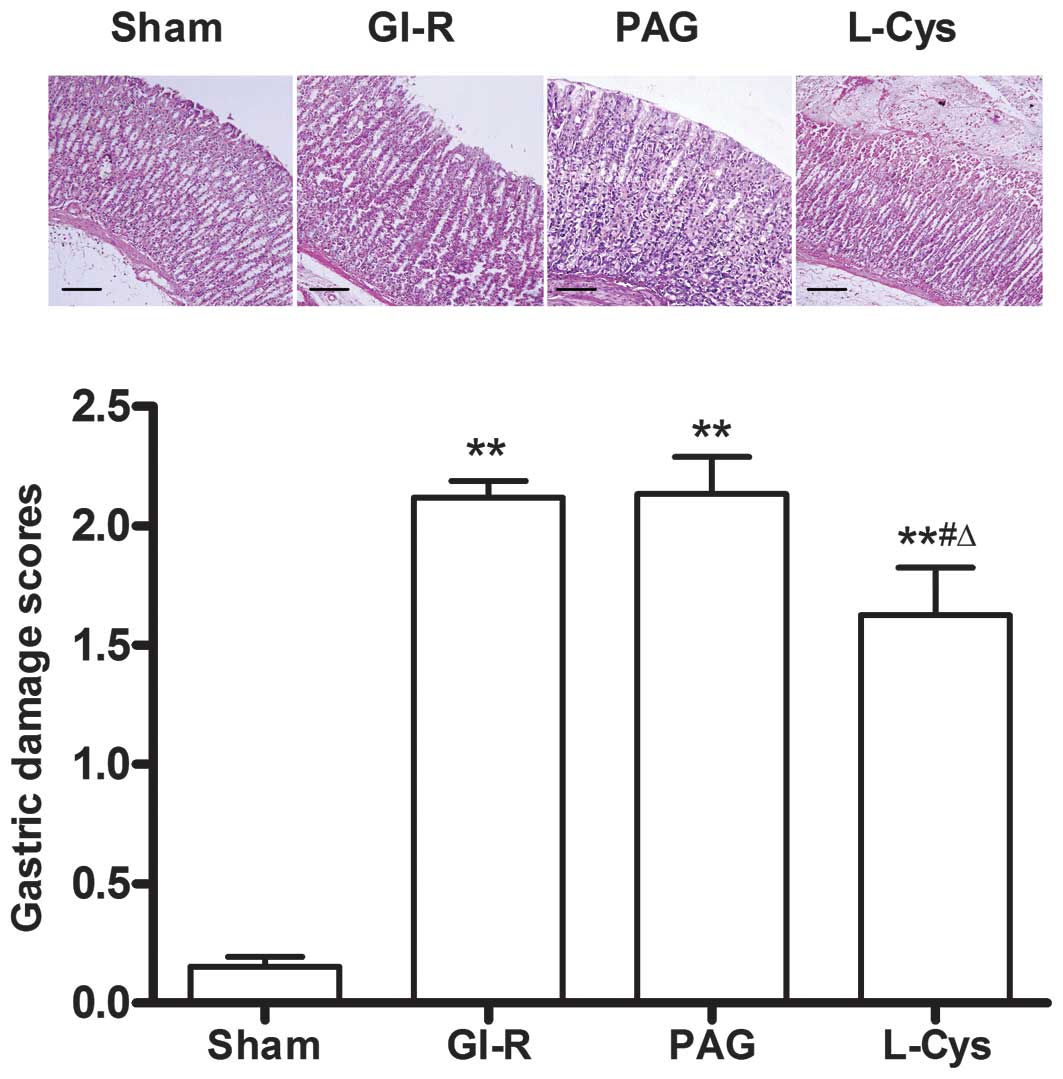 | Figure 2.Effects of PAG and L-cys on gastric
mucosal injury scores in rats subjected to gastric
ischemia-reperfusion (GI-R) (hematoxylin and eosin stain;
magnification, ×100). Male Sprague-Dawley rats were
intraperitoneally injected with the H2S synthetase
blocker PAG (50 mg/kg) or L-cys (50 mg/kg) for 7 days before the
celiac arteries were clamped for 30 min ischemia and then
reperfused for 60 min. Sham group, age-matched rats with
physiological solute treatment but no GI-R procedure; GI-R group,
age-matched rats with physiological solute treatment followed by
the GI-R procedure; PAG group, rats intraperitoneally injected with
PAG and then subjected to the GI-R procedure; L-cys group, rats
intraperitoneally injected with L-cys and then subjected to the
GI-R procedure. Scale bar, 200 μm. Data presented as mean ±
standard deviation (SD), n=8. **P<0.01 vs. the sham
group, #P<0.05 vs. the GI-R group,
ΔP<0.05 vs. the PAG group. PAG, DL-propargylglycine;
L-cys, L-cysteine; H2S, hydrogen sulfide. |
H2S concentration in the serum
and gastric mucosal tissue
Although there were no significant changes in
H2S concentration in the serum (Fig. 3B), compared with the sham and GI-R
groups, PAG decreased the concentration of H2S in the
gastric mucosa (P<0.05) and L-cys attenuated this decrease
(P<0.05). However, there was no significant difference in
H2S level in the gastric mucosa between the GI-R and
sham groups (Fig. 3A).
 | Figure 3.Effects of PAG and L-cys on
H2S concentration in the gastric mucosa (A) and serum
(B) from rats subjected to gastric ischemia-reperfusion (GI-R).
Male Sprague-Dawley rats were intraperitoneally injected with the
H2S synthetase blocker PAG (50 mg/kg) or L-cys (50
mg/kg) for 7 days before the celiac arteries were clamped for 30
min ischemia and then reperfused for 60 min. Sham group,
age-matched rats with physiological solute treatment but no GI-R
procedure; GI-R group, age-matched rats with physiological solute
treatment followed by the GI-R procedure; PAG group, rats
intraperitoneally injected with PAG and then subjected to the GI-R
procedure; L-cys group, rats intraperitoneally injected with L-cys
and then subjected to the GI-R procedure. Data presented as mean ±
standard deviation (SD), n=8. *P<0.05 vs. the sham
group, #P<0.05 vs. the GI-R group,
ΔP<0.05 vs. the PAG group. PAG, DL-propargylglycine;
L-cys, L-cysteine; H2S, hydrogen sulfide. |
CSE expression in the gastric mucosa
Compared with the sham group, GI-R alone had no
effect on CSE expression. PAG significantly inhibited the
expression of CSE (P<0.01) and L-cys increased CSE in the
gastric mucosa to normal levels (P<0.01; Fig. 4).
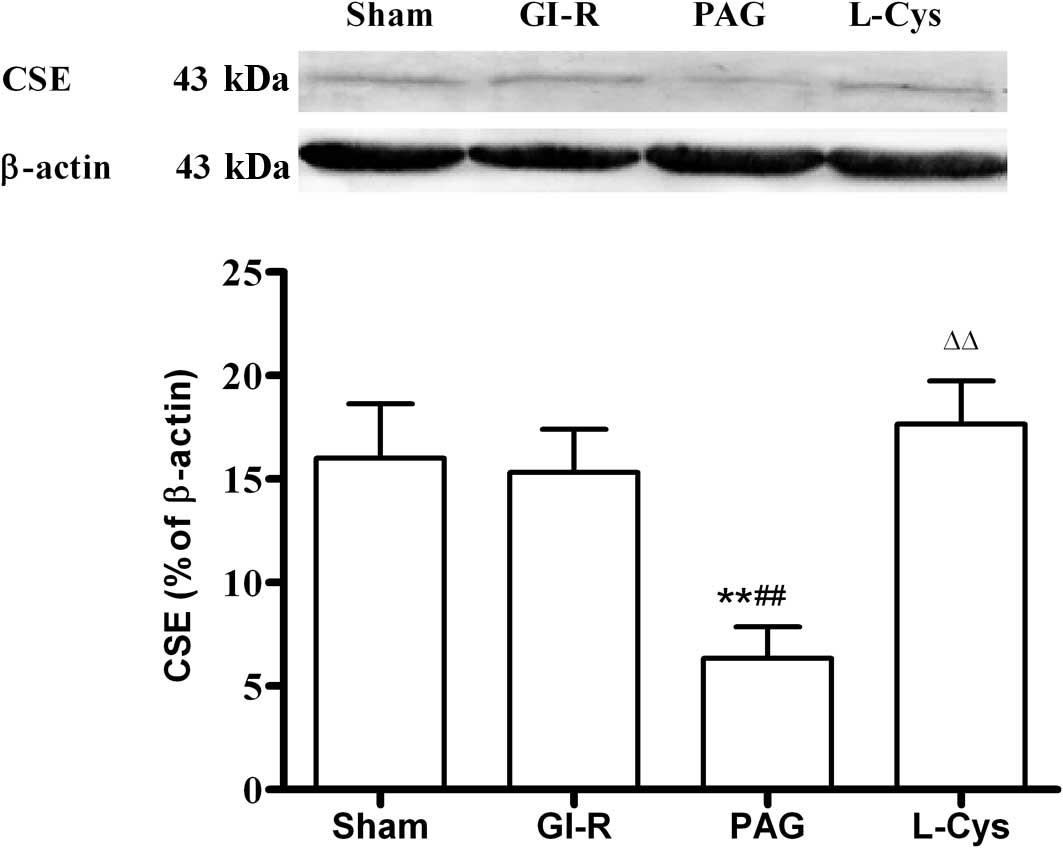 | Figure 4.Effects of PAG and L-cys on CSE
expression in gastric mucosa from rats subjected to gastric
ischemia-reperfusion (GI-R). Male Sprague-Dawley rats were
intraperitoneally injected with the H2S synthetase
blocker PAG (50 mg/kg) or L-cys (50 mg/kg) for 7 days before the
celiac arteries were clamped for 30 min ischemia and then
reperfused for 60 min. Sham group, age-matched rats with
physiological solute treatment but no GI-R procedure; GI-R group,
age-matched rats with physiological solute treatment followed by
the GI-R procedure; PAG group, rats intraperitoneally injected with
PAG and then subjected to the GI-R procedure; L-cys group, rats
intraperitoneally injected with L-cys and then subjected to the
GI-R procedure. Data presented as mean ± standard deviation (SD),
n=3. **P<0.01 vs. the sham group,
##P<0.01 vs. the GI-R group, ΔΔP<0.01
vs. the PAG group. PAG, DL-propargylglycine; L-cys, L-cysteine;
H2S, hydrogen sulfide; CSE, cystathionine γ-lyase. |
MDA and GSH contents in the gastric
mucosa
The MDA content of the mucosa increased following
GI-R (P<0.05) and further increased in the PAG group. However,
L-cys decreased the concentration of MDA in mucosa (Fig. 5A). The GSH content in the mucosa
decreased in the GI-R, PAG and L-cys groups. However, the reduction
of GSH content in the L-cys group was less than that in the PAG
group (P<0.05; Fig. 5B).
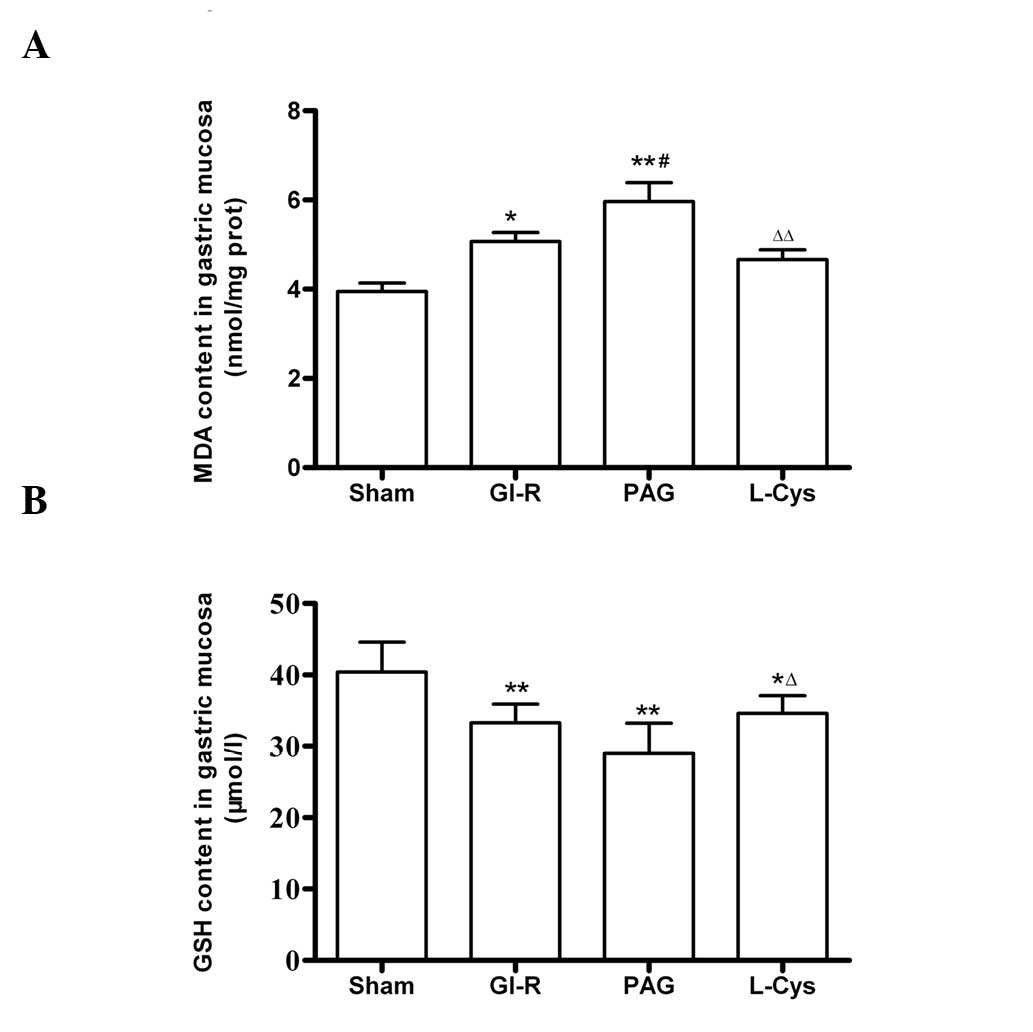 | Figure 5.Effects of PAG and L-cys on (A) MDA
and (B) GSH content in gastric mucosa from rats subjected to
gastric ischemia-reperfusion (GIR). Male Sprague-Dawley rats were
intraperitoneally injected with the H2S synthetase
blocker PAG (50 mg/kg) or L-cys (50 mg/kg) for 7 days before the
celiac arteries were clamped for 30 min ischemia and then
reperfused for 60 min. Sham group, age-matched rats with
physiological solute treatment but no GI-R procedure; GI-R group,
age-matched rats with physiological solute treatment followed by
the GI-R procedure; PAG group, rats intraperitoneally injected with
PAG and then subjected to the GI-R procedure; L-cys group, rats
intraperitoneally injected with L-cys and then subjected to the
GI-R procedure. Data presented as mean ± standard deviation (SD),
n=8. *P<0.05, **P<0.01 vs. the sham
group; #P<0.05 vs. the GI-R group;
ΔP<0.05, ΔΔP<0.01 vs. the PAG group.
PAG, DL-propargylglycine; L-cys, L-cysteine; H2S,
hydrogen sulfide; MDA, malondialdehyde; GSH, glutathione. |
XOD and SOD expression, SOD activity and
inhibition of O2− production in the gastric
mucosa
Compared with the sham group (P<0.01; Fig. 6A), the expression level of XOD
markedly increased in the GI-R and PAG groups; while the SOD
expression in the GI-R and PAG groups decreased compared with the
sham group (P<0.01; Fig. 6B).
Compared with the GI-R group, L-cys downregulated XOD (P<0.01)
and upregulated SOD (P<0.01). Additionally, the activity of SOD
was decreased in the GI-R and PAG groups and the effects of GI-R
were attenuated by L-cys (P<0.01; Fig. 6C), as was the inhibition of
O2− production (Fig. 6D).
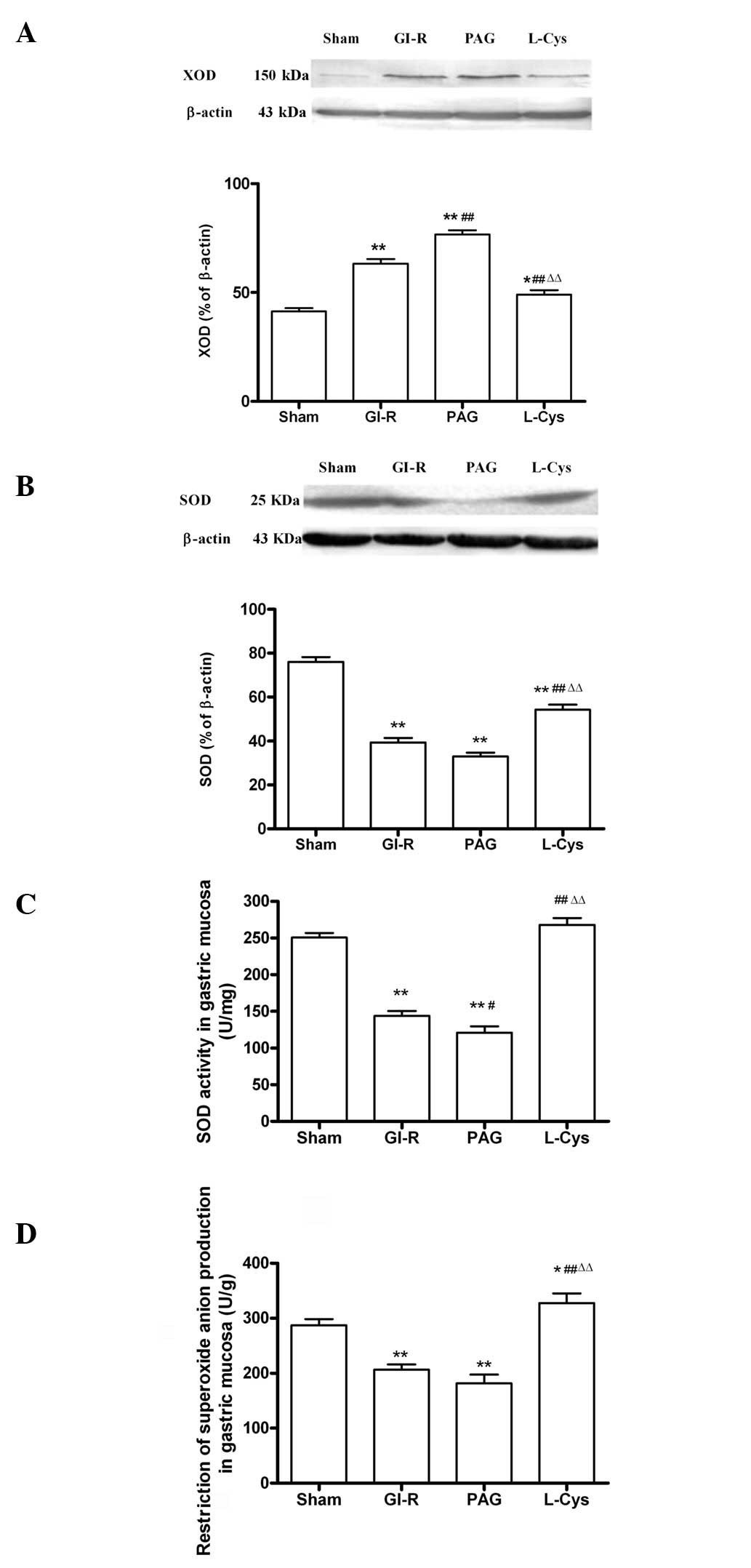 | Figure 6.Alteration of oxidative stress in
gastric mucosal tissue from rats subjected to 30 min gastric
ischemia and 60 min reperfusion (GI-R). (A) Effects of PAG and
L-cys on XOD expression in gastric mucosal tissue induced by GI-R
(n=3). (B) Effects of PAG and L-cys on SOD expression in gastric
mucosal tissue induced by GI-R (n=3). β-actin was used to normalize
for loading variations. (C) Effects of PAG and L-cys on SOD
activity in gastric mucosal tissue induced by GI-R (n=8). (D)
Effects of PAG and L-cys on the alteration of
O2− production in gastric mucosal tissue
induced by GI-R. Male Sprague-Dawley rats (n=8) were
intraperitoneally injected with the H2S synthetase
blocker PAG (50 mg/kg) or L-cys (50 mg/kg) for 7 days before the
celiac arteries were clamped for 30 min ischemia and then
reperfused for 60 min. Sham group, age-matched rats with
physiological solute treatment but no GI-R procedure; GI-R group,
age-matched rats with physiological solute treatment followed by
the GI-R procedure; PAG group, rats intraperitoneally injected with
PAG and then subjected to the GI-R procedure; L-cys group, rats
intraperitoneally injected with L-cys and then subjected to the
GI-R procedure. Data presented as mean ± standard deviation (SD).
*P<0.05, **P<0.01 vs. the sham group,
#P<0.05, ##P<0.01 vs. the GI-R group,
ΔΔP<0.01 vs. the PAG group. PAG, DL-propargylglycine;
L-cys, L-cysteine; H2S, hydrogen sulfide; XOD, xanthine
oxidase; SOD, superoxide dismutase. |
Discussion
The occurrence of GI-R injury is related to a number
of factors, including excessive production of oxygen free radicals
in the mucosa (14), leukocyte
infiltration (15) and decreased
release of nitric oxide (16).
Oxidative stress induced by high levels of active oxygen plays a
major role in GI-R injury. Excess production of oxygen free
radicals is a major initiating factor and an independent pathogenic
factor of GI-R injury (17,18).
Therefore, investigating how to decrease oxidative stress is
essential for developing means of protecting the gastric mucosa
from attack by deleterious factors.
H2S is well-known as a toxic gas with the
smell of rotten eggs. Nevertheless, with the increasing interest in
endogenous gaseous signaling molecules, it has been shown that
endogenous H2S regulates a range of physiological and
pathological processes in the nervous, digestive and cardiovascular
systems (19–22). Hence, H2S is considered
to be a physiologically important molecule and a gaseous mediator.
In mammals, endogenous H2S is generated from L-cys under
the catalysis of total CBS and CSE, 1/3 of which is in the form of
gas while 2/3 is in the form of NaHS. NaHS dissociates in
vivo into sodium ions and sulfhydryl group ions and the latter
bind with hydrogen ions to generate H2S. Thus,
H2S and NaHS are in dynamic equilibrium (17). L-cys administration is
cardioprotective through enhanced myocardial H2S
generation as a result of CSE activation, which is attenuated by
the selective CSE enzyme inhibitor, PAG (23). Meanwhile, H2S protects
brain endothelial cells from oxidative stress (24). In this study, we identified that in
rats pretreated with a H2S synthetase blocker (PAG, 50
mg/kg/day) for 7 days, significant hemorrhage, edema and erosions
were observed in the surface of the gastric mucosa, as well as in
the GI-R group. However, pretreatment with L-cys, the precursor of
H2S, protected the mucosa from the damage induced by
GI-R (Fig. 1). The same results
were observed for gastric damage scores. After blocking the
production of H2S with PAG, gastric mucosal ulceration
and the number of hemorrhage sites significantly increased, a
number of mucosal cells died, erosion sites formed and significant
edema and inflammatory infiltration appeared in the submucosa. The
signs of gastric mucosal injury were markedly alleviated by the
continuous administration of L-cys (Fig. 2). Although there was no significant
change in the H2S concentration in serum, PAG
successfully inhibited H2S production (Fig. 3) and CSE expression (Fig. 4) in the gastric mucosa, while L-cys
did not cause any evident change in H2S. Endogenous
H2S in the gastric mucosa plays an important role in the
protective effect against GI-R injury. Whiteman et al
identified that in the rat brain, H2S protects neurons
from injury by eliminating oxygen free radicals (5).
A number of studies have reported that oxidative
stress plays a pivotal role in GI-R injury. In the GI-R process,
lipid peroxidation is induced by an increased number of oxygen free
radicals. A change of MDA content is a measure of the degree of
damage caused by membrane lipid peroxidation. In our study, when
H2S was inhibited by PAG, lipid peroxidation increased
in the gastric mucosa since the MDA content was higher than that of
the GI-R group. However, when H2S was increased, the MDA
level decreased (Fig. 5A). This
result is consistent with the findings on the effect of
H2S on a rat model of myocardial infarction (25). Furthermore, GSH is the major
endogenous antioxidant produced by mammalian cells, preventing
damage to important cellular components caused by reactive oxygen
species (26). We identified that
L-cys increased the GSH content when compared with the PAG group
(Fig. 5B). When H2S
production was blocked, MDA levels increased. Additionally,
increasing endogenous H2S production reduced MDA content
since GSH levels were enhanced. This suggests that endogenous
H2S protects the integrity of the gastric mucosa by
increasing GSH and decreasing MDA content. This is consistent with
the finding that H2S has anti-oxidative effects by
promoting the transfer of cystine into cells and improving the
cellular synthesis of GSH (27).
The antioxidant action of H2S plays an important role
during GI-R.
During the reperfusion period following ischemia or
hypoxia, large amounts of superoxide and O2−
are generated from xanthine and hypoxanthine under the action of
XOD. XOD is considered the most important source of oxygen free
radicals (28). Meanwhile, living
tissues are endowed with innate antioxidant defense mechanisms,
namely antioxidative enzymes. When the amount of active oxygen
exceeds the scavenging capacity of the antioxidant defensive
system, including SOD, the immunological function of the
gastrointestinal tract is severely damaged by free radicals, which
results in tissue and organ injury (29). In this study, we identified that
blocking the synthesis of endogenous H2S significantly
increased the expression of XOD (Fig.
6A), causing oxygen free radical overproduction in the gastric
mucosa. Nevertheless, the activity of the antioxidative enzyme,
SOD, was enhanced by increasing H2S in rats pretreated
with L-cys (Fig. 6B and C). As a
result, the ability to inhibit O2− production
was clearly reduced due to the reduction of endogenous
H2S by PAG (Fig. 6D).
The protective role of H2S against oxidative stress has
also been clarified in rat gastric mucosal epithelium (30).
In conclusion, this study demonstrates that
endogenous H2S protects the gastric mucosa against
ischemia and reperfusion injury. Selective inhibition of the CSE
enzyme enhances the injury following GI-R. However, L-cys
administration attenuates the harmful effects of GI-R through
activation of CSE to increase H2S generation. The
gastroprotective effect of endogenous H2S against GI-R
may be mediated by enhancing the anti-oxidative capacity through
increasing GSH and SOD to reduce free radical production.
Abbreviations:
|
H2S
|
hydrogen sulfide;
|
|
GI-R
|
gastric ischemiareperfusion;
|
|
PAG
|
DL-propargylglycine;
|
|
MDA
|
malondialdehyde;
|
|
GSH
|
glutathione;
|
|
SOD
|
superoxide dismutase;
|
|
CSE
|
cystathionine γ-lyase;
|
|
XOD
|
xanthine oxidase;
|
|
L-cys
|
L-cysteine
|
Acknowledgements
This study was supported by grants
from the Natural Science Foundation of Jiangsu Province
(BK2009088). The authors thank Dr Jinsong Bian for critical reading
of this manuscript and Dr Iain C Bruce for refining the English
language.
References
|
1.
|
Gross GJ and Auchampach JA: Reperfusion
injury: does it exist? J Mol Cell Cardiol. 42:12–18. 2007.
View Article : Google Scholar
|
|
2.
|
Wang R: Two’s company, three’s a crowd:
can H2S be the third endogenous gaseous transmitter?
FASEB J. 16:1792–1798. 2002.
|
|
3.
|
Tokuda K, Kida K, Marutani E, Crimi E,
Bougaki M, Khatri A, Kimura H and Ichinose F: Inhaled hydrogen
sulfide prevents endotoxin-induced systemic inflammation and
improves survival by altering sulfide metabolism in mice. Antioxid
Redox Signal. 17:11–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Jiang LH, Luo X, He W, Huang XX and Cheng
TT: Effects of exogenous hydrogen sulfide on apoptosis proteins and
oxidative stress in the hippocampus of rats undergoing heroin
withdrawal. Arch Pharm Res. 34:2155–2162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Whiteman M, Armstrong JS, Chu SH, Jia-Ling
S, Wong BS, Cheung NS, Halliwell B and Moore PK: The novel
neuro-modulator hydrogen sulfide: an endogenous peroxynitrite
‘scavenger’? J Neurochem. 90:765–768. 2004.PubMed/NCBI
|
|
6.
|
Sivarajah A, McDonald MC and Thiemermann
C: The production of hydrogen sulfide limits myocardial ischemia
and reperfusion injury and contributes to the cardioprotective
effects of preconditioning with endotoxin, but not ischemia in the
rat. Shock. 26:154–161. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Tay AS, Hu LF, Lu M, Wong PT and Bian JS:
Hydrogen sulfide protects neurons against hypoxic injury via
stimulation of ATP-sensitive potassium channel/protein kinase
C/extra-cellular signal-regulated kinase/heat shock protein 90
pathway. Neuroscience. 167:277–286. 2010. View Article : Google Scholar
|
|
8.
|
Kang K, Zhao M, Jiang H, Tan G, Pan S and
Sun X: Role of hydrogen sulfide in hepatic
ischemia-reperfusion-induced injury in rats. Liver Transpl.
15:1306–1314. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Fu Z, Liu X, Geng B, Fang L and Tang C:
Hydrogen sulfide protects rat lung from ischemia-reperfusion
injury. Life Sci. 82:1196–1202. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Mard SA, Neisi N, Solgi G, Hassanpour M,
Darbor M and Maleki M: Gastroprotective effect of NaHS against
mucosal lesions induced by ischemia-reperfusion injury in rat. Dig
Dis Sci. 57:1496–1503. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Wada K, Kamisaki Y, Kitano M, Kishimoto Y,
Nakamoto K and Itoh T: A new gastric ulcer model induced by
ischemia-reperfusion in the rat: role of leukocytes on ulceration
in rat stomach. Life Sci. 59:PL295–PL301. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Pan TT, Feng ZN, Lee SW, Moore PK and Bian
JS: Endogenous hydrogen sulfide contributes to the cardioprotection
by metabolic inhibition preconditioning in the rat ventricular
myocytes. J Mol Cell Cardiol. 40:119–130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Masuda E, Kawano S, Nagano K, Tsuji S,
Takei Y, Hayashi N, Tsujii M, Oshita M, Michida T, Kobayashi I, et
al: Role of endogenous endothelin in pathogenesis of
ethanol-induced gastric mucosal injury in rats. Am J Physiol.
265:G474–G481. 1993.PubMed/NCBI
|
|
14.
|
Kwiecień S, Brzozowski T and Konturek SJ:
Effects of reactive oxygen species action on gastric mucosa in
various models of mucosal injury. J Physiol Pharmacol. 53:39–50.
2002.PubMed/NCBI
|
|
15.
|
Andrews FJ, Malcontenti-Wilson C and
O’Brien PE: Polymorphonuclear leukocyte infiltration into gastric
mucosa after ischemia-reperfusion. Am J Physiol. 266:G48–G54.
1994.PubMed/NCBI
|
|
16.
|
Wada K, Kamisaki Y, Ohkura T, Kanda G,
Nakamoto K, Kishimoto Y, Ashida K and Itoh T: Direct measurement of
nitric oxide release in gastric mucosa during ischemia-reperfusion
in rats. Am J Physiol. 274:G465–G471. 1998.PubMed/NCBI
|
|
17.
|
Ishii M, Shimizu S, Nawata S, Kiuchi Y and
Yamamoto T: Involvement of reactive oxygen species and nitric oxide
in gastric ischemia-reperfusion injury in rats: protective effect
of tetrahydrobiopterin. Dig Dis Sci. 45:93–98. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Itoh M and Guth PH: Role of oxygen-derived
free radicals in hemorrhagic shock-induced gastric lesions in the
rat. Gastroenterology. 88:1162–1167. 1985.PubMed/NCBI
|
|
19.
|
El Eter E, Hagar HH, Al-Tuwaijiri A and
Arafa M: Nuclear factor-kappaB inhibition by
pyrrolidinedithiocarbamate attenuates gastric ischemia-reperfusion
injury in rats. Can J Physiol Pharmacol. 83:483–492.
2005.PubMed/NCBI
|
|
20.
|
Kimura Y, Dargusch R, Schubert D and
Kimura H: Hydrogen sulfide protects HT22 neuronal cells from
oxidative stress. Antioxid Redox Signal. 8:661–670. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Kimura Y and Kimura H: Hydrogen sulfide
protects neurons from oxidative stress. FASEB J. 18:1165–1167.
2004.PubMed/NCBI
|
|
22.
|
Zhang Z, Huang H, Liu P, Tang C and Wang
J: Hydrogen sulfide contributes to cardioprotection during
ischemia-reperfusion injury by opening K ATP channels. Can J
Physiol Pharmacol. 85:1248–1253. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Elsey DJ, Fowkes RC and Baxter GF:
L-cysteine stimulates hydrogen sulfide synthesis in myocardium
associated with attenuation of ischmia-reperfusion injury. J
Cardiovasc Pharmacol Ther. 15:53–59. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Tyagi N, Moshal KS, Sen U, Vacek TP, Kumar
M, Hughes WM Jr, Kundu S and Tyagi SC: H2S protects
against methionine-induced oxidative stress in brain endothelial
cells. Antioxid Redox Signal. 11:25–33. 2009.
|
|
25.
|
Zhu YZ, Wang ZJ, Ho P, Loke YY, Zhu YC,
Huang SH, Tan CS, Whiteman M, Lu J and Moore PK: Hydrogen sulfide
and its possible roles in myocardial ischemia in experimental rats.
J Appl Physiol. 102:261–268. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Pompella A, Visvikis A, Paolicchi A, De
Tata V and Casini AF: The changing faces of glutathione, a cellular
protagonist. Biochem Pharmacol. 66:1499–1503. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Liu H, Bai XB, Shi S and Cao YX: Hydrogen
sulfide protects from intestinal ischaemia-reperfusion injury in
rats. J Pharm Pharmacol. 61:207–212. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Margaritis EV, Yanni AE, Agrogiannis G,
Liarakos N, Pantopoulou A, Vlachos I, Papachristodoulou A,
Korkolopoulou P, Patsouris E, Kostakis M, Perrea DN and Kostakis A:
Effects of oral administration of l-arginine, l-NAME and
allopurinol on intestinal ischemia/reperfusion injury in rats. Life
Sci. 88:1070–1076. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Wang T, Leng YF and Zhang Y, Xue X, Kang
YQ and Zhang Y: Oxidative stress and hypoxia-induced factor 1alpha
expression in gastric ischemia. World J Gastroenterol.
17:1915–1922. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Yonezawa D, Sekiguchi F, Miyamoto M,
Taniguchi E, Honjo M, Masuko T, Nishikawa H and Kawabata A: A
protective role of hydrogen sulfide against oxidative stress in rat
gastric mucosal epithelium. Toxicology. 241:11–18. 2007. View Article : Google Scholar : PubMed/NCBI
|