Introduction
The landmark study of lin-4 in Caenorhabditis
elegans identified a novel class of molecules called microRNA
(miR), which are small non-coding RNAs consisting of 18–25
nucleotide base pairs (1) These
small nucleic acids regulate gene expression by binding to the 3′
untranslated region (3′-UTR) of mRNA, resulting in translational
repression or transcript degradation (2). Over 2,500 miRs have been identified in
the human genome since their discovery in 1993 and it has been
determined that 30–50% of genes that code for proteins are
controlled by miR in humans (3).
Thus, miRs have emerged as integral components of various
biological processes, including cell proliferation, migration,
differentiation, apoptosis and angiogenesis (4). It has been demonstrated that the
altered expression of miR is associated with tumorigenesis and the
progression of different types of cancer (5,6). By
regulating multiple potential target genes, miR expression may lead
to pathological changes in cells, ultimately contributing to the
development of cancer (7).
One particular family of miR, the miR-200 family,
has been identified to be crucial in tumorigenesis. Members of the
miR-200 family are downregulated in aggressive human tumors and
target different signaling pathways including the Notch, Wnt and
transforming growth factor β (TGF-β) pathways, thus inhibiting
migration, tumor cell adhesion, epithelial-to-mesenchymal
transition (EMT) and angiogenesis (8,9). The
present review will focus on summarizing the roles of the miR-200
family as putative tumor suppressors in tumor progression and
propose that the restoration of miR-200 expression may have
therapeutic implications for the clinical treatment of metastatic
and drug-resistant tumors.
Studies of miR-200 family in human
cancer
The miR-200 family consists of five members
(miR-200a, −200b, −200c, −141 and −429), which are clustered and
expressed as two separate polycistronic pri-miR transcripts
(miR-200b-200c-429 and miR-200a-141) located on human chromosomes 1
and 12, respectively (10).
miR-200b, −200c and −429 contain AAUACUG as their seed sequences,
whereas miR-200a and −141 possess AACACUG (11). Due to the similarity of their seed
sequences, they may have similar target genes (Table I). A series of studies using superior
profiling technologies, such as a combination of miRNA expression
arrays, quantitative polymerase chain reaction assays and mass
spectrometry DNA methylation analysis, have indicated that the
miR-200 family is aberrantly expressed in different types of
cancer. Multiple studies utilizing different detection platforms,
which compared extensive sets of tumor tissues and cell lines to
large sets of normal control samples, identified that the
differential expression of members of the miR-200 family serves an
important role in combating tumor cell invasion, EMT and metastasis
(8,12). Furthermore, numerous studies have
demonstrated that members of the miR-200 family may be associated
with pathogenesis and disease prognosis (12–28)
(Table II). Therefore, the miR-200
family has the potential to become a novel class of biomarker for
tumor prognosis and targets of novel drugs against tumor
progression.
 | Table I.Chromosomal localization and
sequences of the miR-200 family. |
Table I.
Chromosomal localization and
sequences of the miR-200 family.
| miR | Sequences
(5′-3′) | Chromosomal
localization |
|---|
| hsa-miR-200b |
UAAUACUGCCUGGUAAUGAUGAC | Chromosome
1p36 |
| hsa-miR-200c |
UAAUACUGCCGGGUAAUGAUGG | Chromosome
1p36 |
| hsa-miR-429 |
UAAUACUGUCUGGUAAAACCGU | Chromosome
1p36 |
| hsa-miR-200a |
UAACACUGUCUGGUAACGAUGU | Chromosome
12p13 |
| hsa-miR-141 |
UAACACUGUCUGGUAAAGAUGG | Chromosome
12p13 |
| Table II.Studies of miR-200 in various types
of human cancer. |
Table II.
Studies of miR-200 in various types
of human cancer.
| Cancer types | Samples | Conclusions made by
authors | (Refs.) |
|---|
| Bladder cancer | 57 bladder
tumors | miR-200c is
correlated with early stage | (14) |
|
| 11 normal bladder
urothelium | T1 bladder tumor
progression |
|
| Breast cancer | Breast cancer cell
lines | miR-200a and −200b
inhibit EMT characteristics | (15) |
|
| (MCF-7, T47D,
MDA-MB-231) | in the
undifferentiated, non-tumorigenic HC11 cells |
|
|
| HC11 mammary
epithelial cells |
|
|
| Colorectal
cancer | 54 primary
colorectal cancer tissues | miR-200c has an
important role in mediating EMT | (25) |
|
| 54 matched liver
metastasis tissues | and metastatic
behavior in the colon |
|
| Melanoma | 23 primary
melanomas | Decreased miR-200a,
−200c and −203 correlated | (26) |
|
|
| with increasing
thickness in melanomas |
|
| Endometrial
cancer | 17 endometrial
cancer tissues | miR-200 family is
highly expressed in endometrial | (16) |
|
| 11 normal
endometrial tissues | cancer and has an
important role in cancer growth |
|
| Gastric cancer | 35 gastric
neoplastic tissues | miR-141 may be
involved in the development of | (17) |
|
| 35 adjacent
non-neoplastic tissues | gastric cancer
through its inhibitory effect on cell proliferation |
|
|
| 5 gastric cancer
cell lines |
|
|
| Hepatocellular
cancer | 55 hepatocellular
cancer tissues | miR-200 was
deregulated in hepatocellular tumors | (18) |
|
| 30 nontumor liver
samples |
|
|
| Pancreatic
cancer | 99 pancreatic
cancer tissues | miR-200c may have a
role in pancreatic cancer biology | (27) |
|
| 14 pancreatic
cancer cell lines | and may be a novel
marker for the prognosis of pancreatic cancer |
|
| Anaplastic thyroid
carcinomas | 3 anaplastic
thyroid cancer tissues | Inhibition of TGF-β
receptor ١ in thyroid cancer cells induced EMT | (13) |
|
| 3 normal thyroid
sample | and caused an
increase of the miR-200 family |
|
| Head and neck
carcinoma | 45 spindle cell
carcinomas of head and neck | Downregulation of
miR-200 family supports the postulated role of | (12) |
|
| 45 squamous cell
carcinomas of head and neck | EMT in spindle cell
carcinoma of head and neck |
|
| Renal cell
carcinomas | 33 human renal
clear cell cancer tissues | Dysregulation of
miR-200/EMT confers to immortalized pre-tumoral | (24) |
|
| human epithelial
kidney cell line | cells phenotypic
traits of metastatic potential in renal cell carcinoma |
|
| Prostate
cancer | 18 prostate cancer
samples with no relapse | Overexpression of
miR-200a reduced prostate cancer cell growth | (23) |
|
| 18 matched-cancer
samples with relapse |
|
|
| Malignant pleural
mesothelioma | 100 mesothelioma
tumor samples | miR-200 was
downregulated in malignant pleural mesothelioma | (22) |
|
| 32 lung
adenocarcinoma tumor samples | compared to lung
adenocarcinoma |
|
|
| 4 healthy lung
tissue samples |
|
|
| Ovarian cancer | 20 serous ovarian
cancer tissues | Dysregulation of
miR-200 is involved in ovarian carcinogenesis | (21) |
|
| 8 normal ovarian
tissues | and associated with
the prognosis of serous ovarian carcinoma |
|
| OSCC | 25 OSCC cancer
tissues | The suppression of
miR-200 may drive tumor | (20) |
|
| 25 adjacent
non-cancer tissues | expansion and
progression in OSCC |
|
|
| 8 normal control
paired oral stroma and epithelium |
| NPC | C666-1
(EBV-positive NPC cell line) | miR-200a as a
regulatory factor of NPC carcinogenesis and a | (28) |
|
| CNE-1, CNE-2 and
HNE1 (NPC cell lines) | potential candidate
for miRNA-based therapy against NPC |
|
| NPC | C666-1
(EBV-positive NPC cell line) | miR-200a as a
regulatory factor of NPC carcinogenesis and a | (28) |
| Lung cancer | 130 lung squamous
cell carcinoma samples | miR-200 family was
functionally involved in canonical | (19) |
|
| Small airway
epithelial cells | pathways of immune
response, molecular mechanisms |
|
|
| Normal human
bronchial/tracheal epithelial cells | of cancer,
metastasis signaling, cell-cell communication, |
|
|
| H1299 (human
non-small cell lung cancer cell line) | proliferation and
DNA repair in lung cancer |
|
|
| BEAS-2B (human
immortalized lung epithelial cell line) |
|
|
miR-200 family and tumor metastasis
Different types of miR become deregulated in tumors
as a result of various mechanisms, including miR-200c deregulated
in ovarian cancer and miR-21 deregulated in breast cancer (29,30).
Genomic abnormalities, including deletion, amplification and
translocation, are common in tumorigenesis, and miR expression may
also be affected by transcriptional and post-transcriptional
regulation (7). Many pri-miR are
either oncogenes or tumor suppressors and are induced by
transcription factors (7). It has
been demonstrated that various transcription factors including p53,
c-Myc and E2 transcription factor (E2F), are closely associated
with miR in cancer (31). The
processing and stability of miR are also important factors
determining miR expression. Additionally, levels of Dicer or Drosha
expression, the miR processing machinery, are altered and
transformed in various types of cancer, possibly due to the
increase in the copy number of miR (32).
Metastasis is an important characteristic of
malignant tumors and EMT is the initial step of metastasis. The
miR-200 family inhibits EMT and maintains the epithelial phenotype
by directly targeting the transcriptional repressors of E-cadherin
[zinc finger E-box-binding homeobox (ZEB)1 and ZEB2] (33).
The primary step and important characteristics of
tumor metastasis are the disassembly of tight junctions and loss of
apical-basal polarity among cancer cells (8). The loss of epithelial markers and the
gain of mesenchymal morphological features in cancer cells
contributes to the suppression of the transmembrane adhesion
receptor E-cadherin and a gain in the expression of mesenchymal
markers, including vimentin, collagen, fibronectin, and the
E-cadherin transcriptional repressors ZEB1 and ZEB2 (also known as
SMAD-interacting protein 1) (9).
These vital molecules cause the extracellular matrix-induced
stimulation of the integrin signal pathway, resulting in focal
adhesion formation, which facilitates cancer cell migration,
invasion and metastasis (8). The
transcriptional factors ZEB1 and ZEB2 induce EMT by repressing the
expression of E-cadherin and promoting cancer cell migration,
invasion and metastasis (9). TGF-β
also serves an important role in the EMT in epithelial cells as it
commands cell proliferation and differentiation during the process
of embryonic development or cancer progression (34).
miR-200 family and tumor angiogenesis
It is widely accepted that angiogenesis, the
formation of new blood vessels from pre-existing ones, is a
fundamental process required for cancer development and growth
(26). Without angiogenesis, cancer
cells inside the tumor undergo apoptosis. The angiogenesis switch
depends on the balance of angiogenesis activators and inhibitors.
The activation of angiogenesis is initiated when pre-existing
vessels become permeable in response to stimulating factors,
including vascular endothelial growth factor (VEGF), placental
growth factor and angiopoietin-1 (35). VEGF is considered to be the most well
known fundamental factor and modulator of angiogenesis. VEGF
combined with its receptors [fms-like tyrosine kinase 1 (flt1) or
VEGF receptor (VEGFR)-1 and kinase-insert domain containing
receptor (KDR) or VEGFR-2], stimulates endothelial cell migration,
proliferation and survival (36). As
angiogenesis is essential for tumor growth, inhibiting VEGF
signaling using strategies such as small interfering RNA, small
molecule inhibitors, antibodies and VEGF-traps is a promising
therapeutic approach for cancer treatment. Furthermore, research
has demonstrated that miR-200 family members are involved in the
regulation of vascular development and angiogenesis by
downregulating VEGF signaling (37).
By using a clear cell renal cell carcinoma and matched normal
kidney sample, Liu et al (38) identified a negative correlation
between VEGF and the miR-200 family. Choi et al (39) demonstrated that VEGF and its
receptors, Flt1 and KDR, were the key participants in the VEGF
signal pathway and were negatively regulated by miR-200b, which
directly targeted the 3′-UTR of those genes. Tube formation and
phosphorylation of extracellular signal-regulated kinases 1/2 were
inhibited if miR-200b was transected into human umbilical vein
endothelial cells, suggesting that miR-200b has anti-angiogenic
activity (39).
Furthermore, it is understood that intratumoral
ligands establish a complex network of cell-cell interactions
within the tumor microenvironment. Roybal et al (40) determined that Flt1/VEGFR1 was a
candidate miR-200 target gene. Overexpression of miR-200 inhibited
angiogenesis in metastasis-prone lung adenocarcinoma cells derived
from K-ras/p53-mutant mice by inhibiting angiogenesis, thus
decreasing Flt1 levels (40). These
results indicate that lung adenocarcinomas with low levels of
miR-200 expression may be responsive to treatment with anti-VEGF
agents (40). In some chronic
non-cancer non-healing diseases, Chan et al (41) reported that hypoxia inhibited
miR-200b expression in human dermal microvascular endothelial
cells, thus promoting angiogenesis. Furthermore, it was identified
that downregulation of endothelial miR-200b is crucial in
stimulating cutaneous wound angiogenesis by attenuating the
repression of GATA binding protein 2 and VEGFR2 expression
(42). These results identified that
GATA2 was a target of miR-200b, which provided novel insight
regarding the regulation of wound angiogenesis by miR-200 and the
significance of such regulation in the context of chronic cutaneous
wounds (42). Additionally, it was
demonstrated that in lung, renal, ovarian and basal-like breast
cancer, miR-200 inhibits angiogenesis via direct and indirect
mechanisms by targeting interleukin-8 and C-X-C motif chemokine
ligand 1 secretion by tumor endothelial cells (43). Thus, the miR-200 family may provide a
potential anti-angiogenesis therapy to treat cancer and other
diseases dependent on angiogenesis, as inhibition of VEGF signaling
interferes with angiogenesis.
miR-200 family and chemosensitivity
Chemotherapy drug resistance often inhibits the
clinical treatment of cancer, resulting in a poor prognosis for
patients with cancer. MiR-200 family members are involved in
maintaining sensitivity to microtubule targeting agents and DNA
damaging drugs, which are two classes of chemotherapeutics
(44). Gibbons et al
(34) identified an association
between miR expression and cancer chemosensitivity in
cholangiocarcinoma. It has been demonstrated that miR-21 and −200b
are involved in the regulation of tumor cell sensitivity to
gemcitabine by targeting specific genes, including Circadian
Locomotor Output Cycles Kaput, phosphatase and tensin homolog and
protein-tyrosine phosphatase 1B, as well as downstream oncogene
products, including c-Abl, Src and Ras (34,45).
Furthermore, it has been suggested that there are similarities
between drug-resistant cancer cells and enhanced invasiveness or
metastasis, which is consistent with the dysregulation of miR-200
in drug resistant cancer cells (34,44,45). For
example, decreased expression of miR-200b and −200c has been
observed in the acquired cisplatin resistant phenotype of MCF-7
human breast adenocarcinoma cells (46). It was reported that aberrant miR
expression participated in the regulation of cell survival, cell
conductive signaling, invasiveness and DNA methylation, implying
that abnormal miR expression was associated with the unusual
activity of cellular processes in cisplatin-resistant breast cancer
cells (46). In a study
investigating miR expression profiles, the expression of miR-200b
was significantly downregulated in a docetaxel-resistant human
non-small cell lung cancer (NSCLC) cell line (SPC-A1/docetaxel)
compared with a control SPC-A1 cell line (47). The results suggested that measuring
miR-200b expression may provide an explanation for drug sensitivity
or resistance in docetaxel-resistant cells in patients with NSCLC
(47). Additionally, Chen et
al (48) demonstrated that
restoring expression of miR-200b was able to reverse chemoresistant
phenotypes of docetaxel-resistant cells in human lung
adenocarcinoma (LAD) by targeting E2F transcription factor 3 using
histone deacetylase (HDAC) inhibitors. These findings suggest that
the HDAC1/miR-200b/E2F3 signal pathway may be responsible for
chemoresistance in docetaxel-resistant LAD cells (48). Furthermore, members of the miR-200
family may be potential therapeutic targets for disseminated or
chemoresistant cancer cells in ovarian cancer (49).
miR-200 family members serve a vital role in
regulating cancer chemosensitivity. The association between miR-200
and cancer chemoresistance may be explained by a number of factors,
including EMT, cancer stem cell (CSC) maintenance, apoptosis,
angiogenesis and cell cycle distribution (Fig. 1). It has been demonstrated that
miR-200 family members are critical determinants in the EMT process
via targeting ZEB transcription factors (11,50,51).
Following this, the expression of clusters of genes, including
E-cadherin and vimentin, were controlled (11,50,51).
CSCs are defined as a small population of cells that possess the
capacity of self-renewal and generate differentiated progeny in
tumors. The acquisition of an EMT phenotype and the induction of
CSC phenotypes have biological functions in common and these
similarities synergistically contribute to cancer chemoresistance.
Thus, the properties of CSCs partly gave rise to EMT in cancer
metastasis.
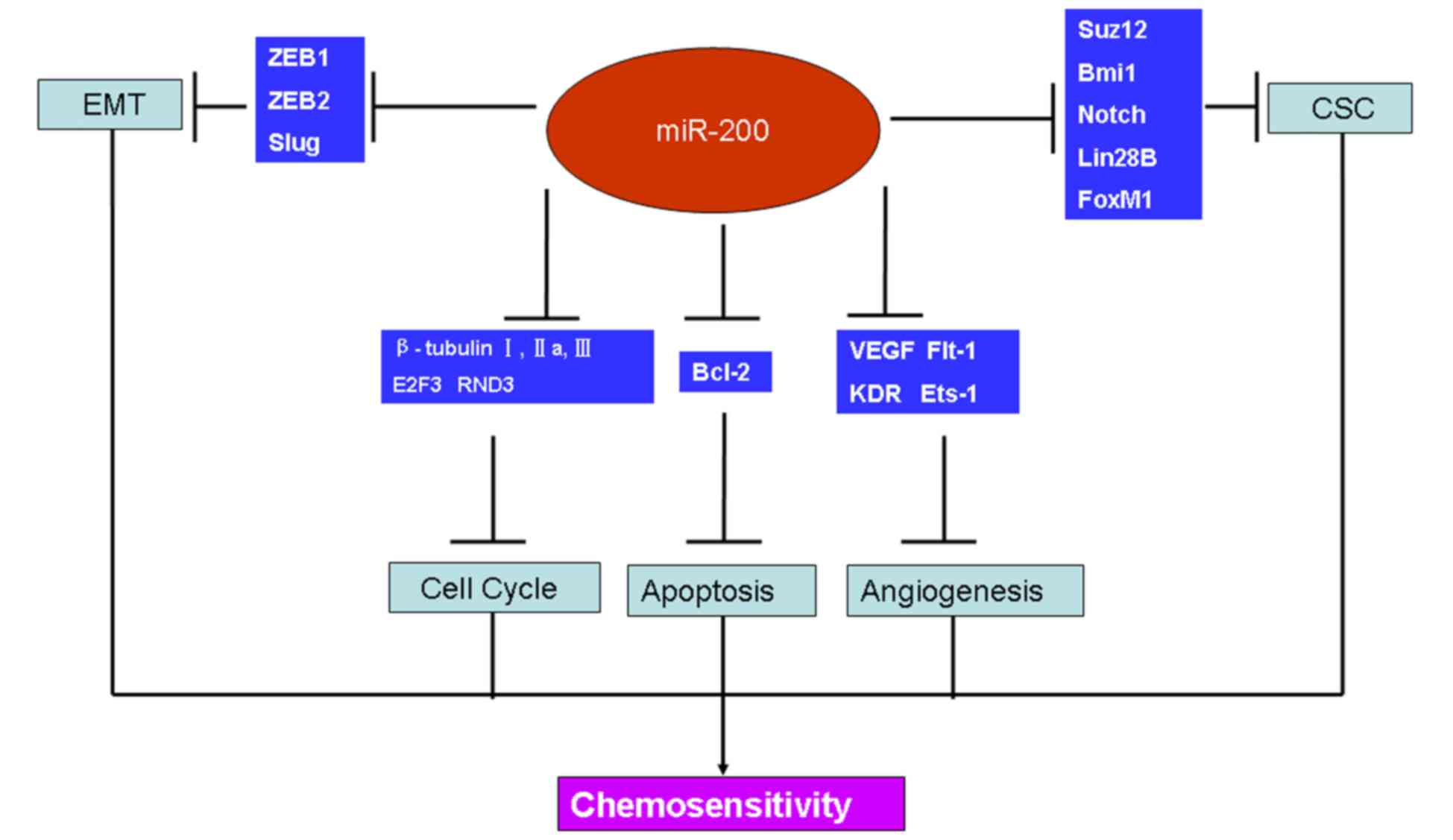 | Figure 1.Targets of miR-200 involved in cancer
chemoresistance. The verified targets of miR-200 are associated
with cancer chemosensitivity through EMT, the cell cycle,
apoptosis, angiogenesis and CSC maintenance. miR, microRNA; EMT,
epithelial-to-mesenchymal transition; CSC, cancer stem cell; ZEB,
zinc finger E-box-binding homeobox; E2F3, E2F transcription factor
3; RND3, Rho family GTPase 3; Bcl-2, B-cell lymphoma 2; VEGF,
vascular endothelial growth factor; Flt-1, fms-like tyrosine kinase
1; KDR, kinase-insert domain containing receptor. |
It is widely understood that angiogenesis is
essential for tumor progression. VEGF is a fundamental regulatory
molecule in tumor angiogenesis and VEGF function may be reduced by
the use of antiangiogenic agents to inhibit VEGFR, which means that
reducing VEGF function may result in a decrease in interstitial
fluid pressure and improved blood vessel morphology recovery
(52). Therefore, decreased tumor
blood supply combined with lower chemotherapy drug delivery may
reduce the sensitivity of cancer cells to chemotherapy, resulting
in the development of chemoresistance. The B-cell lymphoma 2
(Bcl-2) protein family is associated with multi-drug resistance
(53). One of the most important and
common features of tumor cells is resistance to apoptosis
signaling, which generates multi-drug resistance (53). The miR-200 family is able to reduce
resistance to apoptosis and drugs in cancer cells by targeting
Bcl-2 (54). Furthermore, Bcl-2
expression was reduced by miR-200 in gastric carcinoma, which led
to enhanced apoptosis (54).
Chemotherapy resistance is mediated by the cell cycle, and
sensitivity to drugs or agents in cancer treatment may be
attributed to the stage of the cell cycle in which the cells are
(55). The miR-200 family may affect
cell cycle distribution by targeting β-tubulin, E2F3 or Rho family
GTPase 3 (55,56).
Signals regulated by the miR-200 family in
cancer
The signaling of EMT is considered to be active
throughout tumorigenesis and tumor progression. It has been
demonstrated that overexpression of miR-200 inhibits the capacity
of tumor cells to undergo invasion and metastasis, highlighting the
role of miR-200 in the regulation of EMT and subsequent metastasis
(57). Notch signaling is a crucial
pathway involved in embryogenesis and is responsible for the
self-renewal abilities of embryonic stem cells (58). In various solid tumors, including
lung, pancreatic, breast carcinoma and malignant melanoma, aberrant
activation of Notch signaling is frequently observed (58). Mechanistic studies supporting that
Notch signaling serves a crucial role in regulating EMT and
metastasis during cancer progression have also identified that
miR-200 decreases expansion of human metastatic prostate cancer
cells by targeting the Notch ligand, jagged (JAG) 1, and the
mastermind-like transcriptional coactivators (Maml) 2 and 3
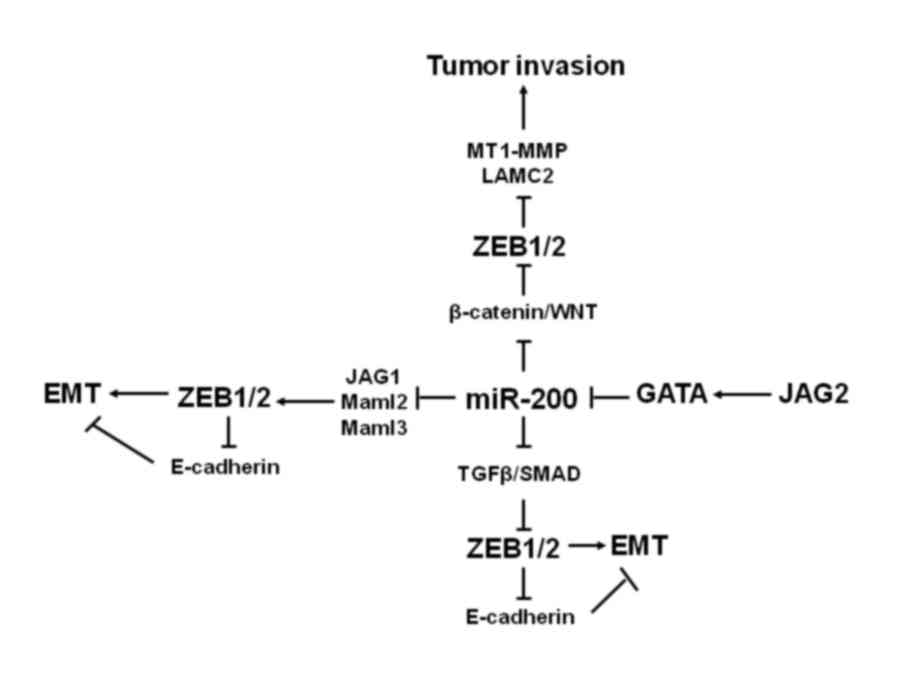
(58,59) Additionally, the Notch ligand JAG2 was
able to inhibit miR-200 family expression at the transcriptional
level by inducing GATA transcription factors, thus stimulating
tumor progression (60).
It has been determined that TGF-β, a profibrotic
cytokine, is the primary pathogenic driver in tubular epithelial
cells and is able to induce EMT (61). The pathway of TGF-β/SMAD signaling is
considered to be a classical pathway that induces EMT. This
inhibits tumor development at the early stage of cancer, yet
promotes tumor progression at the advanced stages of cancer
(61). miR-200 family members
suppress TGF-β/SMAD signaling, promote epithelial gene expression
and suppress cell invasion by regulating a network of genes
(61). In a study on gastric cancer,
miR-200 was downregulated by CpG island methylation and TGF-β
signaling, which increased Zeb1/2 expression and decreased
E-cadherin expression to promote cancer cell migration and
invasion, providing powerful evidence supporting the application of
decitabine in clinical cancer treatment as decitabine inhibits
methylation (62).
The complicated mutual transcriptional regulation of
feedback ZEB/miR-200 loops and TGF-β signaling is involved in the
interaction between ZEB and TGF-β protein (13,63). The
mechanisms of the autocrine TGF-β/ZEB/miR-200 loop signaling
regulatory network to control cell plasticity between the
epithelial and mesenchymal states may include: i) The regulation of
ZEB2 transcription induced by TGF-β by SMAD, which may be directly
targeted by the ZEB2 gene promoter in vivo (13,63) and
ii) the suppression of miR-200 loci was recovered by reversible DNA
methylation of ZEB proteins participating in the process of
recruitment of histone-modifying complexes (64).
Furthermore, it has been demonstrated that Wnt
signaling is functionally correlated with the ZEB/miR-200 loop in
tumors (65). A functional conserved
miR-200a-binding site is contained in the 3′ UTR of β-catenin mRNA,
meaning that miR-200a is able to directly target and bind to this
position to suppress the β-catenin/Wnt signaling pathway. This was
observed in tumorigenesis for human solid tumors, including
hepatocellular carcinoma, melanoma, colon, ovarian and prostate
cancer (66). Nuclear accumulation
of β-catenin is linked with EMT in invasive colorectal tumors, and
the association between the Wnt pathway and EMT has been
investigated (67). Levels of ZEB1
expression have no influence on β-catenin/transcription factor 4
transcriptional signaling in colon cancer cell lines, although ZEB1
immunopositivity has been confirmed in non-invasive colon cancer
cells and in tumor-associated fibroblasts (67). Furthermore, ZEB1 influenced the
expression of other key proteins mediated by the Wnt signaling
pathway, including membrane-type 1 matrix metalloproteinase and
laminin gamma chain 2 (LAMC2) (59).
Therefore, indicating that LAMC2 and all proteins mediated by the
Wnt signaling pathway were implicated in the invasion and
dissection of colorectal cancer cells (67). Additionally, in human meningioma
tumor tissues, the expression of miR-200a was negatively associated
with β-catenin and cyclin D1 (68).
The aforementioned signals regulated by members of the miR-200
family in cancer are summarized in Fig.
2.
Future directions and concerns
Although much insight has been gained regarding the
role of the miR-200 family in tumorigenesis and cancer progression,
there is still a long way to go to fully comprehend and take
advantage of miR utilizations in tumor therapeutics. Identifying
unique patterns of deregulated expression of miR-200 family members
may provide more significant information on the involvement of
miR-200 family members in cancer. For example, miR-200 family
members may act as molecular tumor markers for cancer initial
diagnosis, confirm the degree of risk for cancer patients and to
predict cancer prognosis and clinical responses to certain
therapeutic strategies.
Furthermore, at the mechanistic level, despite some
understanding of the miR-200/ZEB loop and signaling, further
studies are required to define the elusive role of the miR-200
family in cancer pathogenesis, particularly in appropriate cellular
and animal models. At the clinical level, further studies based on
statistically valid experimental designs and selection of highly
characterized case materials are required, which may identify the
appropriate tumor marker and novel therapeutic strategies for
improved cancer diagnosis and treatment.
In conclusion, tumor invasion and metastasis are
responsible for cases of carcinoma-associated mortality. It is
understood that EMT and tumor angiogenesis are critical steps in
tumor invasion and metastasis. Therefore, targeting these processes
may be a promising therapeutic strategy to treat cancer.
Encouragingly, the miR-200 family members are key regulators of the
epithelial phenotype, with targets involved in many aspects of EMT.
In the future, improving understanding of the regulation and
function of miR-200 family members in EMT, tumor angiogenesis and
metastatic processes may aid in the development of a more effective
method of attenuating cancer metastasis. Further understanding
regarding the role of miR-200 in cancer progression and the
development of more efficient miR regulatory molecules to treat
cancer may vastly improve the clinical treatment of tumors.
Acknowledgments
The present study was supported by the National
Natural Science Foundation (grant no. 81302250) and Tianjin Health
Bureau of Science and Technology Funds (grant no. 2012KZ073).
References
|
1
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sorek R and Cossart P: Prokaryotic
transcriptomics: A new view on regulation, physiology and
pathogenicity. Nat Rev Genet. 11:9–16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tomasetti M, Lee W, Santarelli L and
Neuzil J: Exosome-derived microRNAs in cancer metabolism: Possible
implications in cancer diagnostics and therapy. Exp Mol Med.
49:e2852017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paul P, Chakraborty A, Sarkar D, Langthasa
M, Rahman M, Bari M, Singha RS, Malakar AK and Chakraborty S:
Interplay between miRNAs and human diseases: A review. J Cell
Physiol. Feb 9–2017.(Epub ahead of print). View Article : Google Scholar
|
|
7
|
Meltzer PS: Cancer genomics: Small RNAs
with big impacts. Nature. 435:745–746. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang CJ, Chao CH, Xia W, Yang JY, Xiong
Y, Li CW, Yu WH, Rehman SK, Hsu JL, Lee HH, et al: p53 regulates
epithelial-mesenchymal transition and stem cell properties through
modulating miRNAs. Nat Cell Biol. 13:317–323. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim T, Veronese A, Pichiorri F, Lee TJ,
Jeon YJ, Volinia S, Pineau P, Marchio A, Palatini J, Suh SS, et al:
p53 regulates epithelial-mesenchymal transition through microRNAs
targeting ZEB1 and ZEB2. J Exp Med. 208:875–883. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Humphries B and Yang C: The microRNA-200
family: Small molecules with novel roles in cancer development,
progression and therapy. Oncotarget. 6:6472–6498. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zidar N, Boštjančič E, Gale N, Kojc N,
Poljak M, Glavač D and Cardesa A: Down-regulation of microRNAs of
the miR-200 family and miR-205, and an altered expression of
classic and desmosomal cadherins in spindle cell carcinoma of the
head and neck-hallmark of epithelial-mesenchymal transition. Hum
Pathol. 42:482–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Braun J, Hoang-Vu C, Dralle H and
Hüttelmaier S: Downregulation of microRNAs directs the EMT and
invasive potential of anaplastic thyroid carcinomas. Oncogene.
29:4237–4244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wiklund ED, Bramsen JB, Hulf T, Dyrskjøt
L, Ramanathan R, Hansen TB, Villadsen SB, Gao S, Ostenfeld MS,
Borre M, et al: Coordinated epigenetic repression of the miR-200
family and miR-205 in invasive bladder cancer. Int J Cancer.
128:1327–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aydoğdu E, Katchy A, Tsouko E, Lin CY,
Haldosén LA, Helguero L and Williams C: MicroRNA-regulated gene
networks during mammary cell differentiation are associated with
breast cancer. Carcinogenesis. 33:1502–1511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee JW, Park YA, Choi JJ, Lee YY, Kim CJ,
Choi C, Kim TJ, Lee NW, Kim BG and Bae DS: The expression of the
miRNA-200 family in endometrial endometrioid carcinoma. Gynecol
Oncol. 120:56–62. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Du Y, Xu Y, Ding L, Yao H, Yu H, Zhou T
and Si J: Down-regulation of miR-141 in gastric cancer and its
involvement in cell growth. J Gastroenterol. 44:556–561. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ladeiro Y, Couchy G, Balabaud C,
Bioulac-Sage P, Pelletier L, Rebouissou S and Zucman-Rossi J:
MicroRNA profiling in hepatocellular tumors is associated with
clinical features and oncogene/tumor suppressor gene mutations.
Hepatology. 47:1955–1963. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pacurari M, Addison JB, Bondalapati N, Wan
YW, Luo D, Qian Y, Castranova V, Ivanov AV and Guo NL: The
microRNA-200 family targets multiple non-small cell lung cancer
prognostic markers in H1299 cells and BEAS-2B cells. Int J Oncol.
43:548–560. 2013.PubMed/NCBI
|
|
20
|
Wiklund ED, Gao S, Hulf T, Sibbritt T,
Nair S, Costea DE, Villadsen SB, Bakholdt V, Bramsen JB, Sørensen
JA, et al: MicroRNA alterations and associated aberrant DNA
methylation patterns across multiple sample types in oral squamous
cell carcinoma. PloS One. 6:e278402011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nam EJ, Yoon H, Kim SW, Kim H, Kim YT, Kim
JH, Kim JW and Kim S: MicroRNA expression profiles in serous
ovarian carcinoma. Clin Cancer Res. 14:2690–2695. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gee GV, Koestler DC, Christensen BC,
Sugarbaker DJ, Ugolini D, Ivaldi GP, Resnick MB, Houseman EA,
Kelsey KT and Marsit CJ: Downregulated microRNAs in the
differential diagnosis of malignant pleural mesothelioma. Int J
Cancer. 127:2859–2869. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Barron N, Keenan J, Gammell P, Martinez
VG, Freeman A, Masters JR and Clynes M: Biochemical relapse
following radical prostatectomy and miR-200a levels in prostate
cancer. Prostate. 72:1193–1199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Castro-Vega LJ, Jouravleva K, Liu WY,
Martinez C, Gestraud P, Hupé P, Servant N, Albaud B, Gentien D, Gad
S, et al: Telomere crisis in kidney epithelial cells promotes the
acquisition of a microRNA signature retrieved in aggressive renal
cell carcinomas. Carcinogenesis. 34:1173–1180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hur K, Toiyama Y, Takahashi M, Balaguer F,
Nagasaka T, Koike J, Hemmi H, Koi M, Boland CR and Goel A:
MicroRNA-200c modulates epithelial-to-mesenchymal transition (EMT)
in human colorectal cancer metastasis. Gut. 62:1315–1326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
van Kempen LC, van den Hurk K, Lazar V,
Michiels S, Winnepenninckx V, Stas M, Spatz A and van den Oord JJ:
Loss of microRNA-200a and c, and microRNA-203 expression at the
invasive front of primary cutaneous melanoma is associated with
increased thickness and disease progression. Virchows Arch.
461:441–448. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu J, Ohuchida K, Mizumoto K, Sato N,
Kayashima T, Fujita H, Nakata K and Tanaka M: MicroRNA,
hsa-miR-200c, is an independent prognostic factor in pancreatic
cancer and its upregulation inhibits pancreatic cancer invasion but
increases cell proliferation. Mol Cancer. 9:1692010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia H, Ng SS, Jiang S, Cheung WK, Sze J,
Bian XW, Kung HF and Lin MC: miR-200a-mediated downregulation of
ZEB2 and CTNNB1 differentially inhibits nasopharyngeal carcinoma
cell growth, migration and invasion. Biochem Biophys Res Commun.
391:535–541. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pal MK, Jaiswar SP, Dwivedi VN, Tripathi
AK, Dwivedi A and Sankhwar P: MicroRNA: A new and promising
potential biomarker for diagnosis and prognosis of ovarian cancer.
Cancer Biol Med. 12:328–341. 2015.PubMed/NCBI
|
|
31
|
Azrak SS, Ginel-Picardo A, Drosten M,
Barbacid M and Santos E: Reversible, interrelated mRNA and miRNA
expression patterns in the transcriptome of Rasless fibroblasts:
Functional and mechanistic implications. BMC Genomics. 14:7312013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siomi H and Siomi MC: Posttranscriptional
regulation of microRNA biogenesis in animals. Mol Cell. 38:323–332.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Browne G, Sayan AE and Tulchinsky E: ZEB
proteins link cell motility with cell cycle control and cell
survival in cancer. Cell Cycle. 9:886–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gibbons DL, Lin W, Creighton CJ, Rizvi ZH,
Gregory PA, Goodall GJ, Thilaganathan N, Du L, Zhang Y,
Pertsemlidis A and Kurie JM: Contextual extracellular cues promote
tumor cell EMT and metastasis by regulating miR-200 family
expression. Genes Dev. 23:2140–2151. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pożarowska D and Pożarowski P: The era of
anti-vascular endothelial growth factor (VEGF) drugs in
ophthalmology, VEGF and anti-VEGF therapy. Cent Eur J Immunol.
41:311–316. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Welti J, Loges S, Dimmeler S and Carmeliet
P: Recent molecular discoveries in angiogenesis and antiangiogenic
therapies in cancer. J Clin Invest. 123:3190–3200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang HF, Xu LY and Li EM: A family of
pleiotropically acting microRNAs in cancer progression, miR-200:
Potential cancer therapeutic targets. Curr Pharm Des. 20:1896–1903.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu H, Brannon AR, Reddy AR, Alexe G,
Seiler MW, Arreola A, Oza JH, Yao M, Juan D, Liou LS, et al:
Identifying mRNA targets of microRNA dysregulated in cancer: With
application to clear cell renal cell carcinoma. BMC Syst Biol.
4:512010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Choi YC, Yoon S, Jeong Y, Yoon J and Baek
K: Regulation of vascular endothelial growth factor signaling by
miR-200b. Mol Cells. 32:77–82. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Roybal JD, Zang Y, Ahn YH, Yang Y, Gibbons
DL, Baird BN, Alvarez C, Thilaganathan N, Liu DD, Saintigny P, et
al: miR-200 inhibits lung adenocarcinoma cell invasion and
metastasis by targeting Flt1/VEGFR1. Mol Cancer Res. 9:25–35. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chan YC, Khanna S, Roy S and Sen CK:
miR-200b targets Ets-1 and is down-regulated by hypoxia to induce
angiogenic response of endothelial cells. J Biol Chem.
286:2047–20156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chan YC, Roy S, Khanna S and Sen CK:
Downregulation of endothelial microRNA-200b supports cutaneous
wound angiogenesis by desilencing GATA binding protein 2 and
vascular endothelial growth factor receptor 2. Arterioscler Thromb
Vasc Biol. 32:1372–1382. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pecot CV, Rupaimoole R, Yang D, Akbani R,
Ivan C, Lu C, Wu S, Han HD, Shah MY, Rodriguez-Aguayo C, et al:
Tumour angiogenesis regulation by the miR-200 family. Nat Commun.
4:24272013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Knezevic J, Pfefferle AD, Petrovic I,
Greene SB, Perou CM and Rosen JM: Expression of miR-200c in
claudin-low breast cancer alters stem cell functionality, enhances
chemosensitivity and reduces metastatic potential. Oncogene.
34:5997–6006. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rossi L, Bonmassar E and Faraoni I:
Modification of miR gene expression pattern in human colon cancer
cells following exposure to 5-fluorouracil in vitro. Pharmacol Res.
56:248–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pogribny IP, Filkowski JN, Tryndyak VP,
Golubov A, Shpyleva SI and Kovalchuk O: Alterations of microRNAs
and their targets are associated with acquired resistance of MCF-7
breast cancer cells to cisplatin. Int J Cancer. 127:1785–1794.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rui W, Bing F, Hai-Zhu S, Wei D and
Long-Bang C: Identification of microRNA profiles in
docetaxel-resistant human non-small cell lung carcinoma cells
(SPC-A1). J Cell Mol Med. 14:206–214. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen DQ, Pan BZ, Huang JY, Zhang K, Cui
SY, De W, Wang R and Chen LB: HDAC 1/4-mediated silencing of
microRNA-200b promotes chemoresistance in human lung adenocarcinoma
cells. Oncotarget. 5:3333–3349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kinose Y, Sawada K, Nakamura K and Kimura
T: The role of microRNAs in ovarian cancer. Biomed Res Int.
2014:2493932014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol Chem.
283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Winkler F, Kozin SV, Tong RT, Chae SS,
Booth MF, Garkavtsev I, Xu L, Hicklin DJ, Fukumura D, di Tomaso E,
et al: Kinetics of vascular normalization by VEGFR2 blockade
governs brain tumor response to radiation: Role of oxygenation,
angiopoietin-1, and matrix metalloproteinases. Cancer Cell.
6:553–563. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Thomas S, Quinn BA, Das SK, Dash R, Emdad
L, Dasgupta S, Wang XY, Dent P, Reed JC, Pellecchia M, et al:
Targeting the Bcl-2 family for cancer therapy. Expert Opin Ther
Targets. 17:61–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kurashige J, Kamohara H, Watanabe M,
Hiyoshi Y, Iwatsuki M, Tanaka Y, Kinoshita K, Saito S, Baba Y and
Baba H: MicroRNA-200b regulates cell proliferation, invasion, and
migration by directly targeting ZEB2 in gastric carcinoma. Ann Surg
Oncol. 19 Suppl 3:S656–S664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Leskelä S, Leandro-García LJ, Mendiola M,
Barriuso J, Inglada-Pérez L, Muñoz I, Martínez-Delgado B, Redondo
A, De Santiago J, Robledo M, et al: The miR-200 family controls
beta-tubulin III expression and is associated with paclitaxel-based
treatment response and progression-free survival in ovarian cancer
patients. Endocr Relat Cancer. 18:85–95. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Xia W, Li J, Chen L, Huang B, Li S, Yang
G, Ding H, Wang F, Liu N, Zhao Q, et al: MicroRNA-200b regulates
cyclin D1 expression and promotes S-phase entry by targeting RND3
in HeLa cells. Mol Cell Biochem. 344:261–266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Koutsaki M, Spandidos DA and Zaravinos A:
Epithelial-mesenchymal transition-associated miRNAs in ovarian
carcinoma, with highlight on the miR-200 family: Prognostic value
and prospective role in ovarian cancer therapeutics. Cancer Lett.
351:173–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Feng X, Wang Z, Fillmore R and Xi Y:
MiR-200, a new star miRNA in human cancer. Cancer Lett.
344:166–173. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hill L, Browne G and Tulchinsky E:
ZEB/miR-200 feedback loop: At the crossroads of signal transduction
in cancer. Int J Cancer. 132:745–754. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang Y, Ahn YH, Gibbons DL, Zang Y, Lin W,
Thilaganathan N, Alvarez CA, Moreira DC, Creighton CJ, Gregory PA,
et al: The Notch ligand Jagged2 promotes lung adenocarcinoma
metastasis through a miR-200-dependent pathway in mice. J Clin
Invest. 121:1373–1385. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ahn SM, Cha JY, Kim J, Kim D, Trang HT,
Kim YM, Cho YH, Park D and Hong S: Smad3 regulates E-cadherin via
miRNA-200 pathway. Oncogene. 31:3051–3059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhou X, Wang Y, Shan B, Han J, Zhu H, Lv
Y, Fan X, Sang M, Liu XD and Liu W: The downregulation of
miR-200c/141 promotes ZEB1/2 expression and gastric cancer
progression. Med Oncol. 32:4282015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Argast GM, Krueger JS, Thomson S,
Sujka-Kwok I, Carey K, Silva S, O'Connor M, Mercado P, Mulford IJ,
Young GD, et al: Inducible expression of TGFβ, snail and Zeb1
recapitulates EMT in vitro and in vivo in a NSCLC model. Clin Exp
Metastasis. 28:593–614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Brabletz S, Bajdak K, Meidhof S, Burk U,
Niedermann G, Firat E, Wellner U, Dimmler A, Faller G, Schubert J
and Brabletz T: The ZEB1/miR-200 feedback loop controls Notch
signalling in cancer cells. EMBO J. 30:770–782. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sánchez-Tilló E, de Barrios O, Valls E,
Darling DS, Castells A and Postigo A: ZEB1 and TCF4 reciprocally
modulate their transcriptional activities to regulate Wnt target
gene expression. Oncogene. 34:5760–5770. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zaravinos A: The regulatory role of
microRNAs in EMT and cancer. J Oncol. 2015:8658162015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kahlert C, Lahes S, Radhakrishnan P, Dutta
S, Mogler C, Herpel E, Brand K, Steinert G, Schneider M,
Mollenhauer M, et al: Overexpression of ZEB2 at the invasion front
of colorectal cancer is an independent prognostic marker and
regulates tumor invasion in vitro. Clin Cancer Res. 17:7654–7663.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Saydam O, Shen Y, Würdinger T, Senol O,
Boke E, James MF, Tannous BA, Stemmer-Rachamimov AO, Yi M, Stephens
RM, et al: Downregulated microRNA-200a in meningiomas promotes
tumor growth by reducing E-cadherin and activating the
Wnt/beta-catenin signaling pathway. Mol Cell Biol. 29:5923–5940.
2009. View Article : Google Scholar : PubMed/NCBI
|