Introduction
Hair is considered to be an accessory structure of
the integumentary system, along with sebaceous glands, sweat glands
and nails. Hair growth is cyclic, involving a growth (anagen)
phase, an apoptosis-mediated regression (catagen) phase and a
resting (telogen) phase (1,2). Hair loss or alopecia is becoming more
prevalent worldwide (3). Although
hair loss is generally not life-threatening, it has been shown to
have a substantial negative impact on social interactions and
psychological well-being (4). To
date, most marketed anti-hair-loss products are ineffective.
Finasteride and minoxidil are the only two products approved for
hair loss by the US Food and Drug Administration. However, their
use is limited owing to unpredictable efficacy and undesirable side
effects (5). Therefore, it is
urgently necessary to develop novel pharmacological treatments.
Various cytokines and growth factors contribute to
the regulation of hair morphogenesis and the hair growth cycle.
Chemokine (C-X-C motif) ligand 4 (CXCL4; also known as platelet
factor-4) is a member of the CXC chemokine family of small (8–10
kDa) proteins (6). CXCL4 was first
described by Deutsch et al in 1955 and was demonstrated to
be a platelet protein with anti-heparin activity (7). CXCL4 is a 7.8-kDa protein comprising 70
amino acids, that is synthesized in megakaryocytes, expressed in
other cells, and stored in α-granules (8). CXCL4 has been reported to have numerous
biological effects, including immunization, apoptosis, cell
differentiation, survival, proliferation and tissue repair effects
(9). CXCL4 inhibits the spontaneous
apoptosis of monocytes and mediates their differentiation into a
specific subtype of macrophages (9,10). It
also promotes the survival of hematopoietic stem cells and
progenitor cells (11). Moreover,
CXCL4 has been reported to have an antiproliferative effect on
endothelial cells and fibroblasts, in addition to anti-angiogenic
activity (12–15). In addition, CXCL4 participates in
mediation of the cell recruitment and activation necessary for
inflammation and the repair of tissue damage (6). However, to the best of our knowledge,
the role of CXCL4 in the hair growth cycle has not yet been
reported.
Hair shafts are made by the hair follicle, which
undergoes self-renewal when hairs are damaged. The present study is
based on the proposal that homeostatically regulated gene
expression during the hair cycle is essential for hair growth. In
order to identify those genes, a genome-wide gene expression array
using a depilation-induced hair growth mouse model was
performed.
The aim of the present study was to determine
whether CXCL4 is an important gene in hair regeneration.
Previously, the present authors identified that CXCL4 induced
apoptosis of the IEC-6 intestinal epithelial cell line, and that
CXCL4 monoclonal antibody (mAb) reduced the apoptosis of the crypt
epithelia in a 5-fluorouracil-induced mucositis model (16). Therefore, it was hypothesized that
CXCL4 may retard hair growth by exerting an anti-proliferative
effect on hair follicle cells, whereas CXCL4 mAb may promote hair
growth by stimulating follicular proliferation and delaying the
catagen phase. The present study demonstrated that the expression
of CXCL4 was downregulated following the transition from the
telogen to the anagen phase, and returned to the normal level
following the next telogen phase. CXCL4 mAb significantly promoted
the initiation of the hair follicle transition from the telogen to
the anagen phase through upregulation of hair growth-related genes
in vivo. These results indicated that CXCL4 plays an
important role in hair growth.
Materials and methods
Reagents
Antibodies against Bcl-2 (3498), Bax (2772) and the
cleaved form of caspase-3 (9664) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Antibodies against
proliferating cell nuclear antigen (PCNA) (sc-25280) and β-actin
(sc-47778) were from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). Rat immunoglobulin G (IgG) was purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). CXCL4 mAb was produced in the
laboratory, as previously described (16).
Experimental animals
All experiments with mice were conducted in
accordance with the Guide for the Care and Use of Laboratory
Animals (National Academies Press, Washington, DC, USA, 1996) and
were approved by the Animal Research Committee of Shanghai Jiaotong
University (Shanghai, China). Male C57BL/6 mice were purchased from
Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China). The
6-week-old mice (17–20 g) were allowed to adapt to their new
environment for 1 week. Mice were housed in standard animal rooms
with food and water available ad libitum under controlled humidity
(50±15%) and temperature (22±2°C). The room was illuminated by
fluorescent lights that were on from 8:00 a.m. to 8:00 p.m.
Microarray analysis
Microarray analysis was conducted by Shanghai
Biotechnology Corporation (Shanghai, China). Total RNA was
extracted using TRIzol reagent (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) from excised C57BL/6 mouse back skin, followed by
purification, using RNeasy Mini kit (Qiagen GmBH, Hilden, Germany)
and RNase-Free DNase set (Qiagen GmBH). Total RNA was amplified and
labeled using a Low Input Quick Amp Labeling kit, One-Color
(Agilent Technologies, Inc., Santa Clara, CA, USA). Labeled cRNA
was hybridized with Mouse Genome Microarray 4*44K (Agilent
Technologies, Inc.) and washed according to the manufacturer's
recommendations. Slides were scanned using an Agilent Microarray
Scanner (G2565CA; Agilent Technologies, Inc.). The signals were
analyzed using Feature Extraction Software 10.7 (Agilent
Technologies, Inc.), excluding the data from signals that were
recognized as outlier or equal to background. Raw data were
normalized by Quantile algorithm, Gene Spring Software 11.0
(Agilent Technologies, Inc.).
Experimental analysis using CXCL4
mAb
Twenty C57BL/6 mice in two randomized groups (n=10)
were used for studying the activity of CXCL4 mAb. The anagen phase
was induced by depilation on the dorsal skin of the animals at 7
weeks old. This was performed by applying a melted wax/rosin
mixture to the dorsal skin and peeling off this mixture after
hardening, as previously described (17). All depilated mouse hair follicles
were synchronized in the telogen stage. The animals in the test
group were subcutaneously injected with CXCL4 mAb (1 mg/kg body
weight) once a week for three consecutive weeks, and the control
group were subcutaneously injected with rat isotype IgG (1 mg/kg
body weight) once a week for three consecutive weeks. The dorsal
skin of the mice was observed and photographic images were captured
on 0, 7, 10 and 18 days after depilation. Three mice from each time
point were sacrificed to obtain skin specimens. Briefly, after
being euthanized, mice were shaved with mechanical clippers and
total, fully-thickness dorsal skin was dissected free. The skin was
frozen quickly in liquid nitrogen prior to analysis.
Histological preparation
Dorsal skin of the mice was fixed with 10% neutral
buffered formalin. The tissues were dehydrated through an ascending
series of graded ethanol, cleared in xylene, and embedded in
paraffin blocks. Subsequently, samples were cut either
longitudinally or transversely into 5-µm sections. These sections
were stained with hematoxylin and eosin (H&E), and the
histological morphology was examined using light microscopy.
Hair follicle counting
Digital photomicrographs were taken from
representative areas of slides at a fixed magnification of ×40. All
images were cropped in a fixed area with a width of 1,500 µm. The
hair follicles in the deep subcutis were counted manually, as
previously described (5).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the skin using two-step
RNA extraction with TRIzol (Invitrogen; Thermo Fisher Scientific,
Inc.). Reverse transcription (RT) reaction was performed with RNA
(2 µg) at 37°C for 15 min and 85°C for 5 sec according to the
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China). The product was amplified in a reaction volume of 20 µl
containing 9.2 µl RT product, 10 µl SYBR Premix Ex Taq II (2x)
(Takara Biotechnology Co., Ltd.), and 20 pmol of each primer. PCRs
were performed for 40 cycles at 95°C for 10 sec, 60°C for 30 sec
and 72°C for 30 sec using a 7900HT Fast Real Time RT-PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). Three
replicates were performed at each time point. All data were
normalized to β-actin mRNA levels, and the fold change for each
mRNA was calculated using the 2−∆∆Cq method (18). The primer sequences used are shown in
Table I.
 | Table I.Sequences of forward and reverse
primers for reverse transcription-quantitative polymerase chain
reaction. |
Table I.
Sequences of forward and reverse
primers for reverse transcription-quantitative polymerase chain
reaction.
| Genes | Primer | Sequences
(5′-3′) | Product size
(bp) | GenBank accession
no. |
|---|
| β-catenin | Forward |
GTCAGCTCGTGTCCTGTGAA | 148 | NM_007614.3 |
|
| Reverse |
GTGCGCTGAGCTTCAGGT |
|
|
| Ccnd2 | Forward |
CTGAGTCTGGTTGGTGCTGA | 239 | NM_009829.3 |
|
| Reverse |
ACACCCGAGACCACAGAAAC |
|
|
| Lef1 | Forward |
GCCACCGATGAGATGATCCC | 107 | NM_010703 |
|
| Reverse |
TTGATGTCGGCTAAGTCGCC |
|
|
| Wnt10b | Forward |
TCTTGGATTCCAGGGAGGCT | 120 | NM_011718.2 |
|
| Reverse |
ACGTTCCATGGCATTTGCAC |
|
|
| Wnt5a | Forward |
GCTTCGCTTGAATTCCTCGG | 143 | NM_009524.3 |
|
| Reverse |
ACTTGGAAGACATGGCACCT |
|
|
| Bmp4 | Forward |
TGAGCCTTTCCAGCAAGTTT | 180 | NM_007554.2 |
|
| Reverse |
CTTCCCGGTCTCAGGTATCA |
|
|
| Bmp2 | Forward |
TGGAAGTGGCCCATTTAGAG | 166 | NM_007553.3 |
|
| Reverse |
TGACGCTTTTCTCGTTTGTG |
|
|
| Gli1 | Forward |
CCAAGCCAACTTTATGTCAGGG | 130 | NM_010296 |
|
| Reverse |
AGCCCGCTTCTTTGTTAATTTGA |
|
|
| Vcan | Forward |
TTTTACCCGAGTTACCAGACTCA | 106 | NM_001081249 |
|
| Reverse |
GGAGTAGTTGTTACATCCGTTGC |
|
|
| β-actin | Forward |
AGCCTTCCTTCTTGGGTATG | 103 | NM_007393 |
|
| Reverse |
GTGTTGGCATAGAGGTCTTTAC |
|
|
Western blot analysis
Skin tissue (100 mg) was homogenized in 1 ml
radioimmunoprecipitation assay lysis buffer with protease and
phosphatase inhibitors (Beyotime Institute of Biotechnology,
Haimen, China). Supernatants were recovered after 12,000 × g
centrifugation at 4°C for 10 min, and their protein content was
determined using a bicinchoninic acid assay kit (P001; Beyotime
Institute of Biotechnology). Proteins (30 µg) were separated using
12% SDS-PAGE gels, and transferred to a polyvinylidene difluoride
membrane. Membranes were blocked in 5% non-fat dry milk for 2 h at
room temperature and incubated with anti-PCNA (1:1,000 dilution,
sc-25280, Santa Cruz Biotechnology, Inc.), anti-cleaved caspase-3
(1:1,000 dilution, 9664, Cell Signaling Technology, Inc.),
anti-Bcl2 (1:1,000 dilution, 3498, Cell Signaling Technology,
Inc.), anti-Bax (1:1,000 dilution, 2772, Cell Signaling Technology,
Inc.), and anti-β-actin (1:1,000 dilution, sc-47778, Santa Cruz
Biotechnology, Inc.) primary antibodies overnight at 4°C and then
with the corresponding HRP-conjugated secondary antibodies
(1:5,000, 7044 and 7066, Cell Signaling Technology, Inc.) for 2 h
at room temperature. The membranes were washed and visualized using
enhanced chemiluminescence reagents (Thermo Fisher Scientific,
Inc.) and analyzed using a gel imaging system (Tanon 3500; Tanon
Science and Technology Co., Ltd., Shanghai, China). β-actin was
employed as a loading control. Three replicates were performed for
each treatment.
Statistical analysis
All data are presented as the mean ± standard
deviation from three independent experiments. All statistical
analysis was performed using SPSS 18.0 (SPSS, Inc., Chicago, IL,
USA). Differences between experimental groups were evaluated by
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of CXCL4 during the induced
mouse hair cycle
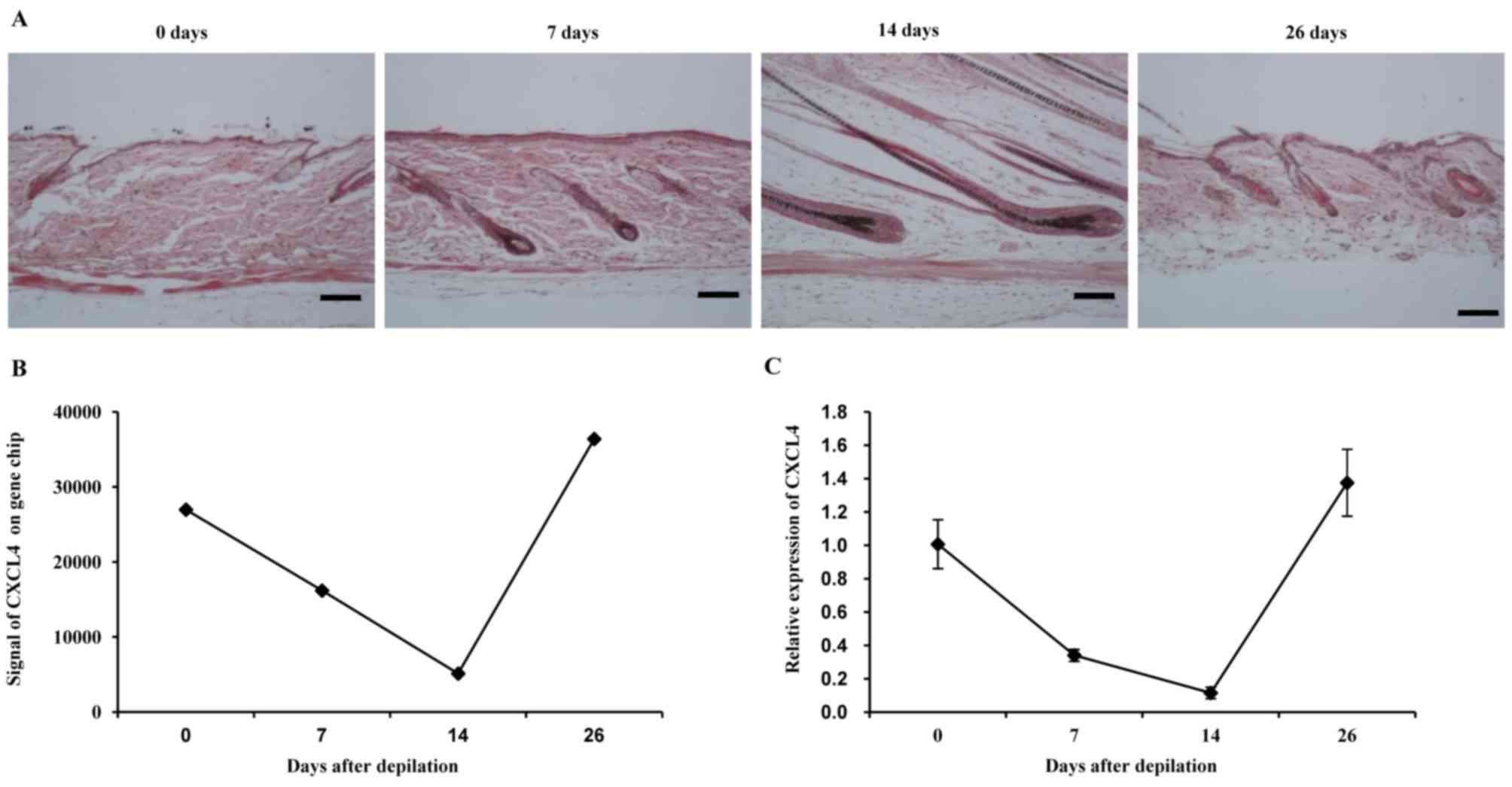
In the mouse model used in the present study, a new
cycle of hair growth was induced by depilation, as previously
described (17). The anagen phase
was fully synchronized over the entire area of depilation. At 0 and
26 days, typical telogen morphology, including the entire hair
follicle residing in the dermis and lacking melanin, was observed
in the hair matrix above the dermal papilla. At 7 and 14 days,
enlarged dermal papillae were observed. In addition, elongated hair
shafts emerged (Fig. 1A). Gene
expression of the total dorsal skin taken from mice at several time
points following depilation (7, 14 and 26 days) was analyzed by DNA
microarray hybridization and compared with skin samples taken at
the telogen phase (0 days). The expression of CXCL4 was identified
to be decreased during the anagen phase (0–14 days) and returned to
the normal level during the telogen phase (26 days; Fig. 1B). The homeostatic regulation of
CXCL4 shown in the expression array was confirmed by RT-qPCR
(Fig. 1C). The expression pattern of
CXCL4 in the hair cycle suggests its participation in hair growth
following depilation.
CXCL4 mAb promotes hair
regeneration
The homeostatic regulation of CXCL4 in the skin
strongly suggests its role in hair growth. A strategy of using a
neutralizing mAb to block CXCL4 was used to evaluate the role of
CXCL4 in hair growth. Adult C57BL/6 mice are useful for testing
hair growth-promoting reagents, as their truncal pigmentation is
dependent on their follicle melanocytes, which produce pigment only
during anagen (19). It has been
reported that the skin color of mice is pink in the telogen phase,
and gradually becomes grey and black along with anagen initiation
(20). Thus, black pigmentation is
taken as evidence of the transition of hair follicles from the
telogen to the anagen phase.
To evaluate the hair growth activity of CXCL4 mAb,
the depilated mice were treated with CXCL4 mAb once a week, for 3
weeks. Each day, the degree of hair growth was evaluated by
observing the skin color. Darker skin was observed in the CXCL4
mAb-treated group at 7 and 10 days, compared with that of the
control mice treated with isotype IgG (Fig. 2). These results indicate that CXCL4
mAb significantly stimulated hair growth.
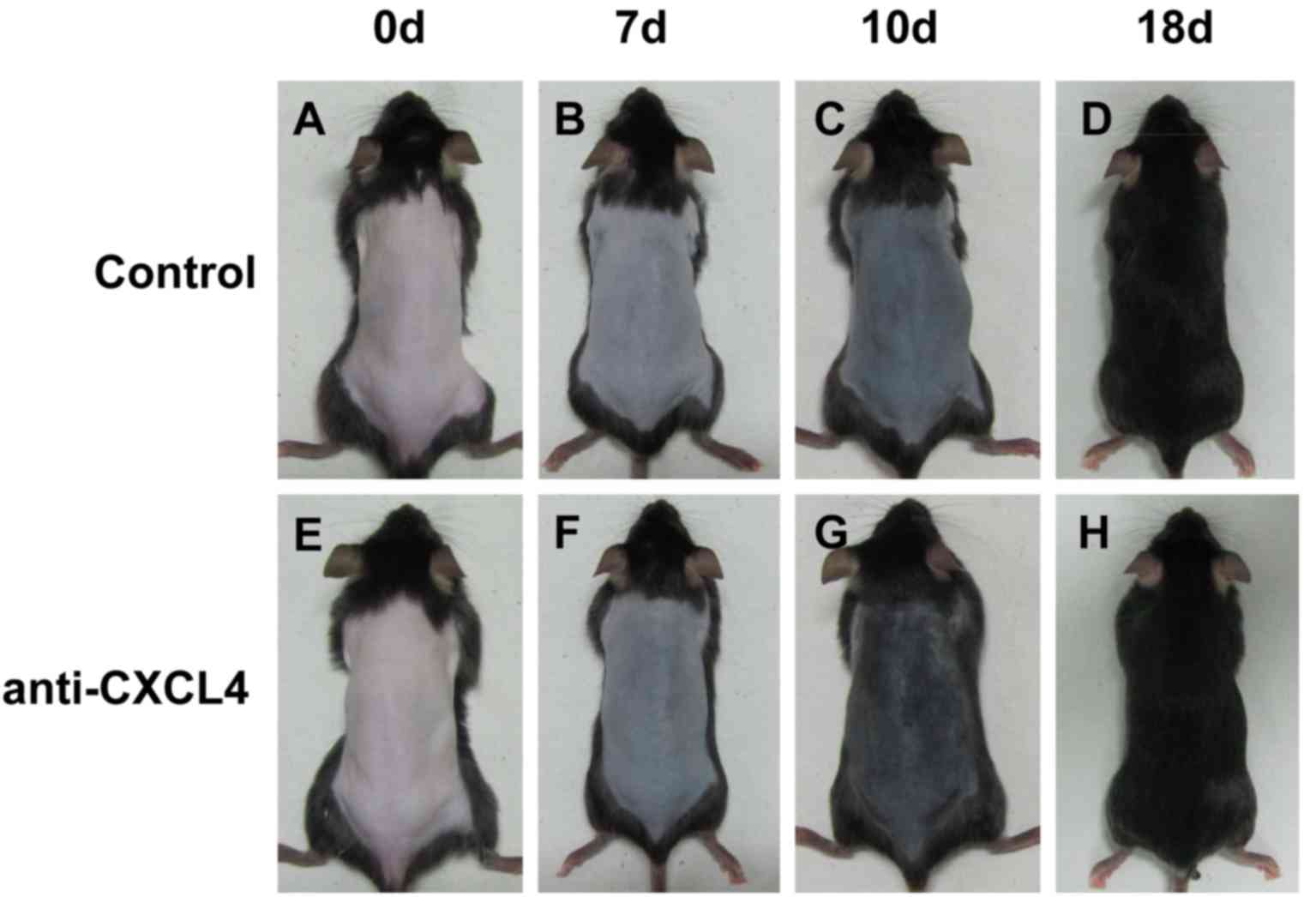 | Figure 2.Anti-CXCL4 mAb promotes hair
regeneration. Telogen-matched, 7-week-old C57BL/6 mice were
depilated and subcutaneously injected with CXCL4 mAb (n=10) or IgG
as the control (n=10) once weekly for 3 weeks. Photographic images
were captured at 0, 7, 10 and 18 days. Darker skin was observed in
the CXCL4 mAb-treated mice compared with the control mice treated
with isotype IgG. Control mice at (A) 0 days, (B) 7 days, (C) 10
days and (D) 18 days, and mice treated with CXCL4 antibody at (E) 0
days, (F) 7 days (G) 10 days and (H) 18 days. CXCL4, chemokine
(C-X-C motif) ligand 4; mAb, monoclonal antibody; IgG,
immunoglobulin G. |
Effects of CXCL4 mAb on the
development and structure of mouse hair follicles
An increase in the number of hair follicles has been
considered as an indicator of the transition of hair growth from
the telogen to the anagen phase. The anagen phase is associated
with an increase in the number of hair follicles that lie in the
deep subcutis compared with that of the telogen phase, in which
hair follicles lie in the dermis only (20). The effect of CXCL4 mAb on hair
follicle density was further assessed via histological analysis of
mouse skin samples following staining with H&E (Fig. 3A-F). CXCL4 mAb significantly
increased the number of hair follicles in mice, compared with that
in the control group at 10 and 18 days (P<0.05; Fig. 3G).
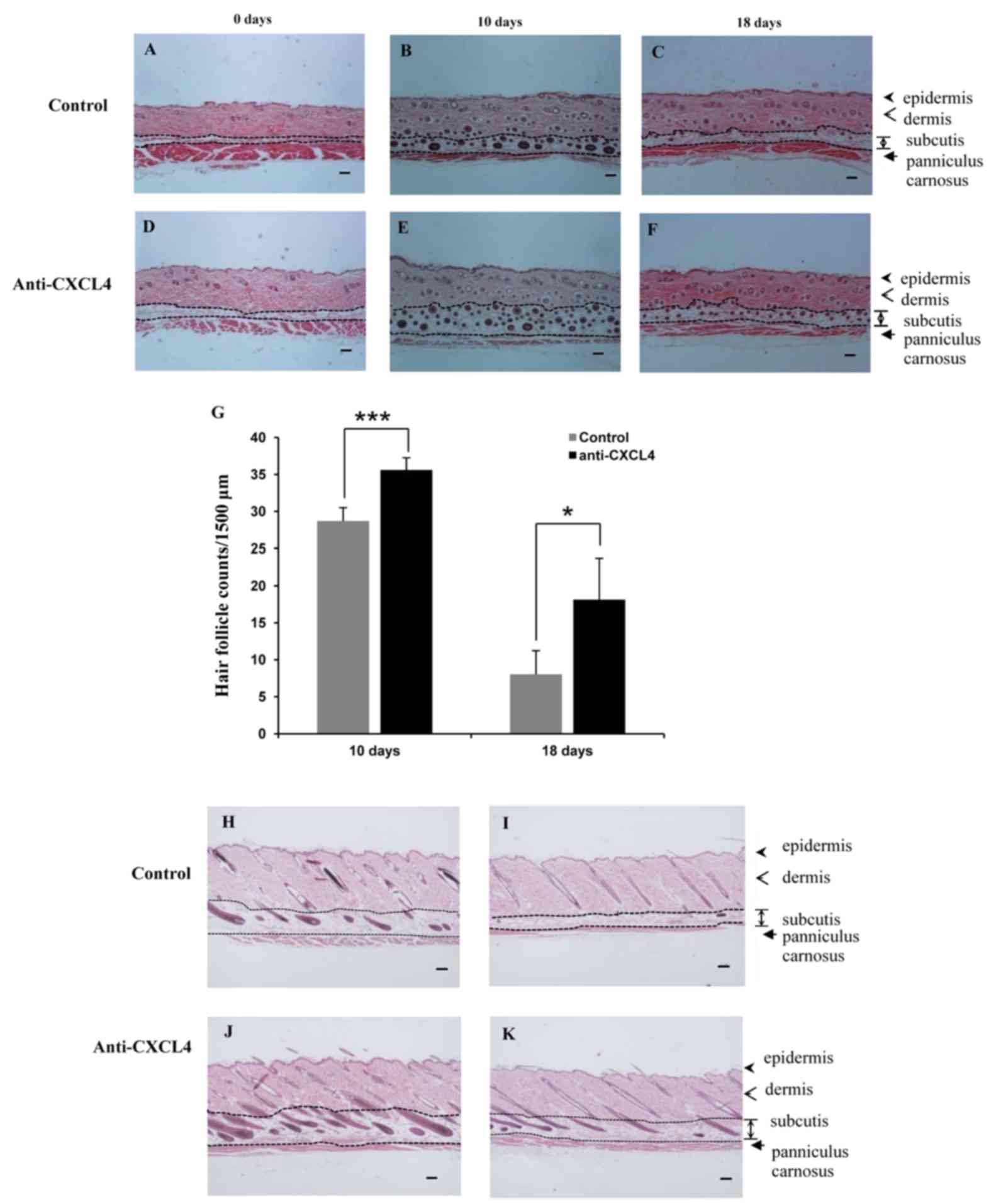 | Figure 3.Hair follicles in the telogen-matched
C57BL/6 mice treated with CXCL4 mAb. The effect of CXCL4 mAb on the
hair follicles was analyzed using hematoxylin and eosin staining.
(A-F) Transverse sections of the dorsal skin at 0, 10 and 18 days
were stained, and representative photomicrographs of skin sections
are shown. Scale bar, 100 µm. Control mice at (A) 0 days, (B) 10
days and (C) 18 days, and mice treated with CXCL4 antibody at (D) 0
days, (E) 10 days and (F) 18 days. (G) The number of hair follicles
in the deep subcutis in five fields per mouse was determined under
a microscope (magnification, ×40). Values are the mean ± standard
deviation (n=5 mice per group); *P<0.05 and ***P<0.001 as
indicated. (H-K) Longitudinal sections of the dorsal skin were
stained, and the images shown are representative of 5 mice. Scale
bar, 100 µm. Control group at (H) 10 days and (I) 18 days and mice
treated with CXCL4 mAb at (J) 10 days and (K) 18 days after
depilation. CXCL4, chemokine (C-X-C motif) ligand 4; mAb,
monoclonal antibody. |
In accordance with the classification of hair cycle
stages in mice (1), hair follicles
at 18 days following depilation are in the catagen phase.
Longitudinal sections of dorsal skins are shown in Fig. 3H-K. It was observed that regression
of the hair follicles pulled the dermal papillae upward into the
dermis in the control group (Fig.
3I). By contrast, almost all of the hair follicles in the CXCL4
mAb-treated group continued to reside in the subcutis (Fig. 3K). These observations indicate that
blockade of CXCL4 induced an earlier anagen phase and prolonged the
mature anagen phase, compared with that in the control group.
CXCL4 mAb upregulates Wnt- and bone
morphogenetic protein (Bmp)-dependent signaling
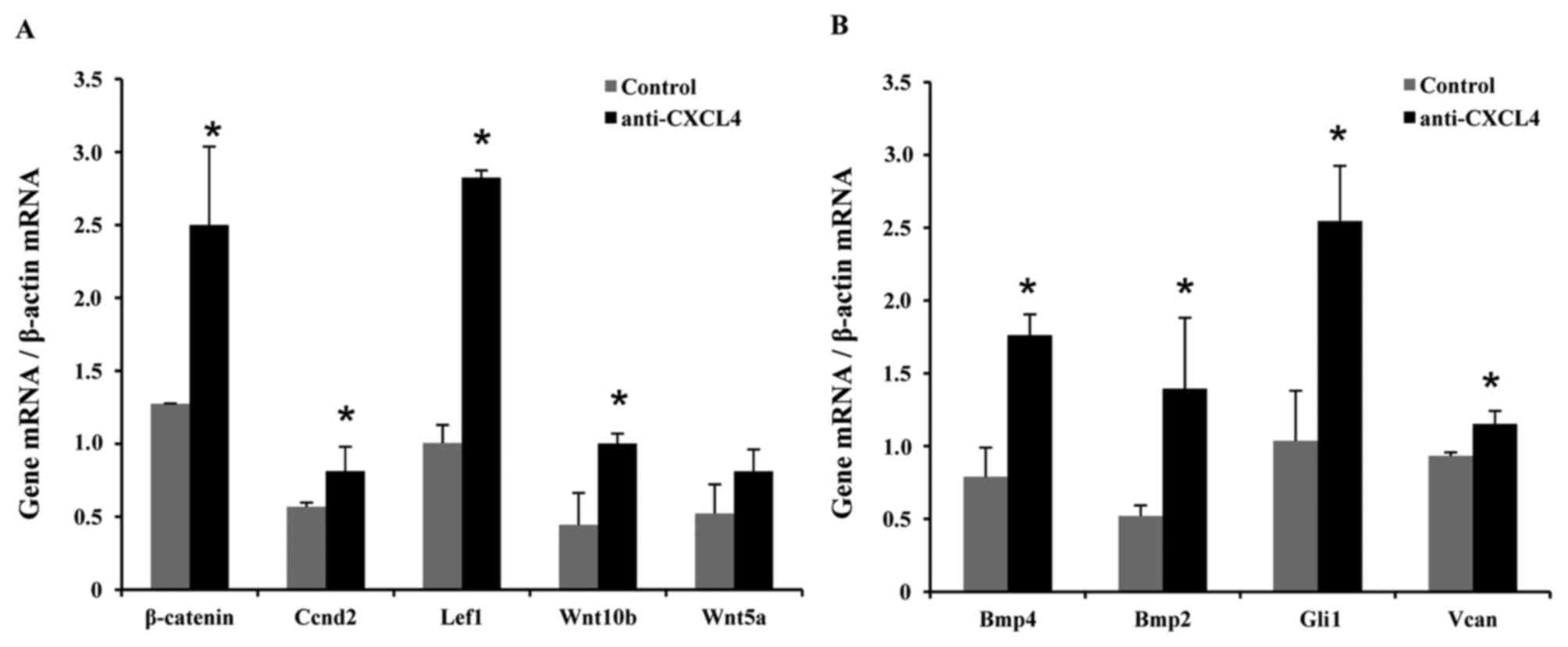
Previous studies demonstrated that transient
activation of Wnt-dependent β-catenin signaling in cutaneous
keratinocytes is sufficient to trigger the active growth phase of
the hair cycle in mice (21,22). Therefore, total RNA was isolated from
C57BL/6 skin samples at 10 days after depilation and expression
levels of Wnt pathway genes were examined using RT-qPCR. Increased
mRNA levels of β-catenin (2.0-fold), cyclin D2 (Ccnd2; 1.4-fold),
lymphoid enhancer binding factor 1 (Lef1; 2.8-fold), Wnt10b
(2.3-fold) and Wnt5a (1.5-fold) were observed in the mice treated
with CXCL4 mAb compared with the control mice (Fig. 4A).
Previous studies have revealed that the Bmp pathway
may provide a molecular target for anagen induction and hair growth
in mice and humans (23), and that
Wnt1a-conditioned medium upregulates hair-induction related genes
(24). Similar to the aforementioned
RT-qPCR experiments, increased mRNA levels of Bmp4 (2.2-fold), Bmp2
(2.7-fold), GLI-Kruppel family member GLI1 (Gli1; 2.5-fold) and
versican (Vcan; 1.2-fold) were found in the mice treated with CXCL4
mAb in comparison with the control mice (Fig. 4B).
Effects of CXCL4 mAb on proliferation-
and apoptosis-associated proteins
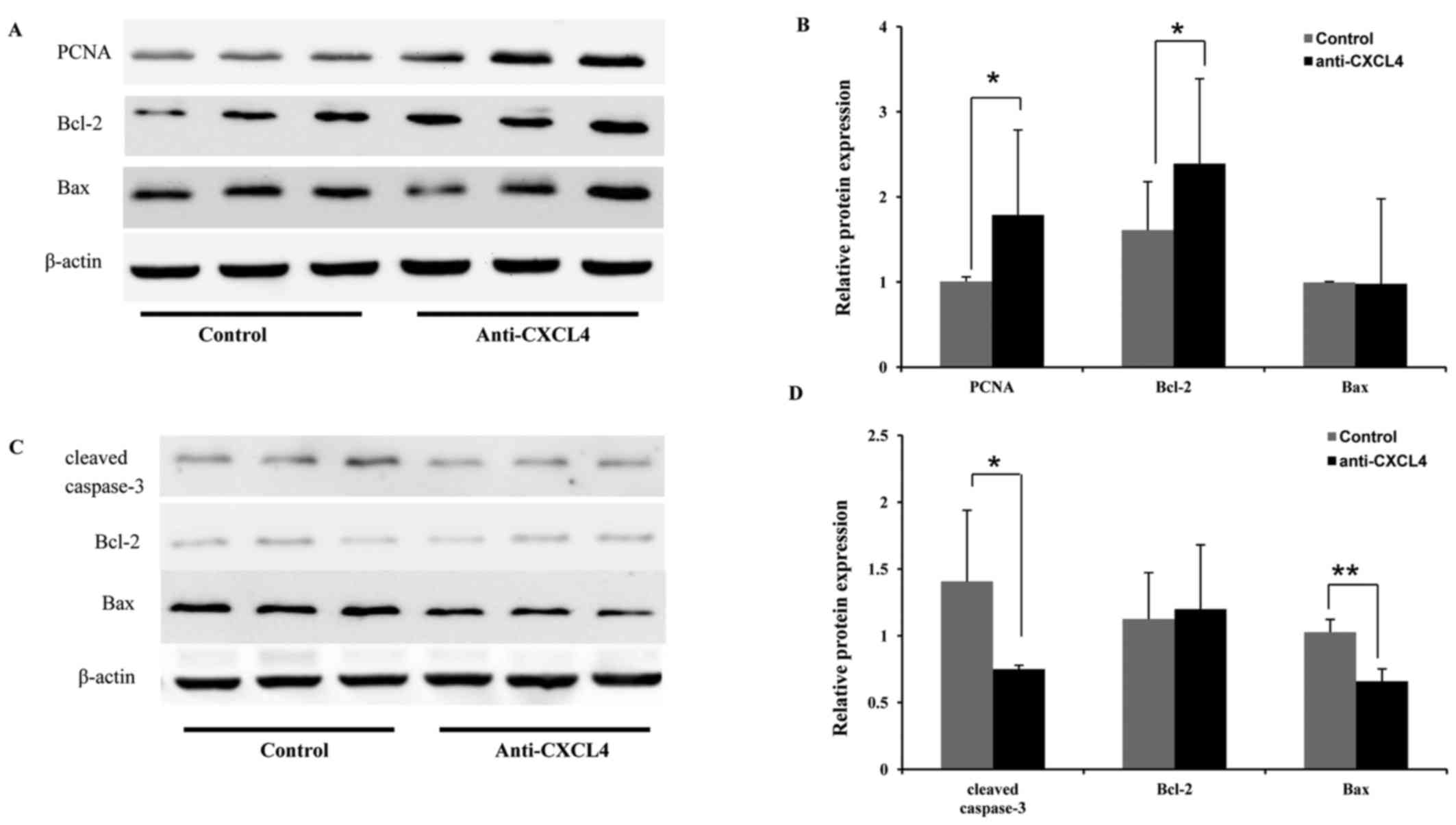
Whole-tissue lysates of dorsal skin tissues were
utilized to investigate the effects of CXCL4 mAb on proliferation-
and apoptosis-associated factors in hair growth and regression. At
10 days, CXCL4 mAb was observed to significantly increase Bcl-2
(1.5-fold) and PCNA (1.8-fold) expression during the anagen phase,
compared with that in the control group (Fig. 5A and B). In addition, at 18 days,
CXCL4 mAb treatment significantly reduced the protein expression of
two apoptosis-associated factors, Bax (1.6-fold) and cleaved
caspase-3 (1.9-fold), compared with that in the control group,
indicating that CXCL4 mAb prevented hair follicles from undergoing
apoptosis (Fig. 5C and D). These
results indicate that CXCL4 mAb promoted hair regrowth and delayed
hair regression.
Discussion
In the present study, a novel role for CXCL4 mAb in
the promotion of hair growth in mice following depilation was
uncovered. To the best of our knowledge, the present study is the
first to report an association between CXCL4 and hair growth. It
demonstrated that dorsal skin expressed low basal level of CXCL4
mRNA during the anagen phase of hair growth, and the expression
level returned to normal following the next telogen phase.
Furthermore, the present study demonstrated that CXCL4 was involved
in the control of hair growth, and that neutralization of CXCL4
using a mAb significantly promoted hair growth in a depilated mouse
model.
Previous studies have shown that CXCL4 inhibits
endothelial cell proliferation and interferes with the cell cycle
(12,25). It has been reported that
CXCL4-stimulated human monocytes induce the apoptosis of
endothelial cells via the release of oxygen radicals (26). This is consistent with our previous
study, in which recombinant human CXCL4 was observed to induce the
apoptosis of intestinal epithelial cells, CXCL4 mAb was
demonstrated to protect mouse intestine from chemotoxicity
(16). To the best of our knowledge,
there are no previous reports concerning the effect of CXCL4 or
anti-CXCL4 antibody on hair growth. In the present study, it was
identified that the expression of CXCL4 was downregulated during
the anagen phase and returned to the basal level at the telogen
phase. The effect of CXCL4 mAb on hair growth was also investigated
in the present study. The results suggest that CXCL4 mAb induces
hair growth by promoting telogen to anagen transition of hair
follicles, and suppresses apoptosis of hair follicles during the
catagen phase in mice.
Hair cycling between the telogen and anagen phases
is precisely regulated by dermal papilla (DP) cells. The Wnt
pathway is an important signaling pathway that activates the hair
induction ability of DP cells (27).
β-catenin and Wnt5a expression are moderate in the early anagen
phase, high in the middle anagen phase, and weak in the catagen and
telogen phases (21,28). Adenovirus-mediated ectopic expression
of Wnt10b was able to activate precocious anagen entry in a
previous mouse model (29). Lef1 is
an essential regulatory protein in the Wnt-signaling pathway that
controls cell growth and differentiation (30). In the stem cell niche, Wnt signaling
and β-catenin stabilization are considered to activate the
Lef1/T-cell factor complex, which binds to its target genes and
promotes stem cell activation and proliferation; these constitute
critical steps required to support hair growth (31). Moreover, the Bmp signal transduction
pathway also plays an important role in anagen induction (23). In the present study, elevated mRNA
levels of Wnt-β-catenin signaling molecules were detected,
including β-catenin, Ccnd2, Lef1, Wnt10b and Wnt5a at 10 days after
depilation in mice (Fig. 4A).
Furthermore, increased levels of Bmp4, Bmp2, Gli1 and Vcan were
also found in mice treated with CXCL4 mAb (Fig. 4B). These findings are consistent with
previous studies in which the expression levels of hair
induction-related genes were upregulated in hair regrowth (24,32). The
data demonstrate that CXCL4 mAb activates the Wnt/β-catenin- and
Bmp-signaling pathways to mediate hair regeneration.
CXCL4 has been demonstrated to induce apoptosis of
epithelia through activation of extrinsic and intrinsic death
pathways (16,26), which is regulated primarily by Bcl-2
family proteins. The Bcl-2 family of proteins are either
anti-apoptotic or pro-apoptotic in nature, notably the
anti-apoptotic Bcl-2 and pro-apoptotic Bax proteins. Bcl-2 is
considered to protect cells from apoptosis, while Bcl-2 activity is
determined by interaction with Bax (33). The present study demonstrated that
CXCL4 mAb increased the expression of the anti-apoptotic Bcl-2,
whereas there was no significant change in the expression level of
Bax in the skin at 10 days after depilation, at the anagen phase of
hair follicles. Notably, the level of PCNA expression was clearly
increased (Fig. 5). These results
indicate that CXCL4 mAb promotes telogen to anagen transition by
the stimulation of proliferation and suppression of apoptosis of
hair follicular cells.
During hair follicle growth and hair production,
factors promoting proliferation, differentiation and survival
predominate in activity. However, hair follicle regression in the
catagen phase is characterized by the activation of a number of
signaling pathways that induce the apoptosis of hair follicle cells
(34). In addition, in the process
of hair follicle regression, caspase activation is required to
induce apoptosis (35). Caspase-3 is
a key executor of the apoptosis signal (36). In the present study, during the
catagen phase (at 18 days after depilation), it was demonstrated
that pro-apoptotic Bax expression was decreased, the expression of
Bcl-2 was unchanged, and the expression of cleaved caspase-3 was
significantly decreased in the skin treated with CXCL4 mAb
(Fig. 5). During the catagen phase,
the dermal papilla transforms into a cluster of quiescent cells
near to the regressing hair follicle epithelium, which moves from
the subcutis to the border of the dermis and subcutis (35). In the present study, suppression of
hair follicle apoptosis prevented the dermal papilla from being
pulled upwards from the subcutis to the dermis in the catagen phase
of hair follicles in the mice treated with CXCL4 mAb (Fig. 3). Thus, CXCL4 mAb attenuated the
apoptosis of hair follicle cells in the catagen phase and delayed
hair follicle regression.
In conclusion, blockade of CXCL4 using CXCL4 mAb
accelerated hair growth through promotion of telogen to anagen
transition, and delayed the regression of hair follicles in the
catagen phase. In-depth studies are necessary to further elucidate
the molecular mechanisms through which CXCL4 mAb imparts its hair
growth activity.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (No. 81273573) and the Science
and Technology Commission of Shanghai Municipality (No.
11431921300).
References
|
1
|
Müller-Röver S, Handjiski B, van der Veen
C, Eichmüller S, Foitzik K, McKay IA, Stenn KS and Paus R: A
comprehensive guide for the accurate classification of murine hair
follicles in distinct hair cycle stages. J Invest Dermatol.
117:3–15. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schneider MR, Schmidt-Ullrich R and Paus
R: The hair follicle as a dynamic miniorgan. Curr Biol.
19:R132–R142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang TL, Zhou C, Shen YW, Wang XY, Ding
XL, Tian S, Liu Y, Peng GH, Xue SQ, Zhou JE, et al: Prevalence of
androgenetic alopecia in China: A community-based study in six
cities. Br J Dermatol. 162:843–847. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cotsarelis G and Millar SE: Towards a
molecular understanding of hair loss and its treatment. Trends Mol
Med. 7:293–301. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang NN, Park DK and Park HJ: Hair
growth-promoting activity of hot water extract of Thuja orientalis.
BMC Complement Altern Med. 13:92013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miller MD and Krangel MS: Biology and
biochemistry of the chemokines: A family of chemotactic and
inflammatory cytokines. Crit Rev Immunol. 12:17–46. 1992.PubMed/NCBI
|
|
7
|
Deutsch E, Johnson SA and Seegers WH:
Differentiation of certain platelet factors related to blood
coagulation. Circ Res. 3:110–115. 1955. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rendu F and Brohard-Bohn B: The platelet
release reaction: Granules' constituents, secretion and functions.
Platelets. 12:261–273. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kasper B and Petersen F: Molecular
pathways of platelet factor 4/CXCL4 signaling. Eur J Cell Biol.
90:521–526. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scheuerer B, Ernst M, Dürrbaum-Landmann I,
Fleischer J, Grage-Griebenow E, Brandt E, Flad HD and Petersen F:
The CXC-chemokine platelet factor 4 promotes monocyte survival and
induces monocyte differentiation into macrophages. Blood.
95:1158–1166. 2000.PubMed/NCBI
|
|
11
|
Han ZC, Lu M, Li J, Defard M, Boval B,
Schlegel N and Caen JP: Platelet factor 4 and other CXC chemokines
support the survival of normal hematopoietic cells and reduce the
chemosensitivity of cells to cytotoxic agents. Blood. 89:2328–2335.
1997.PubMed/NCBI
|
|
12
|
Gupta SK and Singh JP: Inhibition of
endothelial cell proliferation by platelet factor-4 involves a
unique action on S phase progression. J Cell Biol. 127:1121–1127.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luster AD, Greenberg SM and Leder P: The
IP-10 chemokine binds to a specific cell surface heparan sulfate
site shared with platelet factor 4 and inhibits endothelial cell
proliferation. J Exp Med. 182:219–231. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Maione TE, Gray GS, Petro J, Hunt AJ,
Donner AL, Bauer SI, Carson HF and Sharpe RJ: Inhibition of
angiogenesis by recombinant human platelet factor-4 and related
peptides. Science. 247:77–79. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Watson JB, Getzler SB and Mosher DF:
Platelet factor 4 modulates the mitogenic activity of basic
fibroblast growth factor. J Clin Invest. 94:261–268. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao J, Gao J, Qian L, Wang X, Wu M, Zhang
Y, Ye H, Zhu S, Yu Y and Han W: Activation of p38-MAPK by
CXCL4/CXCR3 axis contributes to p53-dependent intestinal apoptosis
initiated by 5-fluorouracil. Cancer Biol Ther. 15:982–991. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paus R, Stenn KS and Link RE: Telogen skin
contains an inhibitor of hair growth. Br J Dermatol. 122:777–784.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Plonka PM, Michalczyk D, Popik M,
Handjiski B, Slominski A and Paus R: Splenic eumelanin differs from
hair eumelanin in C57BL/6 mice. Acta Biochim Pol. 52:433–441.
2005.PubMed/NCBI
|
|
20
|
Datta K, Singh AT, Mukherjee A, Bhat B,
Ramesh B and Burman AC: Eclipta alba extract with potential for
hair growth promoting activity. J Ethnopharmacol. 124:450–456.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huelsken J, Vogel R, Erdmann B, Cotsarelis
G and Birchmeier W: beta-Catenin controls hair follicle
morphogenesis and stem cell differentiation in the skin. Cell.
105:533–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Van Mater D, Kolligs FT, Dlugosz AA and
Fearon ER: Transient activation of beta -catenin signaling in
cutaneous keratinocytes is sufficient to trigger the active growth
phase of the hair cycle in mice. Genes Dev. 17:1219–1224. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mundy G, Gutierrez G, Garrett R, Gallwitz
W, Rossini G, Christiansen C and Langenberg A: Proteasome
inhibitors stimulate both bone formation and hair growth by similar
mechanisms. Ann N Y Acad Sci. 1117:298–301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dong L, Hao H, Xia L, Liu J, Ti D, Tong C,
Hou Q, Han Q, Zhao Y, Liu H, et al: Treatment of MSCs with
Wnt1a-conditioned medium activates DP cells and promotes hair
follicle regrowth. Sci Rep. 4:54322014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gentilini G, Kirschbaum NE, Augustine JA,
Aster RH and Visentin GP: Inhibition of human umbilical vein
endothelial cell proliferation by the CXC chemokine, platelet
factor 4 (PF4), is associated with impaired downregulation of
p21(Cip1/WAF1). Blood. 93:25–33. 1999.PubMed/NCBI
|
|
26
|
Woller G, Brandt E, Mittelstädt J,
Rybakowski C and Petersen F: Platelet factor 4/CXCL4-stimulated
human monocytes induce apoptosis in endothelial cells by the
release of oxygen radicals. J Leukoc Biol. 83:936–945. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Soma T, Fujiwara S, Shirakata Y, Hashimoto
K and Kishimoto J: Hair-inducing ability of human dermal papilla
cells cultured under Wnt/β-catenin signalling activation. Exp
Dermatol. 21:307–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fuchs E and Raghavan S: Getting under the
skin of epidermal morphogenesis. Nat Rev Genet. 3:199–209. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li YH, Zhang K, Yang K, Ye JX, Xing YZ,
Guo HY, Deng F, Lian XH and Yang T: Adenovirus-mediated Wnt10b
overexpression induces hair follicle regeneration. J Invest
Dermatol. 133:42–48. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Andl T, Reddy ST, Gaddapara T and Millar
SE: WNT signals are required for the initiation of hair follicle
development. Dev Cell. 2:643–653. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gat U, DasGupta R, Degenstein L and Fuchs
E: De Novo hair follicle morphogenesis and hair tumors in mice
expressing a truncated beta-catenin in skin. Cell. 95:605–614.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang S, Zhao L, Teklemariam T and Hantash
BM: Small cutaneous wounds induce telogen to anagen transition of
murine hair follicle stem cells. J Dermatol Sci. 60:143–150. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Adams JM and Cory S: The Bcl-2 protein
family: Arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lindner G, Botchkarev VA, Botchkareva NV,
Ling G, van der Veen C and Paus R: Analysis of apoptosis during
hair follicle regression (catagen). Am J Pathol. 151:1601–1617.
1997.PubMed/NCBI
|
|
35
|
Botchkareva NV, Ahluwalia G and Shander D:
Apoptosis in the hair follicle. J Invest Dermatol. 126:258–264.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sabbagh L, Kaech SM, Bourbonnière M, Woo
M, Cohen LY, Haddad EK, Labrecque N, Ahmed R and Sékaly RP: The
selective increase in caspase-3 expression in effector but not
memory T cells allows susceptibility to apoptosis. J Immunol.
173:5425–5433. 2004. View Article : Google Scholar : PubMed/NCBI
|