Introduction
Docosahexaenoic acid (DHA) and eicosapentaenoic acid
(EPA) are two distinct forms of omega-3 fatty acids (FAs). Omega-3
FAs are typically found in fish oil and have a general
polyunsaturated FA structure with the presence of a double bond at
the third carbon (when counting from the methyl end of the chain)
(1). Increased consumption of
omega-3 FAs has been associated with a lower incidence of various
types of cancer, including colon (2), breast (3,4) and
prostate cancer (3,4). It has also been demonstrated that
omega-3 FAs exert a range of anti-tumor effects, including
inhibitory effects on angiogenic mediator production (5), the induction of apoptosis (6), inhibitory effects on tumor invasion and
metastasis (7,8) and regulatory effects on signaling
pathways (9). In particular, the
anti-tumor effects of EPA and DHA have been observed in breast
cancer, whereby higher intakes of these FAs are associated with a
reduced risk of mortality (10,11). It
has also been demonstrated that administration of DHA during
anthracyclin-based chemotherapy against metastatic breast cancer
improves clinical outcomes, suggesting that omega-3 FA may be an
effective adjuvant in the treatment of cancer (12).
Renal cell carcinoma (RCC) represents ~3% of all
adult cancer cases (13). In
patients with RCC, ~30% have metastatic disease at the time of
diagnosis, while another 20–30% develop metastases following
surgery (14). Although the
treatment options available for metastatic RCC have recently
improved due to the clinical development of targeting agents
including sorafenib, sunitinib and everolimus (15), metastatic RCC remains a fatal
disease. RCC cells originate from the renal proximal tubules and
express high levels of multi-drug resistance proteins, thus they
are resistant to most forms of chemotherapy. Previous attempts to
treat patients with RCC using targeted agents have failed in the
majority of cases (14,16). Therefore, studies aiming to identify
a novel therapeutic agent to treat patients with metastatic RCC are
required. The present study used in vitro techniques
including MTS and proliferation assays and flow cytometry analysis
to investigate the anti-tumor activities of DHA on the
proliferative and invasive capacities of RCC cells at clinically
relevant concentrations of 10–200 µM, as previously determined
(17–20). The potential roles of signal
transducer and activator of transcription 3 (STAT3) signaling in
the cellular activities of DHA-treated renal cancer cells were also
evaluated.
Materials and methods
Cell lines and reagents
The human renal cancer cell lines Caki-1 and 786-O
were purchased from the American Type Culture Collection (Manassas,
VA, USA). DHA was purchased from Cayman Chemical Company (Ann
Arbor, MI, USA). Antibodies against epidermal growth factor
receptor (EGFR; cat. no. 4267S), phosphorylated (p)-EGFR
(p-Tyr1068; cat. no. 2234S), STAT3 (cat. no. 9132S), p-STAT3
(p-Tyr705; cat. no. 9145 L), extracellular signal-regulated kinase
(ERK; cat. no. 9102), p-ERK (p-Thr202/Tyr204; cat. no. 9101S), Akt
(cat. no. 9272) and p-Akt (p-Ser473; cat. no. 9271; all from Cell
Signaling Technology, Inc., Danvers, MA, USA) were used in the
present study. Horseradish peroxidase (HRP)-conjugated secondary
antibodies and an Amersham Enhanced Chemiluminescence (ECL) Gel
system (GE Healthcare Life Sciences, Chalfont, UK) were also
used.
Cell viability and proliferation
assays
Cell viability was assessed by an MTS assay using a
CellTiter 96® AQueous Non-Radioactive Cell Proliferation assay kit
(Promega Corporation, Madison, WI, USA), as previously described
(21). Briefly, cells were seeded
into 96-well plates (3×103 cells/well) and following
overnight incubation at 37°C, were treated for 24 h at 37°C with
DHA (0, 25, 50 and 100 µM) prior to addition of MTS solution. The
medium used for Caki-1 cells was 1X minimum essential media (MEM)
with 10% fetal bovine serum (FBS) and the medium for 786-O cells
was RPMI medium 1640 with 10% FBS, both purchased from Gibco;
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). At 2 h post-MTS
addition. Prior to the measurement of absorbance, 20 µl CellTiter
96® Aqueous One Solution Reagent per well was added before 2 h
incubation at 37°C. The absorbance of plates was then measured at a
wavelength of 490 nm with a microplate autoreader. A control group
with 0 µM DHA were used as a comparison.
Cell proliferation was assessed by counting cell
numbers after cells seeded into 6-well plates (1×104
cells/well) had been incubated at 37°C with DHA (0, 50, 100 µM) for
0, 24, 48 and 72 h. The medium used for Caki-1 cells was 1X MEM
with 10% FBS and the medium for 786-O cells was RPMI medium 1640
with 10% FBS, as previously stated. Total cell numbers were then
counted in four fields using a hemocytometer and mean values were
calculated from three replicates. The four fields were the four
corners of each square in the nine large squares of the
hemocytometer, counted using a CK40 phase contrast microscope
(Olympus Corporation, Tokyo, Japan).
Flow cytometry analysis
Cells were incubated for 24 h with 100 µM DHA and a
control group with 0 µM DHA were used as a comparison.
Propidium-iodide (PI)-stained nuclear fractions were obtained and
cell cycle data were acquired with a flow cytometer using
CellQuest™ software, version 5.2.1 (BD Biosciences,
Franklin Lakes, NJ, USA), following the manufacturer's protocol.
Percentages of apoptotic cells were also determined with a
fluorescein isothiocyanate-conjugated Annexin V/PI double-staining
assay, using an Annexin V Apoptosis Detection kit (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), as described previously
(21).
Cell motility and invasion assay
Cells were grown to 90–100% confluence on 6-well
tissue culture plates. Cells were seeded at 0.3×106 in a
6 well flat bottom plate (IWAKI, Co., Ltd., Hong Kong, China) with
MEM (1X) with 10% FBS and RPMI 1640 (1X) with 10% FBS for Caki-1
and 786-O cells, respectively. A wound was then made by scraping
the middle of the cell monolayer with a P200 pipette tip, as
previously described (21). After
floating cells were removed following an extensive wash with 1 ml
ice-cold phosphate-buffered saline, fresh complete MEM and RPMI
medium supplemented with DHA was added to each type of cell, as
previously detailed. Following 12 h, cell migration and movements
throughout the wound area were observed using a CK40 phase contrast
microscope (Olympus Corporation, Tokyo, Japan). ImageJ software
version 1.46 (National Institutes of Health, Bethesda, MA, USA) was
used to analyze the images.
Cell invasion was evaluated using a Matrigel-coated
Transwell system in 24-well plates including a membrane with 8-µm
pores (BD Biosciences), as described previously (21). Briefly, 2×105 cells
suspended in 500 µl serum-free medium (MEM and RPMI, as previously
detailed) were added to the insert and 750 µl serum-free medium
with the indicated concentration of DHA (0, 25, 50 and 100 µM) was
added to the bottom of the well. Following incubation for 24 h at
37°C, the inserts were fixed in 100% methanol, then filters were
stained with 1% toluidine blue in 1% borax for 10 min. Cells that
had invaded through the Matrigel-coated Transwell inserts were
counted at a magnification of ×400. Numbers of cells in at least 10
randomly selected fields/wells were counted in three independent
experiments.
Western blot analysis
Cells at a density of 2×106 were treated
with 0 (control), 50 and 100 µM DHA for 6 h, then lysed in
radioimmunoprecipitation assay buffer composed of 10 mM tris-HCl
(Nacalai Tesque, Inc., Kyoto, Japan), 150 mM NaCl (Kanto Chemical,
Co., Inc., Tokyo, Japan), 1% Triton X-100 (MP Biomedicals LLC,
Santa Ana, CA, USA), 5 µM ethylenediaminetetra acetic acid (Nacalai
Tesque, Inc.), 1%sodium deoxycholate (Difco; BD Biosciences, San
Jose, CA, USA), 0.1% sodium dodecyl sulfate, 1.2% aprotinin, 5 µM
leupeptin, 4 µM antipain, 1 mM phenylmethylsulfonyl fluoride and
0.1 mM Na3VO4 (all; Sigma Aldrich; Merck
KGaA, Darmstadt, Germany) according to a previously described
method (21). Samples were
centrifuged at 16,000 × g for 20 min at 4°C. The amount of protein
was quantified using the DC protein assay kit (BD Biosciences)
according to the manufacturers protocol. Equal amounts (50 µl) of
resulting lysates were separated using a 10% SDS-PAGE gel and
transferred to nitrocellulose membranes. Membranes were then
blocked at room temperature with 5% skimmed milk and tris-buffered
saline and Tween-20 solution for 1 h. The membranes were then
incubated at 4°C overnight with primary antibodies, then incubated
at room temperature for 1 h with the corresponding secondary
antibody (Rabbit Ig HRP-linked Whole Ab, from Donkey; NA934-1ML; GE
Healthcare Japan Corporation, Tokyo, Japan) at a 1:5,000 dilution.
Immunolabelled proteins were subsequently visualized by enhanced
chemiluminescence with an Amersham ECL Gel System, according to the
manufacturer's protocol.
Statistical analysis
Data are presented as the mean ± standard error of
the mean of three independent experiments. One way analysis of
variance followed by a protected Fisher's least significant
difference post hoc test was used to analyze continuous data. The
statistical significance of differences was evaluated using a
paired t-test and P<0.05 was considered to indicate a
statistically significant difference. JMP version 9 was used to
assess all data (SAS Institute, Inc., Cary, NC, USA).
Results
DHA inhibits the growth of renal
cancer cells by inducing apoptosis
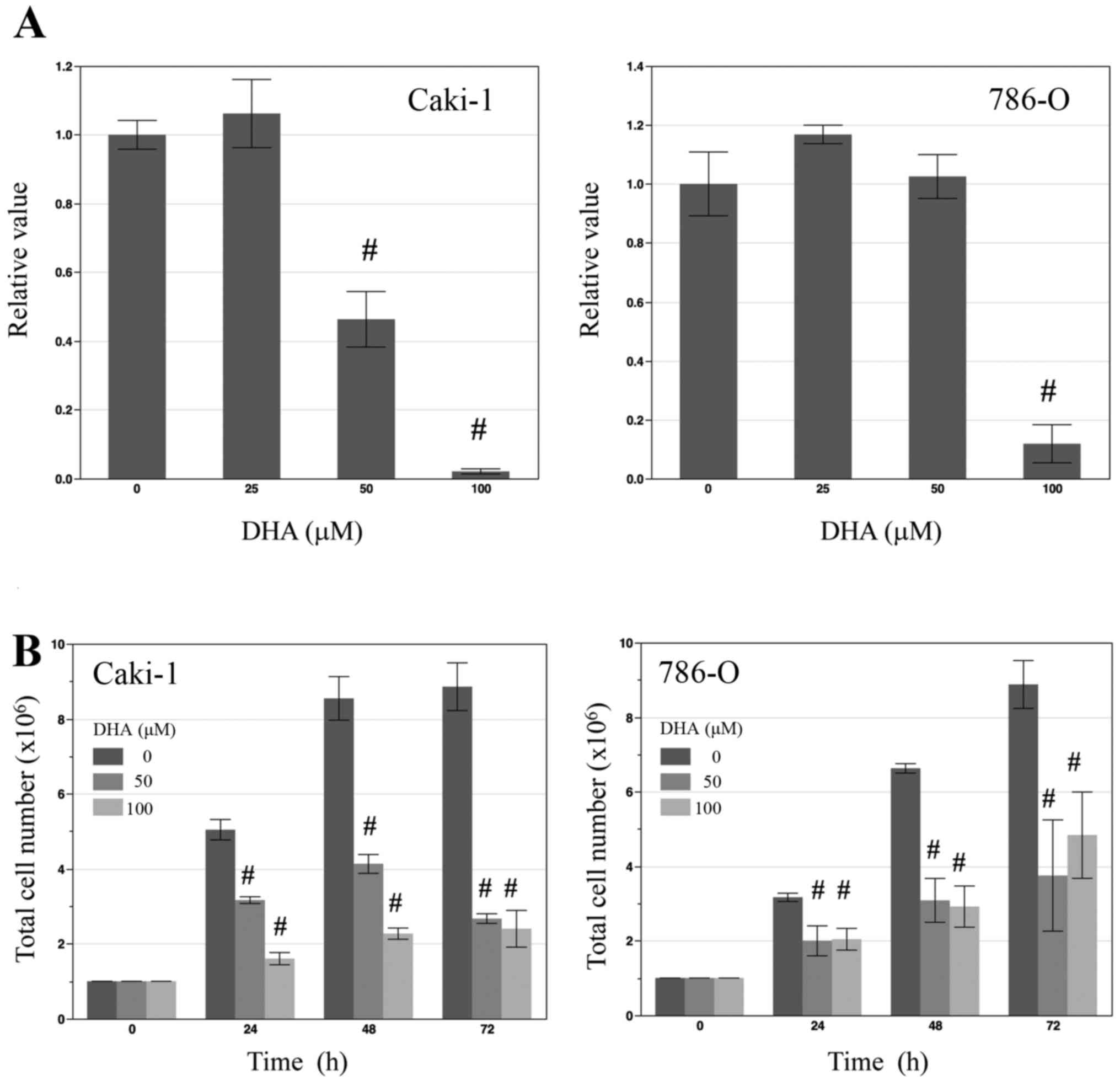
To determine the effect of DHA on the proliferation
of renal cancer cells in vitro, the renal cancer cell lines
Caki-1 and 786-O were treated with various concentrations of DHA
(0, 25, 50 and 100 µM) for 24 h prior to an MTS cell viability
assay. In Caki-1 cells, treatment with 50 and 100 µM DHA led to
significant decreases in cell viability (both P<0.01; Fig. 1A), while in 786-O cells, only 100 µM
DHA induced a significant decrease in cell viability compared with
the control (P<0.01; Fig. 1A). To
determine total cell numbers, Caki-1 and 786-O cells were counted
following treatment with 0, 50, 100 µM DHA for 24, 48 and 72 h. At
each time point, total numbers of DHA-treated cells were
significantly lower than those of untreated control cells (all
P<0.01; Fig. 1B).
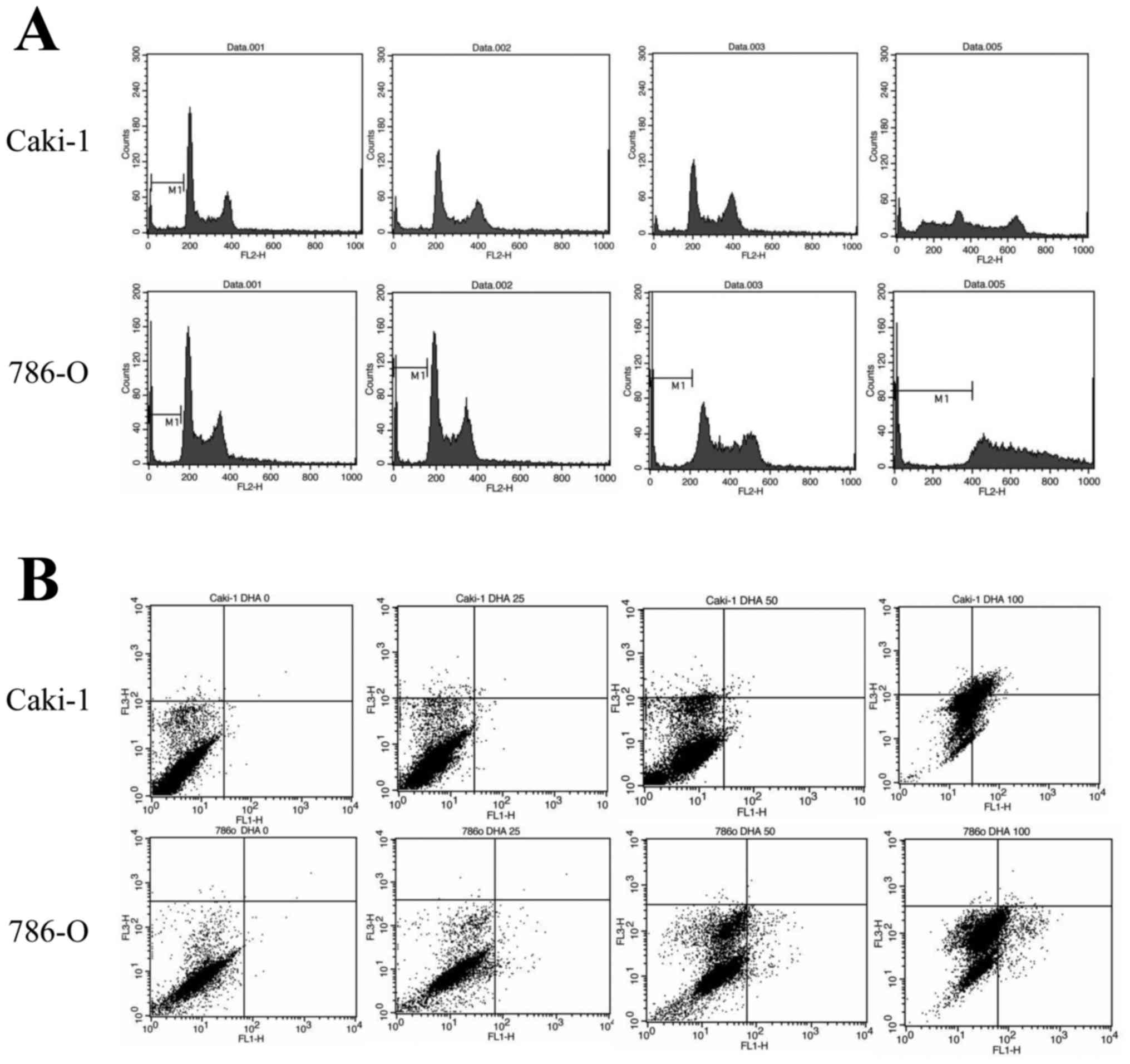
To determine whether the inhibitory action of DHA on
cell proliferation was due to apoptosis-inducing effects, Caki-1
and 786-O cells were treated with 100 µM DHA for 24 h prior to cell
cycle analysis by flow cytometry. Following treatment with DHA, it
was observed that the sub-G1 populations of Caki-1 and 786-O cells
(37.44 and 16.46%, respectively) were greater than those of
untreated controls (7.09% in Caki-1 and 6.26% in 786-O control
cells: Fig. 2A). DHA-treated cell
lines were also double stained with Annexin V and PI and analyzed
by flow cytometry. Following treatment with 100 µM DHA, it was
observed that the percentages of Annexin-positive and PI-negative
(apoptotic) cells in both the Caki-1 and 786-O cell lines were
increased (18.81 and 9.92%, respectively), relative to untreated
controls (0.22% in Caki-1 and 0.14% in 786-O control cells;
Fig. 2B). Collectively, these data
suggest that DHA inhibits the growth of renal cancer cells through
the induction of apoptosis.
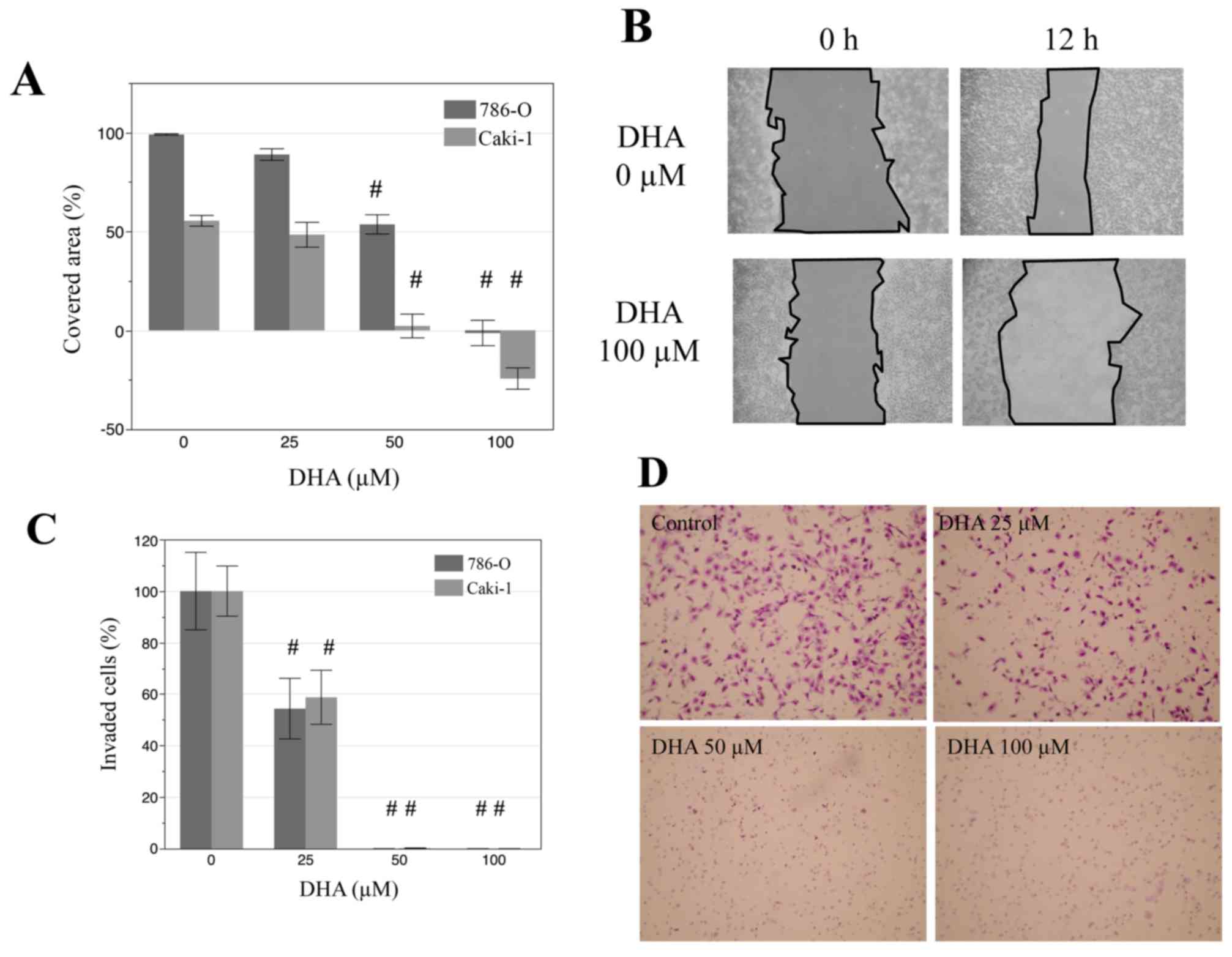
DHA inhibits cell motility and
invasiveness of renal cancer cells
The effects of DHA on the migration and invasion of
renal cancer cells were also evaluated by a wound scratch assay.
Monolayers of Caki-1 and 786-O cells were disrupted to create a
uniform wound and grown for 12 h with 0, 25, 50, 100 µM DHA. In
both cell lines, treatment with 50 and 100 µM DHA lead to
significant decreases in the area covered by cells, relative to
that of untreated controls (Fig 3A and
B; P<0.001). Similarly, in Matrigel invasion assays, DHA
treatment (25, 50 and 100 µM) significantly decreased the invasive
properties of both cell lines (Fig. 3C
and D; P<0.001). In particular, there were markedly low
numbers of invaded cells following treatment with 50 and 100 µM
DHA. These results indicate that DHA may suppress the migration and
invasion of renal cancer cells.
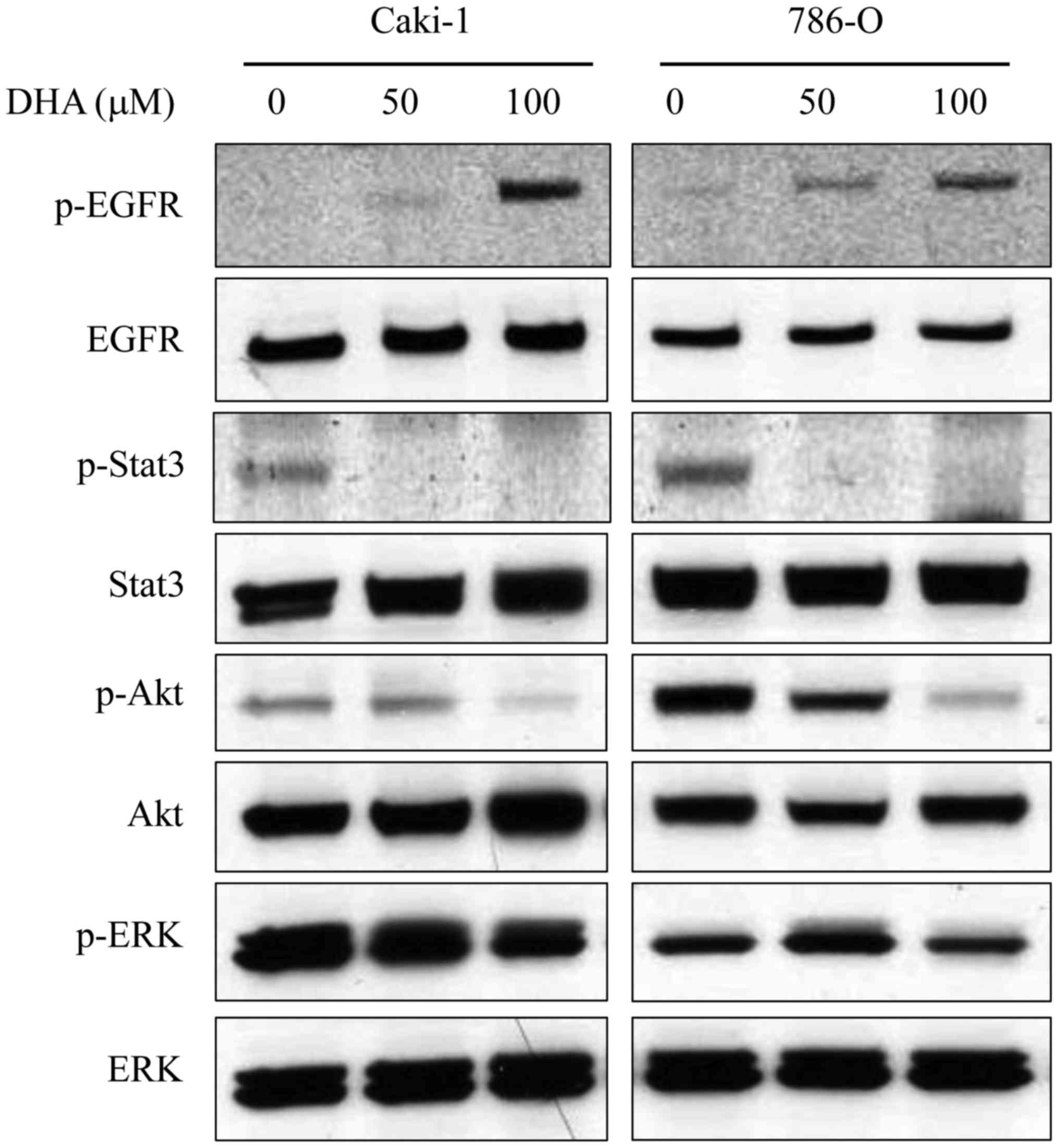
DHA alters EGFR phosphorylation status
and STAT3 signaling
Potential signaling pathways underlying the
anti-tumor activity of DHA in renal cancer cells were subsequently
investigated. Caki-1 and 786-O cells lysates were subjected to
western blotting following incubation of cells for 6 h in the
presence or absence of DHA (50 and 100 µM; Fig. 4). Previous studies have demonstrated
that DHA is incorporated into cellular membranes, where it may
alter the localization and function of EGFR by disrupting its
association with membrane lipid raft microdomains (9). Furthermore, it has been documented that
EGFR is overexpressed in RCC (22).
In Caki-1 and 786-O cells, DHA treatment markedly increased the
phosphorylation of EGFR in a dose-dependent manner, though had no
observable effect on EGFR expression (Fig. 4). The phosphorylation of key
signaling molecules downstream of EGFR, namely STAT3, Akt and ERK,
was also evaluated. Levels of p-STAT3 and p-Akt in both cell lines
were decreased by DHA in a dose-dependent manner (Fig. 4). However DHA had no discernable
effect on the levels of STAT3 and Akt. Furthermore, DHA treatment
had little effect on the levels of ERK or p-ERK (Fig. 4).
Discussion
In the present study, DHA inhibited the
proliferation and invasion of renal cancer cells in vitro,
possibly through regulatory effects on the functions of EGFR, STAT3
and Akt. A previous study in Japan documented that the mean
concentration of serum DHA in healthy controls was 18.52 mg/dl
(563.0 µM) (17) and previous
studies investigating the anti-tumor effects of DHA on colon
(18), breast (19) and prostate cancer cells in
vivo (20) have used DHA
concentrations between 10 and 200 µM. Therefore, the present study
used DHA concentrations ≤100 µM to obtain clinically relevant
results.
Results from epidemiological and preclinical studies
suggest that omega-3 FAs, including DHA, reduce the incidence of
various types of cancer (1,3,4,23). In a mouse model of breast cancer,
tumor size was reduced significantly by dietary DHA in a
dose-dependent manner (24). Indeed,
DHA has been documented to have a range of anti-tumor effects,
including inhibitory effects on tumor angiogenesis (5), apoptosis-inducing activity (6), inhibitory effects on tumor invasion and
metastasis (7,8) and regulatory effects on signaling
pathways (9). In addition, it has
been documented that serum levels of omega-3 FAs in patients with
pancreatic cancer, lung cancer or non-Hodgkin lymphoma were lower
than those in healthy controls (25,26),
with the lowest levels observed in patients with more
advanced-stage cancer (26,27). It has also been demonstrated that DHA
reduces metastasis in animal models using mice (7,8) and
prevents the migration and invasion of human MDA-MB-231 mammary
cancer cells in vitro (7).
Diets high in DHA-rich fish oil prevent breast cancer metastasis to
the bone by reducing the expression of cluster of differentiation
44, regarded as a typical molecular signature of cancer stem cells
(7). Furthermore, in a mouse model
of Lewis lung carcinoma, DHA metabolites were found to inhibit
vascular endothelial growth factor- and fibroblast growth
factor-induced angiogenesis and suppress primary tumor growth and
metastasis (8). Since previous
studies have identified an association between omega-3 FAs and
cancer development, the present study investigated whether DHA
serves a role in the progression and/or metastasis of RCC. DHA is
incorporated into cellular membranes and may disrupt lipid raft
microdomains that act as detergent-resistant signaling platforms
(28), thus leading to a disruption
in signaling pathways associated with numerous cancers, including
EGFR signaling (9). EGFR is a
transmembrane receptor tyrosine kinase and its overexpression is
frequently observed in RCC (22).
Activation of EGFR stimulates downstream signaling pathways that
have been implicated in the regulation of tumor growth, invasion
and metastasis (29). However,
membrane localization of EGFR is essential for its modulatory
effects on downstream signaling pathways, and previous studies have
indicated that DHA displaces EGFR from lipid rafts, thus leading to
a subsequent decrease in its downstream signaling activity
(9). Analogous to previous findings,
the present study observed that DHA treatment led to changes in the
phosphorylation status of EGFR and its downstream signaling
pathways in renal cancer cells. It has previously been observed
that STAT3 and Akt are constitutively activated in RCC patients,
particularly in those with metastatic diseases (30,31). In
turn, constitutively activated Akt in renal cancer cells may confer
cellular resistance to the EGFR inhibitor gefitinib (32). The present study demonstrated that
DHA inhibited the activation of STAT3 and Akt in a dose-dependent
manner. As the STAT3 and Akt pathways serve a key role in
apoptosis, cell migration and invasion (30,31),
these data suggest that these pathways serve a role in cancer
progression.
In conclusion, similar to results from previous
preclinical and clinical studies investigating a range of cancers,
the present results suggest that DHA may be a potential therapeutic
agent for the treatment of renal carcinoma. However, applications
of the present findings are limited, due to a lack of in
vivo and clinical data regarding RCC. Therefore, future
clinical studies into the efficacy of DHA in preventing RCC
metastasis are necessary.
References
|
1
|
Larsson SC, Kumlin M, Ingelman-Sundberg M
and Wolk A: Dietary long-chain n-3 fatty acids for the prevention
of cancer: A review of potential mechanisms. Am J Clin Nutr.
79:935–945. 2004.PubMed/NCBI
|
|
2
|
Sasazuki S, Inoue M, Iwasaki M, Sawada N,
Shimazu T, Yamaji T, Takachi R and Tsugane S; Japan Public Health
Center-Based Prospective Study Group, : Intake of n-3 and n-6
polyunsaturated fatty acids and development of colorectal cancer by
subsite: Japan public health center-based prospective study. Int J
Cancer. 129:1718–1729. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Terry PD, Rohan TE and Wolk A: Intakes of
fish and marine fatty acids and the risks of cancers of the breast
and prostate and of other hormone-related cancers: A review of the
epidemiologic evidence. Am J Clin Nutr. 77:532–543. 2003.PubMed/NCBI
|
|
4
|
Sonoda T, Nagata Y, Mori M, Miyanaga N,
Takashima N, Okumura K, Goto K, Naito S, Fujimoto K, Hirao Y, et
al: A case-control study of diet and prostate cancer in Japan:
Possible protective effect of traditional Japanese diet. Cancer
Sci. 95:238–242. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Spencer L, Mann C, Metcalfe M, Webb M,
Pollard C, Spencer D, Berry D, Steward W and Dennison A: The effect
of omega-3 FAs on tumour angiogenesis and their therapeutic
potential. Eur J Cancer. 45:2077–2086. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Serini S, Trombino S, Oliva F, Piccioni E,
Monego G, Resci F, Boninsegna A, Picci N, Ranelletti FO and
Calviello G: Docosahexaenoic acid induces apoptosis in lung cancer
cells by increasing MKP-1 and down-regulating p-ERK1/2 and p-p38
expression. Apoptosis. 13:1172–1183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mandal CC, Ghosh-Choudhury T, Yoneda T,
Choudhury GG and Ghosh-Choudhury N: Fish oil prevents breast cancer
cell metastasis to bone. Biochem Biophys Res Commun. 402:602–607.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang G, Panigrahy D, Mahakian LM, Yang J,
Liu JY, Lee KS Stephen, Wettersten HI, Ulu A, Hu X, Tam S, et al:
Epoxy metabolites of docosahexaenoic acid (DHA) inhibit
angiogenesis, tumor growth, and metastasis. Proc Natl Acad Sci USA.
110:pp. 6530–6535. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rogers KR, Kikawa KD, Mouradian M,
Hernandez K, McKinnon KM, Ahwah SM and Pardini RS: Docosahexaenoic
acid alters epidermal growth factor receptor-related signaling by
disrupting its lipid raft association. Carcinogenesis.
31:1523–1530. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Patterson RE, Flatt SW, Newman VA,
Natarajan L, Rock CL, Thomson CA, Caan BJ, Parker BA and Pierce JP:
Marine fatty acid intake is associated with breast cancer
prognosis. J Nutr. 141:201–206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vaughan VC, Hassing MR and Lewandowski PA:
Marine polyunsaturated fatty acids and cancer therapy. Br J Cancer.
108:486–492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bougnoux P, Hajjaji N, Ferrasson MN,
Giraudeau B, Couet C and Le Floch O: Improving outcome of
chemotherapy of metastatic breast cancer by docosahexaenoic acid: A
phase II trial. Br J Cancer. 101:1978–1985. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bukowski RM: Prognostic factors for
survival in metastatic renal cell carcinoma: Update, 2008. Cancer.
115 10 Suppl:S2273–S2281. 2009. View Article : Google Scholar
|
|
15
|
Linehan WM, Srinivasan R and Schmidt LS:
The genetic basis of kidney cancer: A metabolic disease. Nat Rev
Urol. 7:277–285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sonpavde G and Choueiri TK: Biomarkers:
The next therapeutic hurdle in metastatic renal cell carcinoma. Br
J Cancer. 107:1009–1016. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghadimi R, Kuriki K, Tsuge S, Takeda E,
Imaeda N, Suzuki S, Sawai A, Takekuma K, Hosono A, Tokudome Y, et
al: Serum concentrations of fatty acids and colorectal adenoma
risk: A case-control study in Japan. Asian Pac J Cancer Prev.
9:111–118. 2008.PubMed/NCBI
|
|
18
|
Horiguchi A, Asano T, Ito K, Sumitomo M
and Hayakawa M: Pharmacological inhibitor of fatty acid synthase
suppresses growth and invasiveness of renal cancer cells. J Urol.
180:729–736. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Calviello G, Resci F, Serini S, Piccioni
E, Toesca A, Boninsegna A, Monego G, Ranelletti FO and Palozza P:
Docosahexaenoic acid induces proteasome-dependent degradation of
beta-catenin, down-regulation of survivin and apoptosis in human
colorectal cancer cells not expressing COX-2. Carcinogenesis.
28:1202–1209. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang KS, Wang P, Yamabe N, Fukui M, Jay T
and Zhu BT: Docosahexaenoic acid induces apoptosis in MCF-7 cells
in vitro and in vivo via reactive oxygen species formation and
caspase 8 activation. PLoS One. 5:e102962010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu Y, Sun H, Owens RT, Gu Z, Wu J, Chen
YQ, O'Flaherty JT and Edwards IJ: Syndecan-1-dependent suppression
of PDK1/Akt/bad signaling by docosahexaenoic acid induces apoptosis
in prostate cancer. Neoplasia. 12:826–836. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Minner S, Rump D, Tennstedt P, Simon R,
Burandt E, Terracciano L, Moch H, Wilczak W, Bokemeyer C, Fisch M,
et al: Epidermal growth factor receptor protein expression and
genomic alterations in renal cell carcinoma. Cancer. 118:1268–1275.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sawada N, Inoue M, Iwasaki M, Sasazuki S,
Shimazu T, Yamaji T, Takachi R, Tanaka Y, Mizokami M and Tsugane S;
Japan Public Health Center-Based Prospective Study Group, :
Consumption of n-3 fatty acids and fish reduces risk of
hepatocellular carcinoma. Gastroenterology. 142:1468–1475. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
El-Mesery M, Al-Gayyar M, Salem H,
Darweish M and El-Mowafy A: Chemopreventive and renal protective
effects for docosahexaenoic acid (DHA): Implications of CRP and
lipid peroxides. Cell Div. 4:62009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zuijdgeest-van Leeuwen SD, van der Heijden
MS, Rietveld T, Van den Berg JW, Tilanus HW, Burgers JA, Wilson JH
and Dagnelie PC: Fatty acid composition of plasma lipids in
patients with pancreatic, lung and oesophageal cancer in comparison
with healthy subjects. Clin Nutr. 21:225–230. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cvetković Z, Vucić V, Cvetković B,
Petrović M, Ristić-Medić D, Tepsić J and Glibetić M: Abnormal fatty
acid distribution of the serum phospholipids of patients with
non-Hodgkin lymphoma. Ann Hematol. 89:775–782. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Macášek J, Vecka M, Žák A, Urbánek M,
Krechler T, Petruželka L, Staňková B and Zeman M: Plasma fatty acid
composition in patients with pancreatic cancer: Correlations to
clinical parameters. Nutr Cancer. 64:946–955. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pike LJ: Rafts defined: A report on the
keystone symposium on lipid rafts and cell function. J Lipid Res.
47:1597–1598. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kalyankrishna S and Grandis JR: Epidermal
growth factor receptor biology in head and neck cancer. J Clin
Oncol. 24:2666–2672. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Horiguchi A, Oya M, Shimada T, Uchida A,
Marumo K and Murai M: Activation of signal transducer and activator
of transcription 3 in renal cell carcinoma: A study of incidence
and its association with pathological features and clinical
outcome. J Urol. 168:762–765. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Horiguchi A, Oya M, Uchida A, Marumo K and
Murai M: Elevated Akt activation and its impact on
clinicopathological features of renal cell carcinoma. J Urol.
169:710–713. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kuroda K, Horiguchi A, Sumitomo M and
Asano T, Ito K, Hayakawa M and Asano T: Activated Akt prevents
antitumor activity of gefitinib in renal cancer cells. Urology.
74:209–215. 2009. View Article : Google Scholar : PubMed/NCBI
|