Introduction
Intestinal microbiota (~1014 microbial
cells/person), contribute to human health and are known as the most
important and complex ecosystem (1).
After birth, the composition of gut microbiota transforms gradually
from aerobic and facultative anaerobic bacteria into predominantly
obligate anaerobes. Therefore, early life is the most significant
period for the structural transformation of the intestinal flora
profile. In principle, the microbial ecosystem reaches a profile
characteristic of adult microbiota in composition and diversity by
the end of the first 3–5 years of life with changes in parameters
such as pH, oxygen content and nutritional consumption (2).
Microbial colonization has a crucial role not only
in the host's acquisition of nutrients and modulation of metabolism
but also in promoting immune system development and maturation. As
a portion of the mucosal barrier, microbiota are helpful in the
protection against intestinal epithelial injury and boost the
immune system to mature in the growth of mucosa-associated lymphoid
tissue, the amount of secreted immunoglobulin A (sIgA), regulation
of cytokines and expansion of T-cell populations. T-helper cells
(Th) have a critical role in amplifying immune responses with Th1
and Th2 cells driving ‘cellular immunity’ and ‘humoral immunity’ of
the adaptive immunity system. Th-cell populations may be classified
by the cytokines they secrete. Th1 cells are heavily reliant on
tumor necrosis factor-γ (IFN-γ) and interleukin-12 (IL-12), while
Th2 cells are characterized by the cytokines IL-4 and IL-10
(3,4).
Except for the interaction between gut microbiota
and the host, which is reflected in the physiological function, it
has long been considered that disorders of the microbial system
have a direct or indirect association with various diseases
(intestinal as well as extra-intestinal), including irritable bowel
syndrome, inflammatory bowel disease, antibiotic-associated
diarrhea (AAD), autoimmune diseases and even central nervous system
disorders (5,6). In children and infants with chronic
diarrhea and allergic disease, a decrease in Lactobacillus
spp. and Bifidobacterium spp. is frequently observed
(7).
Probiotics, live microorganisms that confer a health
benefit on the host when administered clinically to inhabit
inflammatory reactions, abrogate disorders of the intestinal flora
(8), prevent and treat diarrhea and
allergy (9) and alleviate symptoms
of patients with ulcerative colitis (10). However, different species or even
different strains of the same species may have different functions
in the regulation of intestinal flora and development of the
intestinal immune system (11).
Clostridium butyricum has the capacity to produce large
amounts of short-chain fatty acids, which promote intestinal
absorption of water and sodium (12,13).
Studies have found that Clostridium butyricum has a
regulatory effect in the intestinal flora by increasing the amount
of Lactobacillus spp (14)
and preventing AAD (15). The
present study was performed to explore the influence of
Clostridium butyricum [China General Microbiological Culture
Collection Center (CGMCC) no. 0313-1] on the regulation of gut
microflora by assessing the colony counts of Enterococcus
spp., Enterobacter spp., Bifidobacterium spp. and
Lactobacillus spp. and on the development of the immune
system by detecting intestinal sIgA and serum cytokines IFN-γ,
IL-12, IL-4 and IL-10.
Materials and methods
Animals and reagents
A total of 30 female, pregnant BALB/c mice (age, 8
weeks; weight, 25–30 g; one day following pregnancy) were obtained
from the Laboratory Animal Center of Sichuan University. The animal
experiment was approved by the Ethics Committee of Sichuan
University. Clostridium butyricum (CGMCC no. 0313–1) was
provided by Beijing Eastern Baixin Biotechnology Co., Ltd.
(Beijing, China) and was diluted to a concentration of
5×108 colony-forming units (CFU)/0.5 ml with normal
saline. Azide oxygen agar medium, Eosion Methylene Blue agar medium
(EMB), Lactobacillus selection medium (LBS) and
Bifidobacterium medium (BBL) were obtained from LuQiao
Biotechnology Company (Taizhou, China) and were used to culture
Enterobacter spp., Enterococcus spp.,
Lactobacillus spp. and Bifidobacterium spp., respectively.
All media were sterilized at high pressure at 115°C for 20 min. The
sIgA, IL-12, IL-4, IL-10 and IFN-γ ELISA kits were purchased from
R&D Systems, Inc. (Minneapolis, MN, USA). Pilocarpine was
obtained from Shanghai Yuanye Biotechnology Co., Ltd., (Shanghai,
China).
Experimental design
The influence of Clostridium butyricum on the
regulation of gut microflora and immune system development was
assessed by observing and analyzing the level of four bacterial
faces (Enterococcus spp., Enterobacterium spp.,
Lactobacillus spp. and Bifidobacterium spp.), the
amount of intestinal sIgA, cytokines IFN-γ, IL-12 as biomarkers of
Th1 cells and IL-4 and IL-10 as markers of Th2 cells in offspring.
The day of birth was defined as postnatal day 1 (PD1). The mice
were weaned off on PD21, which is the natural time of weaning.
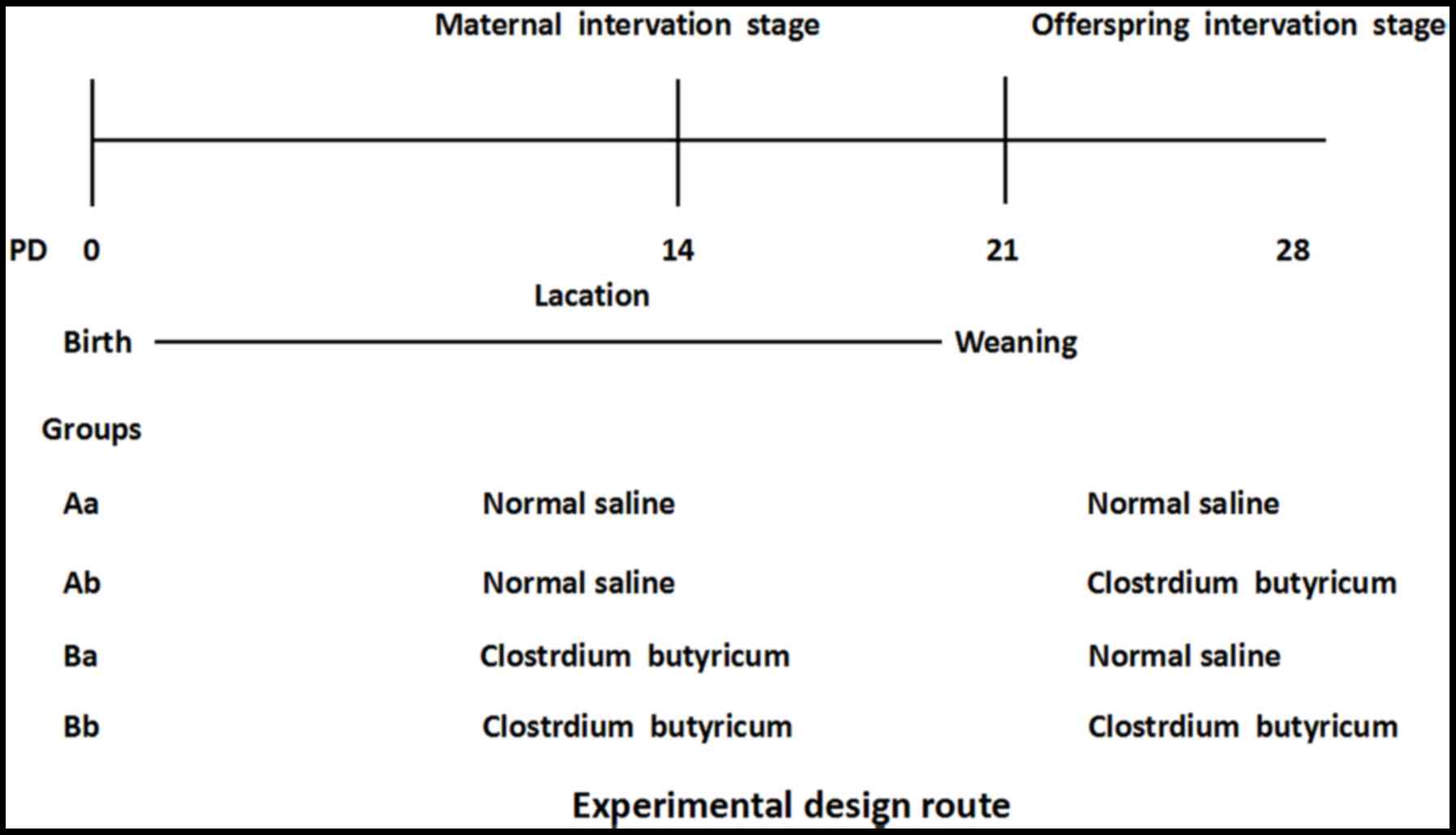
The experimental design is illustrated in Fig. 1. The thirty maternal mice and their
offspring were randomly divided into three groups following
successful delivery (n=10 mouse pups per group): The control group
(Aa), offspring intervention group (Ab), maternal intervention
group (Ba) and maternal and offspring intervention group (Bb). In
the Ba group, maternal mice were treated with Clostridium
butyricum by gavage at a dosage of 5×108 CFU per day
during lactation (from PD1 to PD20) and offspring were given 0.5 ml
normal saline by gavage per day after weaning (from PD21 to PD28).
In the Ab group, maternal mice were given 0.5 ml normal saline per
day by gavage during lactation and offspring were treated with
Clostridium butyricum by gavage at a dosage of
5×108 CFU per day after weaning. In the maternal and
offspring intervention group (Bb), both maternal mice and offspring
were supplemented with Clostridium butyricum at PD0-21 and
at PD21-28. In the control group, maternal mice were given 0.5 ml
normal saline per day by gavage during lactation and offspring were
administered the same amount of saline after weaning. Other
environmental factors and diet were kept consistent among the
groups. The mice were maintained in a 12 h light/dark cycle at a
temperature of 20–25°C in 40–70% humidity. They were given 1–3
g/day food and 4–7 ml/day water each. On PD14, −21 and −28, stool
specimens were collected from the offspring and aliquots were
individually inoculated into the four media mentioned above.
Samples in Azide oxygen agar medium and EMB medium were placed in
an incubator at 37°C and those in LBS and BBL media were placed
into an anaerobic box for bacterial culture at 37°C. Finally,
bacteria were quantified after 48 h. Similarly, sIgA in the
intestinal fluid and serum cytokine (IL-12, IL-4, IL-10 and IFN-γ)
levels were detected using the corresponding ELISA kits at PD14,
−21 and −28. A total of 0.2 ml serum was obtained from blood
obtained from the inner canthal vein. To determine sIgA, intestinal
fluid was gathered by intraperitoneal injection of 10 mg/kg
pilocarpine after administration of an enema without anesthesia as
previously described (16).
Statistical analysis
The numbers of trials between two groups were
analyzed by using an independent-samples t test and the differences
among multiple groups were analyzed by one-way analysis of variance
(ANOVA). A Student-Newman-Keul-Q post hoc test was then used for
individual comparisons when the ANOVA analysis indicated a
significant difference among groups. SPSS, version 18.0 (SPSS,
Inc., Chicago, IL, USA) was used. Values are expressed as the mean
± standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
Clostridium butyricum supplementation
affects the intestinal flora in mouse pups
To investigate the role of Clostridium butyricum in
modulating the gut flora, four common bacterial strains of the
intestine, Enterococcus spp., Enterobacter spp.,
Lactobacillus spp. and Bifidobacterium spp., were
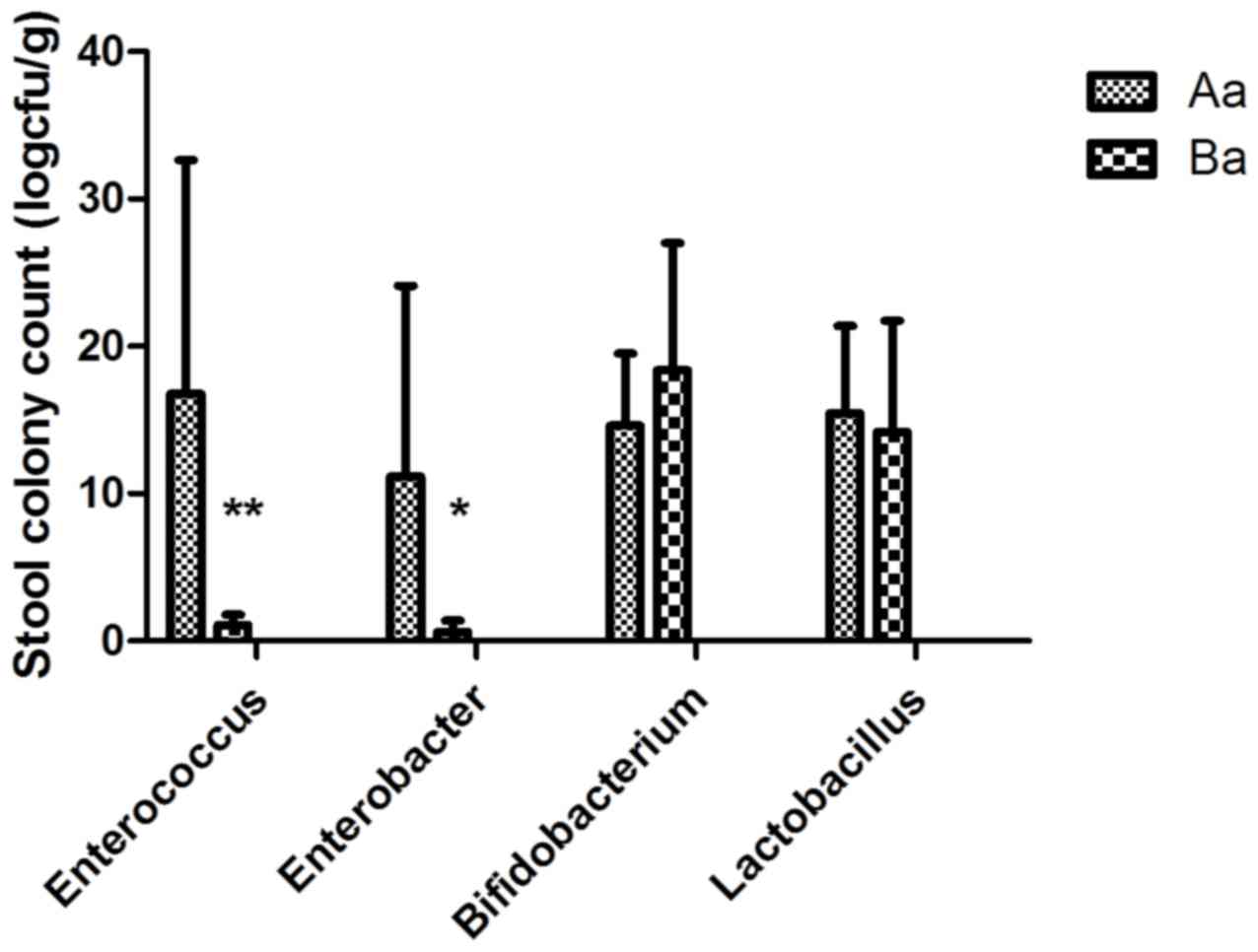
assessed. As revealed in Fig. 2,
supplementation of breast-feeding maternal mice with Clostridium
butyricum (Ba group) significantly decreased the quantity of
intestinal Enterococcus spp. (t=3.123, P<0.01) and
Enterobacter spp. (t=2.563, P<0.05) at PD14 in the
offspring compared with that in the control group. At PD21 the
count of intestinal Enterococcus spp. in the offspring of
the Ba group was further significantly decreased (t=2.292,
P<0.05) compared with the Aa control group, while the count of
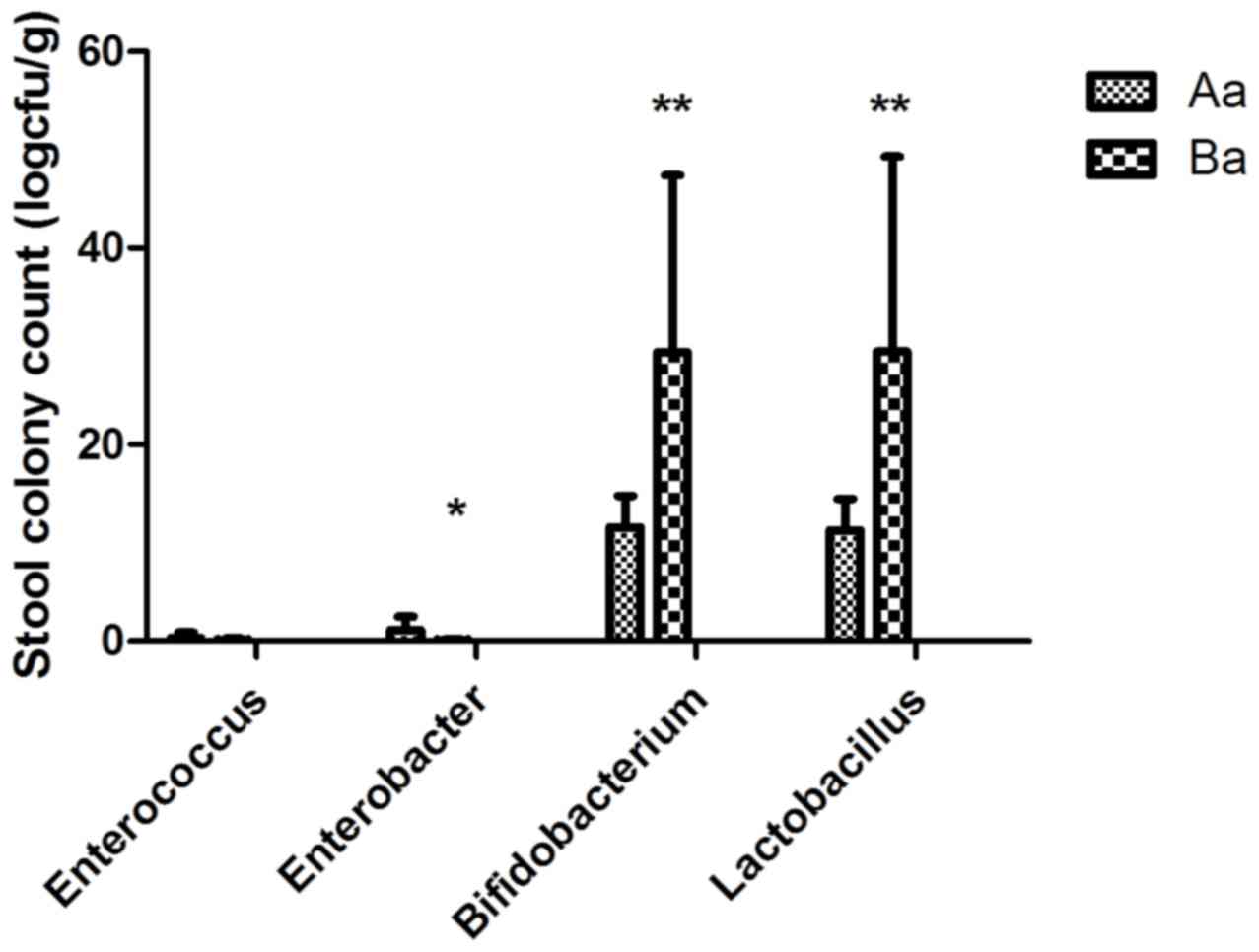
Enterobacter spp. was not (t=1.297, P>0.05; Fig. 3). Additionally, maternal
supplementation significantly increased the quantity of intestinal
Lactobacillus spp. (t=2.8508, P<0.01) and
Bifidobacterium spp. (t=3.085, P<0.01) in the offspring
of the Ba group at PD21 compared with that in the control group,
while it had no significant effect at PD14 (t=1.213, P>0.05 and
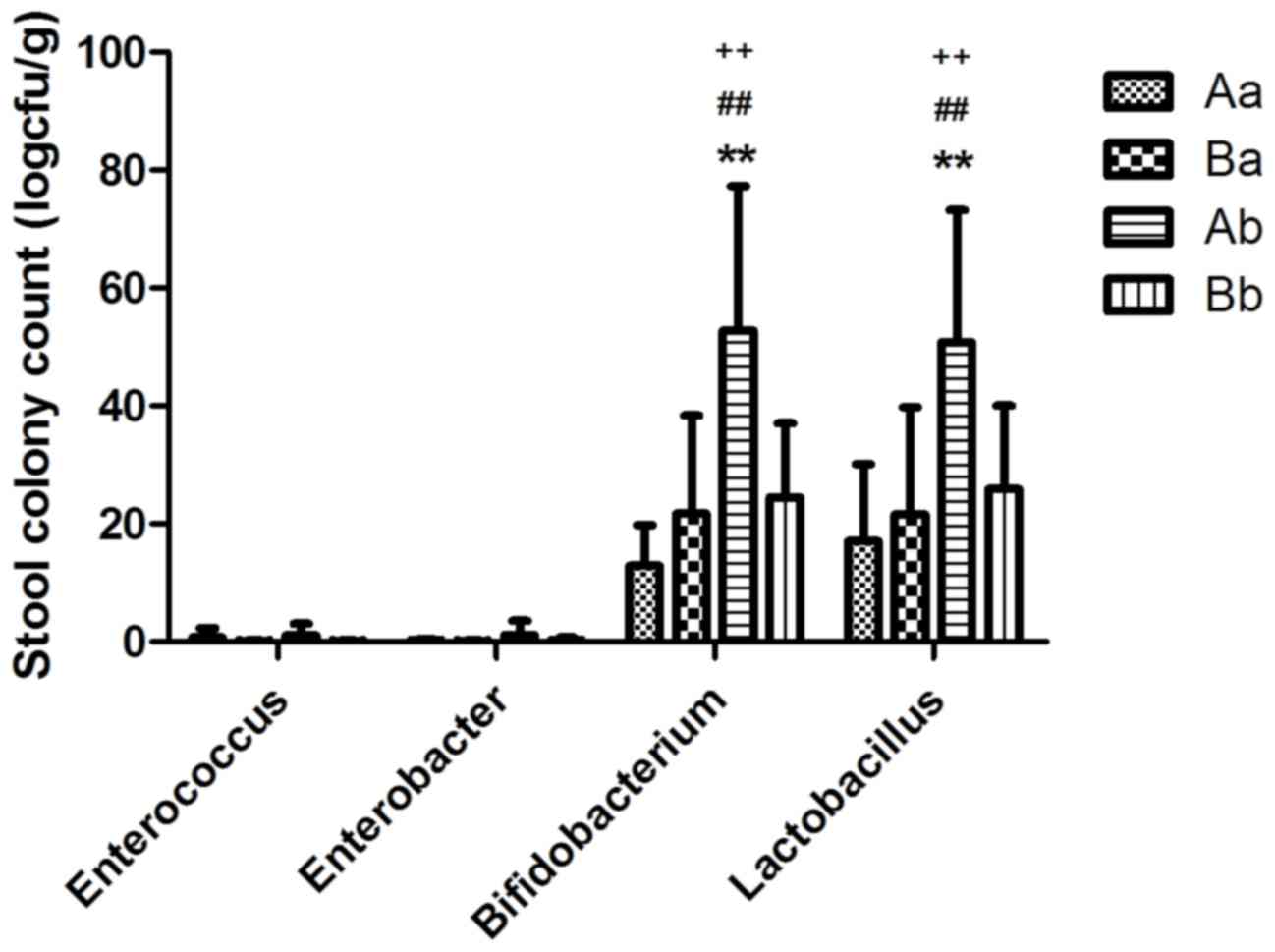
t=0.414, P>0.05 respectively). Following ablactation,
Bifidobacterium spp. and Lactobacillus spp. were
significantly increased in the Ab group compared with the control
group at PD28 (Q=7.679, P<0.01 and Q=6.149, P<0.01,
respectively), however no significant differences were observed in
the quantity of Enterococcus spp. (Q=0.973, P>0.05) and
Enterobacter spp. (Q=1.757, P>0.05) compared with the
control at the same time point (Fig.
4). Bifidobacterium spp. and Lactobacillus spp.
were also significantly increased in the Ab group compared with the
Ba and Bb group (P<0.01). These results suggest that the role of
breast milk on the diet of the offspring requires further
exploration.
Clostridium butyricum supplementation
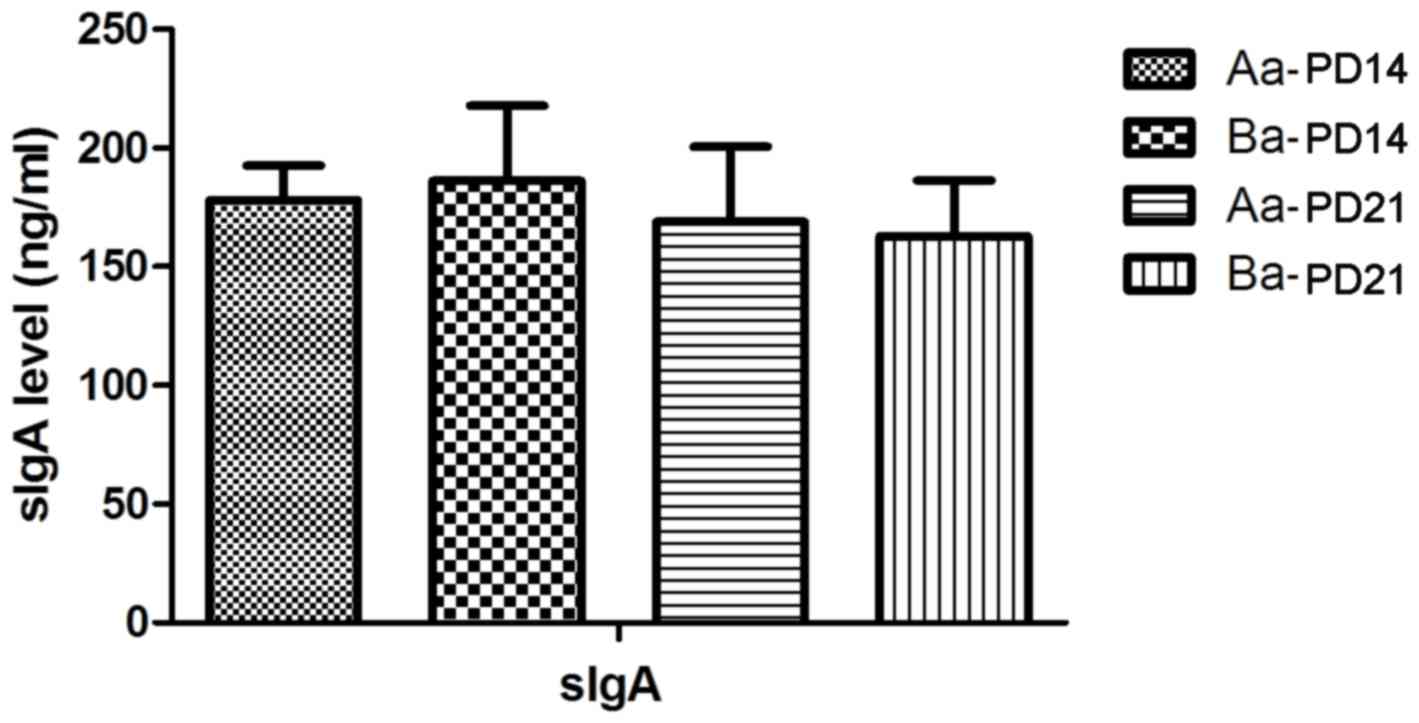
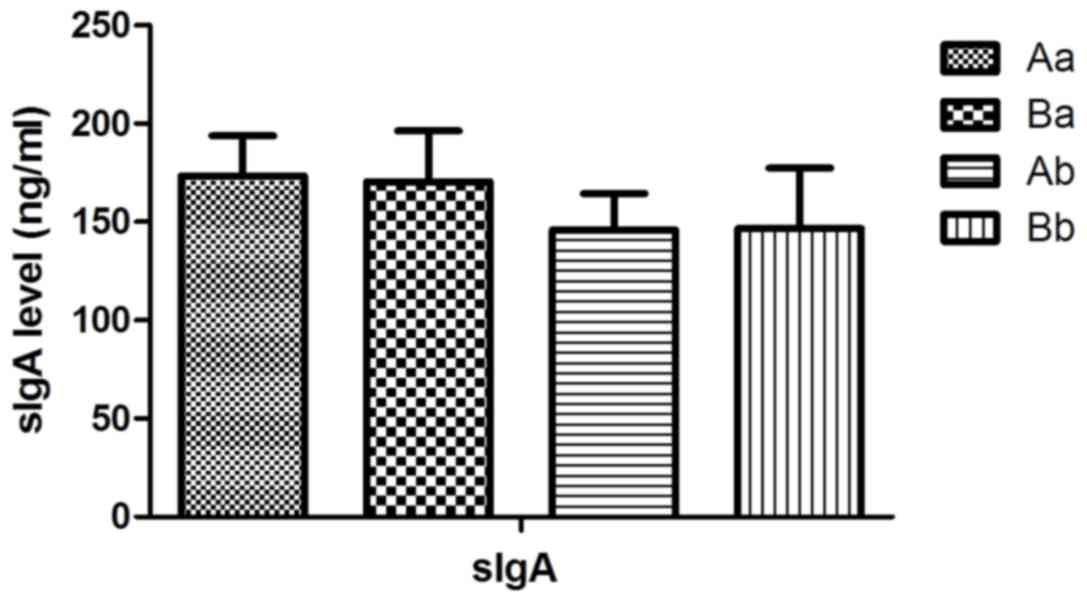
does not affect the intestinal secretion of sIgA in mouse pups
Next, the effects of Clostridium butyricum on
intestinal sIgA in mouse pups were assessed. No statistically
significant differences were observed between the intervention
groups and the control group at PD14 (t =−0.737, P>0.05;
Fig. 5), −21 (t=0.494, P>0.05;
Fig. 5) and −28
(QBa=0.380, P>0.05; QAb=3.564, P>0.05;
QBb=3.461, P>0.05; Fig.
6).
Effect of Clostridium butyricum on
cytokine secretion in offspring
To examine the effects of Clostridium butyricum
supplementation on developing immunity, circulating IL-12 and IFN-γ
were assessed as biomarkers for Th1 cells, while IL-4 and IL-10
served as biomarkers for Th2 cells. As revealed in Table I, a slight but insignificant increase
in the secretion of IL-12 and IFN-γ was detected in the Ba group at
PD21 compared with the control group. Similarly, no significant
differences were observed in the secretion of IL-4 and IL-10 at
PD21 compared with the control group. The Ab group also
demonstrated a slight but insignificant increase in IL-12 and IFN-γ
at PD28 compared with the control group.
 | Table I.Cytokine expression in offspring after
Clostridium butyricum supplementation. |
Table I.
Cytokine expression in offspring after
Clostridium butyricum supplementation.
| Group | Time-point | IFN-γ (pg/ml) | IL-12 (pg/ml) | IL-4 (pg/ml) | IL-10 (pg/ml) |
|---|
| Aa | PD 14 |
273.84±125.12 |
222.52±30.08 |
65.292±27.60 |
86.172±8.61 |
| Ba | PD 14 |
286.85±86.85 |
193.90±29.55 |
62.780±14.57 |
71.130±11.07 |
| Aa | PD 21 |
262.87±62.87 |
219.27±19.27 |
65.80±65.80 |
87.35±17.43 |
| Ba | PD 21 |
362.15±62.15 |
327.97±27.97 |
73.06±3.067 |
88.32±8.327 |
| Aa | PD 28 |
269.05±69.05 |
204.30±04.30 |
43.82±3.82 |
77.08±7.08 |
| Ba | PD 28 |
284.11±84.11 |
203.65±03.65 |
57.51±7.515 |
74.29±4.295 |
| Ab | PD 28 |
334.70±34.70 |
235.97±35.97 |
56.08±6.087 |
68.58±8.58 |
| Bb | PD 28 |
312.63±211.63 |
258.38±52.70 |
57.06±14.3 |
67.49±11.80 |
Discussion
The intestine is an important organ that maintains
the immunity and nutrition of the body, and it is also the biggest
bacterial storeroom in the body. During infancy, the intestinal
flora is not stable. In the present study, supplementation with
Clostridium butyrium reduced the colony count of
Enterobacter spp. and Enterococcus spp. and increased
the colony count of Lactobacillus spp. and
Bifidobacterium spp in the stool of the offspring. Moreover,
when the maternal mice were treated with Clostridium
butyrium by gavage, the decrease in intestinal
Enterobacter spp. and Enterococcus spp. at PD14 in
the offspring occurred earlier than the increase of
Lactobacillus spp. and Bifidobacterium spp. at PD21,
which was in line with the process of gut flora colonization. After
birth, the intestinal microbiota is dominated by aerobic and
facultative anaerobic bacteria such as Enterobacter spp. and
Enterococcus spp., which are then replaced by a complex
anaerobe-dominated flora, including Lactobacillus spp. and
Bifidobacterium spp., an anaerobe-dominated flora that
constitutes a portion of the mucosal barrier to protect against
pathogen invasion and maintain the balance of microbiota. Over the
first few weeks of life, changes in pH and oxygen content and
nutritional consumption occur, which are also affected by feeding
patterns, including breast or formula feeding. In the present
study, the composition and quantity of bacterial strains in the
intestinal flora of neonatal mice changed when the maternal mice
were treated with Clostridium butyrium during lactation. The
above results demonstrated that Clostridium butyrium affects
offspring through breast milk, while having a direct effect when
orally administered after weaning. This is in accordance with the
findings of a previous study on breast-fed subjects, which reported
that infants were able to acquire probiotics via breast milk
(17). Clostridium butyrium
does not colonize in the normal intestinal flora and keeps a
balance of the gut microflora without the Clostridium
butyrium population increasing (14). It may promote the accumulation and
development of beneficial bacteria such as Lactobacillus
spp. and Bifidobacterium spp. by producing low lactose.
A typical example for the association between
intestinal flora and the development of the host's immune system is
the study of germ-free animals in, which a distinct decrease of
Peyer's patches and a reduction of sIgA were found (18). Once germ-free animals were exposed to
bacteria constituting a normal intestinal flora, the development of
immune system occurred. Above of all, the intestinal flora has an
important influence on the normal immune system.
It is accepted that the gastrointestinal tract is
the largest immune organ due to the existence of gut-associated
lymphoid tissue, which has a crucial role in distinguishing
potentially pathogenic microbiota and harmless antigens, and
preventing the invasion of pathogens by intestinal epithelial cells
and sIgA in the innate immune system (19,20).
Once the first line of defense fails, the adaptive immune system
participates in the immune defense through cellular immunity and
humoral immunity mediated by Th1 and Th2 cells (3). Th1 and Th2 cells are differentiated
from the same precursor cell, the Th0 cell. In the process of
balancing between Th1 and Th2 cells, cytokines have an
irreplaceable role. For instance, IL-2, IL-12 and IFN-γ promote the
Th1 cell and inhibit Th2 cell formation, while IL-4 and IL-10
promote the Th2 cell and inhibit Th1 cell formation (21). Autoimmune and allergic diseases may
occur under circumstances where this balance is disturbed.
Furthermore, it has been indicated that an excessive Th1-associated
immune response is linked with autoimmune diseases such as
rheumatoid arthritis, multiple sclerosis and graft vs. host
disease. An excessive Th2-associated immune response has been
associated with allergic diseases such as asthma and atopic
dermatitis (22). Various probiotics
were demonstrated to have different roles in the host's immune
system. For instance, Lactobacillus paracasei (strain
KW3110) may potentially reduce allergy-induced inflammation by
inhibiting Th2 cytokines (11).
The present study demonstrated that the intestinal
sIgA content in mice offspring demonstrated no significant
differences when the maternal mice or their offspring were
supplemented with Clostridium butyrium during lactation,
which implys that the supplement may not affect the levels of sIgA.
Th1-associated cytokine activation was slightly increased in the
two intervention groups, Clostridium butyrium had no
significant effect on the secretion of cytokines. The capacity of
Clostridium butyrium supplementation to improve sIgA and
regulate the microbiotic flora in the intestine suggests that it
may aid in preventing the imbalance of Th1/Th2 and helping to
prevent the occurrence of autoimmune diseases. Clostridium
butyrium did not have a significant effect on intestinal sIgA
content and cytokine secretion, which implies that it may not
directly disrupt the immune systems of mice. In conclusion,
supplementation with Clostridium butyrium (CGMCC no. 0313-1)
regulated the balance of the intestinal floral, manifesting in
decreased colony counts of Enterococcus spp. and
Enterobacterium spp., and increased colony counts of
Lactobacillus spp. and Bifidobacterium spp. in
neonatal mice. Clostridium butyrium may also enhance the
capacity of intestinal resistance to infections by increasing the
secretion of sIgA. However, intervention with Clostridium
butyricum did not significantly affect the balance of Th1/Th2.
Overall, Clostridium butyricum supplementation may be a safe
method for regulating the establisment of the intestinal flora and
acquired immune system development and should be explored in
greater depth.
Acknowledgements
The present study was financially supported by the
Science and Technology Project of Sichuan Province of China for
(no. 2012JY0008). The authors would like to thank the Campus Public
Health Center of Sichuan University (Chengdu, China) for counting
the microbial culture technology and helping with the data
interpretation.
References
|
1
|
Schuijt TJ, van der Poll T, de Vos WM and
Wiersinga WJ: The intestinal microbiota and host immune
interactions in the critically ill. Trends Microbiol. 21:221–229.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rodriguez JM, Murphy K, Stanton C, Ross
RP, Kober OI, Juge N, Avershina E, Rudi K, Narbad A, Jenmalm MC, et
al: The composition of the gut microbiota throughout life, with an
emphasis on earlylife. Microb Ecol Health Dis.
26:260502015.PubMed/NCBI
|
|
3
|
Kidd P: Th1/Th2 balance: The hypothesis,
its limitations, and implications for health and disease. Altern
Med Rev. 8:223–246. 2003.PubMed/NCBI
|
|
4
|
Fujiwara D, Inoue S, Wakabayashi H and
Fujii T: The anti-allergic effects of lactic acid bacteria are
strain dependent and mediated by effects on both Th1/Th2 cytokine
expression and balance. Int Arch Allergy Immunol. 135:205–215.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vandenplas Y: Healthy gut microbiota and
long term health. Benef Microbes. 6:173–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bustos Fernandez LM, Lasa JS and Man F:
Intestinal microbiota: Its role in digestive diseases. J Clin
Gastroenterol. 48:657–666. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Björkstén B, Naaber P, Sepp E and
Mikelsaar M: The intestinal microflora in allergic Estonian and
Swedish 2-year-old children. Clin Exp Allergy. 29:342–346. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Valsecchi C, Marseglia A, Montagna L,
Tagliacarne SC, Elli M, Licari A, Marseglia GL and Castellazzi AM:
Evaluation of the effects of a probiotic supplementation with
respect to placebo on intestinal microflora and secretory IgA
production, during antibiotic therapy, in children affected by
recurrent airway infections and skin symptoms. J Biol Regul Homeost
Agents. 28:117–124. 2014.PubMed/NCBI
|
|
9
|
Edwards CA and Parrett AM: Intestinal
flora during the first months of life: New perspectives. Br J Nutr.
88 Suppl 1:S11–S18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoshimatsu Y, Yamada A, Furukawa R, Sono
K, Osamura A, Nakamura K, Aoki H, Tsuda Y, Hosoe N, Takada N and
Suzuki Y: Effectiveness of probiotic therapy for the prevention of
relapse in patients with inactive ulcerative colitis. World J
Gastroenterol. 21:5985–5994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsai YT, Cheng PC and Pan TM: The
immunomodulatory effects of lactic acid bacteria for improf oving
immune functions and benefits. Appl Microbiol Biotechnol.
96:853–862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakanishi S, Kataoka K, Kuwahara T and
Ohnishi Y: Effects of high amylose maize starch and Clostridium
butyricum on metabolism in colonic microbiota and formation of
azoxymethane-induced aberrant crypt foci in the rat colon.
Microbiol Immunol. 47:951–958. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scheppach W: Effects of short chain fatty
acids on gut morphology and function. Gut. 35 1 Suppl:S35–S38.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ichikawa H, Kuroiwa T, Inagaki A, Shineha
R, Nishihira T, Satomi S and Sakata T: Probiotic bacteria stimulate
gut epithelial cell proliferation in rat. Dig Dis Sci.
44:2119–2123. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seki H, Shiohara M, Matsumura T, Miyagawa
N, Tanaka M, Komiyama A and Kurata S: Prevention of
antibiotic-associated diarrhea in children by Clostridium butyricum
MIYAIRI. Pediatr Int. 45:86–90. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Küster T, Zumkehr B, Hermann C, Theurillat
R, Thormann W, Gottstein B and Hemphill A: Voluntary ingestion of
antiparasitic drugs emulsified in honey represents an alternative
to gavage in mice. J Am Assoc Lab Anim Sci. 51:219–223.
2012.PubMed/NCBI
|
|
17
|
Mitsou EK, Kirtzalidou E, Oikonomou I,
Liosis G and Kyriacou A: Fecal microflora of Greek healthy
neonates. Anaerobe. 14:94–101. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Falk PG, Hooper LV, Midtvedt T and Gordon
JI: Creating and maintaining the gastrointestinal ecosystem: What
we know and need to know from gnotobiology. Microbiol Mol Biol Rev.
62:1157–1170. 1998.PubMed/NCBI
|
|
19
|
Purchiaroni F, Tortora A, Gabrielli M,
Bertucci F, Gigante G, Ianiro G, Ojetti V, Scarpellini E and
Gasbarrini A: The role of intestinal microbiota and the immune
system. Eur Rev Med Pharmacol Sci. 17:323–333. 2013.PubMed/NCBI
|
|
20
|
Kaetzel CS: Cooperativity among secretory
IgA, the polymeric immunoglobulin receptor, and the gut microbiota
promotes host-microbial mutualism. Immunol Lett. 162:10–21. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
de Jong R, Brouwer M, Kuiper HM, Hooibrink
B, Miedema F and van Lier RA: Maturation- and
differentiation-dependent responsiveness of human CD4+ T helper
subsets. J Immunol. 149:2795–2802. 1992.PubMed/NCBI
|
|
22
|
Pessi T, Sütas Y, Hurme M and Isolauri E:
Interleukin-10 generation in atopic children following oral
Lactobacillus rhamnosus GG. Clin Exp Allergy. 30:1804–1808. 2000.
View Article : Google Scholar : PubMed/NCBI
|