Introduction
Syphilis is a sexually transmitted disease with a
complex and long course. The annual worldwide incidence of syphilis
has increased from 8.4% in 2009 to 11.2% in 2014 (1). The infection and incidence rates of
syphilis in China also demonstrated a linear upward trend, and
ranked third among all infectious diseases, after viral hepatitis
and pulmonary tuberculosis (2,3).
Treponema pallidum (Tp), the pathogen of syphilis, has a
strong invasive ability and enters the human body through intact
mucosa or damaged skin at an early stage of the infection. The
local mucosal effects of the body serve an important role against
Tp infection at the early stages. Enhancing the effective
stimulation of the mucosal immune system by vaccination and
immunization strategies may more effectively induce the generation
of protective immunity effect in the body; however, this process
requires a suitable mucosal immune carrier.
CpG-oligodeoxynucleotides (CpG-ODN) exhibit a strong
mucosal adjuvant activity, and is able to promote mucosal immunity
and enhance immunogenicity. As a promising mucosal adjuvant,
CpG-ODN has a synergistic effect with conventional mucosal adjuvant
cholera toxin (CT) and heat labile enterotoxin (LT) and causes no
toxic or side effects (4–6). Following primary immunization with
nucleic acid vaccines, enhanced immunization with corresponding
protein vaccines increases the immune response level and protective
immunity effect (7–11).
Previous studies have demonstrated that
intramuscular primary immunization by nucleic acid vaccine
pcDNA/glycerophosphodiester phosphodiesterase-interleukin-2
(pcDNA/Gpd-IL-2) with mucosal adjuvant CpG-ODN via intranasal
immunization does not significantly protect against Tp infection
compared with multiple intramuscular immunization groups (12,13).
Based on the findings of these studies the present study aimed to
evaluate the immune effect of intramuscular primary immunization by
nucleic acid vaccine pcDNA/Gpd-interleukin-2 (pcDNA/Gpd-IL-2) and
enhanced immunization via nasal feeding with the combination of
mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant protein on skin
infection caused by Tp in New Zealand rabbits.
Materials and methods
Bacterial strain and plasmids
CpG ODN (5′-TCCATGACGTTCCTGACGTT-3′) was
phosphorylated and synthesized by Invitrogen; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Recombinant pcDNA/Gpd-IL-2
plasmid and empty plasmid pcDNA3.1(+) (Thermo Fisher Scientific,
Inc.) were purified, dissolved in sterile phosphate-buffered saline
(PBS) and adjusted to a concentration of 0.2 mg/ml. Recombinant
pET28a/Gpd and pET28a/Gpd-IL-2 plasmids (constructed and stored
successfully at the Pathogenic Biology Institute at University of
South China) were transformed into BL21(DE3) strain following a
previously published protocol (14).
Following expression and purification of Gpd and Gpd-IL-2
recombinant proteins, their concentrations were adjusted to 0.1
mg/ml according to methods described previously (14).
Animals
A total of 108 female New Zealand rabbits (weight,
3.0–3.5 kg; age, 240 days; Department of Laboratory Animals,
University of South China, Hengyang, China) were raised under a
temperature of 18–20°C with 60–65% humidity, a normal 12 h
light/dark cycle, and given every morning and evening (~40 grams
each time) and free access to water without antibiotics. The
rabbits were randomly divided into six groups of 18 animals each
(Table I). At 2 days prior to
immunization, the muscle of inoculation was injected with 0.25%
bupivacaine hydrochloride (0.075 mg/kg; cat no. CAS18010-40-7;
ApexBiotechnology Corp., Taiwan). According to previous studies
(12,13), the vaccines and empty plasmids were
inoculated into the left hind leg quadriceps of rabbits via muscle
multi-point injection for primary immunization, and the
immunization was strengthened via nasal feeding with the
combination of mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant
protein 2 weeks later (Table I). At
week 8 after immunization, 3 rabbits from each group were subjected
to rabbit spleen cell proliferation test, cytokine detection and
secretory immunoglobulin A (SIgA) determination from nasopharyngeal
and vaginal mucosa samples. The remaining 15 rabbits in each group
were used in the syphilis infection model experiments and
protective research. At week 10 following the primary immunization
(day 0 of infection), all rabbits in the experimental groups (n=15)
were inoculated with 105 Tp (Nichols strain) at 8 points
on their back. Every 3 days, alterations in the skin redness,
swelling and ulceration were recorded by calculating the
erythematous diameter for 0–60 days. At day 21 following skin
infection, a positive rate of Tp in the skin lesions and the ulcer
formation rate were examined using dark field (DF)
illuminations/microscopy (cat no. BM-14; Shanghai BM Optical
Instruments Manufacture Company Ltd., Shanghai, China;
magnification, ×1,600) and modified silver staining (Nikon ECLIPSE
E100 light microscope, Nikon Corp., Tokyo, Japan; magnification,
×1,000) at 80°C for 30 sec. All animal experiments were approved by
the Governing Animal Welfare Committee of the University of South
China and conducted in accordance with the regulations of the
institution.
 | Table I.Different vaccine preparations in the
different rabbit groups. |
Table I.
Different vaccine preparations in the
different rabbit groups.
| Group | No. | Immunization
strategy | pcDNA/Gpd-IL-2
vaccinea | Gpd-IL-2 recombinant
proteina | CpG-ODN
adjuvanta |
|---|
| A1 | 18 | Primary: pcDNA3.1
control (im) | 100 µg (pcDNA) | – | – |
|
|
| Secondary: pcDNA3.1
control (im) |
|
|
|
| A2 | 18 | Primary:
pcDNA/Gpd-IL-2 (im) | 100 µg | – | – |
|
|
| Secondary:
pcDNA/Gpd-IL-2 (im) |
|
|
|
| B1 | 18 | Primary:
pcDNA/Gpd-IL-2 (im) | 100 µg | – | – |
|
|
| Secondary:
pcDNA/Gpd-IL-2 (nasal) |
|
|
|
| B2 | 18 | Primary:
pcDNA/Gpd-IL-2 (im) | 100 µg | – | 10 µg |
|
|
| Secondary:
pcDNA/Gpd-IL-2+CpGODN (nasal) |
|
|
|
| C1 | 18 | Primary:
pcDNA/Gpd-IL-2 (im) | 100 µg | 50 µg | – |
|
|
| Secondary: Gpd-IL-2
protein (nasal) |
|
|
|
| C2 | 18 | Primary:
pcDNA/Gpd-IL-2 (im) | 100 µg | 50 µg | 10 µg |
|
|
| Secondary: Gpd-IL-2
protein+CpG ODN (nasal) |
|
|
|
Enzyme-linked immunosorbent assay
(ELISA) for the detection of Tp Gpd antibodies
At weeks 0 (immunization day), 2, 4, 6 and 8 after
primary immunization, 1 ml ear vein blood was collected from
rabbits in each experimental group (n=18) and the serum was
obtained by centrifugation at 37°C at 1,600 × g for 10 min for
ELISA testing (cat no. 20140312; J&L Biological, Shanghai,
China). Briefly, Tp Gpd recombinant protein was diluted with a
coating buffer at a ratio of 1:50 and used to coat 96-well ELISA
plates at 4°C overnight. Next, the plate was blocked with 10%
bovine serum albumin (BSA) and PBS at 37°C for 1 h. Subsequent to
washing with PBS for three times, gradient concentrations of rabbit
serum were added to the plate, using Tp-positive serum as a
positive control. Following incubation at 37°C for 1 h, the plate
was washed, followed by the addition of horseradish
peroxidase-labeled goat anti-rabbit IgG secondary antibody
(1:2,000; cat no. 20140521, Thermo Fisher Scientific, Inc.). After
incubation at 37°C for 1 h, the plate was washed with PBS again and
subjected to color development with 3,3′,5,5′-tetramethylbenzidine
(Shanghai Bioleaf Biotech Co., Ltd. Shanghai, China). The
absorbance was read using a Multiskan reader (Thermo Fisher
Scientific, Inc.) at 450 nm. Each sample was tested in
triplicate.
ELISA for the detection of cytokine
levels
At week 8 after immunization, 3 rabbits were
randomly selected and sacrificed from each group to collect spleen
specimens. The spleen was placed in precooled 5 ml D-Hank's
solution (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) and ground by a 200 mesh copper mesh to obtain
spleen cells. Following centrifugation at 37°C and 1,600 × g for 10
min, the supernatant was discarded, followed by addition of 10 ml
red blood cell lysis buffer (139.6 mM NH4Cl and 16.96 mM
Tris; pH 7.2). Next, the samples were incubated at in a water bath
for 10 min, followed by centrifugation at 1,600 × g and 37°C for 10
min. Subsequent to discarding the supernatant, RPMI-1640 medium
containing 10% fetal bovine serum (FBS) was added to resuspend the
cells for counting. Cells (6×105/well) were added into a
96-well plate and incubated at 37°C and 50 ml/l CO2 for
72 h. Purified Tp Gpd recombinant protein (20 µg/ml) was used as a
stimulating antigen, while concanavalin A (ConA; 10 µg/ml;
Sigma-Aldrich, Merck, Darmstadt, Germany) was used as the positive
control according to a previously described protocol (14). Wells containing cells that were not
stimulated by antigen were used as the negative control. The
supernatant of the cell culture medium was collected for
determining the concentrations of IL-2 and interferon (IFN)-γ using
ELISA kits (J&L Biological, cat nos. 20140502, 20140422).
ELISA for the determination of IgA
level
At week 8 following primary immunization, the
trachea of the rabbits was dissected and 500 µl PBS was used to
wash the nasopharynx. The washing fluid flowing through the
nasopharynx was collected, and the nasopharynx was washed for three
times to obtain 1,500 µl nasopharyngeal washing fluid. In order to
obtain vaginal washing fluid, 500 µl PBS was injected into the
vagina of fluid and the washing fluid was collected. A total of
1,500 µl vaginal washing fluid was obtained, and centrifuged at
1,600 × g and 37°C for 10 min. Purified protein was then diluted
with coating buffer at a ratio of 1:50, and used to coat a 96-well
ELISA plate at 4°C overnight. Next, the plate was blocked with 10%
BSA and PBS at 37°C for 1 h. Subsequent to washing with PBS for 5
min three times, the nasopharyngeal and vaginal washing fluids (100
µl/well) were added to the plate in duplicate. Following incubation
at 37°C for 1 h, the plate was washed with PBS, followed by
addition of a horseradish peroxidase-labeled goat anti-rabbit IgA
secondary antibody (1:2,000; cat no. 20140404, Thermo Fisher
Scientific, Inc.). Following incubation at 37°C for 1 h, the plate
was washed again and subjected to color development and reaction
termination. The absorbance at 492 nm was read using a Multiskan
reader (Thermo Fisher Scientific, Inc.). Each sample was tested in
triplicate.
MTT assay
At week 8 following immunization, spleen cell
suspension was diluted with RPMI-1640 medium containing 10% FBS to
a density of 4×106/ml. The cells (100 µl/well) were
added into a 96-well plate in triplicate. Purified Tp Gpd
recombinant protein was used as a stimulating antigen (8 µg/ml),
while ConA (5 µg/ml) was used as the positive control. Wells
containing cells that were not stimulated by antigen were used as
the negative control. Following incubation for 68 h, T cell
proliferation was determined using MTT assay and the absorbance was
obtained at 570 nm. The results were expressed in terms of the
stimulation index (SI), as follows: SI=absorbance of experimental
group/absorbance of control group.
Statistical analysis
The results were analyzed using SPSS statistical
software (version 19.0; IBM Corp., Armonk, NY, USA). The data are
expressed as the means ± standard deviation. One way analysis of
variance followed by a Tukey's test was used to analyze the
specific antibody titer, cytokine concentration and spleen cell SI
data. The positive rate of Tp infection and ulcer formation rate
were analyzed using χ2 test. P<0.05 indicated that
the difference was statistical significance.
Results
Immunization strategy increases the
level of specific IgG antibody in the serum of rabbits prior to the
establishment of the syphilis model
Following intramuscular primary immunization by
nucleic acid vaccine pcDNA/Gpd-IL-2 and enhanced immunization via
nasal feeding with a combination of mucosal adjuvant CpG-ODN and
Gpd-IL-2 recombinant protein, ELISA was performed to determine the
levels of specific IgG antibody in the serum of rabbits. The data
revealed that the levels of specific IgG antibody against Gpd of
the A2, B1, B2, C1 and C2 groups were significantly increased
compared with that of the A1 group on weeks 4, 6 and 8 after
immunization (P<0.001). Of note, the level of specific IgG
antibody against Gpd of C2 group was significantly higher in
comparison with those of A2, B1, B2 and C1 groups on weeks 4, 6 and
8 after immunization (P<0.05). In addition, the level of
specific IgG antibody against Gpd of B1 group was significantly
higher compared with those of A2, B2, C1 and C2 groups on weeks 4,
6 and 8 after immunization (P<0.05). However, the levels of
specific IgG antibody against Gpd in A2, B2 and C1 were not
significantly different from each other (P>0.05; Fig. 1). These results suggest that
intramuscular primary immunization by nucleic acid vaccine
pcDNA/Gpd-IL-2 and enhanced immunization via nasal feeding with the
combination of mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant
protein increased the level of specific IgG antibody in the serum
of the rabbits.
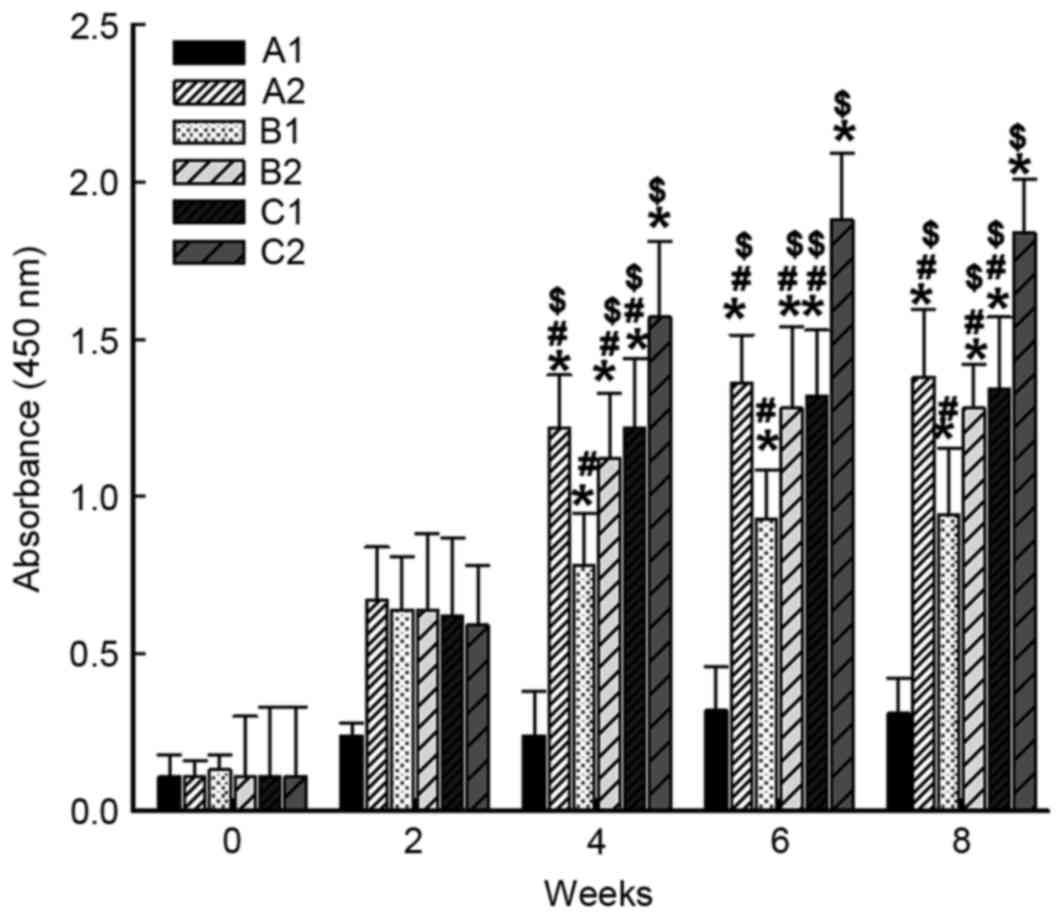 | Figure 1.Alterations in the levels of IgG
antibodies in the serum of experimental rabbits following
immunization. At 0, 2, 4, 6, and 8 weeks after immunization, blood
was collected from the rabbit ear veins and the anti-Tp Gpd
antibody levels in the serum were determined by enzyme-linked
immunosorbent assay. Absorbance was measured at 450 nm. *P<0.05
vs. A1 group at the same time point; #P<0.05 vs. C2
group at the same time point; $P<0.05 vs. B1 group at
the same time point. A1, 2× intramuscular immunization with
pcDNA3.1 (100 µg); A2, 2× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg); B1, 1× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with pcDNA/Gpd-IL-2
(100 µg); B2, 1× intramuscular immunization with pcDNA/Gpd-IL-2
(100 µg) + 1× nasal immunization with pcDNA/Gpd-IL-2 (100 µg) and
CpG-ODN (10 µg); C1, 1× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with Gpd-IL-2
recombinant protein (100 µg); C2, 1× intramuscular immunization
with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with Gpd-IL-2
recombinant protein (100 µg) and CpG-ODN (10 µg); ODN,
oligodeoxynucleotides; TP, Treponema pallidum; IL-2,
interleukin-2; Gpd, glycerophosphodiester phosphodiesterase. |
Immunization strategy elevates the
concentrations of IFN-γ and IL-2 in the supernatant of spleen
cells
To measure the concentrations of IFN-γ and IL-2 in
the supernatant of spleen cell culture, ELISA was employed. The
data demonstrated that the concentrations of IFN-γ and IL-2 in the
A2, B1, B2, C1 and C2 groups were significantly higher as compared
with that in the A1 group on week 8 after immunization (P<0.05).
The concentration of IL-2 in B1 group was not significantly
different from those in B2 (t=14.867, P=0.414) or C1 (t=31.107,
P=0.101), but was significantly lower in comparison with that in
the A2 (t=121.967, P<0.05) and C2 (t=151.533, P<0.05) groups
(Fig. 2A). In addition, the
concentration of IFN-γ in B1 group was significantly lower when
compared with that in the A2 (t=85.267, P=0.002), B2 (t=68.467,
P=0.009), C1 (t=58.833, P=0.021) and C2 (t=118.467, P<0.05)
groups (Fig. 2B). Furthermore, the
concentrations of IFN-γ and IL-2 in C2 group were significantly
higher compared with those in the B2 and C1 groups (P<0.05), but
were not significantly different from those in A2 group (P>0.05;
Fig. 2A and B). These results
indicate that intramuscular primary immunization by nucleic acid
vaccine pcDNA/Gpd-IL-2 and enhanced immunization via nasal feeding
with the combination of mucosal adjuvant CpG-ODN and Gpd-IL-2
recombinant protein elevated the concentrations of IFN-γ and IL-2
in the supernatant of spleen cells obtained from rabbits.
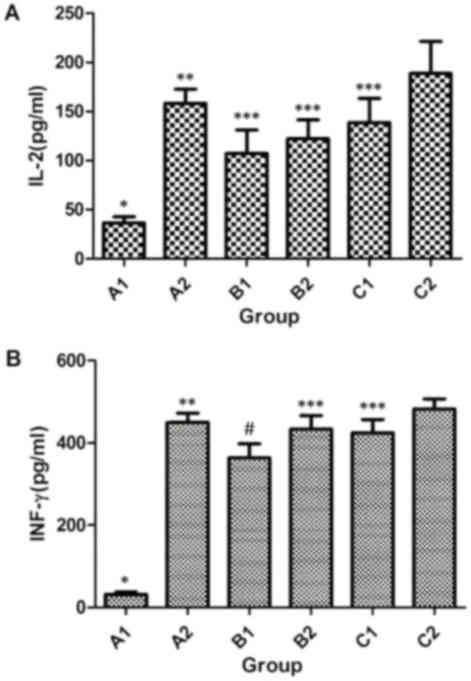 | Figure 2.Concentrations of cytokines (A) IL-2
and (B) IFN-γ secreted from spleen cells following Tp Gpd
stimulation in vitro. One-way analysis of variance was used
for statistical analysis, while pairwise comparison was also
performed among A1 to C2 groups. IL-2 (A1, A2, B1, B2, C1 and C2):
F=17.268, P<0.05; IFN-γ (A1, A2, B1, B2, C1 and C2): F=113.727,
P<0.05. A1, 2× intramuscular immunization with pcDNA3.1 (100
µg); A2, 2× intramuscular immunization with pcDNA/Gpd-IL-2 (100
µg); B1, 1× intramuscular immunization with pcDNA/Gpd-IL-2 (100 µg)
+ 1× nasal immunization with pcDNA/Gpd-IL-2 (100 µg); B2, 1×
intramuscular immunization with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal
immunization with pcDNA/Gpd-IL-2 (100 µg) and CpG-ODN (10 µg); C1,
1× intramuscular immunization with pcDNA/Gpd-IL-2 (100 µg) + 1×
nasal immunization with Gpd-IL-2 recombinant protein (100 µg); C2,
1× intramuscular immunization with pcDNA/Gpd-IL-2 (100 µg) + 1×
nasal immunization with Gpd-IL-2 recombinant protein (100 µg) and
CpG-ODN (10 µg); TP, Treponema pallidum; IL-2,
interleukin-2; IFN-γ, interferon-γ; Gpd, glycerophosphodiester
phosphodiesterase. *P<0.05 vs. A2/B1/B2/C1/C2; **P<0.05 vs.
B1; ***P<0.05 vs. C2; #P<0.05 vs. B2/C1/C2. |
Immunization strategy promotes the
proliferation of spleen cells
In order to examine the proliferation of T cells
following immunization, an MTT assay was conducted. The data
indicated that the SI of rabbit spleen cells in the A2, B1, B2, C1
and C2 groups were significantly higher compared with that in the
A1 group on week 8 after immunization (P<0.001). The SI of
rabbit spleen cells in the A2 group was significantly higher than
that in the B1 group (t=1.1067, P<0.05), although it was not
significantly different from those in B2 (t=0.0.1867, P=0.442), C1
(t=0.4000, P=0.114) or C2 (t=0.2267, P=0.353) groups (Fig. 3). In addition, the SI of rabbit
spleen cells in the B1 group was significantly lower in comparison
with those of the B2, C1 and C2 groups (P<0.05). By contrast,
the SI of rabbit spleen cells in the B2 group was not significantly
different from those of the C1 and C2 groups (P>0.05). In the C2
group, the SI of rabbit spleen cells was significantly higher than
that of C1 group (P<0.05; Fig.
3). These results suggest that intramuscular primary
immunization by nucleic acid vaccine pcDNA/Gpd-IL-2 and enhanced
immunization via nasal feeding with the combination of mucosal
adjuvant CpG-ODN and Gpd-IL-2 recombinant protein promoted the
proliferation of spleen cells.
 | Figure 3.SI of spleen cells. One-way analysis
of variance was used for statistical analysis, while pairwise
comparison was performed among A1 to C2 groups. SI (A1, A2, B1, B2,
C1 and C2): F=74.675, P<0.05. A1, 2× intramuscular immunization
with pcDNA3.1 (100 µg); A2, 2× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg); B1, 1× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with pcDNA/Gpd-IL-2
(100 µg); B2, 1× intramuscular immunization with pcDNA/Gpd-IL-2
(100 µg) + 1× nasal immunization with pcDNA/Gpd-IL-2 (100 µg) and
CpG-ODN (10 µg); C1, 1× intramuscular immunization with
pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with Gpd-IL-2
recombinant protein (100 µg); C2, 1× intramuscular immunization
with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with Gpd-IL-2
recombinant protein (100 µg) and CpG-ODN (10 µg); SI, stimulation
index. *P<0.05 vs. A2/B1/B2/C1/C2; **P<0.05 vs. B1;
***P<0.05 vs. B2/C1/C2; #P<0.05 vs. C2. |
Immunization strategy stimulates the
production of SIgA by the nasopharyngeal and vaginal mucosa
To investigate the levels of SIgA in the
nasopharyngeal and vaginal washing fluids, ELISA was used. The data
demonstrated that the levels of SIgA in the nasopharyngeal and
vaginal washing fluids obtained from A2, B1, B2, C1 and C2 groups
were significantly higher compared with those in the A1 group on
week 8 after immunization (P<0.05; Fig. 4). In addition, the levels of SIgA in
the B1, B2, C1 and C2 groups were significantly higher in
comparison with those in A2 group (P<0.05). Furthermore, the
levels of SIgA in the two washing fluids in B1 group were
significantly lower than those obtained from B2 or C2 groups
(P<0.05), but were not significantly different from the levels
in the C1 group (P>0.05). In B2 group, the levels of SIgA in the
fluids were significantly lower as compared with those in C2 group
(P<0.05), but were not significantly different from the levels
in C1 group (P>0.05). In addition, the levels of SIgA in
nasopharyngeal and vaginal washing fluids from C1 group were
significantly lower compared with those from C2 group (P<0.05;
Fig. 4). These results indicate that
intramuscular primary immunization by nucleic acid vaccine
pcDNA/Gpd-IL-2 and enhanced immunization via nasal feeding with the
combination of mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant
protein stimulated the production of SIgA by the nasopharyngeal and
vaginal mucosa.
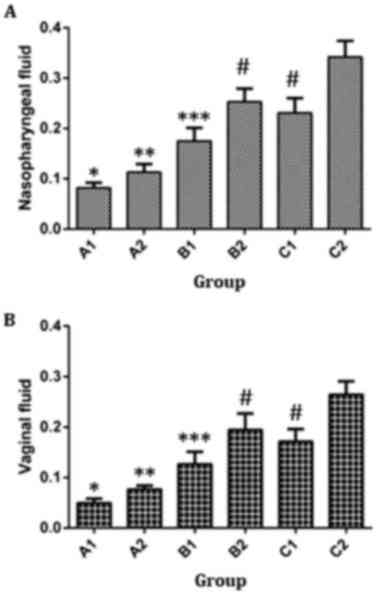 | Figure 4.Levels of SIgA antibody in (A)
nasopharyngeal and (B) vaginal fluid from immunized rabbits.
One-way analysis of variance was used for statistical analysis, and
pairwise comparison was performed among A1 to C2 groups. Levels of
SIgA antibody in nasopharyngeal fluid (A1, A2, B1, B2, C1 and C2):
F=46.703, P<0.05. Levels of SIgA antibody in vaginal fluid (A1,
A2, B1, B2, C1 and C2): F=113.679, P<0.05. A1, 2× intramuscular
immunization with pcDNA3.1 (100 µg); A2, 2× intramuscular
immunization with pcDNA/Gpd-IL-2 (100 µg); B1, 1× intramuscular
immunization with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization
with pcDNA/Gpd-IL-2 (100 µg); B2, 1× intramuscular immunization
with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization with
pcDNA/Gpd-IL-2 (100 µg) and CpG-ODN (10 µg); C1, 1× intramuscular
immunization with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization
with Gpd-IL-2 recombinant protein (100 µg); C2, 1× intramuscular
immunization with pcDNA/Gpd-IL-2 (100 µg) + 1× nasal immunization
with Gpd-IL-2 recombinant protein (100 µg) and CpG-ODN (10 µg);
SIgA, secretory immunoglobulin A. *P<0.05 vs. A2/B1/B2/C1/C2;
**P<0.05 vs. B1/B2/C1/C2; ***P<0.05 vs. B2/C2;
#P<0.05 vs. C2. |
Immunization strategy decreases the Tp
positive rate and ulcer formation rate at Tp infection sites
In order to compare the Tp positive rate and ulcer
formation rate among groups, χ2 test was performed. The
Tp DF positive rate and ulcer lesion formation rate in the A2, B1,
B2, C1 and C2 groups were significantly reduced compared with those
in A1 group (P<0.001). Tp DF positive rate in A2 group was
significantly higher compared with those in B2
(χ2=7.467, P=0.006) and C2 (χ2=24.220,
P<0.05) groups, but was not significantly different from those
in B1 (χ2=0.108, P=0.743) and C1 (χ2=2.092,
P=0.205) groups. However, ulcer lesion formation rate of the A2
group was only significantly higher than that of the C2 group
(χ2=9.808, P=0.002), and not significantly different
from those of the B1, B2 and C1 groups (P>0.05). In addition,
the Tp DF positive rate and ulcer lesion formation rate in the B1
group were significantly higher in comparison with those in the B2
and C2 groups (P<0.05), but were not significantly different
from those of the C1 group (P>0.05). However, the rates in B2
group were not significantly different from those of C1 group
(P>0.05). Furthermore, the Tp DF positive rate of the B2 group
was significantly higher compared with that of the C2 group
(P<0.05), whereas the ulcer lesion formation rate did not differ
significantly between these groups (P>0.05). In C1 group, the Tp
DF positive rate and ulcer lesion formation rate were both
significantly higher when compared with those of the C2 group
(P<0.05; Table II). These
results suggest that intramuscular primary immunization by nucleic
acid vaccine pcDNA/Gpd-IL-2 and enhanced immunization via nasal
feeding with the combination of mucosal adjuvant CpG-ODN and
Gpd-IL-2 recombinant protein decreased the Tp positive rate and
ulcer formation rate at Tp infection sites.
| Table II.Challenge results for immunized
rabbits with Treponema pallidum (Nichols strain)
spirochetes. |
Table II.
Challenge results for immunized
rabbits with Treponema pallidum (Nichols strain)
spirochetes.
| Group | No. of rabbits | No. of
DF+ lesions/total (%)a |
P-valueb | No. of ulcerative
lesions/total (%) |
P-valueb |
|---|
| A1 | 15 | 110/120 (91.7) |
P<0.001c | 106/120 (88.3) |
P<0.001c |
| A2 | 15 | 22/120 (18.3) |
P<0.05d | 18/120 (15.0) |
P=0.002e |
| B1 | 15 | 24/120 (20.0) |
P<0.05d | 22/120 (18.3) |
P<0.001d |
| B2 | 15 | 8/120 (6.7) |
P<0.05e | 10/120 (8.3) |
|
| C1 | 15 | 14/120 (11.7) |
P<0.001e | 12/120 (10.0) |
P<0.05e |
| C2 | 15 | 0/120 (0.0) |
| 4/120 (3.3) |
|
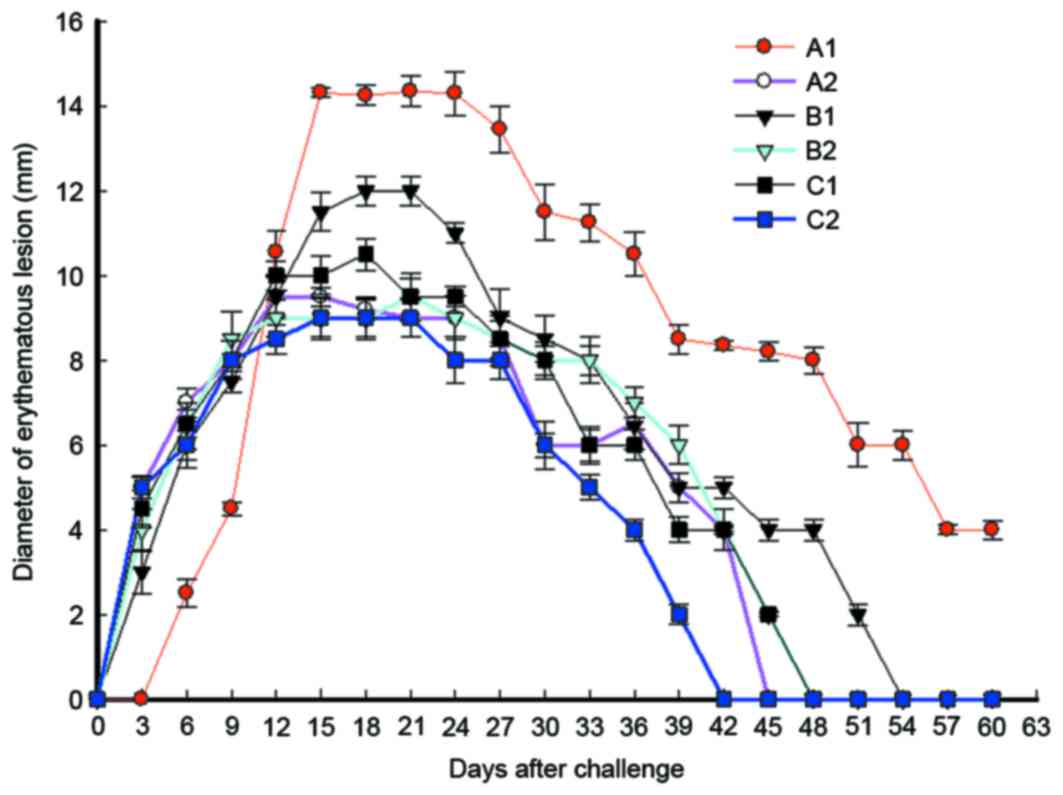
Immunization strategy reduces the
sizes of erythematous lesions in the shortest time following
immunization
To study the effect of immunization on swelling and
ulcer size at different time points in each group, the erythematous
diameter was calculated. The data revealed that erythema at skin
lesions in each group started to appear on day 3 after
immunization, with a diameter of 3–5 mm. The erythematous diameters
in B2 and C2 groups reached a peak of 9.5 and 9.13 mm,
respectively, on day 21, while erythema in the two groups
disappeared on days 48 and 42, respectively (Fig. 5). The erythematous diameters in A2
and C1 groups reached a peak of 9.54 mm and 10.52 mm on days 15 and
18, respectively, and the erythema in the two groups disappeared on
days 45 and 48, respectively. Furthermore, the erythematous
diameter in B1 group reached a peak of 12.13 mm on day 18, and the
erythema disappeared on day 54. Erythema in A1 group (pcDNA)
appeared on day 6, with a diameter of 2.5 mm, however, it was
enlarged quickly and reached a peak of 14.35 mm on day 15. Notably,
the erythematous diameter of A1 group remained at 4 mm on day 60,
and erythema disappeared after this time point (Fig. 5). These results indicate that
intramuscular primary immunization by nucleic acid vaccine
pcDNA/Gpd-IL-2 and enhanced immunization via nasal feeding with the
combination of mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant
protein reduced the sizes of erythematous lesions in the shortest
time following immunization compared with other immunization
strategies.
Discussion
Tp invades the body mainly through the urogenital
tract mucosa, while systemic humoral immunity and cellular immune
response are key mechanisms to prevent Tp diffusion and to finally
eradicate this bacterial infection (15,16).
Specific SIgA antigen in the local mucosa serves an important role
in the early resistance to Tp infection (17). Therefore, Tp vaccines should
stimulate both the systematic immunity and mucosal immunity.
Mucosal vaccines are able to induce mucosal immunity and systematic
immunity (18), and their
development is easier, quicker, more reliable and cheaper in
comparison with other vaccines (19). By contrast, conventional
intramuscular injection of a nucleic acid vaccine is only able to
induce a low level of local mucosal immunity (20).
Although enhanced immunization by nasal feeding
induces the production of a certain level of SIgA in the local
nasopharyngeal and vaginal mucosa, the expression of SIgA is
usually transient since mucosal epithelial cells typically present
a low ability for the uptake and expression of nucleic acids
(21). The results of the present
study demonstrated that this strategy leads to lower levels of
antibody responses and cytokines, as well as to higher levels of
SIgA secretion as compared with multiple intramuscular injection of
pcDNA/Gpd-IL-2. However, the Tp DF positive rate and ulcer
formation rate are not different between the two groups. These
observations suggest that SIgA is not the only factor against early
Tp infection of the skin.
CpG-ODN can be recognized by Toll-like receptor 9 on
dendritic cells and B-cells to induce the production of IFN-α and
chemotactic factors, as well as T-helper 1 cell immune response
(22). CpG-ODN has a target site,
namely pDCs, in mucosal vaccines (23), and it has been reported that CpG-ODN
can increase the production of antibodies by 15 times as a vaccine
adjuvant for hepatitis B vaccines (24). Consistently, the present study
observed that the use of CpG-ODN as an adjuvant (B2 group) enhanced
the levels of specific IgG antibody and cytokines, as well as the
secretion of SIgA by the nasopharyngeal and vaginal mucosa. In
addition, the Tp DF positive rate and ulcer formation rates in the
group where CpG-ODN was used as an adjuvant were reduced. These
results indicate that this immunization strategy induced strong
specific mucosal and systematic immunities, and enhanced the immune
protective effect of enhanced nucleic acid vaccine on early Tp
infection in the mucosa. This further suggests that SIgA is not the
only factor against mucosal Tp infection.
The procedure of immunization by vaccines is divided
into three stages, including the antigen recognition stage,
antigen-presenting cell activation stage, and antigen presenting
and humoral and/or cellular immune response stage (25). CpG-ODN is able to serve a synergistic
effect at all these three stages (26). The immunogenicity of protein
complexes and peptides has been reported to be weak, and adjuvant
is usually required to enhance the immune response (27). Combination of CpG-ODN with a protein
antigen promotes the uptake of antigens by tissues and cells, and
reduces the number of vaccinations required and immunization time
(28). In the present study, it was
demonstrated that the systematic immune effect in C2 group was
better when compared with that in A2 and C1 groups. In addition,
this strategy promoted the secretion of SIgA antibody that is
important in mucosal immunity. As a result, the DF positive rate
and ulcer positive rate in C2 group were lower in comparison with
those in A2 group, and the lesion healing time in C2 group was
shortened in comparison with the A2 group. This suggests that the
protective effect of the strategy used in the C2 group was better
than the strategies used in the A2 or C1 groups.
In conclusion, the present study demonstrated that
intramuscular primary immunization by nucleic acid vaccine
pcDNA/Gpd-IL-2 and enhanced immunization via nasal feeding with the
combination of mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant
protein was able to promote the immune protective effect of
pcDNA/Gpd-IL-2 nucleic acid vaccines on early mucosal infection by
Tp, and induce strong specific humoral immunity and cellular
immunity. However, the optimization of the delivery and release
system, as well as of the immunization methods, dosage, duration
and times, for Tp mucosal nucleic acid vaccines require further
investigation.
Acknowledgements
The present study was supported by grants from the
Preclinical Medicine Hunan Provincial Key Disciplines, Hunan
Provincial Key Laboratory for Special Pathogens Prevention and
Control (no. 2014-5), the Hunan Provincial Cooperative Innovation
Center for Molecular Target New Drug Study (no. 2014-405), the
National Natural Science Foundation (nos. 81373230, 81301470 and
81273322) and the Hunan Provincial Science and Technology
Department Foundation (no. 2014TT2025).
References
|
1
|
Shah BJ, Karia DR and Pawara CL: Syphilis:
Is it making resurgence? Indian J Sex Transm Dis. 36:178–181. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang L, Zeng L, Ren X, Geng M, Li Z and Yu
H: Analysis of morbidity and mortality characteristics of the
notifiable diseases reported in China. Zhonghua Liu Xing Bing Xue
Za Zhi. 36:194–198. 2015.(In Chinese). PubMed/NCBI
|
|
3
|
Zheng N, Guo Y, Padmadas S, Wang B and Wu
Z: The increase of sexually transmitted infections calls for
simultaneous preventive intervention for more effectively
containing HIV epidemics in China. BJOG. 121:35–44. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hiramatsu K, Serada S, Kobiyama K,
Nakagawa S, Morimoto A, Matsuzaki S, Ueda Y, Fujimoto M, Yoshino K,
Ishii KJ, et al: CpG oligodeoxynucleotides potentiate the antitumor
activity of anti-BST2 antibody. Cancer Sci. 106:1474–1478. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li HT, Zhang TT, Chen ZG, Ye J, Liu H, Zou
XL, Wang YH and Yang HL: Intranasal administration of CpG
oligodeoxynucleotides reduces lower airway inflammation in a murine
model of combined allergic rhinitis and asthma syndrome. Int
Immunopharmacol. 28:390–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iho S, Maeyama J and Suzuki F: CpG
oligodeoxynucleotides as mucosal adjuvants. Hum Vaccin Immunother.
11:755–760. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dai Y, Zhu Y, Harn DA, Wang X, Tang J,
Zhao S, Lu F and Guan X: DNA Vaccination by Electroporation and
Boosting with Recombinant Proteins Enhances the Efficacy of DNA
Vaccines for Schistosomiasis Japonica. Clin Vaccine Immunol.
16:1796–1803. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bae JY, Moon SH, Choi JA, Park JS, Hahn
BS, Kim KY, Kim B, Song JY, Kwon DH, Lee SC, et al: Recombinant DNA
and protein vaccines for foot-and-mouth disease induce humoral and
cellular immune responses in mice. Immune Netw. 9:265–273. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Endmann A, Klünder K, Kapp K, Riede O,
Oswald D, Talman EG, Schroff M, Kleuss C, Ruiters MH and Juhls C:
Cationic lipid-formulated DNA vaccine against hepatitis B virus:
Immunogenicity of MIDGE-Th1 vectors encoding small and large
surface antigen in comparison to a licensed protein vaccine. PLoS
One. 9:e1017152014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nchinda G, Amadu D, Trumpfheller C,
Mizenina O, Uberla K and Steinman RM: Dendritic cell targeted HIV
gag protein vaccine provides help to a DNA vaccine including
mobilization of protective CD8+ T cells. Proc Natl Acad Sci USA.
107:pp. 4281–4286. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li W, Wang S and Lu S: Pilot study on the
use of DNA priming immunization to Enhance Y. pestis LcrV-Specific
B cell responses elicited by a recombinant LcrV protein vaccine.
Vaccines (Basel). 2:36–48. 2014. View Article : Google Scholar
|
|
12
|
Zhao F, Wang S, Zhang X, Gu W, Yu J, Liu
S, Zeng T, Zhang Y and Wu Y: Protective efficacy of a Treponema
pallidum Gpd DNA vaccine vectored by chitosan nanoparticles and
fused with interleukin-2. Can J Microbiol. 58:117–123. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao F, Liu S, Zhang X, Yu J, Zeng T, Gu
W, Cao X, Chen X and Wu Y: CpG adjuvant enhances the mucosal
immunogenicity and efficacy of a Treponema pallidum DNA vaccine in
rabbits. Hum Vaccin Immunother. 9:753–760. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao Feijun, Zhang Xiaohong, Yu Jin, Liu
Shuangquan, Zhang Yuejun and Wu Yimou: Expression and
identification of Gpd-IL-2 recombinant protein of treponema
pallidum and its immuno-competence analysis. J Med Sci Central
South China. 40:131–135. 2012.(In Chinese).
|
|
15
|
Syphilis. Nurs Stand. 29:172015.
View Article : Google Scholar
|
|
16
|
Janier M, Hegyi V, Dupin N, Unemo M,
Tiplica GS, Potočnik M, French P and Patel R: 2014 European
guideline on the management of syphilis. J Eur Acad Dermatol
Venereol. 28:1581–1593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stamm LV and Drapp RL: A synthetic lymph
node containing inactivated Treponema pallidum cells elicits
strong, antigen-specifichumoral and cellular immune responses in
mice. Pathog Dis. 70:88–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Khera AK, Afkhami S, Lai R, Jeyanathan M,
Zganiacz A, Mandur T, Hammill J, Damjanovic D and Xing Z: Role of B
cells in mucosal vaccine-induced protective CD8+ T cell immunity
against pulmonary tuberculosis. J Immunol. 195:2900–2907. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stary G, Olive A, Radovic-Moreno AF,
Gondek D, Alvarez D, Basto PA, Perro M, Vrbanac VD, Tager AM, Shi
J, et al: VACCINES. A mucosal vaccine against Chlamydia trachomatis
generates two waves of protective memory T cells. Science.
348:aaa82052015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ye T, Yue Y, Fan X, Dong C, Xu W and Xiong
S: M cell-targeting strategy facilitates mucosal immune response
and enhances protection against CVB3-induced viral myocarditis
elicited by chitosan-DNA vaccine. Vaccine. 32:4457–4465. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Umthong S, Buaklin A, Jacquet A, Sangjun
N, Kerdkaew R, Patarakul K and Palaga T: Immunogenicity of a DNA
and recombinant protein vaccine combining LipL32 and Loa22 for
leptospirosis using chitosan as a delivery system. J Microbiol
Biotechnol. 25:526–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Birk R, Aderhold C, Hörmann K, Wenzel A,
Kramer B, Eschenhagen T and Sommer JU: CpG-Oligodeoxynucleotides in
Chronic Rhinosinusitis Cell Culture. In Vivo. 30:47–52.
2016.PubMed/NCBI
|
|
23
|
Meng Z, Zhang X, Pei R, Zhang E, Kemper T,
Vollmer J, Davis HL, Glebe D, Gerlich W, Roggendorf M, et al:
Combination therapy including CpG oligodeoxynucleotides and
entecavir induces early viral response and enhanced inhibition of
viral replication in a woodchuck model of chronic hepadnaviral
infection. Antiviral Res. 125:14–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiang XX, Zhou XQ, Wang JX, Xie Q, Cai X,
Yu H and Zhou HJ: Effects of CpG-ODNs on phenotype and function of
monocyte-derived dendritic cells in chronic hepatitis B. World J
Gastroenterol. 17:4825–4830. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lei J, Osen W, Gardyan A, Hotz-Wagenblatt
A, Wei G, Gissmann L, Eichmüller S and Löchelt M:
Replication-competent foamy virus vaccine vectors as novel epitope
scaffolds for immunotherapy. PLoS One. 10:e01384582015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
San R, Omán B, Gómez S, Irache JM and
Espuelas S: Co-encapsulated CpG oligodeoxynucleotides and ovalbumin
in PLGA microparticles; an in vitro and in vivo study. J Pharm
Pharm Sci. 17:541–553. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
De Cesare M, Sfondrini L, Pennati M, De
Marco C, Motta V, Tagliabue E, Deraco M, Balsari A and Zaffaroni N:
CpG-oligodeoxynucleotides exert remarkable antitumor activity
against diffuse malignant peritoneal mesothelioma orthotopic
xenografts. J Transl Med. 14:252016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang C, Zheng M, Zhu XH, Li S, Ni J, Li
BL, Liu H, Gao F and Cai JM: Protective effect of
CpG-oligodeoxynucleotides against low- and high-LET irradiation.
Cell Physiol Biochem. 34:1663–1674. 2014. View Article : Google Scholar : PubMed/NCBI
|