Introduction
Azoospermia, which affects ~1% of the male
population and may be responsible for up to 20% of male infertility
situations, is a medical condition in which the semen contains no
sperm (1–3). Azoospermia may be caused by a lack of
spermatozoa production in the testes (secretory or non-obstructive
azoospermia; NOA) or due to spermatozoa being unable to reach the
ejaculate (excretory or obstructive azoospermia; OA) (1). Previous studies have indicated that
genetic factors, including chromosomal abnormalities, are
responsible for azoospermia; however, few studies have explored the
association between microorganisms and azoospermia (1–3).
Microbiomes in the human skin, intestines, oral
cavity, vagina and urethra serve a key role in human health,
disease and other etiologies (4–6). It has
been reported that several bacteria, including Lactobacillus
iners, Gardnerella vaginalis, Escherichia
faecalis, E. coli and Staphylococcus aureus, are
associated with male infertility as demonstrated using polymerase
chain reaction (PCR) or culture-based methods (7–12).
However, the majority of previous studies focused on a few types of
bacteria and relied on qualitative analysis to discover
associations between semen microbiomes and semen quality; as such,
at present there is no comprehensive understanding of bacterial
communities in semen (13).
Recent technological advances in next generation
sequencing technology have enabled the pleiotropic effects of
microorganisms in the human host to be elucidated, providing an
excellent platform to explore the relationship between human and
semen microbiota (14–20). In the present study, the
high-throughput sequencing method was used to compare seminal
bacterial composition in patients with obstructive and
non-obstructive azoospermia. The results may be useful for
developing clinical therapies for the prevention and treatment of
azoospermia.
Materials and methods
Ethical statement
The present study was approved by the Ethical
Committee of the Jiangxi Maternal and Child Health Hospital
(Nanchang, China) and all participants provided written informed
consent. All experiments were carried out in accordance with the
guidelines provided by the Jiangxi Maternal and Child Health
Hospital.
Patients and sampling
Samples of seminal plasma were collected at the
Jiangxi Maternal and Child Health Hospital between October and
December 2016 from three groups of men without leukocytospermia as
follows: The control (C) group (22–36 years old), the obstructive
azoospermia (OA) group (24–38 years old) and the non-obstructive
azoospermia (NOA) group (25–37 years old). A total of 5 fertile
donors with normal sperm parameters (seminal volume, 2–6 ml; sperm
count, 0.6–1.5×1012/l; progressive motile spermatozoa
≥30% or progressive motile spermatozoa + motile spermatozoa ≥50%)
(4) were recruited as the C group. A
total of 12 patients with azoospermia who underwent surgical sperm
retrieval via percutaneous epididymal sperm aspiration or
testicular sperm extraction were recruited and were divided into
the OA group (n=6) and the NOA group (n=6). In OA, sperm are
produced normally inside the testicle, but are prevented from
reaching the ejaculate due to a blockage or obstruction in the
reproductive tubing. In non-obstructive azoospermia group, the
tubes are not blocked; however, there is a spermatogenesis problem
whereby there is a very low level of sperm production or a total
lack of production (8). Patients
with abnormal karyotypes and those who had previously suffered from
an injury to the genitals were excluded.
Prior to sampling, the glans penis of each patient
was washed using soap and water. Semen was obtained by
masturbation, ejaculated into a sterile collection tube and
incubated at 37°C for 30 min for liquefaction. Basic semen
parameters and leukocytospermia were detected as previously
described and the samples were stored at −80°C prior to DNA
extraction (20).
Extraction of genomic DNA and
high-throughput sequencing
Genomic DNA was extracted from each semen sample
using a TIANamp Genomic DNA kit (Tiangen Biotech Co., Ltd.,
Beijing, China) and combined with bead beating as previously
described (14). The concentration
and quality of extracted genomic DNA were tested prior to
sequencing using a spectrophotometer at 230 nm (A 260) and 260 nm
(A 260) (NanoDrop; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The extracted genomic DNA was used as a template to amplify
the V4 region of the 16S rRNA genes of all samples using 515F/806R
primers (515F, 5′-GTGCCAGCMGCCGCGGTAA-3′; 806R,
5′-GGACTACVSGGGTATCTAAT-3′). The PCR reaction conditions were as
follows: 2 min initial denaturation at 94°C and 30 cycles of 30 sec
denaturation at 94°C, 30 sec annealing at 52°C, 30 sec extension at
72°C, followed by a 5 min final extension at 72°C. The samples were
maintained at 4°C when the PCR reaction finished. Each 25 µl
reaction made up of 2.5 µl 10X Ex Taq buffer
(Mg2+ free; Takara Biotechnology Co., Ltd., Dalian,
China), 2 µl (2.5 mM) dNTP mixture, 1.5 µl Mg2+ (2.5
mM), 0.25 µl Ex-Taq DNA polymerase (Takara Biotechnology
Co., Ltd.), 0.5 µl barcode primer V4F (10 µM), 0.5 µl barcode
primer V4R252 (10 µM), 2 µl of fecal DNA and 15.75 µl milli-Q
water. These PCR products were sequenced using an Illumina HiSeq
2000 platform (21).
Bioinformatics and multivariate
statistics
Paired-end reads from the original DNA fragments
were merged using FLASH (version 1.2.7; ccb.jhu.edu/software/FLASH/) when any of the reads
overlapped that generated from the opposite end of the same DNA
fragment. Paired-end reads were assigned to each sample according
to the unique barcodes using Trimmomatic (version 0.33) (22). Sequence analysis was subsequently
performed using the UPARSE software package (version 7.0.100)
(23) with the UPARSE-OTU and Chao1,
the Chao1 estimator and the Shannon index (24) with UPARSE-OTUref algorithms. In-house
Perl scripts were used to analyze α (within samples) and β (among
samples) diversity. Sequences with ≥97% similarity were assigned to
the same OTUs. A representative sequence was selected for each OTU
and the RDP classifier (version 2.2) (25); was used to annotate taxonomic
information for each representative sequence. Cluster analysis was
preceded by weighted UniFrac distance using QIIME software package
(version 1.8.0) (26), partial least
squares discriminate analysis (PLS-DA) was preceded using SIMCA-P
software version 11.5 (Umetrics; Sartorius Stedim Biotech, Malmö,
Sweden) and the species that were differentially abundant were
characterized for their metabolic capacity using Phylogenetic
Investigation of Communities by Reconstruction of Unobserved States
(PICRUSt), Version 1.0.0 (27,28). The
statistical significance was set at P<0.05 for correction of
multiple comparisons.
Results
Seminal bacterial composition at
phylum level
To compare microbes in the OA and NOA groups, the V4
hypervariable region of bacterial genomic DNA was sequenced using
high-throughput sequencing and the effective tags with ≥97%
similarity were considered as one OTU. In total, 675,539 usable
sequences and 5,867 OTUs were obtained from the samples, with a
mean of 345 OTUs per group (data not shown). Furthermore, the Chao1
and Shannon indices were nearly saturated and the rarefaction curve
of every sample entered the plateau phase, which indicated that the
sequencing data was reasonable (OTU number was sufficient for
further analysis) and species composition was highly uniform in
each group (data not shown).
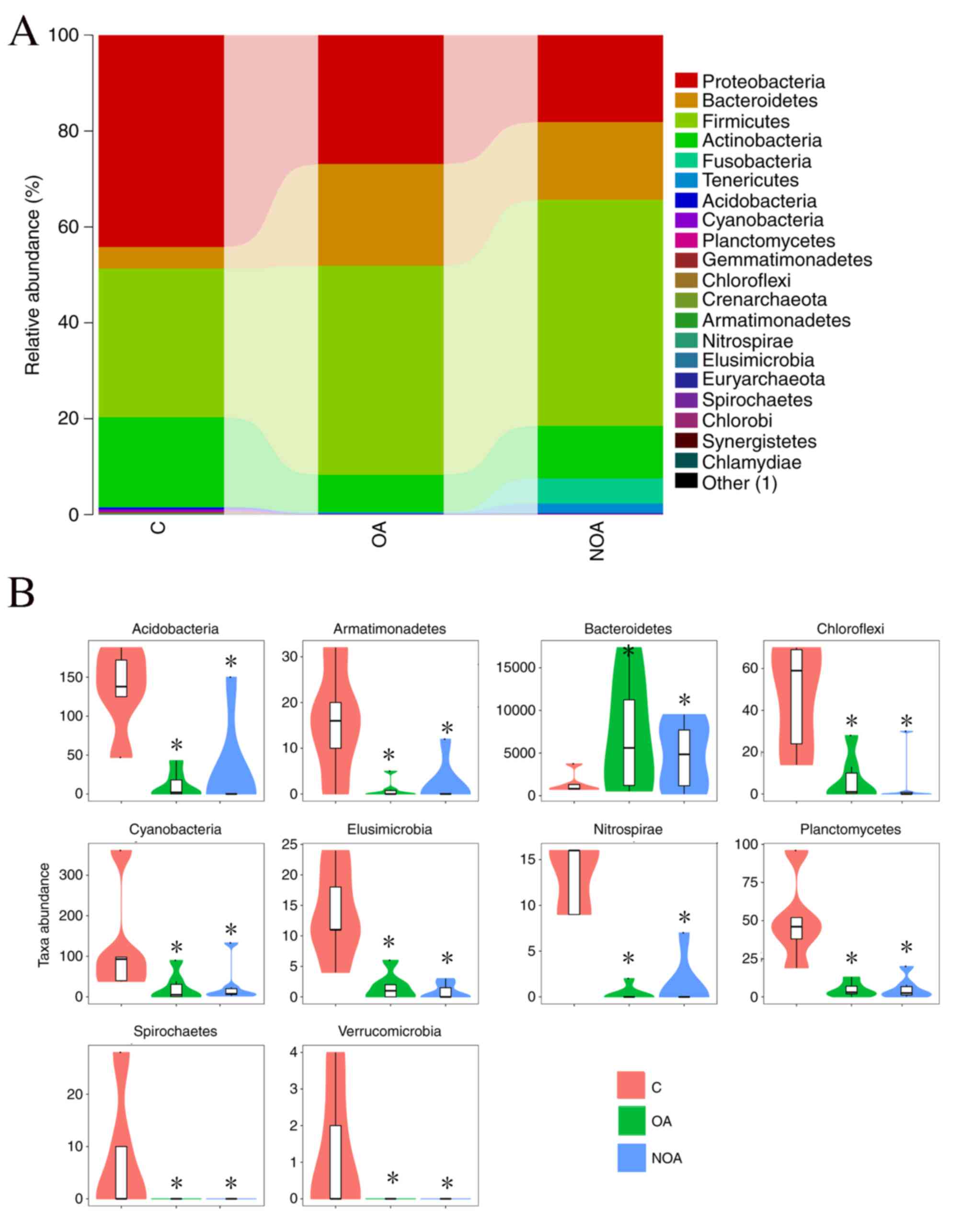
At the phylum level, data from the 20 microorganisms
that were most abundant in each group was analyzed. Firmicutes,
Proteobacteria, Bacteroidetes and Actinobacteria were the
predominant phyla, accounting for 98.14% in the C group, 98.26% in
the OA group and 90.96% in the NOA group (Fig. 1A).
Compared with the C group, azoospermia reduced the
OTU number of Cyanobacteria, Acidobacteria, Gemmatimonadetes,
Planctomycetes, Chloroflexi, Crenarchaeota, Armatimonadetes,
Elusimicrobia, Nitrospirae, Euryarchaeota, Spirochaetes, Chlorobi,
Synergistetes, Chlamydiae and Verrucomicrobia, whereas the number
of Bacteroidetes was increased (P<0.05; Fig. 1B).
Seminal bacterial composition at the
genus level
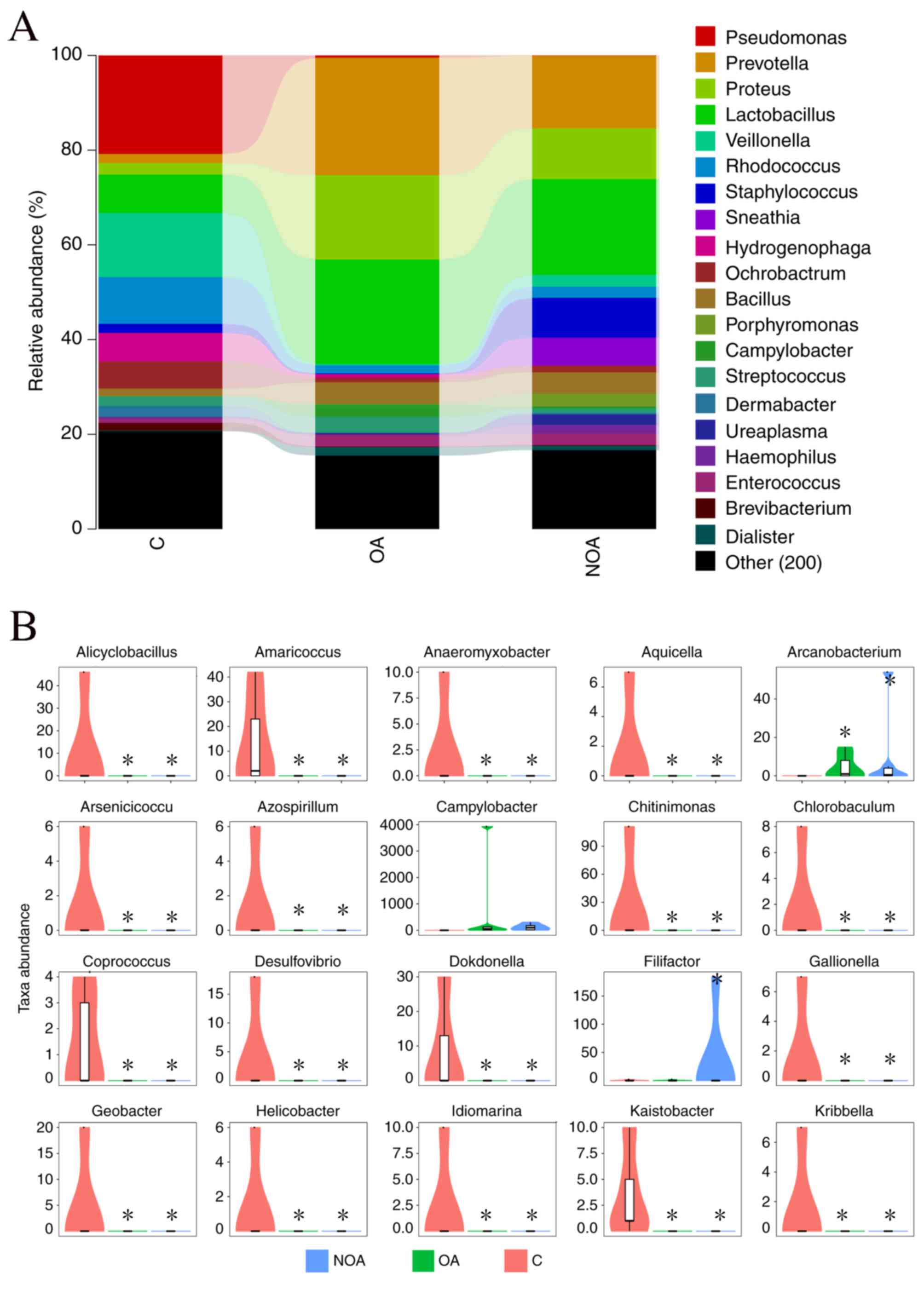
Lactobacillus, Prevotella,
Proteus, Pseudomonas, Veillonella,
Corynebacterium, Rhodococcus, Staphylococcus
and Bacillus were the dominant genera detected in the C
(55.02%), OA (62.54%) and NOA (59.74%) groups (Fig. 2A). The distribution map indicated
that Alicyclobacillus, Amaricoccus,
Anaeromyxobacter, Aquicella, Arsenicicoccus,
Azospirillum, Chitinimonas, Chlorobaculum,
Coprococcus, Desulfovibrio, Dokdonella,
Gallionella, Geobacter, Helicobacter,
Idiomarina, Kaistia and Kribbella were
significantly reduced in the C group compared with the OA and NOA
groups (P<0.05; Fig. 2B).
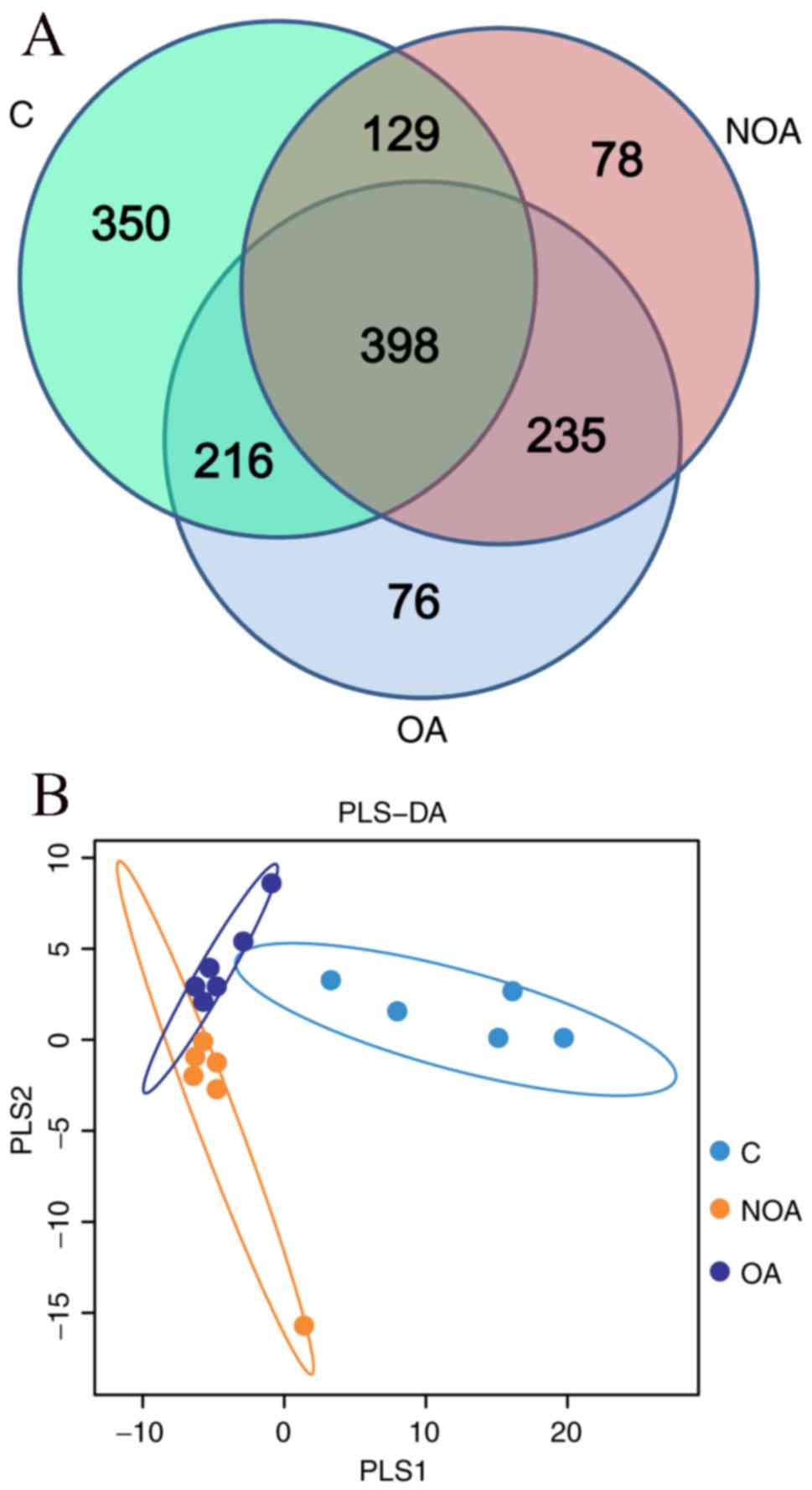
The diversity of the microbial
community in the C, OA and NOA groups
There were 1,093, 925 and 840 OTUs detected in the
C, OA and NOA groups, respectively (Fig.
3A). A total of 398 common OTUs were identified in the C, OA
and NOA groups, of which 27 belonged to the Lactobacillus phylum
(Fig. 3A). In addition, the partial
least squares discriminant analysis (PLS-DA) indicated that
patients in the OA and NOA groups scattered on the left, whereas
the C group gathered together on the right of the coordinate axis
(Fig. 3B). No overlapping of the C
group with the OA or NOA groups was observed; however, some samples
in OA and NOA groups overlapped (Fig.
3B), indicting the microbial diversity between the OA or NOA
groups had a higher degree of similarity than the C group.
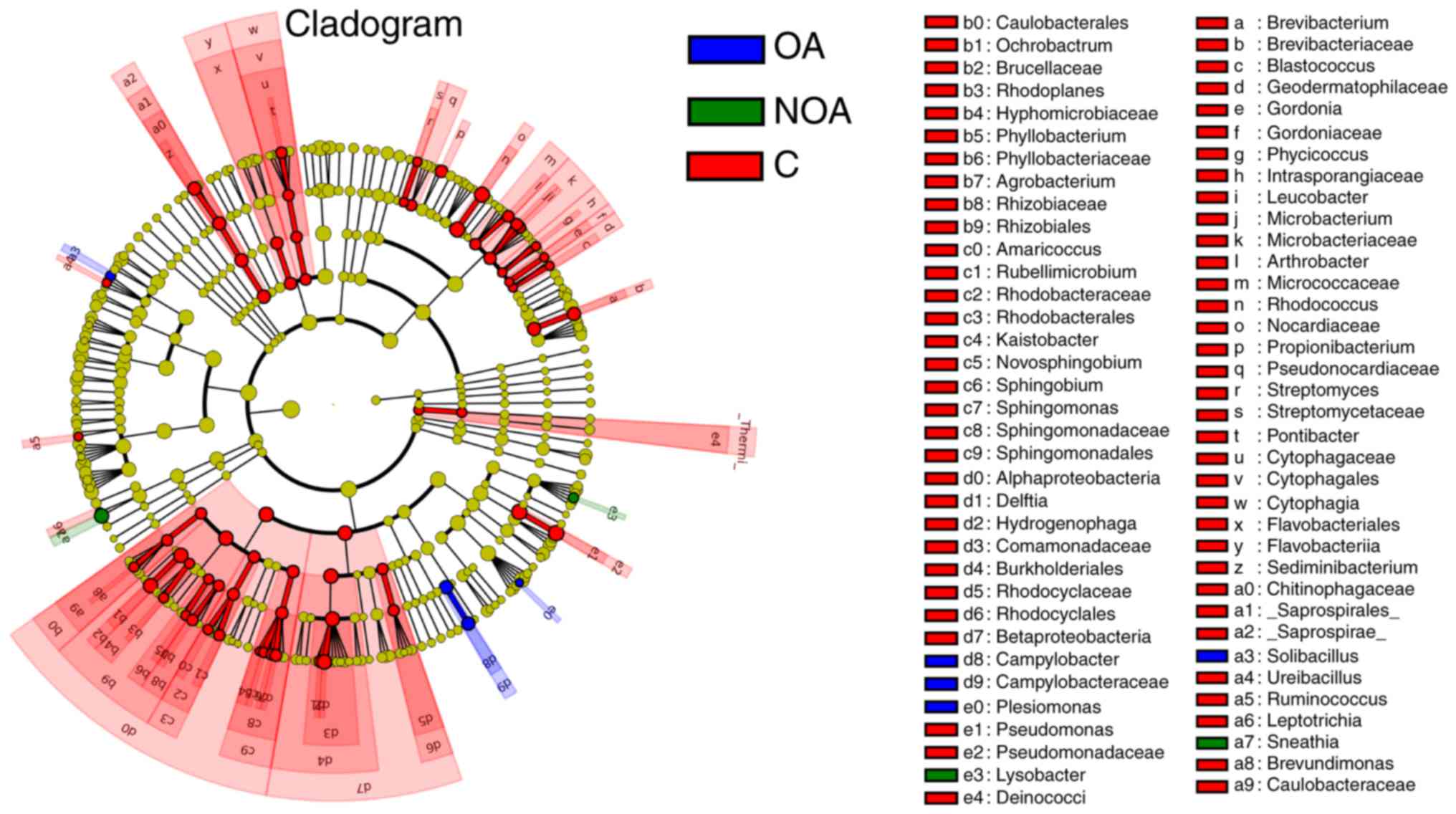
Composition of the bacterial
communities in C, OA and NOA groups
Significantly more Sneathia and Lysobacter
sp. were present in the NOA group compared with the C and OA
groups (P<0.05; Fig. 4). The
number of Solibacillus, Campyiobacter,
Campyiobacteraceae and Plesiomonas OTUs in the OA
group was significantly higher compared with the NOA and C groups
(P<0.05; Fig. 4). The number of
bacteria in red-labeled genera was significantly higher in the C
group compared with the OA and NOA groups (P<0.05; Fig. 4).
Prediction of bacterial functions in
C, OA and NOA groups
Correlation analyses were performed for various
Kyoto Encyclopedia of Genes and Genomes pathways using the PICRUSt
software. The association between seminal bacteria and health was
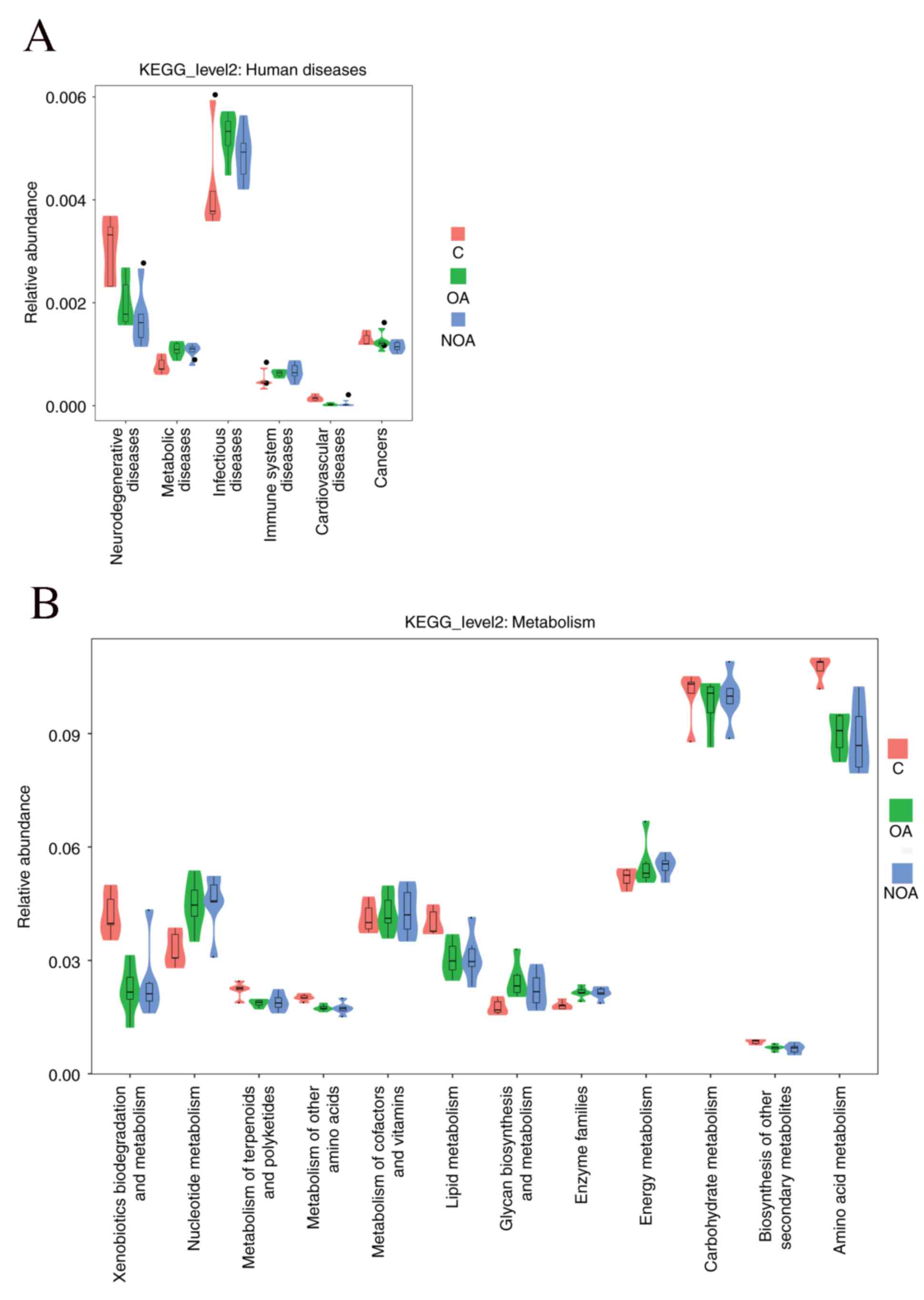
investigated (Fig. 5A). The results
indicated that patients in the OA and NOA groups were at an
increased risk of having metabolic, infectious and immune diseases
(Fig. 5A). Furthermore, the results
indicate that azoospermia enhances the protein expression of genes
associated with nucleotide metabolism, metabolism of cofactors and
vitamins, glycan biosynthesis and metabolism, enzyme families and
energy metabolism; however, the expression of genes associated with
xenobiotic biodegradation and metabolism, metabolism of terpenoids
and polyketides, lipid metabolism and amino acid metabolism was
downregulated (Fig. 5B).
Discussion
In the present study, semen samples were obtained
from patients with azoospermia and healthy control subjects,
following which the 16S rRNA gene fragments were sequenced using
high-throughput sequencing method. The results revealed the
composition of polymicrobial communities in semen samples and the
most prevalent phyla were Firmicutes, Proteobacteria, Bacteroidetes
and Actinobacteria, comprising 98.14, 98.26 and 90.96% of total
phyla in the C, OA and NOA groups, respectively. Patients in the OA
and NOA groups had a greater number of Bacteroidetes and Firmicutes
phyla, whereas the number of Proteobacteria and Actinobacteria was
decreased compared with the C group. Firmicutes is a phylum of
bacteria, the majority of which produce endospores, are resistant
to desiccation, are able to survive extreme conditions and are
typically found in various environments; this group includes some
notable pathogens (29). In
addition, the phylum Bacteroidetes comprises three large classes of
Gram-negative, non-spore-forming, anaerobic or aerobic and
rod-shaped bacteria; furthermore, some members of the genus
Bacteroides are opportunistic pathogens (29). The increased number of Bacteroidetes
and Firmicutes species in patients with azoospermia may therefore
increase the risk of gynecological inflammation in their sexual
partners.
At the genus level, nine dominant bacteria of the
Lactobacillus, Prevotella, Proteus,
Pseudomonas, Veillonella, Corynebacterium,
Rhodococcus, Staphylococcus and Bacillus genera
accounted for 55.02, 62.54 and 59.74% of total OTUs in the C, OA
and NOA groups, respectively. The OTU number of
Lactobacillus was 6.79% in the C group, whereas it accounted
for 17.98 and 17.24% in the OA and NOA groups, respectively. The
vagina and its microflora form a balanced ecosystem; vaginal
dysbacteriosis therefore results in obstetric and gynecological
diseases, as well as complications characterized by a reduced
number or total absence of Lactobacilli (30,31). It
may therefore be of use to further study the role of
Lactobacillus in the semen of patients with azoospermia.
To further compare the microbial diversity across
groups, PLS-DA and Venn methods were used. The Venn results
indicated that there were 398 OTUs common to the C, OA and NOA
groups, of which 27 were Lactobacillus. Furthermore, PLS-DA
indicated that the bacterial community was more similar when
comparing the OA and NOA groups than when comparing either
azoospermia group with control subjects. This suggests that sperm
may be an important factor in determining microbial
composition.
Patients with azoospermia exhibited a reduction in
many seminal bacteria compared with healthy controls; however, the
OTU number of Solibacillus, Campylobacter,
Campyiobacteraceae and Plesiomonas in the OA group,
as well as Sneathia and Lysobacter in the NOA group,
were increased. The majority of Campylobacter species cause
diseases that affect humans and other animals, so this increase
poses a potential health risk to patients with azoospermia
(32).
PICRUSt software as used to assess bacterial
functions and the results indicated that patients with azoospermia
had an increased expression of genes associated with nucleotide
metabolism, metabolism of cofactors and vitamins, glycan
biosynthesis and metabolism, enzyme families and energy metabolism,
as well as a reduced expression of genes associated with xenobiotic
biodegradation and metabolism, metabolism of terpenoids and
polyketides, lipid metabolism and amino acid metabolism. These
changes in expression increase the risk of metabolic, infectious
and immune diseases in patients with azoospermia compared with the
C group.
In the present study, the seminal bacterial
composition in patients with OA and NOA was explored using
high-throughput sequencing and the results indicate that the semen
of patients with azoospermia contains less beneficial microbes
compared with healthy controls. The increased number of
Bacteroidetes and Firmicutes species in patients with azoospermia
puts them at a greater risk of metabolic, infectious and immune
diseases. To the best of our knowledge, this is the first study
comparing seminal microbiomes in patients with OA or NOA and
healthy men using Illumina sequencing. The small sample size used
in the present study may be insufficient to fully reflect the
actual microbiota composition of semen from patients with OA or NOA
and, as such, future studies are required.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81501317, 81503364,
31560264), Excellent Youth Foundation of Jiang Xi Scientific
Committee (grant nos. 20171BCB23028).
References
|
1
|
Luo T, Zou QX, He YQ, Wang HF, Wang T, Liu
M, Chen Y and Wang B: Matrine compromises mouse sperm functions by
a [Ca(2+)]i-related mechanism. Reprod Toxicol. 60:69–75. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
He Y, Zou Q, Li B, Chen H, Du X, Weng S,
Luo T and Zeng X: Ketamine inhibits human sperm function by
Ca(2+)-related mechanism. Biochem Biophys Res Commun. 478:501–506.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He Y, Zou Q, Chen H, Weng S, Luo T and
Zeng X: Lead inhibits human sperm functions by reducing the levels
of intracellular calcium, cAMP and tyrosine phosphorylation. Tohoku
J Exp Med. 238:295–303. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hooper LV, Littman DR and Macpherson AJ:
Interactions between the microbiota and the immune system. Science.
336:1268–1273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nelson MH, Diven MA, Huff LW and Paulos
CM: Harnessing the microbiome to enhance cancer immunotherapy. J
Immunol Res. 2015:3687362015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakatsuji T, Chen TH, Narala S, Chun KA,
Two AM, Yun T, Shafiq F, Kotol PF, Bouslimani A, Melnik AV, et al:
Antimicrobials from human skin commensal bacteria protect against
Staphylococcus aureus and are deficient in atopic dermatitis. Sci
Transl Med. 9:pii: eaah46802017. View Article : Google Scholar
|
|
7
|
De Croo I, Van der Elst J, Everaert K, De
Sutter P and Dhont M: Fertilization, pregnancy and embryo
implantation rates after ICSI in cases of obstructive and
non-obstructive azoospermia. Hum Reprod. 15:1383–1388. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Franasiak JM and Scott RT Jr: Reproductive
tract microbiome in assisted reproductive technologies. Fertil
Steril. 104:1364–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Javurek AB, Spollen WG, Ali AM, Johnson
SA, Lubahn DB, Bivens NJ, Bromert KH, Ellersieck MR, Givan SA and
Rosenfeld CS: Discovery of a novel seminal fluid microbiome and
influence of estrogen receptor alpha genetic status. Sci Rep.
6:230272016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tuddenham S and Ghanem KG: A microbiome
variable in the HIV-prevention equation. Science. 356:907–908.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Francesco MA, Negrini R, Ravizzola G,
Galli P and Manca N: Bacterial species present in the lower male
genital tract: A five-year retrospective study. Eur J Contracept
Reprod Health Care. 16:47–53. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Younes JA, Lievens E, Hummelen R, van der
Westen R, Reid G and Petrova MI: Women and their microbes: The
unexpected friendship. Trends Microbiol. 26:16–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weng SL, Chiu CM, Lin FM, Huang WC, Liang
C, Yang T, Yang TL, Liu CY, Wu WY, Chang YA, et al: Bacterial
communities in semen from men of infertile couples: Metagenomic
sequencing reveals relationships of seminal microbiota to semen
quality. PLoS One. 9:e1101522014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu X, Wu X, Qiu L, Wang D, Gan M, Chen X,
Wei H and Xu F: Analysis of the intestinal microbial community
structure of healthy and long-living elderly residents in Gaotian
village of liuyang city. Appl Microbiol Biot. 99:9085–9095. 2015.
View Article : Google Scholar
|
|
15
|
Sivan A, Corrales L, Hubert N, Williams
JB, Aquino-Michaels K, Earley ZM, Benyamin FW, Lei YM, Jabri B,
Alegre ML, et al: Commensal Bifidobacterium promotes antitumor
immunity and facilitates anti-PD-L1 efficacy. Science.
350:1084–1089. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhernakova A, Kurilshikov A, Bonder MJ,
Tigchelaar EF, Schirmer M, Vatanen T, Mujagic Z, Vila AV, Falony G,
Vieira-Silva S, et al: Population-based metagenomics analysis
reveals markers for gut microbiome composition and diversity.
Science. 352:565–569. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Derrien M and van Hylckama Vlieg JE: Fate,
activity and impact of ingested bacteria within the human gut
microbiota. Trends Microbiol. 1–366. 2015.
|
|
18
|
Xin F, Xin W, Yang S, Meng F, Wang X, Wei
H and Chen T: Evaluation of the microbial diversity in amyotrophic
lateral sclerosis using high-throughput sequencing. Front
Microbiol. 7:14792016.PubMed/NCBI
|
|
19
|
Cui M, Xiao H, Li Y, Zhou L, Zhao S, Luo
D, Zheng Q, Dong J, Zhao Y, Zhang X, et al: Faecal microbiota
transplantation protects against radiation-induced toxicity. EMBO
Mol Med. 4:448–461. 2017. View Article : Google Scholar
|
|
20
|
Chen T, Wu Q, Zhou H, Deng K, Wang X, Meng
F, Yang S, Wang X, Shah NP and Wei H: Assessment of commercial
probiotic products in China for labelling accuracy and probiotic
characterisation of selected isolates. Int J Dairy Technol.
70:119–126. 2017. View Article : Google Scholar
|
|
21
|
Xu J, Lian F, Zhao L, Zhao Y, Chen X,
Zhang X, Guo Y, Zhang C, Zhou Q, Xue Z, et al: Structural
modulation of gut microbiota during alleviation of type 2 diabetes
with a Chinese herbal formula. Isme J. 9:552–562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bolger A, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Edgar R: UPARSE: Highly accurate OTU
sequences from microbial amplicon reads. Nat Methods. 10:996–998.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Steege HT: Measuring biological diversity.
Environ Ecol Stat. 1:95–103. 2004.
|
|
25
|
Cole JR, Wang Q, Fish JA, Chai B,
McGarrell DM, Sun Y, Brown CT, Porras-Alfaro A, Kuske CR and Tiedje
JM: Ribosomal database project: Data and tools for high throughput
rRNA analysis. Nucleic Acids Res. 42(Database Issue): D633–D642.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Caporaso JG, Kuczynski J, Stombaugh J,
Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich
JK, Gordon JI, et al: QIIME allows analysis of high-throughput
community sequencing data. Nat Methods. 7:3352010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Javurek AB, Spollen WG, Johnson SA, Bivens
NJ, Bromert KH, Givan SA and Rosenfeld CS: Consumption of a
high-fat diet alters the seminal fluid and gut microbiomes in male
mice. Reprod Fert Devel. 29:1602–1612. 2017. View Article : Google Scholar
|
|
28
|
Langille MG, Zaneveld J, Caporaso JG,
McDonald D, Knights D, Reyes JA, Clemente JC, Burkepile DE, Vega
Thurber RL, Knight R, et al: Predictive functional profiling of
microbial communities using 16S rRNA marker gene sequences. Nat
Biotechnol. 31:814–821. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wolf M, Müller T, Dandekar T and Pollack
JD: Phylogeny of Firmicutes with special reference to Mycoplasma
(Mollicutes) as inferred from phosphoglycerate kinase amino acid
sequence data. Int J Syst Evol Microbiol. 54:871–875. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Donders GG, Bosmans E, Dekeersmaeckerb A,
Vereecken A, Van Bulck B and Spitz B: Pathogenesis of abnormal
vaginal bacterial flora. Am J Obstet Gyneco. 182:872–878. 2000.
View Article : Google Scholar
|
|
31
|
Kovachev SM: Obstetric and gynecological
diseases and complications resulting from vaginal. dysbacteriosis.
68:1–184. 2014.
|
|
32
|
Bae HS, Im WT and Lee ST: Lysobacter
concretionis sp. nov., isolated from anaerobic granules in an
upflow anaerobic sludge blanket reactor. Int J Syst Evol Microbiol.
55:1155–1161. 2005. View Article : Google Scholar : PubMed/NCBI
|