Introduction
Premature ovarian failure (POF), which is
characterized by high gonadotropin and low estrogen levels, has a
serious impact on women's reproductive and psychological health.
The most common presentation is secondary amenorrhea and the main
consequence is infertility. Infertility may be the consequence of
follicle depletion or a poor response to gonadotropin, which
reduces the effectiveness of ovulation induction programs (1). Women with POF not only have premature
aging symptoms but are also subject to a significantly increased
risk of osteoporosis and cardiovascular disease (2).
Autoimmune abnormality is one of the main causes of
POF. Previous studies by Chernyshov et al (3) and Ban (4) indicated that immunoregulation disorder
involving an imbalance between CD4+ T and
CD8+ T is a leading cause of POF. The researchers also
revealed increased autoantibodies of peripheral blood
CD5+ CD19+ B lymphocytes in early cases and
high levels of zona pellucida (ZP) antibodies in patients with POF
(5). Another study demonstrated that
the levels of regulatory T cells (Tregs) in patients with POF were
significantly lower than those in normal controls (6). Thus, the abnormal regulation of Tregs
lead to an autoimmune response that harmed the ovaries, which may
be responsible for the pathogenesis of POF.
Women with POF are usually treated with hormone
replacement therapy to compensate for the decreased ovarian
production of sex steroids. However, long-term use of hormone
replacement therapy may increase a female's risk of breast and
ovarian cancer, venous thromboembolism and stroke (7). Glucocorticoids are used clinically for
immunosuppression to treat autoimmune POF; however, there is no
known prospective randomized placebo controlled studies that
demonstrating the safety and efficacy of immunosuppressive therapy
for POF (8). Therefore, the
exploitation of effective drugs and treatments is particularly
important for the prevention and therapy of POF.
Bu Shen Huo Xue formula (BSHXF) is a Chinese herbal
formulation. Clinical results revealed that BSHXF restored the
functions of the ovary and markedly improved the clinical symptoms
of POF (9). Early results indicated
that BSHXF may protect the ovaries from autoimmune destruction
(10). However, the mechanisms
require further investigation.
In the present study, an animal model of autoimmune
POF was established using internationally recognized ZP3 fragments
(11). The protective immunity
effects of BSHXF were investigated on the expression of Treg cells
and CD4+ T lymphocytes in mice, anti-ZP antibodies and
associated inflammatory cytokines.
Materials and methods
Reagents and instruments
The amino acid sequence of the murine ZP3 330–342
peptide used in the present study (NSSSSQFQIHGPR) was synthesized
by GL Biochem, Ltd. (Shanghai, China), and the amino acid
composition was verified by amino acid analysis. Mycobacterium
tuberculosis H37RA was purchased from Difco (BD Biosciences,
Franklin Lakes, NJ, USA). Incomplete Freund's adjuvant (IFA) was
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Qualitative analysis was performed using a HPLC-MS/MS system
consisting of Waters 2695 HPLC instrument and Quattro Premier XE
MircoMass triple quadrupole tandem mass spectrometer (Waters Co.,
Milford, MA, USA). Enzyme-linked immunosorbent assay (ELISA) kits
for mouse interleukin-10 (IL-10) (#BMS614-2, IL-10 Mouse ELISA kit)
and interferon-γ (IFN-γ) (#BMS6027, IFN alpha Mouse ELISA kit) were
purchased from eBioscience (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), EnVisionTM horseradish peroxidase rabbit/mouse
reagent (#K5007) was purchased from Agilent Technologies, Inc.
(Santa Clara, CA, USA). 3,3′-Diaminobenzidine (DAB) was purchased
from Medchemexpress Co., Ltd. (Monmouth Junction, NJ, USA). Mouse
anti-CD3 allophycocyanin (APC) (#17-0031-82), anti-CD4 fluorescein
isothiocyanate (FITC) (#11-0042-86), anti-CD8 phycoerythrin (PE)
(#85-12-0081-82), anti-CD25 FITC (#11-0250-42), anti-mouse forkhead
box P3 (FoxP3) PE (#12-4771-82), mouse IgG1 isotype control APC
(#MA5-18093), FITC (#GM4992) and PE (#GM4993) antibodies were
purchased from eBioscience (Thermo Fisher Scientific, Inc.). Red
blood cell lysing buffer (#c3702) and bovine serum albumin (BSA)
were purchased from Beyotime Biotechnology Co. (Shanghai, China).
RPMI 1640, new born calf serum (NCS), fetal calf serum (FBS) and
N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid (HEPES) were
purchased from Invitrogen (Thermo Fisher Scientific, Inc.). Mouse
IL-2 (#P04351) was purchased from R&D Systems, Inc.
(Minneapolis, MN, USA). Carboxyfluorescein succinimidyl ester
(CFSE) Cell Proliferation kit (#C34554) was purchased from
Invitrogen (Thermo Fisher Scientific, Inc.). Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) kits
and reverse transcription kits were purchased from MBI Fermentas
(Thermo Fisher Scientific, Inc.). PCR primers were synthesized by
Jinsirui Biotechnology Co., Ltd. (Nanjing, China).
Preparation of BSHXF
The herbal medicine was produced by Jiangsu Yadong
Biological Pharmacy Co., Ltd. (Nanjing, China). BSHXF was composed
of Rehmanniae Radix Preparata, Radices Paeoniae Alba, Angelica
sinensis, Ligusticum wallichii, Semen Cuscutae, Herba Epimedii,
Rhizoma Anemarrhenae, Golden Cypress and Radix Bupleuri. The
proportions of these herbs were 10:10:10:10:10:10:10:10:6,
respectively. The herbs (1 kg) were extracted twice with boiling
water, and the combined aqueous extracts were concentrated to yield
a crude extract (233 g) under reduced pressure, and stored at
4°C.
Identification of active ingredients
in BSHXF using high-performance liquid chromatography (HPLC)
Methanol (Tianjin Concord Science Co., Ltd.,
Tianjin, China) and Milli-Q Academic ultrapure water system (Waters
Corp., Milford, MA, USA). All other reagents were of analytical
grade (Tianjin Concord Science Co., Ltd.). HPLC with diode-array
detection was used to detect peoniflorin (#23180-57-6), ferulic
acid (#1135-24-6) and icariin (#489-32-7; Must Bio-Technology Co.,
Ltd., Chengdu, China). The samples were separated on an Alltima C18
column (5 µm, 250×4.6 mm; Waters Corp.) with a mobile phase that
consisted of methanol-0.1% phosphoric acid (gradient elution) at
25°C. The injection volume was 20 µl with the flow rate of 1
ml/min. The average recoveries of peoniflorin, ferulic acid and
icariin were 96.7, 97.5 and 99.6%, and the relative standard
deviations were 1.37, 3.13 and 0.96%, respectively. The linear
ranges of peoniflorin, ferulic acid and icariin were 0.159–3.970 µg
(r=0.9997), 0.048–1.200 µg (r=0.9999), 0.207–5.175 µg (r=0.9999).
Therefore, the samples were analyzed by HPLC. The contents of the
chief components in BSHXF, peoniflorin, ferulic acid and icariin
(derived from Radices Paeoniae Alba, Angelica sinensis and
Herba Epimedii) were 0.842, 0.032 and 1.608 mg/g, respectively.
Experimental animals
A total of 70 female C57BL/6 mice, 20 male A/J mice
(aged 7–9 weeks; weighing, 16.0–18.0 g) and 10 female Wistar rats
(aged 7–9 weeks; weighing, 190.0–210.0 g) were purchased from
Academy of Military Medical Sciences (Beijing, China)
[certification no. SCXK (JUN) 2007-004]. The mice were fed in the
laboratory animal room of the Jiangsu Research Institute of
Traditional Chinese Medicine [Nanjing, China; certification no.
SYXK (SU) 2011–0017]. The animals were housed in a controlled
environment (temperature, 23±2°C; relative humidity, 42±5% with a
12-h light/dark cycle) and had ad libitum access to drinking
water and food. Mice were allowed to be acclimatized to the
laboratory environment for at least 1 week prior to commencement of
testing. The first generation (F1) of hybrid mice was obtained by
mating C57BL/6 and A/J mice. B6AF1 mice were studied at 8 weeks of
age. Vaginal smears were normal in the cell smear screening. The
present study was ethically approved by the Animal Care Ethics
Committee of Southeast University (Nanjing, China).
Model establishment and experimental
treatment
A total of 75 female B6AF1 mice were randomly
divided into three groups: Control (n=25), model (n=25) and
BSHXF-treated (n=25) groups. ZP3 peptides were dissolved in
double-distilled water to a concentration of 1 mM and sterilized by
ultrafiltration. They were then emulsified in an equal volume of
IFA containing M. tuberculosis. POF model mice were
established by injection subcutaneously (s.c.) in the hind footpad
and tail root with 0.1 ml emulsion containing 165 µg M.
tuberculosis in 500 µl IFA and 72 µg ZP3 peptides in 500 µl
ddH2O. The same volume of PBS was injected into a
parallel group of mice to represent the Control group. Mice were
sacrificed 14 days after immunization in the pre-experiment, which
demonstrated POF mouse models were successful. The mice in the
BSHXF-treated group were then administered an oral suspension of
31.53 g/kg BSHXF once a day for the following 28 days. Mice in the
control and model groups were treated with equal amounts of
distilled water.
BSHXF-derived serum preparation
A total of 10 Wistar rats were randomly divided into
two groups: Blank and BSHXF groups (5 mice per group). At the same
time, the mice were orally administrated treatments continuously 2
weeks. BSHXF group rats were administered 18.63 g/kg BSHXF whereas
the blank group rats were administrated equal volumes of double
distilled water. A total of 1 h after the last BSHXF intragastric
administration, serum was acquired from the heart. It was
inactivated at 56°C for 30 min, filtered through at 0.2-mm sized
filter, and stored at −80°C until use.
Estrous cycle staging
Stages of the estrous cycle were determined by
examining vaginal cytology. Vaginal washes in saline were
transferred to a glass slide, which was air-dried, fixed in
methanol for 30 min at room temperature and stained with
Papanicolaou (Hologic, Inc., San Diego, CA, USA) for 10 min at room
temperature. The estrous cycle was staged according to the
proportions of leukocytes, nucleated epithelial cells and cornified
squamous epithelial cells (12). The
sections were observed using light microscopy (Nikon Eclipse 80i;
Nikon Corporation, Tokyo, Japan). Images were captured using a
digital camera (COOLPIX 950; Nikon Corporation).
Sample collection
The vaginal exfoliate cell smear of mice was
observed from day 10 and continued for 4 days. The mice sacrificed
on day 14 were examined using hematoxylin and eosin (H&E)
staining in order to determine whether modeling was successful.
After 28 days of administration, all remaining mice were sacrificed
for analysis. Blood was collected from the mouse orbit. The thymus,
spleen, ovary and uterus were extirpated and weighed for the
calculation of organ indices. Additionally, the spleens of mice in
each group were removed and prepared for flow cytometric analysis,
and the left-side ovary and uterus were prepared for H&E
staining for histopathological investigation. The right-side ovary
and uterus were immediately frozen to detect ZP using a direct
immunofluorescence assay.
Ovarian and uterus
histomorphology
Ovaries were fixed in Bouin's fluid (MXB
Biotechnologies, Fuzhou, China; 1:1) fixative for 24 h at room
temperature and embedded in paraffin. Serial sections (5 µm) were
stained with H&E for 24 h at room temperature and examined as
coded specimens by an independent observer who was blind to the
experimental details. The sections were observed using light
microscopy (Nikon Eclipse 80i; Nikon Corporation). Images were
captured using a digital camera (COOLPIX 950, Nikon
Corporation).
Ovarian immunohistochemistry
The ovaries were isolated from the mice, fixed in
10% paraformaldehyde for 48 h at room temperature and
paraffin-embedded. Samples were embedded in paraffin wax, treated
with xylene and infused into PBS buffer for 4–8 h in a water bath
at 60°C. Subsequently, samples were rehydrated in a graded alcohol
series. Antigen retrieval was accomplished by heating sections to
180°C for 10 min in Citrate buffer (pH 6.2) (Dako; Agilent
Technologies, Inc.) and allowed to cool to room temperature.
Consecutive 4-µm sections were cut and processed for
immunohistochemistry with CD4 (#RMA-0620), estrogen receptor (ER)
(#kit-0012) and progesterone receptor (PR) (#kit-0013) at 1:100
concentration (MXB Biotechnologies, Fuzhou, China). Each section
was treated with 50 µl of primary antibody (CD4/ER/PR) and
incubated overnight at 4°C. Subsequently, each section was treated
with 50 µl EnVisionTM horseradish peroxidase rabbit/mouse reagent
as a secondary antibody overnight at 4°C. The antibody-binding
sites were visualized using DAB for 10 min at room temperature and
the cell nuclei were counterstained with hematoxylin for 10 sec at
room temperature. The estrogen/progesterone-receptor-positive
clinical breast cancer specimen was used as a positive control and
estrogen/progesterone-receptor-negative breast tissue was
considered a negative control. The tonsil tissue section served as
CD4 positive control and PBS was used in place of the first
antibody to serve as a negative control. The positive cells were
stained brown. The slides were examined under a light microscope,
and representative images were captured from a minimum of 4 or 5
slides from each group. The sections were observed using light
microscopy (Nikon Eclipse 80i; Nikon Corporation). Images were
captured using a digital camera (COOLPIX 950; Nikon Corporation).
All slides were examined in a blinded manner.
Detection of antibodies to ovarian ZP
by immunofluorescence
Sections (5 µm) of snap-frozen mouse ovaries, which
had been fixed in cold acetone for 15 min, incubated by using 3%
bovine serum albumin in PBS for 30 min at room temperature.
Following washing in PBS, slices were dried and treated with 1 ml
Alexa Fluor® 488 donkey anti-mouse IgG (H+L) (1:200;
#R37114; Invitrogen; Thermo Fisher Scientific, Inc.) antibody at
37°C for 45 min. After washing three times with PBS, slides were
viewed and images were captured with an inverted fluorescence
microscope (Carl Zeiss AG, Oberkochen, Germany).
Isolation of CD4+
CD25+ Tregs
CD4+ T cells and CD4+
CD25+ Tregs were isolated from murine spleen using
CD4+ T cell isolation and CD4+
CD25+ Treg isolation kits (Thermo Fisher Scientific,
Inc., Waltham, MA, USA), respectively, following the manufacturer's
protocols.
Flow cytometry (FCM) analysis
To assess the changes in splenic Treg cells in each
group of mice, the levels of CD4+ CD25+
FoxP3+ T cells were measured using FCM. Spleens of mice
were harvested, passed through a 70-µm cell strainer, centrifuged
at 300 × g for 5 min at 4°C and resuspended in a red blood cell
lysing buffer at 37°C for 2 min to lyse the red blood cells,
whereby erythrocytes from the spleen were lysed in ammonium
chloride buffer and the remaining living cells washed and
resuspended in PBS. A single-cell suspension was prepared. Two sets
of experiments were performed. The first set used a total of
1×106 cells stained with the following conjugated
antibodies: CD3-APC, CD4-FITC and CD8-PE. The second set used a
total of 1×106 cells stained with CD4-FITC, CD25-APC and
FoxP3-PE. FoxP3 and cytokine staining were performed according to
the manufacturer's intracellular staining protocol, and appropriate
isotype antibodies were used as controls.
In vitro CFSE proliferation assay
The culture media consisted of RPMI 1640
supplemented with 2% NCS, 1% HEPES, 1% anti-CD3/28 mAbs and 1% FBS.
Culture media was freshly composed before usage and stored in a
refrigerator at 4°C. Isolated CD4+ CD25+ Treg
cells were seeded into 96 well plate, each well containing
1×105 Tregs. The cells were labeled with CFSE
(Invitrogen; Thermo Fisher Scientific, Inc.), stimulated with IL-2
(40 U/well) and BSHXF-derived serum/PBS (10 µl/well). T cells were
cultured in media at 37°C under 5% CO2 saturated
humidity for 2 days. After 24 h, the media were replaced with fresh
media. A total of 48 h later, the cells were harvested and analyzed
using flow cytometry. Detection of
CD4+CD25+Treg proliferation using a CFSE kit
(#C34554; Invitrogen, Thermo Fisher Scientific, Inc.) following the
manufacturer's protocols.
Determination of cytokine levels in
mouse serum
The blood of each mouse was centrifuged at 1,000 × g
for 10 min at 4°C to obtain serum for cytokine ELISA. ELISA kits
were used to detect changes in the levels of IFN-γ and IL-10
cytokines in the mice. The aforementioned ELISA kits were used for
quantitative determination of the aforementioned cytokines in the
serum samples of mice. Cytokine concentrations were determined
using the relevant standard curves.
Expression levels of FoxP3 mRNA
Total RNA was isolated from ~30 mg of spleen tissue
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. The RNA concentration was
quantified with a Spectrophotometer (Eppendorf, Hamburg, Germany).
RNA was reversed transcribed into cDNA using reverse transcriptase
(Takara Shuzo Co., Tokyo, Japan) at 37°C for 1 h. A total of 500 ng
of RNA was reverse transcribed in a 20-µl system using a Revert Aid
First-Strand cDNA synthesis kit (Fermentas; Thermo Fisher
Scientific, Inc.) and the following thermocycling conditions were
used: 25°C for 10 min, 50°C for 15 min, 85°C for 5 min, followed by
a holding temperature of 4°C. Subsequently, 1 µl complementary DNA
was used as the template for qPCR using SYBR-Green PCR master mix
(with ROX; Invitrogen; Thermo Fisher Scientific, Inc.) and a 7300
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Each qPCR reaction contained 0.1 mM each primer, 5 µl
SYBR-Green PCR master mix including AmpliTaq Gold DNA polymerase
with buffer, dNTP mix, SYBR-Green I and ROX reference dyes and 10
mM MgCl2, and 1 µl template cDNA in a 10-µl total
reaction volume. The typical amplification program included
activation of the enzyme at 95°C for 10 min, followed by 45 cycles
of denaturation at 95°C for 15 sec, then annealing and extension at
60°C for 30 sec. The Cq (cycle threshold) value for each gene was
determined using the automated threshold analysis function of the
PCR instrument. The relative expression levels of FoxP3 against
GAPDH were calculated using the 2−ΔΔCq method (13). The primer sequences for PCR designed
using Primer Premier 5.0 software (Premier Biosoft International,
Palo Alto, CA, USA) on the basis of GenBank sequences, were as
follows: FoxP3 forward, 5′-CTGGACAACCCAGCCATGAT-3′ and reverse,
5′-ACATTGATCCCAGGTGGCAG-3′; GAPDH forward,
5′-CTGAAAATCAATAGCACGAAC-3′ and reverse,
5′-ATGGAGCCACCGATCCACA-3′.
Statistical analysis
Each experiment was performed as least thrice, and
the data are presented as the mean ± standard error where
applicable. All statistical comparisons were analyzed by means of
one-way analysis of variance followed by Tukey's test using SPSS
21.0 software (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
General status of mice following model
establishment
Symptoms of the model group, including a flaccid
tail, staggering gait and irritability, appeared sequentially in
experimental mice from day 8. The ovaries of mice in the model
group were smaller than those of controls and the uterine volume
was also reduced.
Examination of the vaginal smears in the control
group (Fig. 1A) demonstrated that
the mice had a regular estrous cycle whereas the model group
presented irregular, prolonged and even arrested estrous cycles.
Regular sex cycles were identified and the ovulation of mice was
increased in the BSHXF-treated group compared with the model
group.
The results of ovarian H&E staining demonstrated
that compared with the control group, there were more primordial
follicles, and less typical growing and mature follicles in the
cortex of mouse ovaries in the model group, and a large amount of
apoptosis was observed in all these follicles (Fig. 1B).
Changes of indices following administration of
BSHXF
Body weight
The body weight of mice in each group decreased
markedly at >2 weeks following model establishment. However, as
the time was extended the body weight gradually increased. Compared
with the model group, the body weight of the BSHXF-treated group
increased more rapidly, but the most rapid increase was observed in
the control group. Notably, BSHXF may have a protective effect on
autoimmunity damage in mice with POF (Fig. 2A).
 | Figure 2.(A) Changes in mouse body weights
prior to and following BSHXF administration. Prior to modeling, the
weights of the mice in the control, model and BSHXF-treated groups
were 21.024±1.353, 20.700±1.287 and 20.588±1.113 (P>0.05).
Following modeling, the body weights were markedly reduced in the
model group compared with the control. At the end of the 6-week
experimental period, the body weights were 24.108±0.861,
23.612±0.564 and 22.984±0.903 for the control, model and
BSHXF-treated groups, respectively, and the body weight in the
model group was significantly decreased compared with that in the
control and BSHXF-treated groups (P<0.01). (B) Changes of organ
indices prior to and following BSHXF administration. The index
values in the control, model and BSHXF-treated groups were: Thymus
index: 2.619±0.371, 2.220±0.235 and 2.400±0.305 mg; spleen index:
3.543±0.227, 3.767±0.476 and 3.520±0.409 mg; uterus index:
3.561±0.149, 2.854±0.825 and 3.018±0.640 mg; ovary index:
0.498±0.164, 0.382±0.043 and 0.465±0.137 mg. *P<0.05 and
**P<0.01 vs. the control group; ##P<0.01 vs. the
model group. BSHXF, Bu Shen Huo Xue formula. |
Organ indices
The spleen index in the model group mice was
increased but the thymus, uterus and ovary indices were decreased
compared with those in the control group (P<0.05). Compared with
the model group, the ovarian and thymus indices in the
BSHXF-treated group mice increased significantly (P<0.01),
whereas the spleen index decreased and uterus index increased to a
non-statistically significant extent (P>0.05; Fig. 2B).
Pathological changes on the ovary and
uterus
The results of ovarian H&E staining revealed
that the number of follicles, particularly developing follicles,
was greater in the BSHXF-treated group compared with the model
group, and the lymphocytes were less involved in the follicle with
some lymphocyte infiltration in the interstitium. The results of
H&E staining of the uterus demonstrated that the uterine
endometrium in the model group was thinner than that in the control
group, secretory material was decreased in the epithelium, the
basic layer of the uterine gland was decreased, the gland cavity
decreased and reticulations of the inner membrane were also
reduced. However, the endometrium was thickened in the
BSHXF-treated group. It is noteworthy that BSHXF induced changes in
endometrial structure, and endometrial hyperplasia and thickening
(Fig. 3).
Immunohistochemical detection of
changes in ER and PR expression in the ovaries
Immunohistochemical studies indicated that the
expression of the ER and PR in the ovary in the model group was
decreased relative to that in the control group. After 4 weeks of
continuous administration, the expression of ER and PR in the ovary
increased. Estrogen and progesterone exert their functions by
combining with their corresponding receptors. It appears that the
expression of ER and PR in mice ovary may be promoted by BSHXF
(Fig. 4).
Immunofluorescence detection of
ovarian ZP
No strong immunofluorescence was exhibited in the
control group whereas bright green immunofluorescence was observed
in the model group. In the BSHXF-treated group, a few spots of
immunofluorescence were observed, and the expression of ZP antibody
was clearly decreased compared with that in the control group
(Fig. 5).
Regulation of CD4+ T
lymphocyte immune response
Immunohistochemical results for CD4+ are
depicted in Fig. 6A. The model mice
exhibited increased lymphocyte infiltration around the follicle
compared with the control mice. Lymphocyte infiltration was clearly
reduced following treatment with BSHXF.
 | Figure 6.(A) Immunohistochemical staining of
CD4+ in the ovaries (magnification, ×200). (B)
CFSE-based proliferative assay for CD4+ CD25+
Treg lymphocytes. (C) ELISA analysis of IL-10 and IFN-γ expression
in the spleen of the three groups. In the control, model and
BSHXF-treated groups, the levels of IL-10 were 55.647±9.185,
111.918±11.282 and 85.742±6.166, respectively, and the levels of
IFN-γ were 28.831±11.024, 76.844±13.736 and 55.183±11.595,
respectively. ***P<0.001 vs. the control group;
###P<0.001 vs. the model group. BSHXF, Bu Shen Huo
Xue formula; IL, interleukin; IFN-γ, interferon-γ. |
Proliferation of T cells
FBS or medicated serum was added to CFSE-labeled
CD4+ CD25+ Treg cells following separation,
and the cells were then cultured. After 2 days, the effect on cell
proliferation was detected by FCM. This revealed greater
proliferation in the medicated serum group compared with the
control group (Fig. 6B) indicating
that BSHXF in the serum was able to induce mouse CD4+
CD25+ Treg cell proliferation.
ELISA analysis of inflammation-related
cytokines in the serum
The levels of IL-10 and IFN-γ in the serum of model
mice was significantly increased compared with that in the control
group mice. However, the levels of these cytokines were
significantly decreased in BSHXF-treated mice compared with the
model mice (P<0.001; Fig.
6C).
FCM analysis of splenic
CD4+ CD25+ FoxP3+ T cells
The results of the FCM analysis are presented in
Fig. 7A. A significant reduction in
CD4+ CD25+ FoxP3+ T cells in the
model mice compared with the control group was observed
(P<0.01), and treatment with BSHXF significantly increased the
number of these cells compared with the model group
(P<0.05).
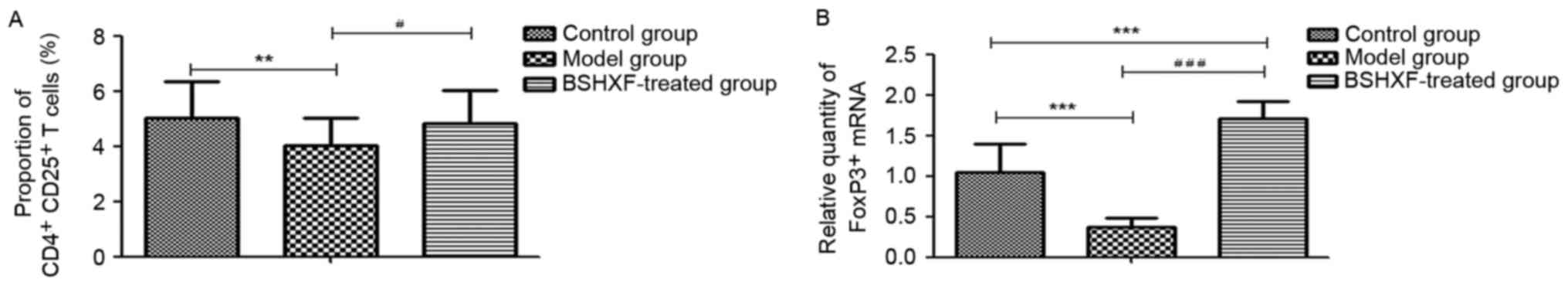 | Figure 7.(A) Effect of BSHXF on the expression
of CD4+ CD25+ FoxP3+ T cells in
the spleen of mice. The average Treg proportions in the control,
model and BSHXF-treated groups were 5.033±1.312, 4.034±0.975 and
4.810±1.222%, respectively. Following administration of BSHXF, the
Treg level was higher than that of the model groups. (B) Effect of
BSHXF on the expression of FoxP3+ T cell mRNA in the
spleen of mice. The FoxP3+ mRNA level in the spleen in
model mice was 0.373±0.108, which was significantly decreased
compared with that in the control group mice (P<0.001).
Furthermore, the FoxP3+ mRNA expression level in the
spleen in the BSHXF-treated group was 1.704±0.222, which was
significantly increased compared with that in the control and model
groups (P<0.001). **P<0.01 and ***P<0.01 vs. the control
group; #P<0.05 and ###P<0.001 vs. the
model group. BSHXF, Bu Shen Huo Xue formula; Treg, regulatory T
cells; FoxP3, forkhead box P3. |
RT-qPCR analysis of FoxP3 mRNA
expression in the spleen
As demonstrated in Fig.
7B, FoxP3 mRNA in the spleen of model mice was significantly
decreased compared with that of the control group mice
(P<0.001). Treatment with BSHXF significantly increased FoxP3
mRNA levels in the spleen compared with those in the model and
control groups (P<0.001).
Discussion
Autoimmunity may be closely associated with the
pathogenesis of POF. A previous study indicated that human
autoimmune oophoritis is a pathological entity encountered in 5% of
women with POF (14). Autoimmune
oophoritis is the main mechanism of ovarian immune damage. ZP
antigen immunization results in increasing atretic follicles and
disorders of the local immune milieu in the ovaries, which
facilitates depletion of follicles in different phases and leads to
POF. The induction of autoimmune oophoritis in B6AF1 mice using ZP3
has been described in the literature (11,15,16).
Therefore, this model with the same dosage was selected for use in
the present study. The results indicate that the estrous cycle was
dysfunctional in the model mice, with a lower ovarian and uterine
viscera coefficient and an atrophied ovary and endometrium. Ovarian
pathology revealed inflammatory cell expression, which generated
more atretic follicles. Ovarian follicles were infiltrated by
inflammatory cells and anti-ZP antibody was expressed.
Additionally, immunohistochemical results for the ovaries revealed
that ER and ER expression was downregulated. Furthermore, ovarian
follicles were surrounded by a large number of infiltrating
CD4+ T lymphocytes. These indicated that the model of
autoimmune POF was successfully established.
CD4+ T lymphocytes are the main cells
involved in cellular immunity, and their activation can cause
immune damage (3,4). Additionally, CD4+
CD25+ Tregs belong to a T-lymphocyte subset with an
immunity regulation function. They have an important function in
the maintenance of a stable internal environment and self-immune
tolerance. Furthermore, the reduction or dysfunction of these cells
may be correlated with autoimmune diseases (17,18). A
previous study demonstrated that Tregs are able to prevent and even
reverse autoimmune disease progression in mice (19). FoxP3, which is closely associated
with the development and function of Tregs, is essential for Treg
cell differentiation and the functioning of transcription factors
(20). In the present study, FCM
methods demonstrated that CD4+ CD25+ Treg
cells were decreased in model mice. Compared with the control group
mice, FoxP3+ mRNA levels in the spleen were
significantly decreased in model mice. The results of the present
study imply that autoimmune POF may be associated with T lymphocyte
levels, and be particularly closely correlated with Treg
levels.
Inflammation is important in the progression of
autoimmune POF (21). IL-10
significantly inhibits the synthesis of mononuclear cells and the
release of inflammatory mediators (22). Furthermore, IFN-γ, a potent
immunomodulatory cytokine, is a member of the IFN family that
regulates immune responses through the activation of mononuclear
macrophages (23). The results of
the present study demonstrate that serum levels of IL-10 and IFN-γ
increased in the model mice. The data of the present study
indicated that increases in inflammatory and anti-inflammatory
factors increased with the occurrence of the inflammatory response.
Similar results have been observed for other autoimmune diseases
(24).
Experimental studies have revealed that Bu Shen Huo
Xue inhibits the specific immune damage of residual follicles in
the ovaries, thereby restoring ovarian function in mice with POF,
with additional preventive effects on the incidence of POF
(25,26). Earlier studies have demonstrated that
traditional Chinese medicine is able to attenuate immune injury.
Angelica Sinensis, Rehmanniae Radix Preparata, Radix
Paeoniae Alba and Ligusticum Wallichii in the Bu Shen Huo
Xue formulation are the main components of Siwutang.
Pharmacological experiments have revealed that Siwutang enhances
immune function, improves the spleen and the thymus index of aging
mice, and regulates multiple functional states of the immune system
(27). Additionally, an Angelica
Sinensis extract is able to increase the number of immune cells
and regulate immune functions (28).
The peoniflorin in Radix Paeoniae Alba and ligustrazine and ferulic
acid in Ligusticum Wallichii have been demonstrated to
promote the transformation and proliferation of lymphocytes in mice
(29,30). Furthermore, Semen Cuscutae and Herba
Epimedii have immunoregulatory functions (31,32).
These findings indicate that numerous compounds present in BSHXF
contribute to immune function.
The results of the present study demonstrated that
BSHXF elevated the ovarian index, increased the number of mature
follicles, reduced the levels of anti-ZP antibodies, promoted the
expression of ER and PR and improved ovarian functions. In
addition, it decreased the spleen index in mice with autoimmune POF
(although not significantly), and effectively inhibited the
activation of CD4+ T lymphocytes, increased Treg cells
and promoted the mRNA expression of FoxP3. Serum of mice treated
with BSHXF was used to cultivate CD4+ CD25+
Treg cells separated from the spleen, and the proliferation of the
cells was increased. Therefore, it is considered that BSHXF may
improve the immune function and status of autoimmune POF mice by
increasing Treg cells.
The present study provides a basis for further
research into the efficacy of BSHXF in the treatment of POF.
However, the BSHXF formulation is complex and the components exert
multi-targeted effects. Thus, other mechanisms by which BSHXF
influences disease in POF mice remain poorly understood and require
further investigation.
In the present study, the mechanisms of autoimmune
POF are indicated to be associated with a reduction in the number
of CD4+ CD25+ Treg cells and increased
activation of CD4+ T lymphocytes, leading to an
excessive immune response of the ovaries. BSHXF may exert
significant protective immunity on POF in mice by increasing the
number of CD4+ CD25+ Treg cells to inhibit
the activation of CD4+ T lymphocytes.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81303136), Jiangsu
Province Natural Science Foundation of China (grant no.
SBK20151606) and Jiangsu Province Bureau of Traditional Chinese
Medicine (grant no. LZ13088). The authors would like to thank
Professor Jian Zhang, Professor Peng Cao and Professor Meng Cao for
their invaluable help in the processing of the experiment.
References
|
1
|
Beall SA and DeCherney A: History and
challenges surrounding ovarian stimulation in the treatment of
infertility. Fertil Steril. 97:795–801. 2016. View Article : Google Scholar
|
|
2
|
Wu X, Cai H, Kallianpur A, Li H, Yang G,
Gao J, Xiang YB, Ji BT, Yu Tang, Zheng W and Shu XO: Impact of
premature ovarian failure on mortality and morbidity among Chinese
women. PLoS One. 9:e895972014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chernyshov VP, Radysh TV, Gura IV,
Tatarchuk TP and Khominskaya ZB: Immune disorders in women with
premature ovarian failure in initial period. Am J Reprod Immunol.
46:220–225. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ban H: Peripheral blood cell immune
detection and its clinical significance of premature ovarian
failure patients. China Modern Med. 17:84–85. 2010.
|
|
5
|
Shi X, Shu Q and Zhang J: Detection and
analysis of autoimmunity data on patients of premature ovarian
failure. Prog Obstet Gvnecol. 16:591–593. 2007.(In Chinese).
|
|
6
|
Xie J, Dong Y, Liang Z and He W: Changes
and significances of CD4+CD25+T regulatory
cells in patients with premature ovary failure. Reprod Contracep.
33:224–227. 2013.
|
|
7
|
Farquhar C, Marjoribanks J, Lethaby A,
Suckling JA and Lamberts Q: Long term hormone therapy for
perimenopausal and postmenopausal women. Cochrane Database Syst
Rev: CD004143. 2009. View Article : Google Scholar
|
|
8
|
Kalantaridou SN, Braddock DT, Patronas NJ
and Nelson LM: Treatment of autoimmune premature ovarian failure.
Hum Reprod. 14:1777–1782. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu N: Traditional Chinese medicine
treatment of premature ovarian failure in clinical research. Hunan
J Tradit Chin Med. 24:94–96. 2006.
|
|
10
|
Tang CL, Li F, Sun L and Li DJ:
Therapeutic Effect of Bushen Huoxue recipe on autoimmune premature
ovarian failure mice established by immunization with recombinant
porcine zona pellucida 4 antigen. Chin J Integr Med. 19:439–445.
2003. View Article : Google Scholar
|
|
11
|
Fu L, Zhao Y and Li S: The establishment
of the animal model of premature ovarian failure. J Reprod Med.
15:179–183. 2006.(In Chinese).
|
|
12
|
Cora MC, Kooistra L and Travlos G: Vaginal
cytology of the laboratory rat and mouse: Review and criteria for
the staging of the estrous cycle using stained vaginal smears.
Toxicol Pathol. 43:776–793. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
La Marca A, Brozzetti A, Sighinolfi G,
Marzotti S, Volpe A and Falorni A: Primary ovarian insufficiency:
Autoimmune causes. Curr Opin Obstet Gynecol. 22:277–282.
2010.PubMed/NCBI
|
|
15
|
Rhim SH, Millar SE, Robey F, Luo AM, Lou
YH, Yule T, Allen P, Dean J and Tung KS: Autoimmune disease of the
ovary induced by a ZP3 peptide from the mouse zona pellucida. J
Clin Invest. 89:28–35. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kimura S, Matsumoto T, Matsuyama R, Shiina
H, Sato T, Takeyama K and Kato S: Androgen receptor function in
folliculogenesis and its clinical implication in premature ovarian
failure. Trends Endocrinol Metab. 18:183–189. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Belkaid Y: Regulatory T cells and
infection: A dangerous necessity. Nat Rev Immunol. 7:875–888. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mottet C, Uhlig HH and Powrie F: Cutting
edge: Cure of colitis by CD4+CD25+ regulatory
T cells. J Immunol. 170:3939–3943. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vojdani A and Erde J: Regulatory T cells,
a potent immunoregulatory target for CAM researchers: Modulating
tumor immunity, autoimmunity and alloreactive immunity (III). Evid
Based Complement Alternat Med. 3:309–316. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shevach EM: Mechanisms of
foxp3+ T regulatory cell-mediated suppression. Immunity.
30:636–645. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Said RS, El-Demerdash E, Nada AS and Kamal
MM: Resveratrol inhibits inflammatory signaling implicated in
ionizing radiation-induced premature ovarian failure through
antagonistic crosstalk between silencing information regulator 1
(SIRT1) and poly(ADP-ribose) polymerase 1 (PARP-1). Biochem
Pharmacol. 103:140–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Polz-Dacewicz M, Strycharz-Dudziak M,
Dworzański J, Stec A and Kocot J: Salivary and serum IL-10, TNF-α,
TGF-β, VEGF levels in oropharyngeal squamous cell carcinoma and
correlation with HPV and EBV infections. Infect Agent Cancer.
11:452016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J and Ivashkiv LB: IFN-γ abrogates
endotoxin tolerance by facilitating Toll-like receptor-induced
chromatin remodeling. Proc Natl Acad Sci USA. 107:pp. 19438–19443.
2010; View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nie X, Deng R, Xiang L, Jiang P and Xue Q:
Reno-protective effect and mechanism study of Huang Lian Jie Du
Decoction on lupus nephritis MRL/lpr mice. BMC Complement Altern
Med. 16:4482016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cai LR, Li DJ and Sun XX: Experimental
study on preventive and therapeutic effect of Bushen Huoxue recipe
on autoimmune premature ovarian failure model. Zhongguo Zhong Xi Yi
Jie He Za Zhi. 21:126–129. 2001.(In Chinese). PubMed/NCBI
|
|
26
|
Dong L, Jiang L, Men W and Zhu NS:
Preventive and therapeutic effects of Bushen Huoxue Recipe on
autoimmune premature ovarian failure in mice. Zhong Xi Yi Jie He
Xue Bao. 6:294–297. 2008.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang H, Zhu M, Sun Y, Wang X and Xu J:
The immunoregulatory research progress of Sijunzi decoction and
siwu decoction. Inf Traditional Chin Med. 29:136–137. 2012.(In
Chinese).
|
|
28
|
Liu SP, Dong WG, Wu DF, Luo HS and Yu JP:
Protective effect of angelica sinensis polysaccharide on
experimental immunological colon injury in rats. World J
Gastroenterol. 9:2786–2790. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu Y and Dong Y: Radix paeoniae alba total
glycosides on mice lymphocytes in vitro value-added and active
research. Chin Arch Traditional Chin Med. 31:914–917. 2013.
|
|
30
|
Zhang T, Zhao R, Liu Z, Shang Y, Wang M
and Wang X: Ligustrazine effects on T lymphocyte activation value.
Chin J Gerontol. 29:1658–1659. 2009.(In Chinese).
|
|
31
|
Zhou P: Epimedium immune pharmacological
research progress. Chin J Traditional Med Sci Technol. 17:279–280.
2010.
|
|
32
|
Lin HB, Lin JQ, Lin Jian-Qun, Lu N and Yi
XY: Comparative study on immune enhancement effects of four kinds
of dodder seeds in Shandong province. Zhong Xi Yi Jie He Xue Bao.
1:51–53. 2003.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|


















