Introduction
Ischemic stroke is the primary cause of
cerebrovascular diseases and remains to be one of the leading
causes of death and disability in patients worldwide (1,2). Despite
tremendous research efforts leading to improvements in ischemic
stroke treatment, the available therapeutic strategies are
currently limited, and several promising agents evaluated in
extensive preclinical trials failed to enter clinical trials
(3,4). At present, thrombolytic therapy remains
to be the gold standard for ischemic stroke treatment; however, it
is only efficacious within a narrow therapeutic window (1,2,4,5). Hence,
beyond the therapeutic window, management of stroke mainly depends
on supportive therapy, secondary prevention and rehabilitation
(1). Thus, an enhanced understanding
regarding the pathological processes may provide novel therapeutic
options to promote stroke recovery.
Ischemic stroke is a highly complex pathological
process, which induces a series of cellular and molecular events
and may be divided into three stages: The acute phase (hours), the
subacute phase (hours to days) and the chronic phase (days to
months) (6). In the acute stage,
metabolic disturbances and excitotoxicity are involved in the
progression of neuronal damage; in the subacute period,
inflammation and cell death are the dominant events; in the chronic
phase, brain repair is the major response and includes
microgliosis, glial scar formation, angiogenesis, as well as
neuronal regeneration (6–8). In addition, during the pathological
process of ischemic stroke, the subacute and chronic phases are
generally termed as late (subacute/chronic) phases (6,9). In the
late phase, one of the dominant responses is reactive astrogliosis
with subsequent glial scar formation, which not only protects the
neurons against harmful substances by isolating the injury area but
also obstructs neuronal regeneration by suppressing axonal
sprouting (10,11). Another important event is the
activation of microglia, which endangers neuronal survival by
releasing numerous proinflammatory and neurotoxic mediators
(8,12,13).
Recently, considerable attention has been paid to
Traditional Chinese Medicine, as it includes sources of
neuroprotective components (14).
Natural products, particularly medicinal plants, may provide an
ideal choice for the development of safe and effective drugs for
stroke treatment (14). Paeonol, an
active component isolated from the Chinese herbal medicine Cortex
Moutan, which is the root bark of Paeonia suffruticosa
Andr., has been demonstrated to possess diverse pharmacological
activities, including anti-oxidant (15), anti-atherosclerotic (16), anti-tumor (17), anti-diabetic (18), and anti-inflammatory effects
(19). Regarding its applicability
for central nervous system diseases, previous studies have
demonstrated that paeonol exerts neuroprotective actions against
acute ischemic stroke, as well as Parkinson's and Alzheimer's
disease in animal models, (19–22).
These studies indicate that paeonol may be a promising drug for the
treatment of neurological disorders. However, to the best of our
knowledge, the potential effects of paeonol on subacute/chronic
ischemic stroke have remained to be determined. The present study
pursued to investigate the therapeutic potential of paeonol in
middle cerebral artery occlusion (MCAO)-induced subacute/chronic
cerebral ischemia. Furthermore, the present study focused on the
post-ischemic microglial and astrocyte responses in the rat brain
in an attempt to elucidate the underlying mechanisms.
Materials and methods
Experimental animals
A total of 125 adult male Sprague Dawley rats
weighing 230–280 g (10–12 weeks old) were obtained from the Animal
Science Center of Zhejiang Academy of Medical Sciences, [Hangzhou,
China; certificate no. SCXK (Zhe) 2014-0001]. Animals were housed
in an animal center at a constant temperature of 22±2°C, a relative
humidity of 50±10% and a 12-h light/dark cycle. They were allowed
free access to food and water. Behavioral experiments were arranged
at 10:00 a.m.-05:00 p.m. during the day. All efforts were made to
minimize their suffering. All experimental procedures were
performed in accordance with the National Institutes of Health
Guidelines for the Care and Use of Experimental Animals and were
approved by the Animal Care and Use Committee of Zhejiang
University (Hangzhou, China).
Induction of transient focal cerebral
ischemia
Transient focal cerebral ischemia was induced by
MCAO according to previous methods (23) with certain modifications, such as
using the nylon suture with a poly-L-lysine coated and without
performing the ligation of the pterygopalatine artery (9,24). In
brief, after animals were anesthetized by intraperitoneal (i.p.)
injection chloral hydrate (400 mg/kg), and the left common carotid
artery, the left external carotid artery (ECA) and the left
internal carotid artery (ICA) were isolated. A nylon suture coated
with poly-L-lysine was inserted into the ICA from the ECA until a
slight resistance was felt, which indicated blockage of the MCA
origin. Occlusion was performed for a period of 30 min, after which
the suture was removed to allow for reperfusion. After the surgery,
the animals were placed in a warm box and allowed to recover from
anesthesia. The sham animals underwent the same surgical procedures
without insertion of the suture. During the experiment, the change
in regional cerebral blood flow (rCBF) was monitored as described
previously with the steady-state baseline prior to the operation
being regarded as 100% (9,25). In addition, the blood pressure,
PaO2, PaCO2, glucose and pH were continuously
monitored as described previously during the entire course of the
operation (9,25). At the designated endpoint, all
animals were euthanized by inhalation of CO2, and
additional animals were allocated to ensure that a sufficient
number of animals survived until the designated endpoint. Animals
exhibiting signs including subarachnoid hemorrhage, or a moribund
or comatose state with labored respiration, were excluded and
euthanized with CO2 (26,27).
Drug administration and groups
Paeonol was obtained from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany) with a purity of >99% and dissolved in
saline at a concentration of 2 mg/ml. Based on a preliminary
experiment, a dose of 25 mg/kg paeonol (i.p.) was used in the
present study, which exerted the best neuroprotective effect in
acute cerebral ischemia [2,3,5-triphenyltetrazolium chloride (TTC)
staining and histopathological results; data not shown]. The
regimen of drug administration applied in the present study was
established in previous studies (9,24).
To evaluate the effects of paeonol on subacute
ischemic injury, the animals were randomly divided into a sham
group (saline, n=16), a model group (MCAO + saline, n=23) and a
paeonol-treated group (MCAO + Pae at 25 mg/kg, n=21). For drug
delivery, the same volume of saline or paeonol solution was
injected i.p. at the onset of MCAO (when the nylon suture was
inserted and the MCA origin was blocked), and then injected once a
day for 3 days. During the experiment, one part of the animals
(n=8) were used for determination of infarct volume, and the
remaining animals were prepared to cut frozen sections for
immunostaining. The 10-µm frozen sections at 2-mm intervals from
the frontal to the occipital poles were cut by cryomicrotomy
(CM1900, Leica, Wezlar, Germany).
To evaluate the effects of paeonol on chronic
ischemic injury, the animals were randomly divided into a sham
group (saline, n=16), a model group (MCAO + saline, n=26) and a
paeonol-treated group (MCAO + Pae at 25 mg/kg, n=23) in another
separate experiment. For drug delivery, the same volume of saline
or paeonol was injected i.p. at the onset of MCAO, then injected
once a day from days 2–7, and then once every 2 days from days
8–28.
Behavioral assessment
The neurological deficit score was determined at the
indicated time-points according to the established scoring system:
In the absence of neurological deficits, the score of 0 was given,
upon failure to extend right paw fully, the score of 1 was given,
animals circling to the right received the score of 2, falling to
the right was scored as 3, and no spontaneous walking and depressed
levels of consciousness was scored as 4 (23). The test was repeated for three times
and the average value was recorded.
The inclined board test was performed to evaluate
the balance and coordination of animals according to previous
research (28) with certain
modifications, including the material and size of the board, and
the rotation angle (24). Animals
were placed on the board (50×30 cm), and once they were stable, the
board was inclined from horizontal to vertical at a rate of 2°/sec.
The holding angle was defined as the angle at which the animal fell
off the board. The test was repeated for three times and the
average value was calculated.
One observer who was blinded to the experimental
groups performed all of the assessments.
Determination of infarct volume
After the neurological assessment, one part of the
animals was re-anesthetized, and the brains were quickly removed
and coronally sliced into 6 sections of 2 mm in thickness. The
slices were then incubated in 0.5% 2,3,5-triphenyltetrazolium
chloride (TTC) at 37°C for 20 min, followed by fixation in 4%
paraformaldehyde for 2 h. Subsequently, images of the fixed slices
were captured. The infarction area was identified as the unstained
area in the brain sections. The infarct volume was calculated by
summing up the volumes of the 6 slices, and represented as the
percentage of infarction in the total brain hemisphere.
Histological analysis
Following the behavioral tests, the remaining
animals were re-anesthetized and then transcardially perfused with
4% paraformaldehyde after a pre-flush with saline. The brains were
quickly removed, and were immediately immersed in 4%
paraformaldehyde for 24 h, followed by dehydration in 30% sucrose
for 3 days, then images were captured with a digital camera.
Finally, a series of sections (10 µm) from the frontal to the
occipital poles were cut by cryomicrotomy (CM1900; Leica
Microsystems, Wetzlar, Germany). The sections were prepared for
immunostaining.
To determine the changes in the number of different
cell types, immunofluorescence staining was performed. The 10-µm
slides were rinsed with PBS and then incubated with 10% normal goat
serum (Zhongshan Belling Biotechnology Co., Ltd., Beijing, China)
for 2 h at room temperature. Subsequently, the sections were
incubated at 4°C overnight with the following primary antibodies:
Mouse anti-glial fibrillary acidic protein (GFAP; cat. no. MAB3402;
1:800 dilution; EMD Millipore, Billerica, MA, USA), mouse
anti-neuronal nuclei (NeuN; cat. no. MAB377; 1:200 dilution; EMD
Millipore) and rabbit anti-ionized calcium binding adaptor molecule
1 (Iba1; cat. no. 019-19741; 1:1,000 dilution; Wako Pure Chemical
Industries, Ltd., Wako, Japan). Thereafter, sections were washed
three times in PBS and incubated with fluorescein
isothiocyanate-conjugated secondary antibody (1:200 diluton; cat.
nos. AP132F or AP124F; EMD Millipore) for 2 h at room temperature.
For the negative control, PBS was applied instead of the primary
antibodies. Images of the stained sections were captured using a
fluorescence microscope (Olympus BX51: Olympus, Tokyo, Japan).
Eight non-overlapping images for each site of the same rats were
randomly selected, and the average value was determined. The
neurons and the microglia were calculated according to the average
number of stained cells and the astrocytes were calculated
according to the average fluorescence intensity.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Significance of differences was assessed by one-way
analysis of variance, followed by Dunnett's post-hoc test (SPSS
10.0 for Windows; SPSS, Inc., Chicago, IL, USA). The results of the
behavioral assessments were analyzed using the Kruskal-Wallis test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Physiological changes after the
operation
No significant differences in the physiological
parameters, including mean arterial blood pressure, partial
pressure of CO2 (PaCO2), PaO2,
blood glucose, pH and weight were identified between 30 min prior
to and after the operation. After cerebral ischemia, the rCBF was
~70% decreased at 30 min after occlusion, and then returned to
baseline levels after reperfusion (P<0.01; Table I). Paeonol did not affect these
physiological variables, including the reduction of rCBF (Table I). During the 28 days, all of the
sham animals were survived. However, no significant difference in
the survival rate was identified between the model group (61.54%)
and the paeonol-treated group (69.57%), and these rates were
consistent with those reported by previous studies (29).
 | Table I.Physiological variables prior to and
after operation. |
Table I.
Physiological variables prior to and
after operation.
|
|
| Ischemia |
|---|
|
|
|
|
|---|
| Variable | Sham | Model | Pae |
|---|
| Body weight
(g) | 262.3±11.4 |
272.8±14.3 | 264.4±11.6 |
| MABP (mmHg) |
|
|
|
|
Baseline | 105.2±10.1 | 102.3±8.9 | 110.0±14.7 |
| 30 min
after reperfusion | 111.8±11.2 |
110.1±14.3 | 118.5±10.2 |
| PaO2
(mmHg) |
|
|
|
|
Baseline | 109.2±4.1 | 105.2±4.5 | 103.8±5.3 |
| 30 min
after reperfusion | 104.7±3.0 |
99.8±4.1 | 101.2±5.6 |
| PaCO2
(mmHg) |
|
|
|
|
Baseline | 38.3±5.1 |
40.4±6.2 | 39.4±4.1 |
| 30 min
after reperfusion | 41.2±3.9 |
42.8±4.3 | 44.3±4.9 |
| pH |
|
|
|
|
Baseline |
7.45±0.38 |
7.38±0.41 |
7.39±0.43 |
| 30 min
after reperfusion |
7.40±0.42 |
7.48±0.39 |
7.46±0.36 |
| Glucose (g/l) |
|
|
|
|
Baseline |
6.58±0.81 |
6.85±0.64 |
6.12±0.53 |
| 30 min
after reperfusion |
7.10±0.75 |
6.99±0.68 |
7.08±0.65 |
| rCBF (%) |
|
|
|
|
Baseline | 100 | 100 | 100 |
| 30 min
after ischemia | 98.5±6.2 |
35.8±7.5a |
34.6±5.7a |
| 30 min
after reperfusion | 98.9±5.8 |
93.6±8.3 | 95.1±7.9 |
Effect of paeonol on behavioral
impairment
In the subacute experiment, behavioral impairments
were evaluated at 24 (data not shown) and 72 h after reperfusion
(Fig. 1). It was demonstrated that
the neurological deficit scores and the holding angle were
aggravated at 72 h after reperfusion. Compared with those in the
sham group, the neurological deficit scores were significantly
increased in the MCAO rats, whereas the holding angle in the
inclined board test was significantly decreased (P<0.01;
Fig. 1A and C). By contrast, a
significant alleviation in the behavioral impairment was observed
in the paeonol-treated group compared with that in the model group
(P<0.05; Fig. 1A and C).
In the chronic experiment, the measures of
behavioral impairment, including the neurological deficit score and
the holding angle, were aggravated at 72 h after reperfusion, and
then gradually recovered at 7, 14 and 28 days after reperfusion
(data not shown). Compared with the model group, paeonol
administration led to a more efficient improvement in neurological
scores and the holding angle in rats at 72 h after reperfusion.
However, no significant difference in the behavioral impairments
was identified between these groups from days 28 after reperfusion
due to the chronic recovery (P>0.05; Fig. 1B and D).
Effect of paeonol on infarct volumes
and ischemic/contralateral hemispheric ratio
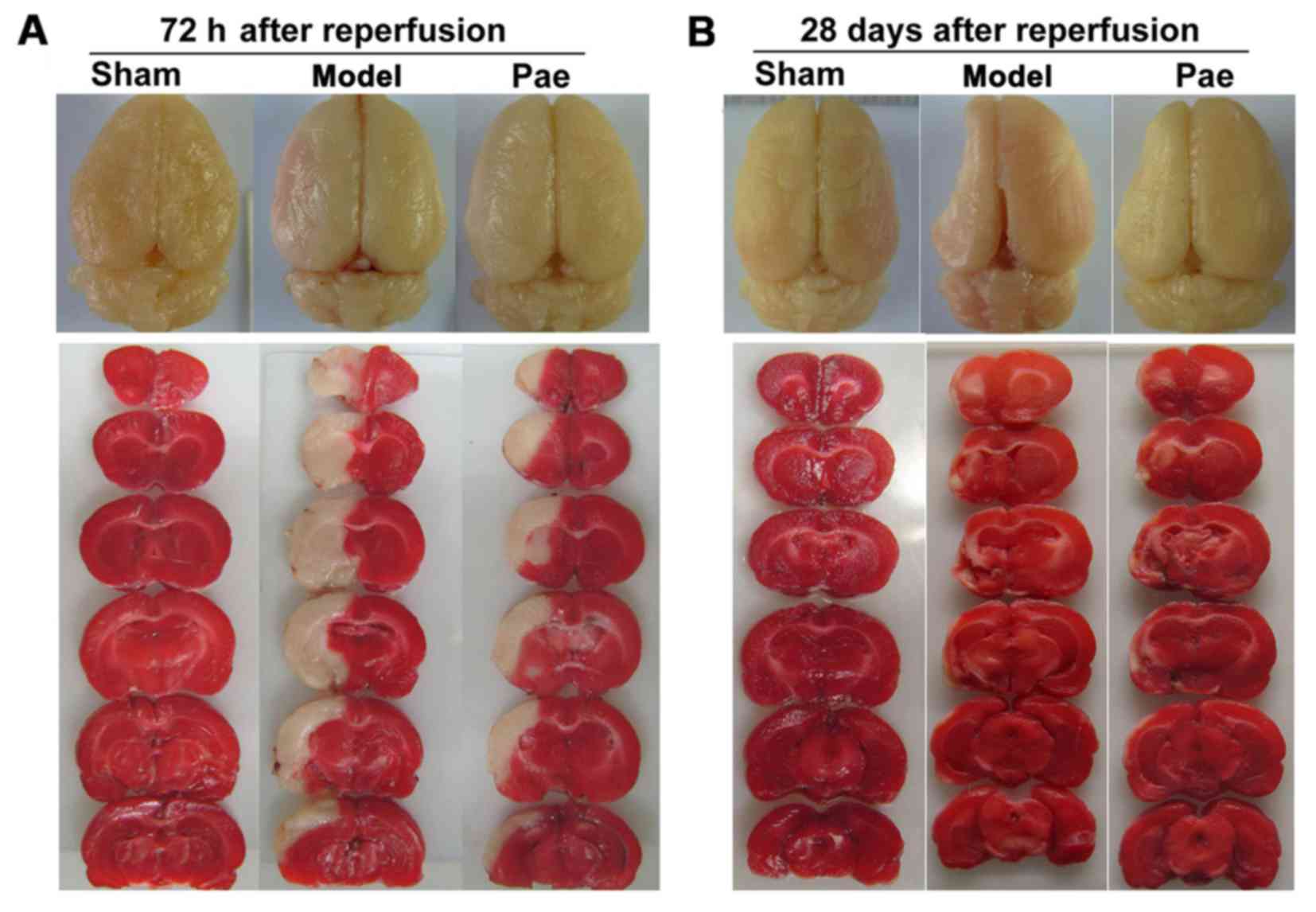
The representative images of total brains and the
TTC-stained coronal slices indicated swelling/atrophy on the
surface and infarctions in the hemispheres after cerebral ischemia,
respectively (Fig. 2A-B).
The effects of paeonol on subacute brain injury at
72 h after reperfusion were assessed. Compared with those in the
sham group, the infarct volume and the ischemic/contralateral
hemispheric ratio, an index of brain edema, were significantly
increased at 72 h after reperfusion (P<0.01; Fig. 1E and G, respectively, and Fig. 2A). However, compared with that in the
model group, paeonol treatment significantly alleviated the
increase of infarct volume and ischemic/contralateral hemispheric
ratio at 72 h after reperfusion (P<0.05 or P<0.01; Fig. 1E and G, respectively, and Fig. 2A). Furthermore, the effects of
paeonol on chronic brain injury were assessed at 28 days after
reperfusion. Compared with that in the sham group, the
ischemic/contralateral hemispheric ratio was markedly decreased at
28 days after reperfusion (P<0.01; Figs. 1H and 2B), indicating brain atrophy, which was
consistent with the results of previous studies (9,29,30). By
contrast, administration of paeonol markedly alleviated chronic
brain atrophy in comparison with that in the model group
(P<0.05; Figs. 1H and 2B). However, no significant differences
were identified in the infarct volumes between the paeonol-treated
and model groups (P>0.05; Figs.
1F and 2B). This may have been
due to the apparent atrophy, which hindered the observation of
further changes, and the chronic functional recovery. Collectively,
these results indicated that paeonol exerted neuroprotective
effects on subacute and chronic cerebral ischemic injury.
Effect of paeonol on neuronal
damage
To assess the effect of paeonol on ischemic neuronal
injury, the changes of neuronal damage in the ischemic brains after
paeonol treatment were observed (Fig.
3). Cerebral ischemia induced evident ischemic lesions
exhibiting neuronal injury, which included the shrinkage of cell
bodies that were deeply stained using Nisssl staining, and the
disappearance of Nissl bodies (data not shown). In the ischemic
core, compared with that in the sham group, the density of neurons
identified by immunofluorescent staining for NeuN was markedly
decreased at 72 h after reperfusion, and disappeared at 28 days
after reperfusion (P<0.01; Fig. 3A, B
and D). In the boundary zone, neuronal density was markedly
decreased at 72 h and 28 days after reperfusion (P<0.01;
Fig. 3A, C and E). Paeonol treatment
significantly ameliorated neuronal loss in the ischemic core, as
well as in the boundary zone at 72 h after reperfusion in
comparison with that in the model group (P<0.01; Fig. 3A-C). In addition, paeonol treatment
markedly inhibited neuronal loss in the boundary zone at 28 days
after reperfusion (P<0.01; Fig. 3A
and E). However, no significant differences in the neuronal
density in the ischemic core area were observed between the
paeonol-treated and model groups, as neurons disappeared at 28 days
after reperfusion in the ischemic core area (Fig. 3A and D). However, a limited number of
degenerated neurons or background staining may be observed
(Fig. 3A and D).
Effect of paeonol on astrocyte
proliferation
To determine the underlying mechanisms involved in
the neuroprotective effects of paeonol on subacute and chronic
ischemic injury, the present study further assessed the astrocyte
responses in the ischemic hemisphere after paeonol administration
(Fig. 4). The results indicated that
the density of GFAP-positive astrocytes was not significantly
changed at 24 h after reperfusion (data not shown). However, at 72
h after reperfusion, astrocytes in the boundary zone were
significantly increased and hypertrophied, and a glial scar
surrounding the ischemic core area had formed at 28 days after
reperfusion (P<0.01; Fig. 4A, C and
E). By contrast, in the ischemic core area, astrocytes were
initially decreased at 72 h after reperfusion, and eventually
disappeared at 28 days after reperfusion (Fig. 4A, B and D). Paeonol treatment
significantly reduced the density of GFAP-positive astrocytes in
the boundary zone at 72 h and 28 days after reperfusion compared
with that in the model group (P<0.01; Fig. 4A, C and E). However, no significant
differences were observed in the density of GFAP-positive
astrocytes in the ischemic core area between the paeonol-treated
and model group at 72 h and 28 days after reperfusion, and the
astrocytes gradually disappeared from this area (Fig. 4A, B and D). Overall, the results
indicated that paeonol exerted its beneficial effects by reducing
post-ischemic astrocyte proliferation.
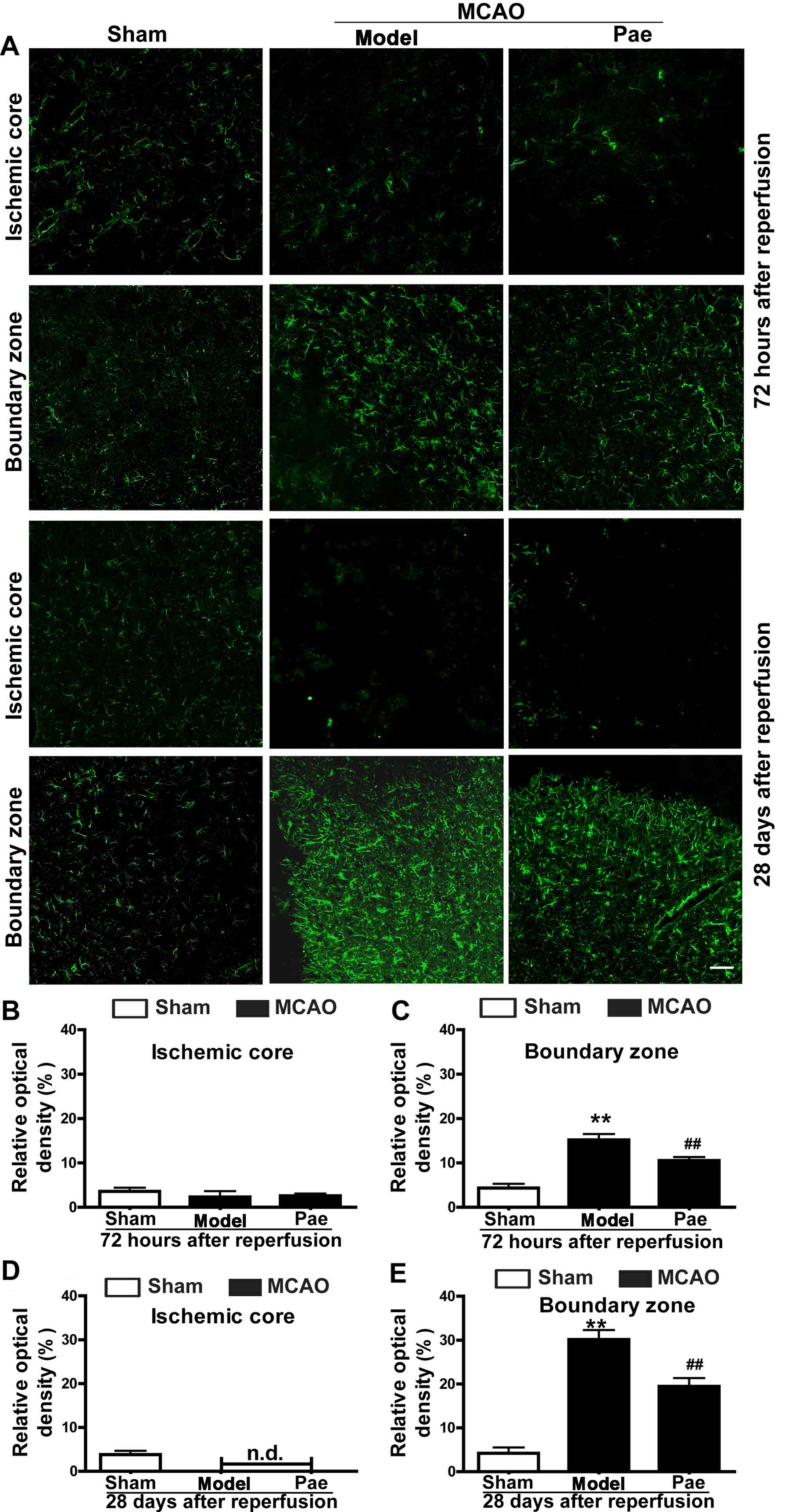 | Figure 4.Effects of paeonol on astrocyte
proliferation at 72 h and 28 days after reperfusion in rats. (A)
Representative photomicrographs indicating glial fibrillary acidic
protein-immunopositive astrocytes at 72 h and 28 days after
reperfusion (scale bar, 100 µm) and (B-E) quantification results.
At 72 h after reperfusion, astrocytes were initially increased in
the boundary zone, and paeonol treatment significantly suppressed
the changes in the boundary zone, but not those in the ischemic
core. At 28 days after reperfusion, astrocytes were significantly
increased in the boundary zone, and paeonol treatment significantly
attenuated the astrogliosis in the boundary zone, but not that in
the ischemic core. Values are expressed as the mean ± standard
error of the mean from eight rats per group. **P<0.01 compared
with sham group, ##P<0.01 compared with model group.
Pae, paeonol; MCAO, middle cerebral artery occlusion; n.d., not
detectable. |
Effect of paeonol on microglial
activation
To evaluate whether microglial activation was
associated with the neuroprotective effects of paeonol on
post-ischemic injury, immunostaining analysis for microglia was
performed. Microglia are inflammatory cells that are rapidly
activated after brain injury. Activation of microglia involves
their proliferation, migration into the injured area, upregulation
of various immunomodulators and phagocytosis of the damaged cells
and debris. In the cerebral cortex of sham rats, ramified
Iba-1-positive microglial cells were diffusely distributed
(Fig. 5A). At 24 h after
reperfusion, the number of Iba-1-positive microglia was not
particularly changed, but the of Iba-1-positive microglia
morphology became irregular (data not shown). However, the number
of Iba-1-positive cells, including that of the ramified microglia
and activated (ameboid/round) microglia/macrophages, was gradually
increased at 72 h after reperfusion compared with that in the sham
group (P<0.01; Fig. 5A-C). In
addition, at 28 days after reperfusion, the number of
Iba-1-positive cells, particularly that of activated
(ameboid/round) microglia/macrophages, was markedly increased in
the ischemic core and boundary zone regions compared with that in
the sham group (P<0.01; Fig. 5A, D
and E). By contrast, compared with that in the model group,
paeonol treatment significantly inhibited the increase in
Iba-1-positive microglia and ameliorated the morphological changes
in the ischemic core, as well as the boundary zone at 72 h and 28
days after reperfusion (P<0.01; Fig.
5A-E). These results indicated the protective effects of
paeonol, which were associated with post-ischemic microglial
activation.
Discussion
Traditional Chinese Medicine has been increasingly
recognized and has demonstrated a therapeutic significance in the
treatment of ischemic stroke (14).
Paeonol, an active phenolic component of Cortex Moutan, has been
demonstrated to possess diverse biological properties (16,17,19–22).
Paeonol has been proved to be effective in treating acute
experimental ischemic stroke by inhibiting the excitotoxicity,
calcium overload and oxidative stress (19,20).
However, to date, the effects of paeonol against subacute/chronic
cerebral ischemic injury have remained elusive. Therefore, the
present study investigated the effects of paeonol on
subacute/chronic cerebral ischemic injury and further determined
the underlying mechanisms.
The amphiphilic structure and low molecular weight
of paeonol facilitate the easy penetration of the blood brain
barrier (19). Thus, in the present
study, paeonol was administered to rats by intraperitoneal
injection, and its effect against subacute/chronic cerebral
ischemic injury was explored. Numerous animal studies on ischemia
have established a model where the injury at 24 h following
ischemia/reperfusion is regarded as acute injury of cerebral
ischemia, that at 72 h to 7 days after reperfusion is regarded as
subacute injury and that at 14–35 days following
ischemia/reperfusion is regarded as chronic injury (6,9,29,31).
Thus, in the present study, the ischemic injury in rats was
observed at 72 h and 28 days after cerebral ischemia/reperfusion.
The behavioral impairments in these rats were also evaluated in 24
h after reperfusion (data not shown). Consistent with the previous
studies, the behavioral impairments were aggravated at 24 h after
reperfusion, and paeonol treatment improved their neurological
deficits. The major results of the present study demonstrated that
the increase in the infarct volume and ischemic/contralateral
hemispheric ratio (edema) in the ischemic hemisphere at 72 h after
reperfusion and the decrease in the ischemic/contralateral
hemispheric ratio (atrophy) at 28 days after reperfusion were
significantly attenuated by paeonol treatment. Furthermore, paeonol
treatment greatly ameliorated the behavioral impairment at 72 h
after reperfusion compared with that in the model group. In
addition, paeonol treatment significantly ameliorated neuronal
damage in the ischemic core and the boundary zone regions at 72 h
after reperfusion, and in the boundary zone at 28 days after
reperfusion. Taken together, it was indicated that paeonol had
protective effects on subacute and chronic cerebral ischemic
injury.
In addition to the above changes, the present study
further investigated the potential mechanisms underlying the
protective effects of paeonol on subacute and chronic ischemic
stroke. As is known, astrocytes and microglia have pivotal roles in
the progression of ischemic stroke (10,32).
Accumulating evidence suggests that glial responses require hours
to days to fully develop, and may in turn provide potential targets
for stroke recovery with longer therapeutic windows compared with
those of other treatments (3,13). In
addition, post-ischemic inflammation is a crucial step during the
process of ischemic stroke (13,33).
Thus, the present study subsequently focused on the effects of
paeonol on inflammation-associated glial cells.
During the post-ischemic phase, one important event
in subacute/chronic ischemic brain injury is reactive astrogliosis
and glial scar formation (10,11). As
a vital part of the neurovascular unit, astrocytes have an
important role in the physiology of the normal brain, which
includes blood circulation, extracellular ionic homeostasis,
release of energy substrates and growth factors, as well as
neurotransmission (3,11). Numerous studies have demonstrated
that astrocytes protect the neurons from glutamate excitotoxicity
during ischemic stroke (10,11). However, increasing evidence has
indicated that astrocytes paradoxically exacerbate ischemic injury
with morphological and phenotypic changes; this process is termed
reactive gliosis or astrogliosis (7,10).
Following focal ischemic injury, reactive astrocytes migrate
towards the boundary zone and then eventually organize into the
glial scar that separates the ischemic core region from healthy
tissue (3,7,10). The
formation of a glial scar is a critical event in the brain repair
responses after ischemic injury, which act as a physical and
biochemical barrier that separates the viable and dead tissues, but
also obstructs neuronal regeneration by suppressing axonal
sprouting (3,10,11).
Growing evidence suggests that modulation of reactive astrocytes
may be a promising strategy for the management of stroke (11). In the present study, paeonol
treatment significantly reduced GFAP-positive astrocytes in the
boundary zone at 72 h and 28 days after reperfusion compared with
those in the model group, but not in the ischemic core, which may
be due to the gradual disappearance of astrocytes in the area.
Different from the mechanisms observed during acute cerebral
ischemic injury, paeonol treatment may regulate the astrocyte
responses and inhibit undesired outcomes, including glial scar
formation, because of its association with the obstruction neuronal
regeneration by suppressing axonal sprouting. Thus, the present
study provided valuable details regarding the role of paeonol in
modulating post-ischemic astrocyte proliferation.
With regard to the changes in the late phase,
another important event in post-ischemic brain injury is
microgliosis and the associated inflammatory responses (13,32,34).
Microglial cells are the resident immune cells and are the first
and major line of active immune defense in the intact or injured
brain (12,13). During neuropathological conditions,
microglial cells are rapidly activated to restore homeostasis of
the central nervous system (12,35). The
activation of microglia involves their proliferation and migration
into the injured area, upregulation of various immunomodulators,
phagocytose of the damaged cell debris and antigenic substances
(8,33,34).
However, microglial activation has dual effects, and uncontrolled
or overactivated microglia are detrimental for the pathological
conditions (35). Activated
microglia may aggravate neuronal damage through the release of
nitric oxide, reactive oxygen species, protease and proinflammatory
cytokines such as tumor necrosis factor-α, as well as
interleukin-1β and −6 (35,36). Chronic microglial activation has been
considered to be the possible underlying mechanism in the neuronal
damage and is associated with numerous neurodegenerative diseases,
including ischemic stroke (12,13).
Therefore, appropriate identification of agents that inhibit
microglial activation may act as effective treatment strategies for
providing neuroprotection (12,35). In
the present study, paeonol treatment significantly inhibited the
increase in Iba-1-positive microglia and ameliorated the
morphological changes in the ischemic core and boundary zone areas
at 72 h and 28 days after reperfusion. Among the various biological
properties, the anti-inflammatory activities of paeonol have been
demonstrated in macrophages, microglia and in the BV2 cell line
in vitro, which was consistent with the present in
vivo results (37,38). It was indicated that paeonol
treatment may regulate microglial activation, suggesting the
potential use of paeonol in the treatment of post-ischemic
microgliosis.
In summary, the present study indicated the
protective effects of paeonol and its involvement in the treatment
of subacute/chronic cerebral ischemia, particularly in the late
phase of microglial activation and astrocyte proliferation. During
subacute cerebral ischemia, paeonol effectively alleviated
neurological impairment, reduced the infarct volume, cerebral edema
and neuronal loss, and suppressed microglial activation and
astrocyte proliferation. During chronic cerebral ischemia, paeonol
markedly ameliorated brain atrophy and neuronal loss, and inhibited
microglial proliferation and astrocyte proliferation. Based on the
above results obtained in rats, understanding the long-term
neuroprotective effects of paeonol in the process of chronic
cerebral ischemia may lead to the development of a novel clinical
treatment for ischemic stroke. Overall, the present study broadened
the current knowledge on the range of pharmacological properties of
paeonol for the treatment of ischemic stroke. However, the precise
molecular mechanisms of the neuroprotective effects of paeonol
require elucidation in future studies.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation (grant nos. 81401566, 31301933,
81301122 and 81400594), the Zhejiang Provincial Natural Science
Foundation (grant no. LQ15H090005) and the Science and Technology
Planning Project of Zhejiang Province (grant nos. 2015KYB076 and
2015ZB012).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BZ, QJS and HW designed the research. BZ, QJS, ZZZ,
HW performed the experiments and data collection. BZ, SYW and XW
analyzed the data. BZ, QJS and HW wrote the manuscript. All authors
have read and approved this version of the article, and ensure the
integrity of the work.
Ethics approval and consent to
participate
All experimental procedures were performed in
accordance with the National Institutes of Health Guidelines for
the Care and Use of Experimental Animals and were approved by the
Animal Care and Use Committee of Zhejiang University (Hangzhou,
China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hankey GJ: Stroke. Lancet. 389:641–654.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jauch EC, Saver JL, Adams HP Jr, Bruno A,
Connors JJ, Demaerschalk BM, Khatri P, McMullan PW Jr, Qureshi AI,
Rosenfield K, et al: Guidelines for the early management of
patients with acute ischemic stroke: A guideline for healthcare
professionals from the American Heart Association/American Stroke
Association. Stroke. 44:870–947. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li L and Stary CM: Targeting glial
mitochondrial function for protection from cerebral ischemia:
Relevance, mechanisms, and the role of MicroRNAs. Oxid Med Cell
Longev. 2016:60323062016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hacke W, Kaste M, Bluhmki E, Brozman M,
Dávalos A, Guidetti D, Larrue V, Lees KR, Medeghri Z, Machnig T, et
al: Thrombolysis with alteplase 3 to 4.5 h after acute ischemic
stroke. N Engl J Med. 359:1317–1329. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shichita T, Ago T, Kamouchi M, Kitazono T,
Yoshimura A and Ooboshi H: Novel therapeutic strategies targeting
innate immune responses and early inflammation after stroke. J
Neurochem. 123 Suppl 2:S29–S38. 2012. View Article : Google Scholar
|
|
6
|
Fagan SC, Hess DC, Hohnadel EJ, Pollock DM
and Ergul A: Targets for vascular protection after acute ischemic
stroke. Stroke. 35:2220–2225. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Burda JE and Sofroniew MV: Reactive
gliosis and the multicellular response to CNS damage and disease.
Neuron. 81:229–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hanisch UK and Kettenmann H: Microglia:
Active sensor and versatile effector cells in the normal and
pathologic brain. Nat Neurosci. 10:1387–1394. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao B, Zhao CZ, Zhang XY, Huang XQ, Shi
WZ, Fang SH, Lu YB, Zhang WP, Xia Q and Wei EQ: The new P2Y-like
receptor G protein-coupled receptor 17 mediates acute neuronal
injury and late microgliosis after focal cerebral ischemia in rats.
Neuroscience. 202:42–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Choudhury GR and Ding S: Reactive
astrocytes and therapeutic potential in focal ischemic stroke.
Neurobiol Dis. 85:234–244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Z and Chopp M: Astrocytes, therapeutic
targets for neuroprotection and neurorestoration in ischemic
stroke. Prog Neurobiol. 144:103–120. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim JY, Kim N and Yenari MA: Mechanisms
and potential therapeutic applications of microglial activation
after brain injury. CNS Neurosci Ther. 21:309–319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jin X and Yamashita T: Microglia in
central nervous system repair after injury. J Biochem. 159:491–496.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bu Y, Lee K, Jung HS and Moon SK:
Therapeutic effects of traditional herbal medicine on cerebral
ischemia: A perspective of vascular protection. Chin J Integr Med.
19:804–814. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding Y, Li Q, Xu Y, Chen Y, Deng Y, Zhi F
and Qian K: Attenuating oxidative stress by paeonol protected
against Acetaminophen-induced hepatotoxicity in mice. PLoS One.
11:e01543752016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu YR, Chen JJ and Dai M: Paeonol
protects rat vascular endothelial cells from ox-LDL-induced injury
in vitro via downregulating microRNA-21 expression and TNF-α
release. Acta Pharmacol Sin. 35:483–488. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li M, Tan SY and Wang XF: Paeonol exerts
an anticancer effect on human colorectal cancer cells through
inhibition of PGE, synthesis and COX-2 expression. Oncol Rep.
32:2845–2853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu J, Feng L, Ma D, Zhang M, Gu J, Wang
S, Fu Q, Song Y, Lan Z, Qu R and Ma S: Neuroprotective effect of
paeonol on cognition deficits of diabetic encephalopathy in
streptozotocin-induced diabetic rat. Neurosci Lett. 549:63–68.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liao WY, Tsai TH, Ho TY, Lin YW, Cheng CY
and Hsieh CL: Neuroprotective effect of paeonol mediates
anti-inflammation via suppressing Toll-like receptor 2 and
Toll-like receptor 4 signaling pathways in cerebral
ischemia-reperfusion injured rats. Evid Based Complement Alternat
Med. 2016:37046472016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao Y, Fu B, Zhang X, Zhao T, Chen L,
Zhang J and Wang X: Paeonol pretreatment attenuates cerebral
ischemic injury via upregulating expression of pAkt, Nrf2, HO-1 and
ameliorating BBB permeability in mice. Brain Res Bull. 109:61–67.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi X, Chen YH, Liu H and Qu HD:
Therapeutic effects of paeonol on
methyl-4-phenyl-1,2,3,6-tetrahydropyridine/probenecid-induced
Parkinson's disease in mice. Mol Med Rep. 14:2397–2404. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou A, Wu H, Pan J, Wang X, Li J, Wu Z
and Hui A: Synthesis and evaluation of paeonol derivatives as
potential multifunctional agents for the treatment of Alzheimer's
disease. Molecules. 20:1304–1318. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi QJ, Wang H, Liu ZX, Fang SH, Song XM,
Lu YB, Zhang WP, Sa XY, Ying HZ and Wei EQ: HAMI 3379, a CysLT2R
antagonist, dose- and time-dependently attenuates brain injury and
inhibits microglial inflammation after focal cerebral ischemia in
rats. Neuroscience. 291:53–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yano T, Anraku S, Nakayama R and Ushijima
K: Neuroprotective effect of urinary trypsin inhibitor against
focal cerebral ischemia-reperfusion injury in rats. Anesthesiology.
98:465–473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Graham SM, McCullough LD and Murphy SJ:
Animal models of ischemic stroke: Balancing experimental aims and
animal care. Comp Med. 54:486–496. 2004.PubMed/NCBI
|
|
27
|
Rewell SS, Churilov L, Sidon TK, Aleksoska
E, Cox SF, Macleod MR and Howells DW: Evolution of ischemic damage
and behavioural deficit over 6 months after MCAo in the rat:
Selecting the optimal outcomes and statistical power for
multi-centre preclinical trials. PLoS One. 12:e01716882017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yonemori F, Yamaguchi T, Yamada H and
Tamura A: Evaluation of a motor deficit after chronic focal
cerebral ischemia in rats. J Cereb Blood Flow Metab. 18:1099–1106.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu LQ, Lou Q, Guo HG, Zhou WW, Ying HZ,
Wei EQ and Shi QJ: Neuroprotective effect of HAMI 3379, a CysLT2R
receptor antagonist, on chronic brain injury after focal cerebral
ischemia in rats. Int J Clin Exp Pathol. 10:4123–4136. 2017.
|
|
30
|
Shen LH, Li Y, Chen J, Cui Y, Zhang C,
Kapke A, Lu M, Savant-Bhonsale S and Chopp M: One-year follow-up
after bone marrow stromal cell treatment in middle-aged female rats
with stroke. Stroke. 38:2150–2156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garbuzova-Davis S, Haller E, Tajiri N,
Thomson A, Barretta J, Williams SN, Haim ED, Qin H, Frisina-Deyo A,
Abraham JV, et al: Blood-spinal cord barrier alterations in
subacute and chronic Stages of a rat model of focal cerebral
ischemia. J Neuropathol Exp Neurol. 75:673–688. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma Y, Wang J, Wang Y and Yang GY: The
biphasic function of microglia in ischemic stroke. Prog Neurobiol.
157:247–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu L, He D and Bai Y: Microglia-mediated
inflammation and neurodegenerative disease. Mol Neurobiol.
53:6709–6715. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Benarroch EE: Microglia: Multiple roles in
surveillance, circuit shaping, and response to injury. Neurology.
81:1079–1088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee Y, Lee SR, Choi SS, Yeo HG, Chang KT
and Lee HJ: Therapeutically targeting neuroinflammation and
microglia after acute ischemic stroke. Biomed Res Int.
2014:2972412014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kawabori M and Yenari MA: The role of the
microglia in acute CNS injury. Metab Brain Dis. 30:381–392. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nam KN, Woo BC, Moon SK, Park SU, Park JY,
Hwang JW, Bae HS, Ko CN and Lee EH: Paeonol attenuates
inflammation-mediated neurotoxicity and microglial activation.
Neural Regen Res. 8:1637–1643. 2013.PubMed/NCBI
|
|
38
|
Himaya SW, Ryu B, Qian ZJ and Kim SK:
Paeonol from Hippocampus kuda Bleeler suppressed the
neuro-inflammatory responses in vitro via NF-kB and MAPK signaling
pathways. Toxicol In Vitro. 26:878–887. 2012. View Article : Google Scholar : PubMed/NCBI
|