Introduction
Two subsets of Th cells, named Th1 and Th2, play a
key role in the regulation of the adaptive immunity (1). Th1 cells majorly present
pro-inflammatory cytokines, including interferon (IFN)-γ,
interleukin (IL)-2 and tumor necrosis factor (TNF)-α, which
activate CD8+ T cells and macrophages, thus promoting cellular
immune reactions. Conversely, Th2 cells largely generate
anti-inflammatory cytokines, such as IL-4, IL-10 and IL-13, as well
as stimulate mast cells, eosinophils and B cells, thereby enhancing
allergic responses and humoral immunity (2). The imbalance between Th1 and Th2 cells
in development and regulation participates in various immune
disorders including hypersensitivity (3).
Delayed-type hypersensitivity (DTH), known as type
IV hypersensitivity reaction, has been demonstrated to be a
Th1-driven immune response, and is associated with contact allergy
with the activity of inflammatory Th1 lineage secreting IFN-γ
(4). Because the hypersensitivity is
also a useful approach for evaluating cell-mediated immune
responses that migrates to the inflammatory site, the experimental
model of ovalbumin (OVA)-induced DTH is widely used to assess the
immunomodulation and anti-inflammatory effects of a serious of new
substances and materials on T-cell immune response (5,6).
As the principal bioactive ingredient, puerarin
generally is isolated from the roots of Pueraria lobate.
Until now, puerarin has been widely indicated to possess a broad
range of pharmacological activities, including anti-cardiotoxicity
anti-neurotoxicity, anti-oxidative, anti-diabetic, anti-tumor and
anti-inflammatory effects (7,8). A
previous study showed that puerarin inhibited the activation of
NF-kB pathway and TNF-α productio in peripheral blood mononuclear
cells in asthma patients, suggesting its protective effect in
allergic disease (9). Utilizing a
mouse model of allergic asthma, a recent study indicated that
puerarin treatment attenuated OVA-induced airway inflammation by
the regulation of eotaxin-3, which suggested its potential
application in allergic inflammation although the detailed
mechanism was unknown (10).
In this study, our results indicated that puerarin
treatment alleviated OVA-induced DTH by modulating Th1/Th2 balance,
resulting from the limitation of Th1 response accompanying
inhibitive production of IFN-γ. These findings suggest puerarin
would be effective alternative to eliminate inflammation associated
with DTH.
Materials and methods
Animals
Female C57BL/6 mice weighing 18–20 g, were purchased
from Hubei Research Center of Laboratory Animals (Wuhan, China) and
housed in an air-conditioned room (23±0.5°C, 12 h light/dark cycle)
with free access to food and water. All experimental procedures
were carried out in accordance with the National Institutes of
Health Guidelines for the Care and Use of Laboratory Animals (NIH
Publication no. 80–23, revised in 1996), and animal handling
followed the dictates of the Animal Care and Use Committee of Hubei
University of Chinese Medicine (SYXK-2012-0067).
DTH
Mice were immunized s.c. with 50 µg OVA (Worthington
Biochemical Corp., Lakewood, NJ, USA) in 100 µl PBS-CFA emulsion
(50 µl complete freund's adjuvant) (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). After 6 days, mice were challenged s.c. 200 µg
heat-aggregated OVA in 20 µl of PBS in the left hind footpad and in
the right footpad with PBS only. After 24 h, OVA-specific DTH was
determined by footpad swelling according to the difference in the
footpad thickness between OVA-challenged footpad and PBS-injected
control footpad.
Groups
48 Mice were randomly assigned to six different
groups: Control group (not immunized), DTH group (OVA-induced DTH),
Puerarin (Pue) + DTH group (low-, middle- and high-dose puerarin
was used during repeated OVA application, respectively),
Dexamethasone (Dex) + DTH group (dexamethasone treatment during
repeated OVA application).
Puerarin treatment
In the therapeutic treatment protocol (Fig. 1A), mice were treated with puerarin
(Sigma-Aldrich; Merck KGaA) via intraperitoneal injection for
continuous 7 days (8 mg/kg for the low dose, 40 mg/kg for the
middle dose and 200 mg/kg for the high dose) (11,12). As
a positive control, dexamethasone (1 mg/kg) (Sigma-Aldrich; Merck
KGaA) was administered intraperitoneally from day 1 to day 7.
Histological analysis
The left hind footpads of mice in the different
group were removed and fixed in 4% paraformaldehyde. Fixed tissues
were embedded in paraffin for 24 h and serially sectioned to a
thickness of 5 µm for histological analysis. Tissue sections were
stained with hematoxylin and eosin (H&E; Sigma-Aldrich; Merck
KGaA) and examined for histopathological change. Pictures were
captured using a Nikon Eclipse Ti-S microscope (Nikon, Tokyo,
Japan). A minimum of three sections per animal experimentation was
examined, and ten visual fields of each sample were randomly
selected to observe the inflammatory cell infiltration in a blinded
manner. The evaluation was performed at ×200 magnification.
Sear collection and anti-OVA IgG
assay
Sera of mice in different groups were collected at
the termination of experiment for the detection of anti-OVA IgG.
Blood from the internal veins of mice in the different group was
collected. After centrifugation at 12,000 × g for 10 min at 4°C,
the serum was used for the ELISA assay of anti-OVA IgG. The levels
of serum anti-OVA IgG were determined using the ELISA kit (3011)
(Chondrex, Seattle, WA, USA) with a micro-plate reader (Thermo
Fisher Scientific, Inc., Waltham, MA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
The left footpad samples of different group were
weighed and homogenized in 1 ml of tissue protein extraction
reagent containing a protease inhibitor cocktail (Pierce; Thermo
Fisher Scientific, Inc.). Homogenates were then centrifuged at
12,000 × g for 15 min at 4°C to obtain the supernatant. Cytokine
such as IFN-γ (MIF00) and IL-4 (M4000B) were measured with the
ELISA kit (R&D Systems, Inc., Minneapolis, MN, USA) according
to the manufacturer's instruction.
Preparation of spleen lymphocytes
Spleen was collected and the mononuclear cells were
isolated based on the method of density gradient centrifugation
using the Ficoll-Hypaque (Sigma-Aldrich; Merck KGaA) (13–15).
Spleen mononuclear cells were isolated by density gradient
centrifugation (2,000 × g for 30 min) (Ficoll-Hypaque density 1.077
g/ml) and incubated overnight at 37°C in complete RPMI-1640 medium
containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.).
Nonadherent cells were harvested as spleen lymphocytes, which
majorly includes T cells.
ConA stimulation assay
Spleen lymphocytes (2×105) were isolated
and co-cultured with ConA (5 mg/ml) (Sigma-Aldrich; Merck KGaA) at
37°C. After 24 h, the ConA stimulation assay was analyzed.
MTT assay
Spleen lymphocytes (5×105) were separated
and co-cultured with OVA (100 µg/ml) at 37°C for 24 h. MTT
(Sigma-Aldrich; Merck KGaA) working solution (0.5 mg/ml) was added
to each well. After 4 h, the plates were centrifugated and dimethyl
sulfoxide (Sigma-Aldrich; Merck KGaA) was added for MTT assay. The
absorbance was detected with a plate reader (hermo Fisher
Scientific, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA from spleen lymphocytes in the different group
was extracted using a Total RNA kit (Tiangen Biotechnology Co.,
Ltd., Beijing, China). The super Moloney Murine Leukemia Virus
Reverse Transcriptase (M-MLV; BioTeke, Beijing, China) was used for
DNA synthesis. qPCR was performed using the SYBR-Green Master Mix
system (Tiangen Biotechnology Co., Ltd.). Following primers were
used: T-bet: Forward, 5′-TGCCTGCAGTGCTTCTAACA-3′ and Reverse,
5′-TGCCCCGCTTCCTCTCCAACCAA-3′; GATA-3: Forward,
5′-GAAGGCATCCAGACCCGAAAC-3′ and Reverse,
5′-ACCCATGGCGGTGACCATGC-3′; IFN-γ: Forward,
5′-AGCGGCTGACTGAACTCAGATTGTAG-3′ and Reverse,
5′-GTCGCTTCGTTGATCACAA-3′; IL-4: Forward,
5′-TCAACCCCCAGCTAGTTGTC-3′ and Reverse, 5′-TGTTCTTCGTTGCTGTGAGG-3′;
GAPDH: Forward, 5′-AACTTTGGCATTGTGGAAGG-3′ and Reverse,
5′-ACACATTGGGGGTAGGAACA-3′. The thermal cycling parameters for the
PCR: denaturation at 95°C for 5 min followed by 40 cycles at 95°C
for 30 sec, 55°C for 30 sec and 72°C for 30 sec. The relative
expression of mRNA was calculated according to 2−∆∆Cq
method.
Western blotting
Spleen lymphocytes were isolated and lyzed with the
buffer (Cell Signaling Technology, Inc., Beverly, Massachusetts,
USA). The total protein concentration in the supernatants was
determined using a BCA Protein Assay kit (Pierce; Thermo Fisher
Scientific, Inc.). The protein (30 µg) were separated by 12% (w/v)
sodium dodecyl sulphate-polyacrylamide gel electrophoresis and
transferred to polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA) using a wet transfer method. The membranes were
blocked with 5% skim milk for 2 h at room temperature and then
incubated overnight at 4°C with the primary rabbit anti-mouse
antibodies against T-bet (sc-21003), GATA-3 (sc-9009) or GAPDH
(sc-25778; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA).
The membranes were incubated with secondary antibody conjugated to
horseradish peroxidase for 2 h (sc-2317; Santa Cruz Biotechnology,
Inc.) for 2 h at room temperature, and the label proteins were
detected with chemiluminescence detection regent (Cell Signaling
Technology, Inc.) and visualized with the detection system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Optical density for each
band was assessed using ImageJ analysis software (National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All data were expressed as mean ± standard
deviation. Statistical analysis was performed with one-way analysis
of variance followed by Duncan's multiple range tests. SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA) was used for all
calculations. P<0.05 was considered to indicate a statistically
significant difference.
Results
Puerarin treatment attenuates
OVA-induced DTH in mice
Firstly, mice model with OVA-induced DTH was
established with or without puerarin treatment (Fig. 1A). In comparison with control group,
repeated OVA application induced an obvious amplification of
footpad swelling (Fig. 1B). Compared
to the DTH group, puerarin treatment, middle and high dose,
inhibited the increment in the swelling thickness (Fig. 1C). Additionally, puerarin treatment
also significantly lowered the levels of OVA-IgG in serum (Fig. 1D), suggesting the protective role of
puerarin on DTH. As the positive control, Dex showed obvious
inhibitory effect on the footpad swelling and the level of
OVA-specific antibody in serum compared with those in DTH
group.
Puerarin decreases local inflammatory
cell infiltration
Inflammatory cells were evidently increased
following OVA-challenge in the dermal tissue (Fig. 2). Compared to the DTH group, puerarin
evidently inhibited the increment of epidermis thickness (Fig. 2B) and reduced inflammatory cell
infiltration (Fig. 2C), which
diminished the number of inflammatory cells recruited to the
footpad associated with the local inflammation.
Puerarin suppresses inflammation
associated with OVA-induced DTH
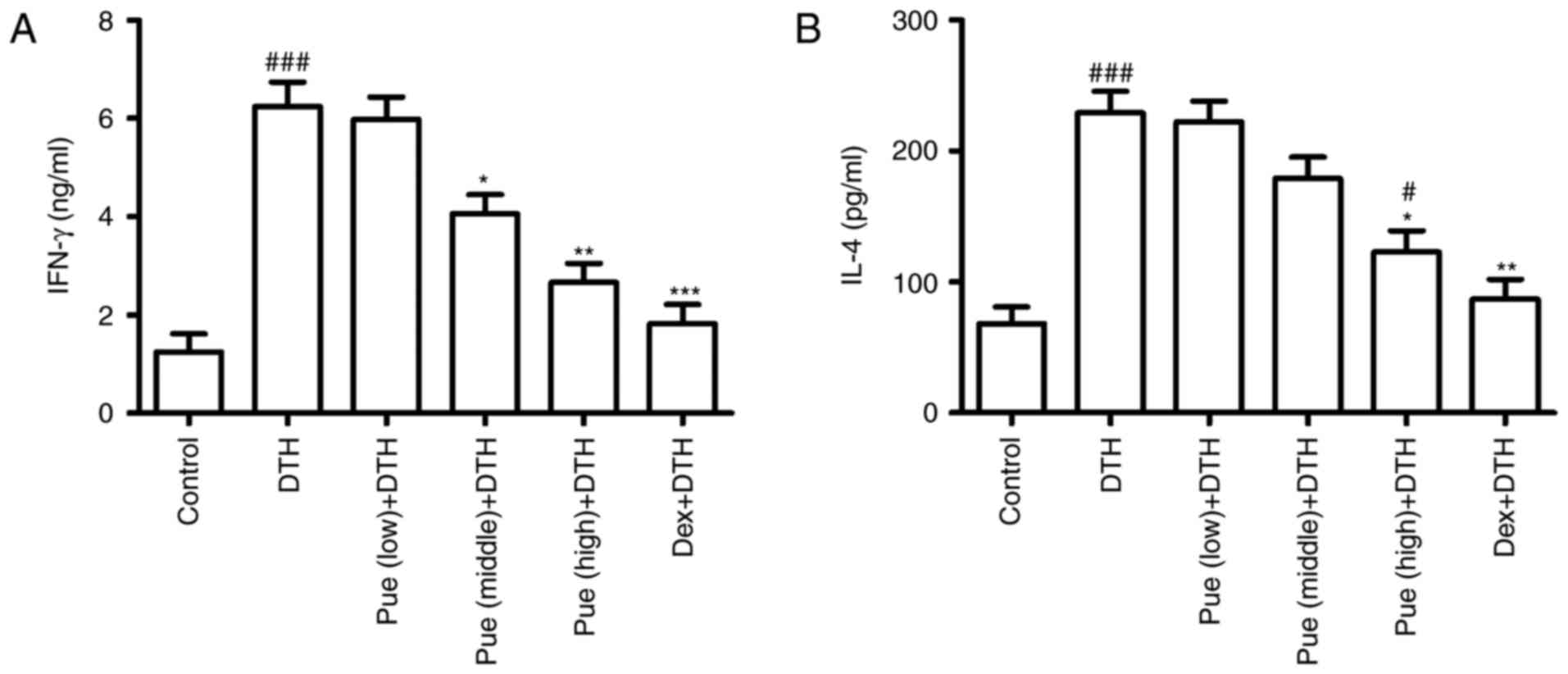
Following puerarin treatment, cytokine associated
with inflammation in OVA-induced DTH was investigated. IFN-γ, the
classic Th1-type cytokines, was found to be greatly down-regulated
in puerarin group compared to that in the DTH group (Fig. 3A), suggesting the anti-inflammatory
effect of puerarin on DTH. Interestingly, only high dose of
puerarin showed suppressive effect in IL-4 production (Fig. 3B), suggesting its anti-allergic
potential of puerarin.
Puerarin inhibits Th1 response
Further, we observed the effect of puerarin on Th1
response based on spleen lymphocytes. Following ConA stimulation,
IFN-γ was increased in the DTH group, but puerarin treatment
significantly inhibited the secretion of IFN-γ (Fig. 4A). In addition, cell proliferation
was no difference between DTH group and puerarin group by the MTT
assay (Fig. 4B). These results
suggest that puerarin can restrain Th1 response, which inhibited
the activation of Th1 cells and led to deduce IFN-γ production,
thereby the regulation of inflammation in OVA-induced DTH.
Effect of puerarin in Th1 and Th2
differentiation
Lastly, the effect of puerarin on the transcription
factors such as T-bet and GATA-3 were investigated, which directly
or indirectly regulate Th1 and Th2 differentiation. As shown in
Fig. 5A-C, middle and high
concentration of puerarin inhibited the expression of T-bet in mRNA
level, whereas only high dose of puerarin showed suppressive effect
for GATA-3 (Fig. 5A-C). In addition,
the similar tendency was found for the expression of IFN-γ and IL-4
in the molecular level (Fig. 5D-F).
Compared with that in the control group, the expression of T-bet
protein was clearly increased in the DTH group. However, high dose
of puerarin significantly reduced the level of T-bet and GATA-3
protein (Fig. 6A and B). These
results suggest that puerarin could regulate a shift of Th1
polarization to Th1/Th2 balance, which is related to the decrement
of T-bet in mRNA and protein levels and subsequently inhibitive
IFN-γ production.
Discussion
Although further development of puerarin has drawn
substantial attention in natural product research, the direct
targets and the underlying mechanisms of its bioactivity remain
unclear. The present study shows that puerarin alleviates
OVA-induced DTH via cytokine inhibition by modulating Th1/Th2
balance, which might provide a new perspective for comprehensive
understanding of its biological effects.
Increasing evidence indicates the anti-inflammatory
effect of puerarin in different disease. Using a murine model of
Parkinson's disease, puerarin significantly alleviated injury in
dopaminergic neurons, which was attributed to its anti-apoptotic
and anti-inflammatory activities, resulting in inhibition of
cleaved Caspase-3 and Bax as well as reduction of inducible nitric
oxide synthase (16). In addition,
puerarin attenuated inflammation and oxidation in mice with
collagen antibody-induced arthritis, which was associated with
reducing production of pro-inflammatory cytokines such as IL-6 and
TNF-α (17). In a sepsis model using
H9c2 cardiomyocytes stimulated with lipopolysaccharide (LPS),
puerarin treatment attenuated the expression of TNF-α and IL-β in
mRNA and protein levels. Cytokine inhibition, especially Th1-type
cytokines including TNF-α and IL-β, contributed to the
anti-inflammatory effect of puerarin (18). In line with above results, our study
showed puerarin restrained the footpad swelling (Fig. 1B and C), inflammatory cell
infiltration in skin tissues (Fig.
2) triggered by OVA application. Importantly, puerarin
treatment significantly hampered IFN-γ production (Fig. 3A), classic Th1-type cytokine, which
suggested its anti-inflammatory effect was associated with
inhibiting Th1 response. The speculate also was confirmed by the
ConA stimulation assay and MTT assay (Fig. 4). Following puerarin treatment, IFN-γ
was decreased and Th1 response was hindered in the presence of
ConA.
As we know, IFN-γ is involved in the allergic
inflammation, such as asthma and DTH (19,20).
However, it is worth noting that IL-4 also participates in the
immune progress (21,22). The Th1/Th2 paradigm assumes that a
dominance of Th1 cell activation and an inadequate Th2 cell
response are responsible for the development of inflammation,
suggesting the balance of Th1/Th2 is important to regulate immune
response (23–25). As the regulatory factor on Th
differentiation, T-bet and GATA-3 has shown the modulating effect
in Th1 and Th2 cells, respectively. T-bet controls the development
and differentiation of Th1 cells as well as induces the synthesis
of IFN-γ, but presents a negative regulation on Th2 cells (26). In this study, puerarin treatment
(middle and high dose, respectively) declined the expression of
T-bet and IFN-γ in mRNA and protein levels, but middle dose of
puerarin showed few effect on the expression of GATA-3 and IL-4
(Fig. 5). Additionally, the ratio of
T-bet/GATA-3 also showed the similar trend as IFN-γ/IL-4 ratio
(Th1/Th2 ratio) (Fig. 6). Based on
these results, we speculated that puerarin treatment could inhibit
the expression of T-bet in mRNA and protein levels, resulting in
the repression of Th1 differentiation, which was responsible for
the immune-switch between Th1 and Th2 cells. Interestingly, high
dose of puerarin also restrained the expression of GATA-3 mRNA and
IL-4 secretion as the similar effect of dexamethasone, suggesting
its anti-allergic potential of puerarin on DTH. If other T-cell
subtype (such as regulatory T cells), transcription factors (such
as NF-κB) or signaling pathway (such as MAPK pathway) participates
in the immuno-shift between Th1 and Th2 cells in the DTH responses
triggered by repeated-OVA administration, further studies are
required to clarify the facts.
In summary, puerarin treatment plays a regulator
role between Th1 and Th2 cells, which restores Th1/Th2 balance
through facilitating an immune-shift from Th1 cells to Th2 cells on
OVA-induced DTH, accompanying regulation of footpad swelling,
anti-OVA IgG, IFN-γ/IL-4 production, Th1 response and T-bet/GATA-3
ratio. These findings suggest the potential therapeutic utility of
puerarin as a candidate in treating Th1-mediated inflammation
disorders such as DTH, although its detailed mechanism needs
further elucidation.
References
|
1
|
Abbas AK, Murphy KM and Sher A: Functional
diversity of helper T lymphocytes. Nature. 383:787–793. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Neurath MF, Finotto S and Glimcher LH: The
role of Th1/Th2 polarization in mucosal immunity. Nat Med.
8:567–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kidd P: Th1/Th2 balance: The hypothesis,
its limitations, and implications for health and disease. Altern
Med Rev. 8:223–246. 2003.PubMed/NCBI
|
|
4
|
Kobayashi K, Kaneda K and Kasama T:
Immunopathogenesis of delayed-type hypersensitivity. Microsc Res
Tech. 53:241–245. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hernández A, Yager JA, Wilkie BN, Leslie
KE and Mallard BA: Evaluation of bovine cutaneous delayed-type
hypersensitivity (DTH) to various test antigens and a mitogen using
several adjuvants. Vet Immunol Immunopathol. 104:45–58. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jacysyn JF, Abrahamsohn IA and Macedo MS:
Modulation of delayed-type hypersensitivity during the time course
of immune response to a protein antigen. Immunology. 102:373–379.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou YX, Zhang H and Peng C: Puerarin: A
review of pharmacological effects. Phytother Res. 28:961–975. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Z, Lam TN and Zuo Z: Radix
Puerariae: An overview of its chemistry, pharmacology,
pharmacokinetics and clinical use. J Clin Pharmacol. 53:787–811.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu XJ, Zhao J and Gu XY: The effects of
genistein and puerarin on the activation of nuclear factor-kappaB
and the production of tumor necrosis factor-alpha in asthma
patients. Pharmazie. 65:127–131. 2010.PubMed/NCBI
|
|
10
|
Wang J, Zhang T, Ma C and Wang S: Puerarin
attenuates airway inflammation by regulation of eotaxin-3. Immunol
Lett. 163:173–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mahdy HM, Mohamed MR, Emam MA, Karim AM,
Abdel-Naim AB and Khalifa AE: The anti-apoptotic and
anti-inflammatory properties of puerarin attenuate
3-nitropropionic-acid induced neurotoxicity in rats. Can J Physiol
Pharmacol. 92:252–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang D, Ma G, Hou M, Zhang T, Chen L and
Zhao C: The Neuroprotective effect of Puerarin in acute spinal cord
injury rats. Cell Physiol Biochem. 39:1152–1164. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Z, Ouyang L, Liang Z, Chen J, Yu Q,
Jeza VT, Gong Y, Shen G, Weng X and Wu X: CD8(low)CD28(-) T cells:
A human CD8 T-suppressor subpopulation with alloantigen specificity
induced by soluble HLA-A2 dimer in vitro. Cell Transplant.
24:2129–2142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu Q, Zhang L, Ouyang L, Gong Y, Liang Z,
Shen G, Weng X and Wu X: A similarity in peptide cross-reactivity
between alloantigen- and nominal antigen-induced CD8+ T cell
responses in vitro. Immunogenetics. 65:173–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weng X, Lu S, Zhong M, Liang Z, Shen G,
Chen J and Wu X: Allo-restricted CTLs generated by coculturing of
PBLs and autologous monocytes loaded with allogeneic
peptide/HLA/IgG1-Fc fusion protein. J Leukoc Biol. 85:574–581.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang X, Xiong J, Liu S, Wang L, Huang J,
Liu L, Yang J, Zhang G, Guo K, Zhang Z, et al: Puerarin protects
dopaminergic neurons in Parkinson's disease models. Neuroscience.
280:88–98. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang C, Wang W, Jin X, Shen J, Hu W and
Jiang T: Puerarin attenuates inflammation and oxidation in mice
with collagen antibody-induced arthritis via TLR4/NF-κB signaling.
Mol Med Rep. 14:1365–1370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan Y, Zhou H, Wu QQ, Li FF, Bian ZY,
Deng W, Zhou MQ and Tang QZ: Puerarin attenuates the inflammatory
response and apoptosis in LPS-stimulated cardiomyocytes. Exp Ther
Med. 11:415–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Raundhal M, Morse C, Khare A, Oriss TB,
Milosevic J, Trudeau J, Huff R, Pilewski J, Holguin F, Kolls J, et
al: High IFN-γ and low SLPI mark severe asthma in mice and humans.
J Clin Invest. 125:3037–3050. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z and Chen Z, Chen T, Yi T, Zheng Z,
Fan H and Chen Z: Berberine attenuates inflammation associated with
delayed-type hypersensitivity via suppressing Th1 response and
inhibiting apoptosis. Inflammation. 40:221–231. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hirose K, Iwata A, Tamachi T and Nakajima
H: Allergic airway inflammation: Key players beyond the Th2 cell
pathway. Immunol Rev. 278:145–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ashraf MI, Shahzad M and Shabbir A:
Oxyresveratrol ameliorates allergic airway inflammation via
attenuation of IL-4, IL-5 and IL-13 expression levels. Cytokine.
76:375–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hirahara K and Nakayama T: CD4+ T-cell
subsets in inflammatory diseases: Beyond the Th1/Th2 paradigm. Int
Immunol. 28:163–171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bui TT, Piao CH, Kim SM, Song CH, Shin HS,
Lee CH and Chai OH: Citrus tachibana leaves ethanol extract
alleviates airway inflammation by the modulation of Th1/Th2
imbalance via inhibiting NF-κB signaling and histamine secretion in
a mouse model of allergic asthma. J Med Food. 20:676–684. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peine M, Rausch S, Helmstetter C, Fröhlich
A, Hegazy AN, Kühl AA, Grevelding CG, Höfer T, Hartmann S and
Löhning M: Stable T-bet(+) GATA-3(+) Th1/Th2 hybrid cells arise in
vivo, can develop directly from naive precursors and limit
immunopathologic inflammation. PLoS Biol. 11:e10016332013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yagi R, Zhu J and Paul WE: An updated view
on transcription factor GATA3-mediated regulation of Th1 and Th2
cell differentiation. Int Immunol. 23:415–420. 2011. View Article : Google Scholar : PubMed/NCBI
|