Introduction
Post-traumatic epilepsy (PTE) is a common type of
acquired epilepsies secondary to traumatic brain injury (TBI),
accounting for approximately 10–25% of patients with moderate to
severe injuries (1). After TBI, the
brain carries out a series of pathological processes, including
neuronal loss, gliosis, axonal sprouting, neurogenesis,
inflammatory response, neurotransmitter release, and mitochondrial
dysfunction, all of which have been connected to the
epileptogenesis (2,3). Evidence indicate that the risk of PTE
increases obviously 10 years after TBI and it usually develops
refractory to medical management (4). At present, there are still many
difficulties in PTE treatment partially because of the pathological
mechanism has not been fully illustrated.
The mammalian target of rapamycin (mTOR) signaling
pathway has been investigated extensively in recent decades.
Experimental data suggest that mTOR pathway is involved in a number
of physiological processes including cell proliferation, protein
synthesis and cortical development (5–8).
However, in recent years, it has also been reported to implicate in
several neurological diseases, such as epilepsy (9). Evidences indicate that the mTOR pathway
is activated in multiple type of epilepsies. For instance,
epileptogenesis of cortical dysplasia, a common type of pediatric
epilepsies, has been attributed to the over activation of mTOR
signaling (9), which is also proven
to involve in the development of temporal lobe epilepsy, the most
common epilepsy in adults (10). The
activation of mTOR protein kinase is regulated by multiple
signaling pathways including PI3K/AKT, Ras/MAPK and AMPK.
Phosphorylation level of downstream protein, such as p70 ribosomal
S6 kinase (P70S6K), increased notably when the mTOR signaling is
triggered (11). Up to now, only few
research concentrates on the relationship between the development
of PTE and the activation of mTOR. Although previous study
demonstrated that mTOR-mediated epileptogenesis was activated via
PI3K-Akt signaling in vitro model of PTE (12). However, they did not further explore
the activation status of mTOR in animal model of PTE, nor did
others.
Anti-epileptic drugs (AEDs) is the predominant
therapy for epilepsy patients, however, about 30% of them are not
sensitive to the treatment. Similarly, therapeutic effects of AEDs
on patients with PTE is close to 35% (13). Additionally, some patients even
suffer from side effects caused by AEDs such as nausea, dizziness,
lethargy and weight gain. Therefore, it is essential to find a
novel therapeutic target for epilepsy patients. Fortunately,
rapamycin, a macrolide antibiotic drug, has been demonstrated
effectively via deactivation of mTOR signaling and then provides
neuroprotective effect against epileptic injury in previous studies
(9,14,15).
Furthermore, a published study also confirmed that administration
of rapamycin also reduced epileptic activity, cell death and axon
sprouting in organotypic hippocampal culture model of PTE (12).
Therefore, we performed this study to explore
whether there is a abnormal activation of mTOR signaling in a rat
model of PTE, and to determine the potential effect of rapamycin on
mTOR-mediated epilepsic injury, aiming to find a novel therapeutic
target for PTE management.
Materials and methods
Animals
Male Sprague-Dawley rats weighing between 180–220 g
were purchased from the SLAC Laboratory Animal Co., Ltd. (Shanghai,
China). The rats were housed in a 12 h of light and dark cycle, and
were allowed free access to water and food. All the procedures
performed were approved by the Animal Care and Use Committee of
Fujian Medical University and in accordance with the guidelines of
National Institutes of Health.
Establishment and assessment of PTE
model
Procedures to induce the rat model of PTE has been
described detailedly in previous studies (16,17).
Briefly, rats were anaesthetized with an intraperitoneal injection
of chloral hydrate (350 mg/kg) and placed in a stereotaxic frame.
Animals were injected with a 10 µl of 100 mM FeCl2 or an
equal volume of normal saline (NS) at a rate of 1 µl per minute
through a small burr hole with the following coordinates: 2.0 mm
anterior, 3 mm lateral to bregma, 2 mm depth. The sham-operated
group was inserted a neddle without receiving any solution. After
the operation, the rats were kept in single cage. An experienced
observer who was blinded to the experimental protocols graded the
scale of seizures using modified Racine criteria, in which seizure
stages were identified technically according to animal behaviors
(18). Grade evaluation began at 1–2
h after the surgery.
Experimental design and
pharmacological administration
In experiment 1, 129 rats were divided into three
groups: Sham group (n=10), NS group (n=52), and PTE group (n=67).
Brain tissues were collected at baseline, 1, 24 h, 1, 2 and 4
weeks, respectively. Each time point contained 10 rats. The
expression levels of p-mTOR and p-P70S6K were determined using
western blotting and immunohistochemistry. In experiment 2, 127
rats were randomly assigned to two groups: the PTE + vehicle group
(n=62) and the PTE + rapamycin group (n=65). For drug
administration, rapamycin was dissolved in solution containing 5%
Tween 80, 5% PEG 400 and 4% ethanol, and then administered
intraperitoneally at a dose of 6 mg/kg subsequently after induction
of PTE. The time and dose of rapamycin injection was based on
previous studis with slight modifications (19). The vehicle group received an equal
volume of solution. The potential role of rapamycin was estimated
also by western blotting and immunohistochemistry. In addition, the
frequency and number of behavioral seizures were also
evaluated.
Western blot analysis
Rats were decapitated separately at each time point
after operation. Perilesional cortex and ipsilateral hippocampus
were collected and processed for western blot assays as previously
described (20). Protein
concentrations were measured with a BCA kit (Beyotime, China).
Equal amounts of protein were run on 10% SDS-PAGE and then
transferred to polyvinylidene fluoride membranes at 100 V for 70
min. After blocking with 5% nonfat dry milk in PBS for 2 h at room
temperature, the membranes were incubated overnight at 4°C with
primary antibodies for rabbit anti-p-mTOR (1:200; Wuhan Boster
Biological Technology, Ltd., Wuhan, China) and mouse anti-p-P70S6K
(1:100; Wuhan Boster Biological Technology, Ltd.). The membranes
were washed with PBST and further incubated with horseradish
peroxidase (HRP) conjugated secondary antibodies at room
temperature for 1 h. Enhanced chemiluminescence reagent kit was
used to detected the band' reactivitiy according to the
manufacturer' protocol (Beyotime Institute of Biotechnology,
Haimen, China). Band densities were quantified with ImageJ software
(http://rsb.info.nih.gov/ij/). The
results are presented as the relative densities to β-actin.
Immunohistochemistry
Immunohistochemistry was carried out as previously
described (21). Briefly, rats were
anaesthetized with chloral hydrate and perfused transcardially with
saline followed by 4% paraformaldehyde in phosphate-buffered saline
(PBS; 0.l M, pH 7.4). After postfixation and dehydration, the
brains were embedded in paraffin and cut into 4 µm slices. The
sections were deparaffinized and rehydrated in graded
concentrations of ethanol to distilled water. and were further
received antigen retrieval. Sections were incubated with 3%
H2O2 in PBS for 10 min to block the
endogenous peroxidase activity. After blocking with 5% fetal bovine
serum in PBS, the sections were incubated with primary antibodies
overnight at 4°C. Next day, the sections were incubated with
HRP-conjugated second antibodies at room temperature for 1 h. After
that, diaminobenzidine and hematoxylin were used to stain
successively. Sections were observed under an Olympus microscope
and five non-overlapping fields (magnification, ×400) per section
were selected. Three sections per rat were used for quantification.
Immunoreactivity densities were analyzed with ImageJ software by an
investigator blinded to the treatment groups.
Statistical analysis
Data are presented as means ± SD. The statistical
analyses were performed using SPSS 13.0 (SPSS, Inc., Chicago, IL,
USA). Student's t-test was used for comparison between groups. For
behavioral data, Kruskal-Wallis test was applied. P<0.05 was
considered to indicate a statistically significant difference.
Results
General condition and epilepsy
evaluation
A total of 194 rats received intracortical injection
of FeCl2, and 150 of 176 (85.2%) developed spontaneous
behavioral seizures, including mouth and facial movement, head
nodding, stiff tail, forelimb contraction and full motor seizures.
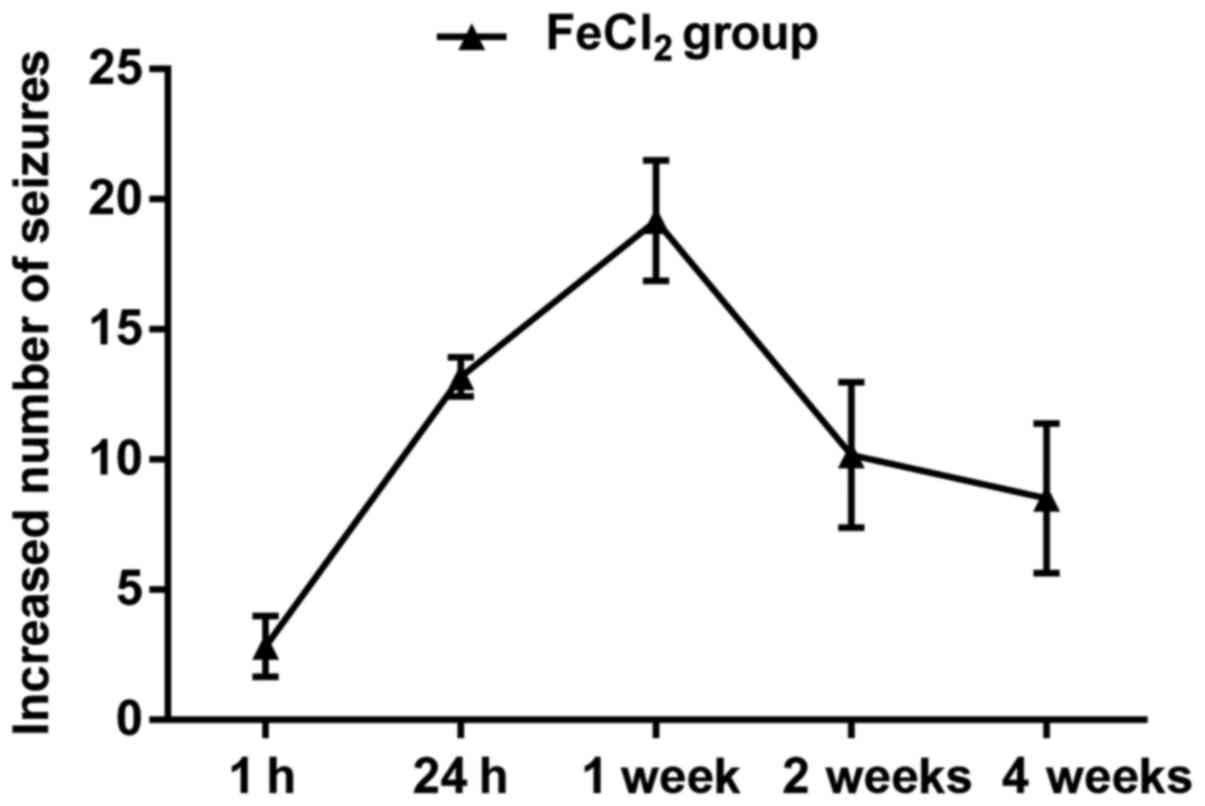
In general, the frequency and number of behavioral seizures
increased beginning at 1 h and peaked at 3–4 h. About 24 h later,
generalized seizures almost dispeared, however, focal seizures
could still be seen in most of the FeCl2-injected rats.
Over the next four weeks, epileptic duration and frequency
decreased continuously, in which rats primarily performed
shivering, head nodding, facial movements and few limb clonus
(Fig. 1). Additionally, 18 rats
subjected to FeCl2 injection died during the
observation, of which 10 rats died for status epilepticus, 8 rats
for high seizures frequency. The remaining 26 rats failed to
develop behavioral seizures and were excluded from final analyses.
For animals injected with NS, only three rats developed focal
seizures including head nodding and facial movements, but none of
these rats died (data not shown). As expected, none of rats died or
showed abnormal performance in the sham group.
Cortical and hippocampal injury in
rats subjected to FeCl2 injection
As shown in Fig. 2A,
granule cells, molecular and polymorphic layer of the dentate gyrus
in the sham group showed normal arrangements and distributions.
Moreover, there was also no neuronal degeneration or cell loss in
frontal cortex (Fig. 2D). Slight
cellular changes in frontal lobe and hippocampus were found in the
NS-injected rats including neuronal degeneration, nuclear pyknosis
and granule cells proliferation (Fig. 2B
and E). However, more obvious cellular changes in perilesional
cortex and hippocampus were confirmed in the
FeCl2-injected rats than that in the NS-injected rats,
such as plenty of neuronal loss, swelling, degeneration and death,
as well as inflammatory cell infiltration and glial cell
proliferation (Fig. 2C and F).
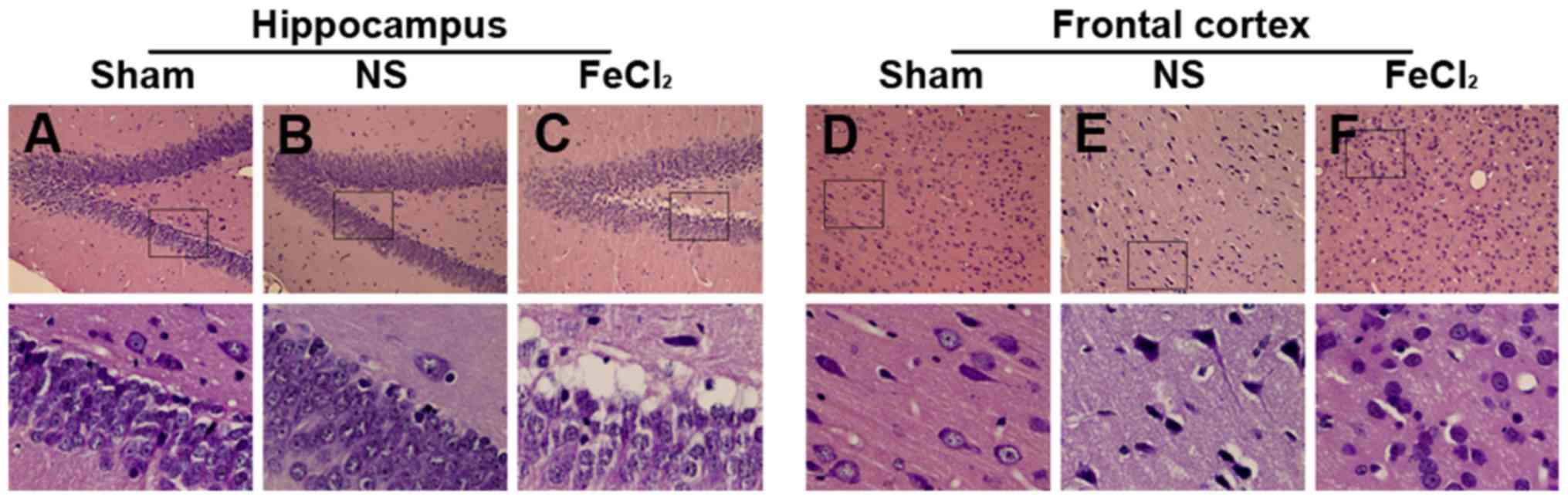 | Figure 2.Hematoxylin and eosin of hippocampus
and frontal cortex. (A) Cells in dentate gyrus show normal
arrangements and distributions in the sham group. (B) Slight
hippocampal changes in the NS-injected rats including neuronal
degeneration, nuclear pyknosis and granule cells proliferation. (C)
Hippocampal changes in the FeCl2-injected rats including
plenty of neuronal loss, swelling, degeneration and death, as well
as inflammatory cell infiltration and glial cell proliferation. (D)
Cells in frontal lobe show normal arrangements and distributions in
the sham group. (E) Slight cellular changes in frontal lobe of the
NS-injected rats including neuronal degeneration, nuclear pyknosis
and granule cells proliferation. (F) Cellular changes in frontal
lobe of the FeCl2-injected rats including plenty of
neuronal loss, swelling, degeneration and death, as well as
inflammatory cell infiltration and glial cell proliferation. Bottom
graphs are enlarged view of the boxs above (magnification, ×400),
respectively. NS, normal saline. |
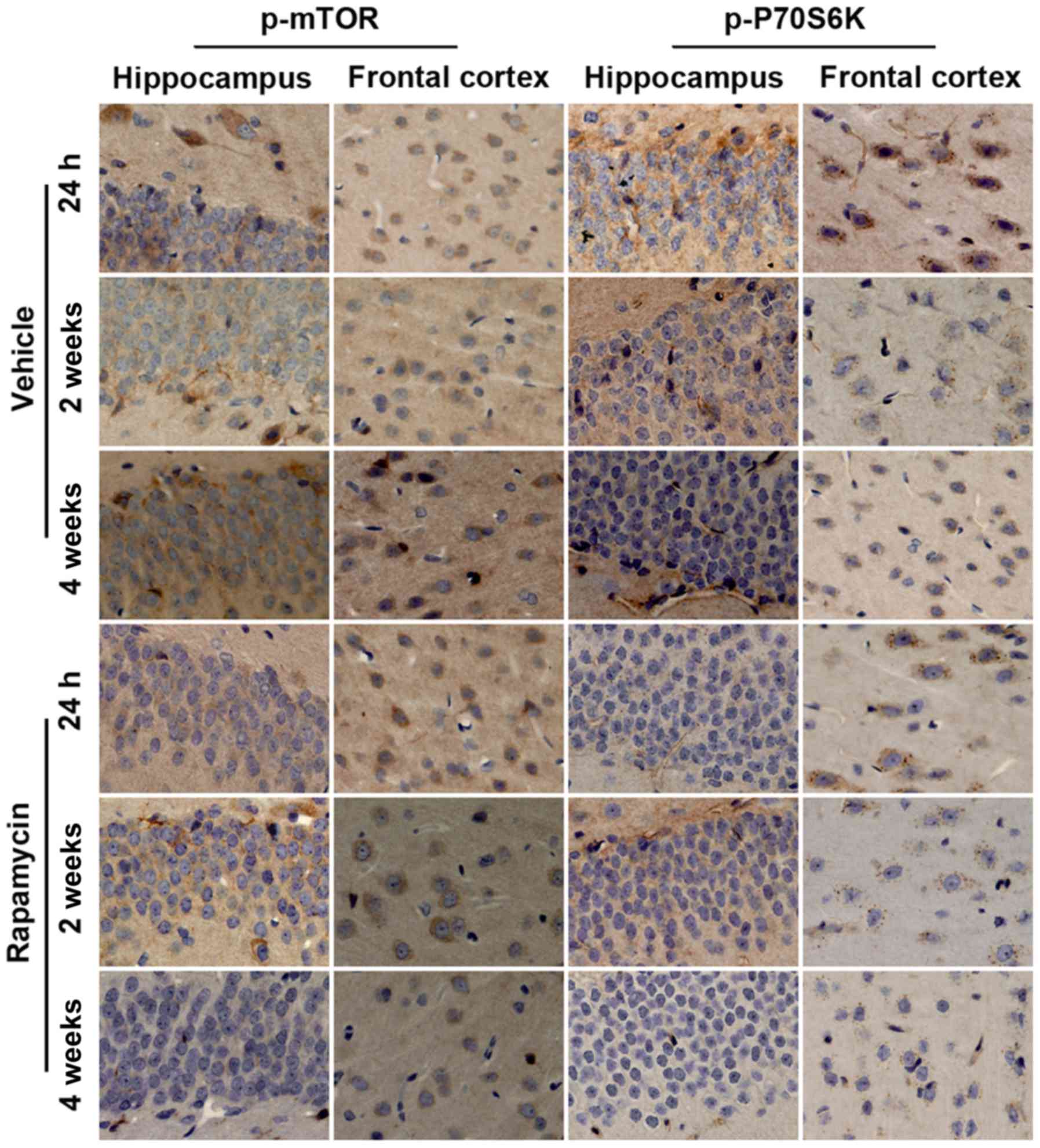
Increased expression of p-mTOR and
p-P70S6K in rats after intracortical injection of
FeCl2
To examine whether mTOR signaling was upregulated in
rats after PTE induction, we first carried out a temporal course of
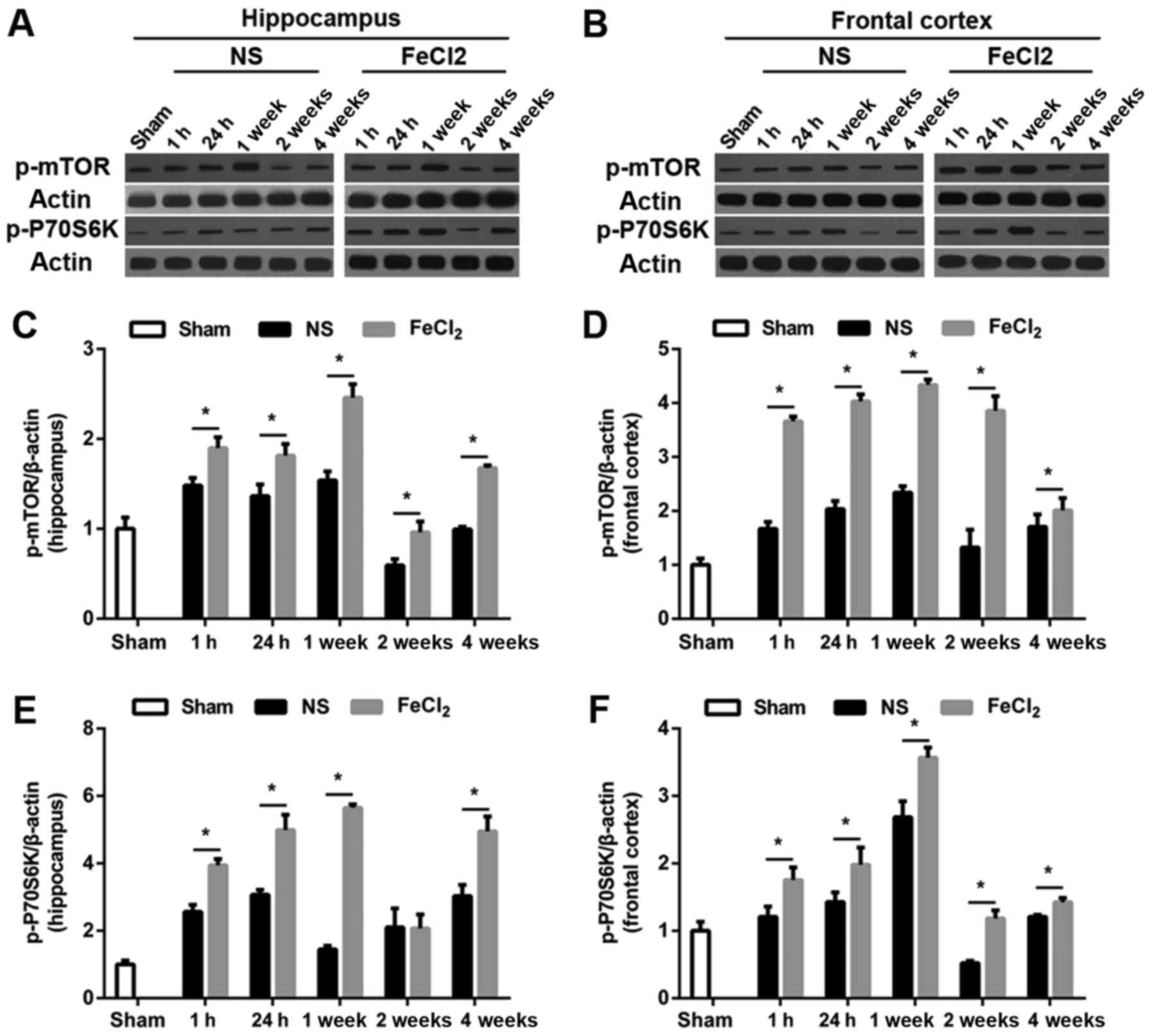
experiment with western blotting. As visible in Fig. 3A and B, at baseline, there was a
small amount of p-mTOR and p-P70S6K expression in frontal lobe and
hippocampus. Of note, both proteins increased significantly at 1 h
after the surgery (FeCl2 or NS injection) and peaked at
1 week in the abovementioned regions. Then protein levels of p-mTOR
and p-P70S6K decreased to the bottom at 2 weeks (except p-mTOR
expression in frontal cortex) while elevated again at 4 weeks
(Fig. 3C-F). Furthermore, compared
with rats injected with NS, perilesional and hippocampal p-mTOR
levels in rats after PTE induction were significantly increased at
all time points. Similarly, in addition to p-P70S6K expression in
hippocampus at 2 week after surgery, p-P70S6K levels at each time
point were higher than those in the NS-injected rats. Consistence
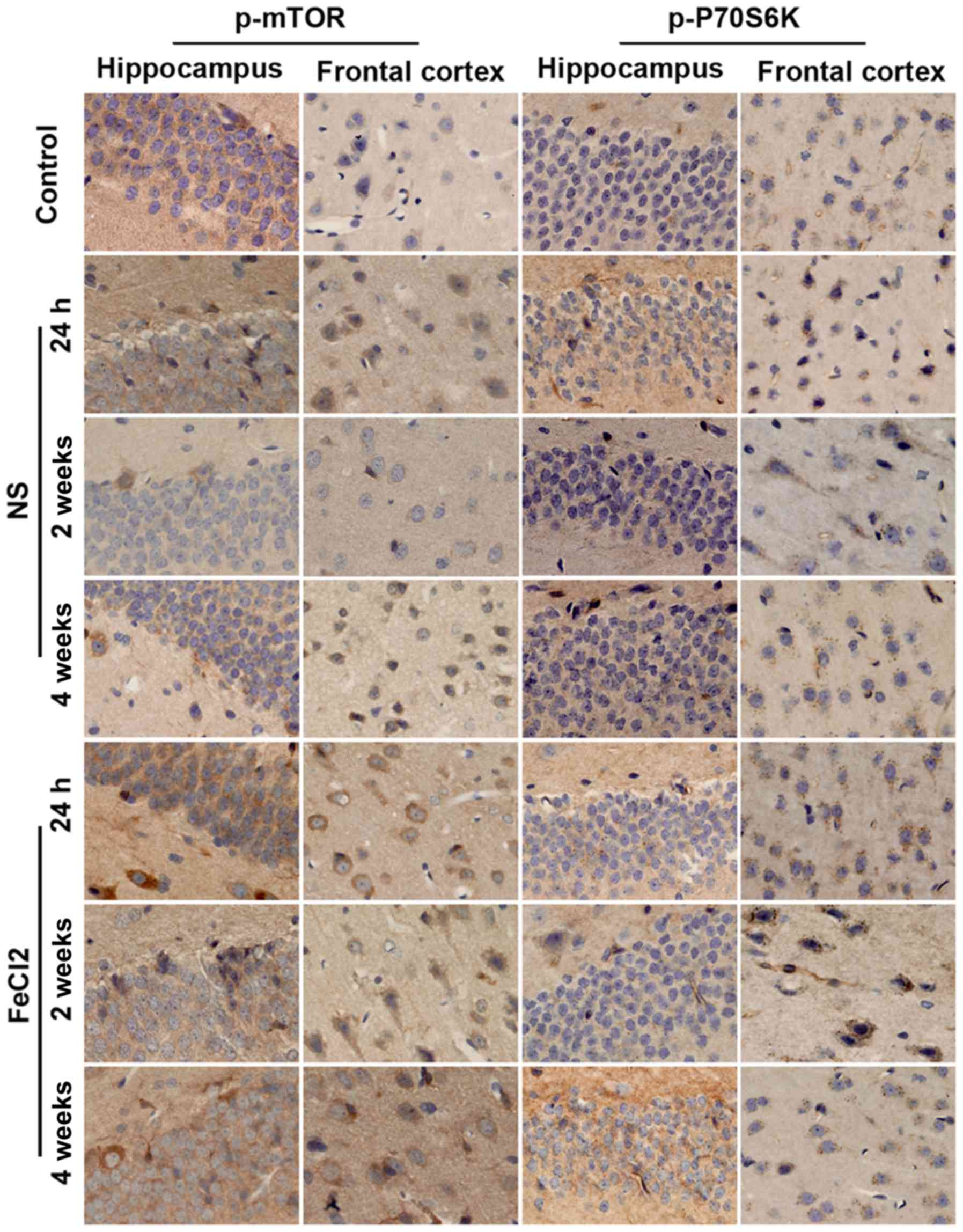
with the results of western blotting, immunohistochemistry further
demonstrated the higher increased levels of p-mTOR and p-P70S6K in
frontal lobe and hippocampus following PTE (Fig. 4). Moreover, both proteins primarily
expressed in cytoplasm.
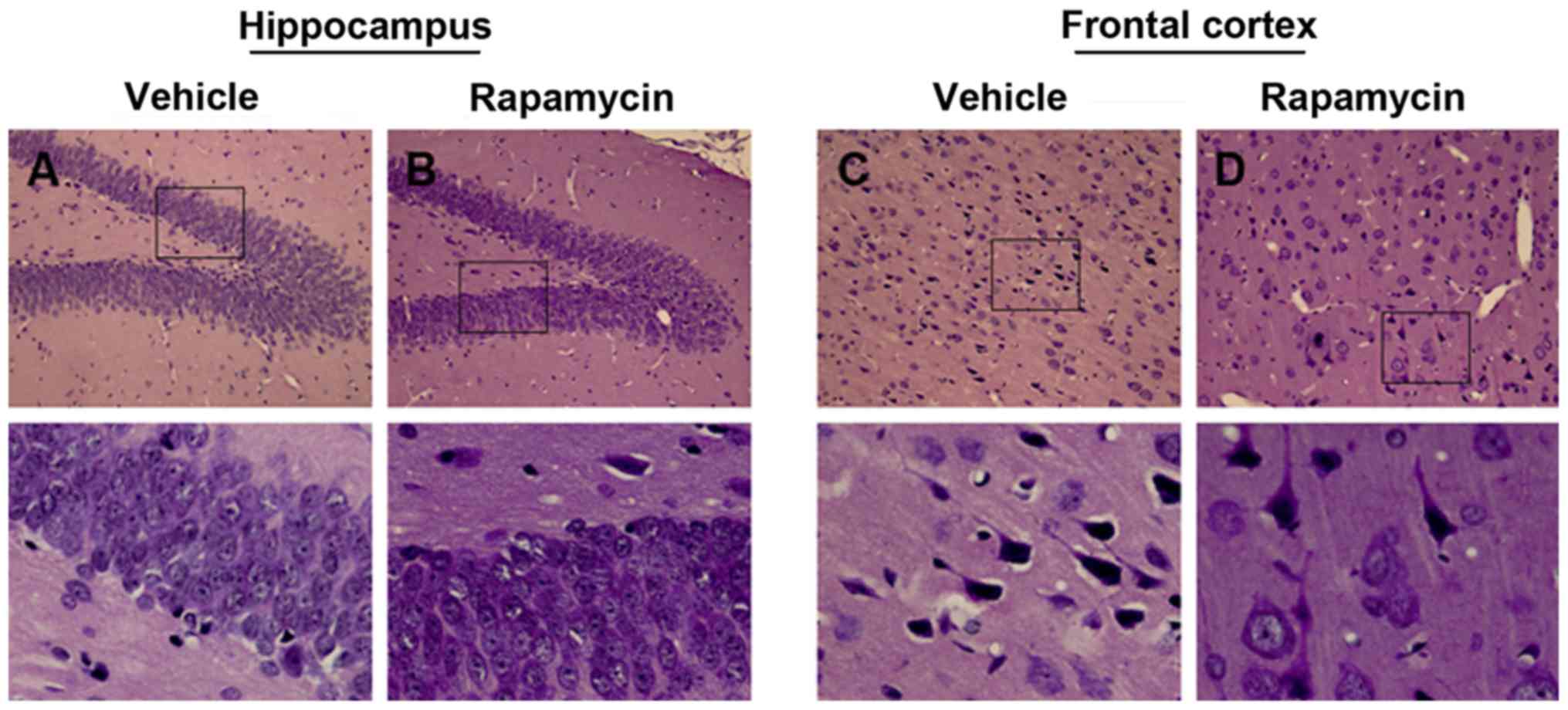
Rapamycin reduced behavioral seizures
and attenuated epileptic injury
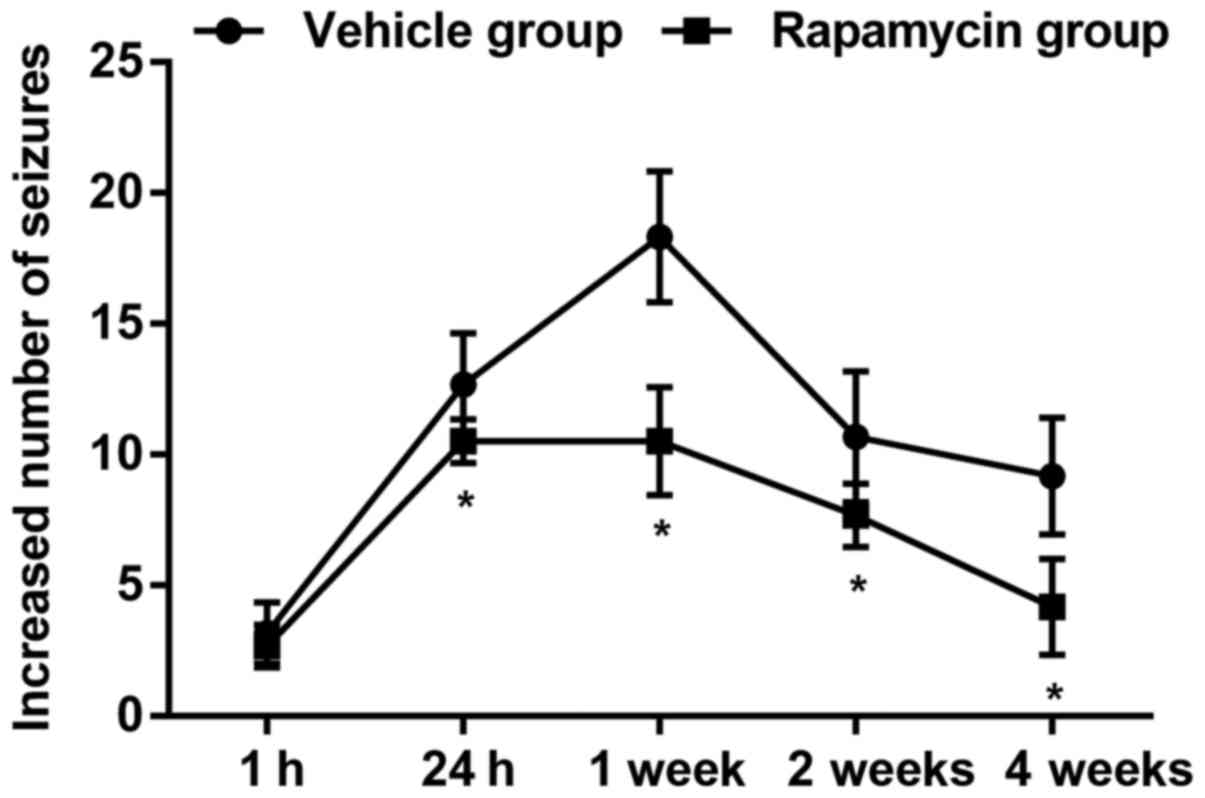
Similar pattern of seizures was found in the vehicle
group as compared with the PTE + rapamycin group, but actually, the
PTE rats exerted significant decline of behavioral seizures ranging
from 24 h to 4 weeks after treatment with rapamycin (Fig. 5; P<0.05). Additionally, there
still existed slight disruptions of cellular arrangement and
structure in the PTE + vehicle group, while which were also
alleviated greatly both in frontal lobe and hippocampus after
rapamycin treatment (Fig. 6A-D).
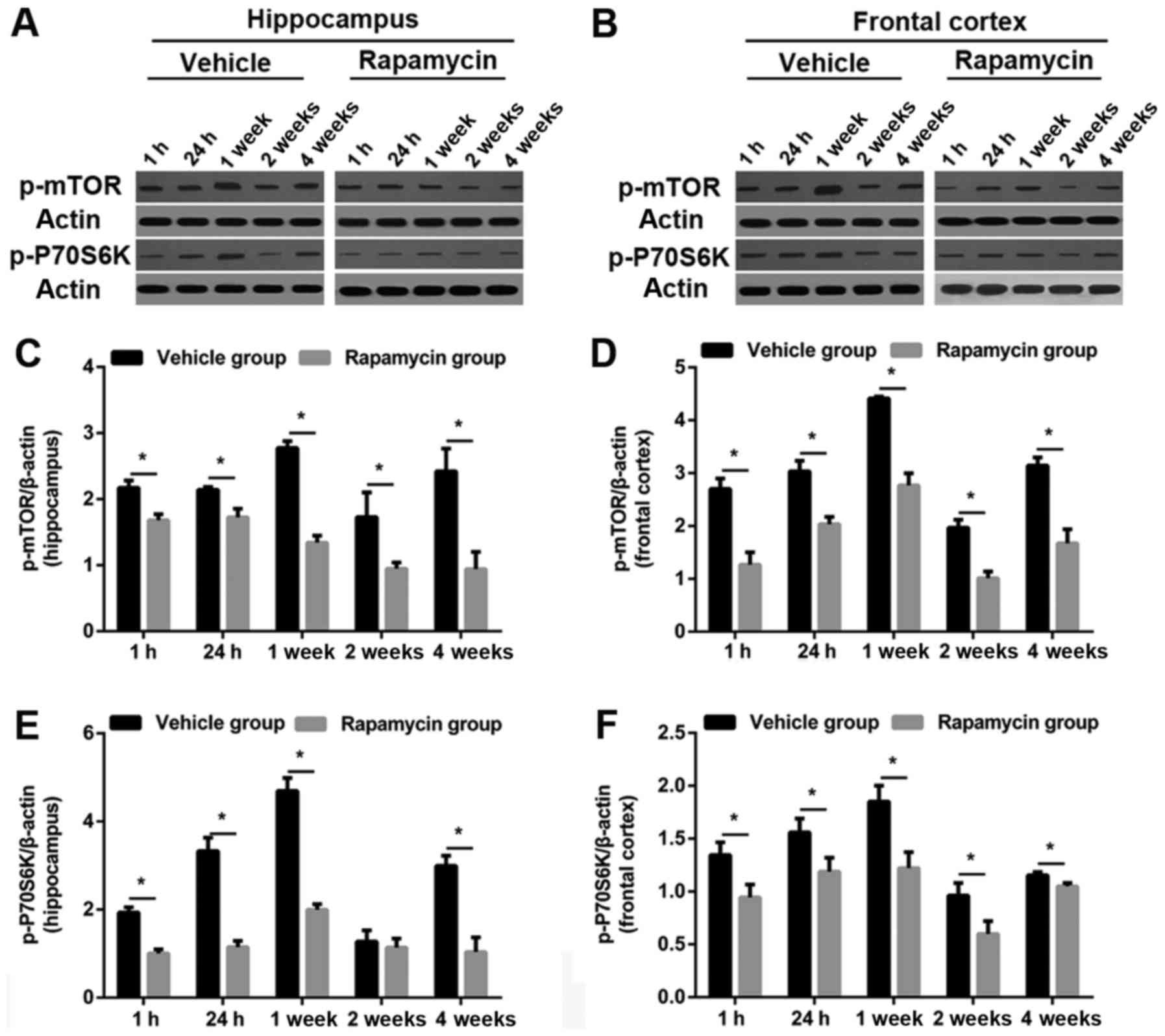
Rapamycin decreased the level of
p-mTOR and p-P70S6K in rats after PTE induction
As predicted, the levels of p-mTOR and p-P70S6K were
obviously increased in frontal lobe and hippocampus in the vehicle
group. Both proteins were significantly reduced in the PTE +
rapamycin group as compared to the PTE + vehicle group (except
hippocampal p-P70S6K level at 2 weeks). Specifically, perilesional
and hippocampal p-mTOR levels decreased significantly at all time
points after rapamycin treatment. Similar changes were found in the
levels of perilesional p-P70S6K. Moreover, hippocampal p-P70S6K
levels at 1 and 24 h, as well as 1 and 4 weeks were also decreased
significantly after rapamycin treatment in addition to that at 2
weeks (Figs. 7 and 8).
Discussion
The main finding of this study was that the levels
of p-mTOR and p-P70S6K, biomarkers of activated mTOR signaling,
increased significantly in rats subjected to PTE induction, which
was accompanied by obvious behavioral seizures and cellular injury.
Of interest, both elevated indicators were decreased remarkably
after rapamycin treatment. Moreover, inhibition of mTOR with
rapamycin also greatly reduced behavioral seizures over a longer
time-period as compared to previously studies (12,19). Our
results suggest that the over activation of mTOR signaling may be a
crucial pathogenic mechanism participated in the development of
FeCl2-induced PTE. Rapamycin provides a
antiepileptogenic effect through the inhibition of mTOR
signaling.
At present, there are several animal models of PTE,
such as controlled cortical impact model (19,22), low
Mg2+ model (23),
cortical undercut model (24), and
FeCl2-induced model (16). In the present study, we adopted the
frontal cortex injection of FeCl2-induced PTE model for
the following reasons: i) Frontal cortex damage is the most common
type of injuries in clinical practice and the pathological change
caused by FeCl2 injection is similar to human TBI
(17); ii) The model has high
success rate, stability and repeatability; and iii) The latency of
seizures is short and beneficial to behavioral evaluation and
pharmacological intervention. Most of the reasons were further
verified in our study. For example, 85.2% rats developed to
epileptogenesis after injection of FeCl2. Moreover,
behavioral seizures could be induced within as early as an hour and
sustained at least 4 weeks after FeCl2 injection.
The mTOR pathway has been reported to involve in
several neurological diseases including depressive disorder
(25), ischemia brain injury
(11,26), brain tumor (27) and epilepsy (28). Here, we provide evidence that the
expression of p-mTOR and p-P70S6K, over activation biomarkers of
mTOR signaling, strengthened heavily in hippocampus and frontal
cortex after PTE induction. Consistence with our results, a study
in vitro also verified the upregulation of mTOR signaling
after the induction of PTE despite of without investigating the
downstream proteins. In addition, literature concerning other type
of epilepspy also determined the hyperactivation of mTOR and
further found the increased expression of downstream proteins, such
as p-P70S6K (28). PTE is a common
sequela following craniocerebral injury. However, there is limited
effectivity for its treatment. In recent years, rapamycin has been
widely applied in experimental researches and neuroprotective
effects are found in a number of diseases including TBI (29), hypoxic-ischemic encephalopathy
(26), and epilepsy (28,30). In
line with the results of these studies, our results indicate that
the increased levels of p-mTOR and p-P70S6K were reduced
significantly after rapamycin treatment. That is, the over
activation of mTOR pathway was markedly inhibited after
administration of rapamycin. Meanwhile, the frequency and number of
behavioral seizures and epileptic injury were also greatly
weakened.
Multiple potential mechanisms may contribute to the
antiepileptogenic effects of rapamycin. First, hyperactivation of
mTOR/P70S6K signaling promotes oligodendrocyte proliferation and
myelination (25), which is
essential to the axonal sprouting susceptible to the development of
epilepsy (2). Therefore,
antiepileptogenic effect was obtained via blocking the mTOR
signaling and decreasing the expression of downstream p-P70S6K.
Second, induction of synaptic plasticity, resulting from the
elevated proteins synthesis in hippocampus following activation of
mTOR signaling is a important mechanism for epileptogenesis
(31). Rapamycin weaken the
frequency and number of seizures through decreasing the levels of
certain translation initiation factors and reducing the rate of
protein synthesis. Third, as previously reported (32), rapamycin reduce the activation of
microglia, a hallmak of neuroinflammation, which can be attibuted
to its intrinsic anti-inflammatory effect. Last but no least,
although the role of mTOR signaling on neuronal apoptosis is
controversial, several studies have demonstrated that inhibition of
seizure-induced mTOR signaling with rapamycin could reduce the
amount of apoptosis (33).
Meanwhile, we need to notice the limitations of our
study. First, electrophysiological examination is still the golden
standard for epilepsy diagnosis and is crucial for the response
evaluation. So it should be included in the future study. Second,
the distribution of p-mTOR and p-P70S6K have not been illustrated
in this study. Further tests would be conducted to identify the
specific cell types that express the abovementioned biomarkers of
activated mTOR signaling.
In summary, our results suggest that there exists
over activation of mTOR signaling in a rat model of FeCl2-induced
PTE. Inhibition of mTOR signaling with rapamycin significantly
reduces the behavioral seizures and then ameliorates epileptic
brain injury. It suggests that mTOR signaling pathway may be a
potential therapeutic target, and rapamycin can be considered as a
promising agent for PTE treatment.
Acknowledgements
This study was sponsored by the Natural Science
Foundation of Fujian Province (no. 2015J01387), and the Medical and
Health Science and Technology Innovation Program of Fujian Province
(no. 2014-CXB-14), and the Key Clinical Specialty Discipline
Construction program of Fujian, China (no. 2012-SLCZD-2).
Glossary
Abbreviations
Abbreviations:
|
mTOR
|
mammalian target of rapamycin
|
|
PTE
|
post-traumatic epilepsy
|
|
TBI
|
traumatic brain injury
|
|
AEDs
|
anti-epileptic drugs
|
|
NS
|
normal saline
|
|
HRP
|
horseradish peroxidase
|
|
PBS
|
phosphate buffer
|
References
|
1
|
Gupta PK, Sayed N, Ding K, Agostini MA,
Van Ness PC, Yablon S, Madden C, Mickey B, D'Ambrosio R and
Diaz-Arrastia R: Subtypes of post-traumatic epilepsy: Clinical,
electrophysiological, and imaging features. J Neurotrauma.
31:1439–1443. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bhuyan P, Patel DC, Wilcox KS and Patel M:
Oxidative stress in murine Theiler's virus-induced temporal lobe
epilepsy. Exp Neurol. 271:329–334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Irimia A and Van Horn JD: Epileptogenic
focus localization in treatment-resistant post-traumatic epilepsy.
J Clin Neurosci. 22:627–631. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Christensen J, Pedersen MG, Pedersen CB,
Sidenius P, Olsen J and Vestergaard M: Long-term risk of epilepsy
after traumatic brain injury in children and young adults: A
population-based cohort study. Lancet. 373:1105–1110. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong M: Mammalian target of rapamycin
(mTOR) pathways in neurological diseases. Biomed J. 36:40–50. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cho CH: Frontier of epilepsy research-mTOR
signaling pathway. Exp Mol Med. 43:231–274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Crino PB: The mTOR signalling cascade:
Paving new roads to cure neurological disease. Nat Rev Neurol.
12:379–392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nguyen LH, Brewster AL, Clark ME,
Regnier-Golanov A, Sunnen CN, Patil VV, D'Arcangelo G and Anderson
AE: mTOR inhibition suppresses established epilepsy in a mouse
model of cortical dysplasia. Epilepsia. 56:636–646. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sha LZ, Xing XL, Zhang D, Yao Y, Dou WC,
Jin LR, Wu LW and Xu Q: Mapping the spatio-temporal pattern of the
mammalian target of rapamycin (mTOR) activation in temporal lobe
epilepsy. PLoS One. 7:e391522012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu P, Yang X, Hei C, Meli Y, Niu J, Sun T
and Li PA: Rapamycin reduced ischemic brain damage in diabetic
animals is associated with suppressions of mTOR and ERK1/2
Signaling. Int J Biol Sci. 12:1032–1040. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Berdichevsky Y, Dryer AM, Saponjian Y,
Mahoney MM, Pimentel CA, Lucini CA, Usenovic M and Staley KJ:
PI3K-Akt signaling activates mTOR-mediated epileptogenesis in
organotypic hippocampal culture model of post-traumatic epilepsy. J
Neurosci. 33:9056–9067. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sancho-Rieger J and Parra-Martinez J:
Preventive and therapeutic attitude in post-traumatic epileptic
seizures. Rev Neurol. 35 Suppl 1:S39–S42. 2002.(In Spanish).
PubMed/NCBI
|
|
14
|
Talos DM, Sun H, Zhou X, Fitzgerald EC,
Jackson MC, Klein PM, Lan VJ, Joseph A and Jensen FE: The
interaction between early life epilepsy and autistic-like
behavioral consequences: A role for the mammalian target of
rapamycin (mTOR) pathway. PLoS One. 7:e358852012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galanopoulou AS, Gorter JA and Cepeda C:
Finding a better drug for epilepsy: The mTOR pathway as an
antiepileptogenic target. Epilepsia. 53:1119–1130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Willmore LJ and Triggs WJ: Effect of
phenytoin and corticosteroids on seizures and lipid peroxidation in
experimental posttraumatic epilepsy. J Neurosurg. 60:467–472. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ueda Y, Willmore J and Triggs WJ:
Amygdalar injection of FeCl3 causes spontaneous recurrent seizures.
Exp Neurol. 153:123–127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Racine RJ: Modification of seizure
activity by electrical stimulation. II. Motor seizure.
Electroencephalogr Clin Neurophysiol. 32:281–294. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Butler CR, Boychuk JA and Smith BN:
Effects of rapamycin treatment on neurogenesis and synaptic
reorganization in the dentate gyrus after controlled cortical
impact injury in mice. Front Syst Neurosci. 9:1632015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rankin-Gee EK, McRae PA, Baranov E, Rogers
S, Wandrey L and Porter BE: Perineuronal net degradation in
epilepsy. Epilepsia. 56:1124–1133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou C, Xie G, Wang C, Zhang Z, Chen Q,
Zhang L, Wu L, Wei Y, Ding H, Hang C, et al: Decreased progranulin
levels in patients and rats with subarachnoid hemorrhage: A
potential role in inhibiting inflammation by suppressing neutrophil
recruitment. J Neuroinflammation. 12:2002015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kelly KM, Miller ER, Lepsveridze E,
Kharlamov EA and Mchedlishvili Z: Posttraumatic seizures and
epilepsy in adult rats after controlled cortical impact. Epilepsy
Res. 117:104–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gong XW, Li JB, Lu QC, Liang PJ and Zhang
PM: Effective connectivity of hippocampal neural network and its
alteration in Mg2+-free epilepsy model. PLoS One. 9:e929612014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chauvette S, Soltani S, Seigneur J and
Timofeev I: In vivo models of cortical acquired epilepsy. J
Neurosci Methods. 260:185–201. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lloyd BA, Hake HS, Ishiwata T, Farmer CE,
Loetz EC, Fleshner M, Bland ST and Greenwood BN: Exercise increases
mTOR signaling in brain regions involved in cognition and emotional
behavior. Behav Brain Res. 323:56–67. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee HJ, Koh SH, Song KM, Seol IJ and Park
HK: The Akt/mTOR/p70S6K pathway is involved in the neuroprotective
effect of erythropoietin on hypoxic/ischemic brain injury in a
neonatal rat model. Neonatology. 110:93–100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fan W, Wang W, Mao X, Chu S, Feng J, Xiao
D, Zhou J and Fan S: Elevated levels of p-Mnk1, p-eIF4E and
p-p70S6K proteins are associated with tumor recurrence and poor
prognosis in astrocytomas. J Neurooncol. 131:485–493. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lippman-Bell JJ, Rakhade SN, Klein PM,
Obeid M, Jackson MC, Joseph A and Jensen FE: AMPA receptor
antagonist NBQX attenuates later-life epileptic seizures and
autistic-like social deficits following neonatal seizures.
Epilepsia. 54:1922–1932. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding K, Wang H, Wu Y, Zhang L, Xu J, Li T,
Ding Y, Zhu L and He J: Rapamycin protects against apoptotic
neuronal death and improves neurologic function after traumatic
brain injury in mice via modulation of the mTOR-p53-Bax axis. J
Surg Res. 194:239–247. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Buckmaster PS and Wen X: Rapamycin
suppresses axon sprouting by somatostatin interneurons in a mouse
model of temporal lobe epilepsy. Epilepsia. 52:2057–2064. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stoica L, Zhu PJ, Huang W, Zhou H, Kozma
SC and Costa-Mattioli M: Selective pharmacogenetic inhibition of
mammalian target of Rapamycin complex I (mTORC1) blocks long-term
synaptic plasticity and memory storage. Proc Natl Acad Sci USA.
108:3791–3796. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van Vliet EA, Otte WM, Wadman WJ, Aronica
E, Kooij G, de Vries HE, Dijkhuizen RM and Gorter JA: Blood-brain
barrier leakage after status epilepticus in rapamycin-treated rats
II: Potential mechanisms. Epilepsia. 57:70–78. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zeng LH, Rensing NR and Wong M: The
mammalian target of rapamycin signaling pathway mediates
epileptogenesis in a model of temporal lobe epilepsy. J Neurosci.
29:6964–6972. 2009. View Article : Google Scholar : PubMed/NCBI
|