Introduction
Gastric cancer (GC) is one of the most common types
of cancer and the second most common cause of cancer-associated
mortalities worldwide (1). In
certain cases, recurrence and metastasis may occur despite
comprehensive surgical and medical treatment (2). Although previous studies of GC
tumorigenesis demonstrated close associations between
overexpression of oncogenes/oncoproteins and inactivation of
anti-oncogenes/anti-oncoproteins (3), the underlying mechanisms remain to be
investigated. Exploration of molecular mechanisms, prognosis and
rational clinical/medical treatments are important for the
discovery of new genes involved in GC development and behavior and
for improvements in treatment.
Members of the cadherin superfamily of cell adhesion
factors are expressed as cell surface glycoproteins and regulate
calcium-mediated cell adhesion, influence cell polarity and
morphogenesis and direct cell recognition and signal transduction
mechanisms (4). Classical cadherins,
including E- and N-cadherin, comprise an extracellular
calcium-binding domain and a transmembrane domain (5). By contrast, the non-classical truncated
(T)-cadherin lacks the transmembrane domain and binds cytomembranes
via glycosyl-phosphatidyl inositol (GPI) (6). Previous studies have reported
associations between deletion or mutation of classical cadherins
and proliferation, migration and invasion of GC, breast cancer and
lung cancer cells (7–10). Tryndyak et al (11) observed that transfection of tumor
cells with E-cadherin decreased proliferation and invasiveness
significantly. Ivanov et al (12) revealed that T-cadherin upregulation
correlates with cell cycle progression and promotes proliferation
of vascular cells. Notably, T-cadherin downregulation was observed
in GC (13), breast cancer (14), lung cancer (15), colon cancer (16), skin squamous carcinoma (17) and other types of cancer (18), suggesting a potential role as an
anti-oncoprotein.
Defects, including aberrant promoter methylation and
improper histone modification of CDH13, which encodes
T-cadherin, may contribute to downregulation of protein expression
(16,19). A study on non-small cell lung cancer
cell lines and tumor tissues revealed that T-cadherin deletion
increased tumorigenicity (15).
Furthermore, a defect in CDH13 was demonstrated to promote
tumor progression in human prostate cancer cells, whereas
restoration of T-cadherin expression inhibited both cell
proliferation and invasion (20). In
neuroblastoma, transduction with CDH13 was revealed to
inhibit tumor growth by reducing endothelial growth factor receptor
expression (21). An in vitro
study by Lee (22) demonstrated that
transduction of CDH13 cDNA into breast cancer cells reduced
growth and invasiveness of tumor cells. Furthermore, tumor volumes
observed in mice implanted with T-cadherin-overexpressing MCF-7
human breast cancer cells were significantly reduced, suggesting
that T-cadherin expression inhibits tumorigenesis in vivo
(22,23). In a previous study, it was
demonstrated that mRNA levels and T-cadherin protein expression
were significantly downregulated in GC tissues compared with
adjacent noncancerous tissues, suggesting that T-cadherin may be
important in GC cell proliferation and metastasis and serve as a
target for treatment of GC (24).
The current study aimed to investigate functions and
underlying mechanisms of T-cadherin and to provide a basis for
usage of this protein in clinical diagnosis and treatment of GC. A
5-year follow-up study of survival among patients with GC was
conducted to determine the association between T-cadherin
expression and GC prognosis. A T-cadherin-overexpressing cell line
was generated from HGC-27 cells and used to investigate
associations between T-cadherin expression and GC cell
proliferation, invasiveness and metastasis.
Materials and methods
Patients
Eighty-one patients with Stage I–III GC who
underwent surgical treatment at the Department of Surgical
Oncology, Second Affiliated Hospital of Fujian Medical University
(Quanzhou, China) between August 2011 and August 2015, were
followed for 2–60 months. Overall survival was estimated using the
Kaplan-Meier method, as described in a previous report (25). T-cadherin-negative disease was
defined as the tissue section exhibiting ≤10% or no positive cancer
cells. Patients included 49 males and 32 females with a mean age of
62.5±18.6 years (range, 26–76 years). All experiments were
performed in accordance with the relevant guidelines and written
informed consent was obtained from all patients. Study protocols
were approved by the Ethical Committee of the Second Affiliated
Hospital of Fujian Medical University.
HGC-27 cell culture and
transfection
Human gastric carcinoma cell line HGC-27 was
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China). Cells were cultured in RPMI-1640
medium (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), penicillin (100 U/ml) and streptomycin (100
µg/ml) and incubated in a 5% CO2 incubator at 37°C.
Plasmids and cell transfection
The plasmids for pcDNA3.1 (400 ng/µl) and
pcDNA-T-cadherin (400 ng/µl; pcDNA-Tcad; Invitrogen; Thermo Fisher
Scientific, Inc.) were purchased and respectively transfected into
HGC-27 cell lines using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Cell transfection efficiency was determined as the
percentage of fluorescent cells among total cells in ten regions,
using a 200-fold phase contrast fluorescence microscope. Two weeks
following transfection, positive colonies (with GFP expression)
were obtained and cells were resuspended in RPMI-1640 medium and
cultured to yield T-cadherin overexpression cells. Cells
transfected with pcDNA3.1 (400 ng/µl; empty vector) were used as
negative controls. Untransfected HGC-27 cells were used as blank
controls.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.)
was used to extract total RNA from transfected HGC-27 cells
following 24 h incubation according to the manufacturer's protocol.
RNA was quantified spectrophotometrically as ratio of absorbance at
260 over 280 nm. cDNA was synthesized from total RNA using the
TIANScriptII cDNA first chain synthesis kit (Tiangen Biotech, Co.,
Ltd., Beijing, China) according to the manufacturer's protocol.
Primers for qPCR are listed in Table
I. RT-qPCR was performed using a PCR-iQ5 detection system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) with a SYBR-Green
PCR Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.;
4309155). The thermocycling conditions were as follows: 1 min at
95°C, followed by 40 cycles of 95°C for 5 sec and 60°C for 20 sec.
Gene expression was normalized to the expression of β-actin using
the 2−ΔΔCq method (26).
Each experiment was performed in triplicate.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Name | Sequence
(5′-3′) |
|---|
|
T-cadherin_Forward |
GATGTTGGCAAGGTAGTCGAT |
|
T-cadherin_Reverse |
GCTCCCTGTGTTCTCATTGAT |
|
β-actin_Forward |
GACGATATCGCTGCGCTG |
|
β-actin_Reverse |
GTACGACCAGAGGCATACAGG |
Western blotting
Proteins were extracted from tissue samples
according to the manufacturer's protocol in HGC-27 cells using the
Total Protein Extraction kit (BestBio, Co., Shanghai, China) and
were separated on 10% SDS-PAGE gels and transferred to
polyvinylidene difluoride membranes, which were subsequently
blocked with 5% skimmed milk for 1 h at 37°C. Membranes were
incubated overnight at 4°C with rabbit antibodies specific for
phosphorylated (p)-S6K (1:1,000; ab32529; Abcam), β-actin (1:5,000;
ab6276; Abcam), anti-p-protein kinase B (AKT; 1:1,000; 9271; Cell
Signaling Technology, Inc.), anti-AKT (1:1,000; 9272; Cell
Signaling Technology, Inc.), anti-mammalian target of rapamycin
(mTOR; 1:1,000; 2972; Cell Signaling Technology, Inc.) or
anti-p-mTOR (1:1,000; 5536; Cell Signaling Technology, Inc.).
Following washing, membranes were incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies
(1:3,000; ab6721; Abcam). Membranes were washed, incubated for 2 h
at room temperature with an enhanced chemiluminescence substrate
(Abcam) and analyzed. To quantify, signal intensities of specific
bands were measured using Image Lab 3.0 software (Bio-Rad
Laboratories, Inc.).
Flow cytometry analysis of cell
cycle
HGC-27 cells transfected with pcDNA3.1-Tadherin or
pcDNA3.1 were harvested following 24 h transfection by
trypsinization, fixed and permeabilized at 4°C for 30 minusing
Cytofix/Cytoperm™ Fixation/Permeabilization Solution kit (BD
Biosciences, San Jose, CA, USA) and stored at 4°C. At the time of
analysis, cells were stained with propidium iodide (Sigma-Aldrich,
Merck KGaA) at room temperature for 15 min and analyzed on a flow
cytometer (Aria II, BD Biosciences). Data were analyzed using
FlowJo Software version 7.6.1 (FlowJo LLC, Ashland, OR, USA).
Invasion and migration assay
pcDNA3.1-Tadherin- and pcDNA3.1-transfected HGC-27
cells were harvested in the logarithmic growth phase following 24 h
transfection and resuspended at a density of 5×10 cells/ml in
RPMI-1640 serum-free medium. Matrigel matrix (BD Biosciences) was
used to coat 24-well invasion chambers and rehydrated with
serum-free culture medium for 30 min at 37°C prior to adding 200 µl
of cell suspension to upper chambers and 600 µl of RPMI-1640 medium
with 10% FBS to lower chambers. Plates were incubated for 22 h at
37°C. Following discarding of excess matrix and non-invading cells,
cells that had invaded the Matrigel were fixed with 95% ethanol and
stained with crystal violet at room temperature for 10 min
according to the manufacturer's instructions (BD Biosciences).
Invaded and migrated cells were observed using a microscope
(magnification, ×10). Cell migration assays were performed using
the same procedure in the absence of Matrigel coating.
Cell proliferation assay
A suspension of HGC-27 cells transfected with
pcDNA3.1-Tadherin and pcDNA3.1 (200 µl; density,
1×104/ml) were added to each well of a 96-well plate.
Following a 24-h incubation at 37°C, cell viability was quantified
every day over 5 days using an MTT assay (American Type Culture
Collection, Manassas, VA, USA). MTT reagent (10 µl) was added to
each well and cells were incubated at room temperature in the dark
for 2 h. Detergent reagent (100 µl, American Type Culture
Collection) was added to each well and following an additional 2-h
incubation at room temperature, absorbance at 570 nm was measured
using a microplate reader. Insulin-like growth factor-1 (IGF-1) was
added to the RPMI-1640 medium. The growth inhibition rates were
defined as the OD negative value-OD positive value/OD value
negative.
Statistical analysis
All data are presented as the mean ± standard
deviations of at least five independent repeats. Survival rates of
patients with GC were estimated using the Kaplan-Meier method and
differences in survival were compared using the log-rank test. Data
were analyzed by analysis of variance (ANOVA) using GraphPad Prism
6 (GraphPad Software, Inc., La Jolla, CA, USA). In the event that
ANOVA justified post hoc comparisons, these were conducted using
Neuman-Keuls or Tukey's multiple-comparisons test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Association between T-cadherin
expression and survival
To investigate how T-cadherin expression affected
the prognosis of patients with GC, the Kaplan-Meier method was used
to evaluate the association of overall survival and T-cadherin
expression levels (Fig. 1). A total
of 81 patients with GC, including 30 with T-cadherin-negative
disease (≤10% or no positive cancer cells in tissue sections) and
51 with T-cadherin-positive disease (>10% positive cancer
cells), were followed for 2–60 months. The T-cadherin-negative
group had a significantly worse prognosis compared with the
T-cadherin-positive group (median survival: 18 months vs. 43
months, P<0.05).
Effect of T-cadherin on cell
growth
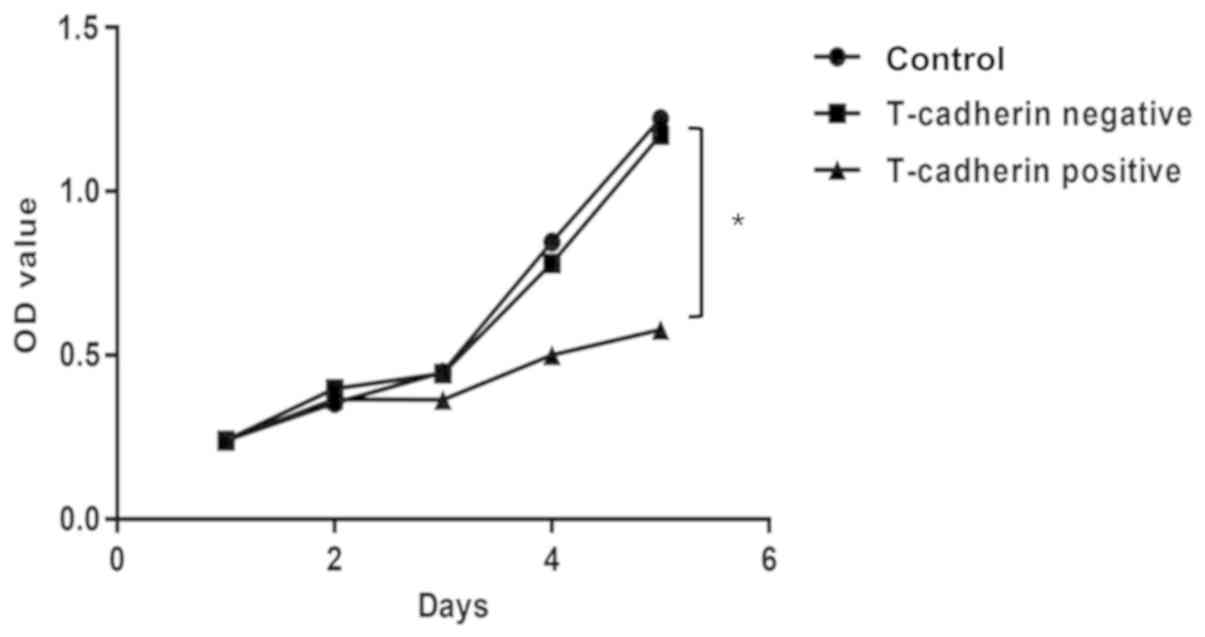
To assess roles of T-cadherin in GC cells, a stable
T-cadherin-overexpressing HGC-27 cell line was established and
T-cadherin expression was confirmed using RT-qPCR (data not shown).
T-cadherin expression increased in cells transfected with
pcDNA3.1-Tadherin but not in cells transfected with empty pcDNA3.1.
An MTT cell proliferation assay was conducted to investigate the
effect of T-cadherin on HGC-27 cell growth. Growth curves
demonstrated that T-cadherin-overexpressing cells exhibited
significant growth suppression compared with cells transfected with
empty plasmid, with growth inhibition rates of 31.09% at 5 days
post-transfection (Fig. 2).
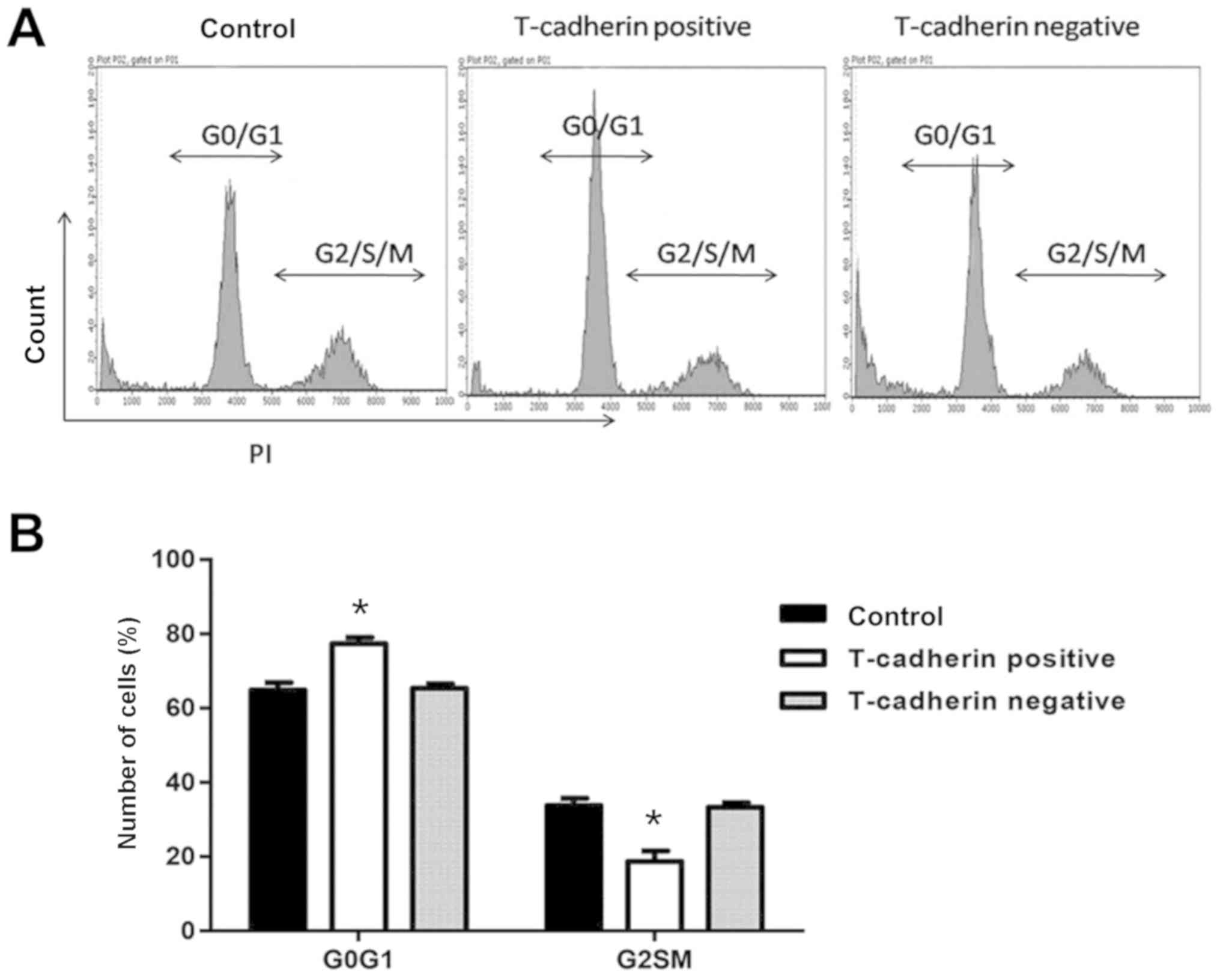
Effect of T-cadherin on cell
cycle
The effect of T-cadherin on the cell cycle of HGC-27
cells was determined using flow cytometry. Of HGC-27 cells
transfected with pcDNA3.1-Tadherin, 77.4% remained in the
G0/G1 phase, an increased percentage compared
with cells transfected with empty vector (65.3%). Furthermore, the
percentage of T-cadherin-overexpressing cells in the S/G2/M phase
decreased significantly to 18.7%, compared with 33.2% for
vector-transfected cells (P<0.05; Fig. 3), suggesting that T-cadherin
overexpression induced cell cycle arrest in the
G0/G1 phase of HGC-27 cells.
Effect of T-cadherin on cell invasion
and migration
To examine whether T-cadherin overexpression may
inhibit cell mobility, a Transwell migration assay was conducted.
Significantly fewer T-cadherin-overexpressing HGC-27 cells migrated
compared with empty vector-transfected cells (P<0.05; Fig. 4A). An invasion assay yielded a
similar trend, with a 64.6% reduction in invasiveness among
T-cadherin-overexpressing HGC-27 cells compared with control and
vector-transfected cells (P<0.05; Fig. 4B). These findings suggest that
T-cadherin ameliorated malignant phenotypes of HGC-27 cells by
inhibiting cell migration and invasion.
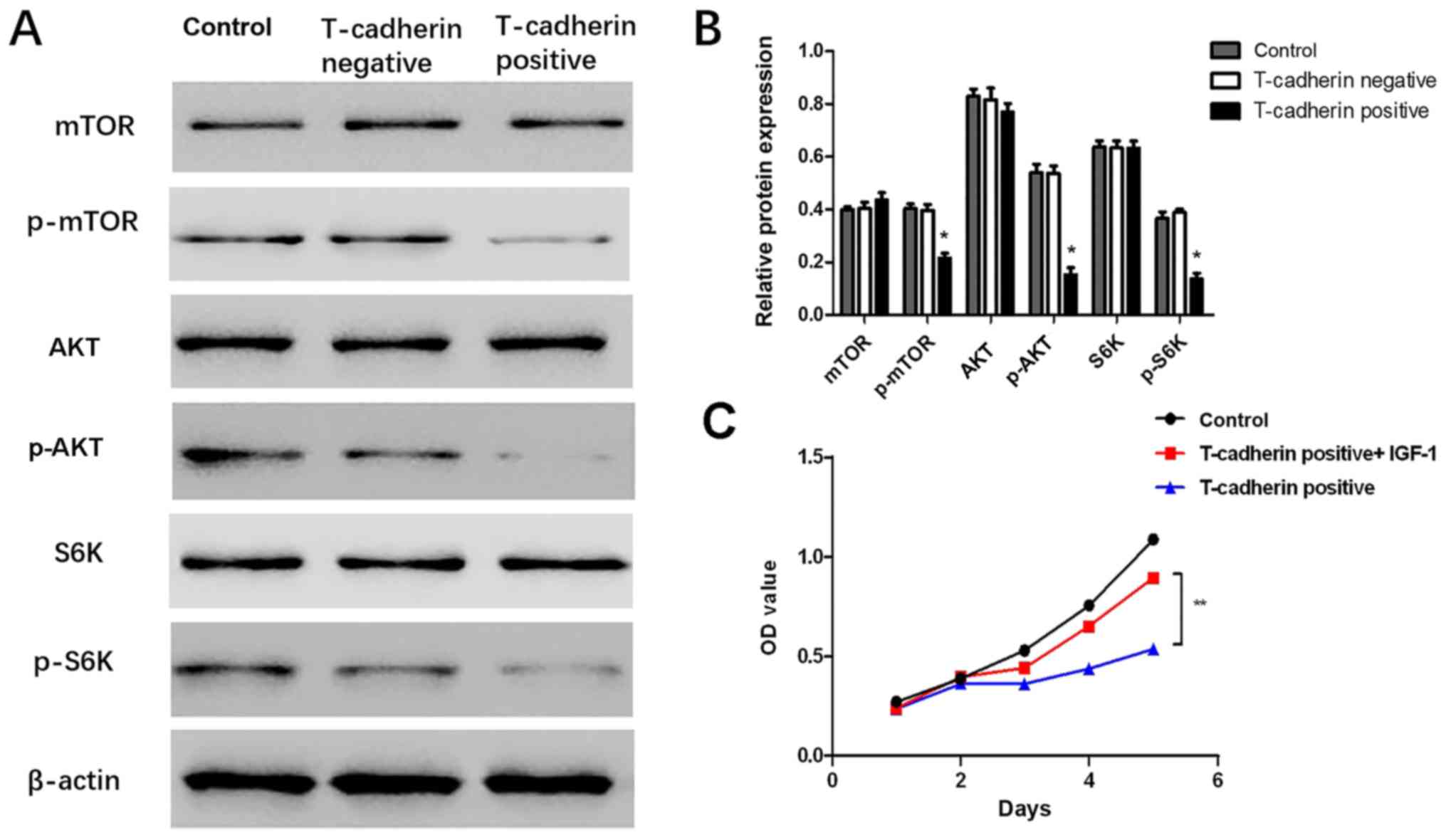
T-cadherin overexpression inhibits
AKT/mTOR activity
To uncover potential mechanisms for
T-cadherin-associated regulation of GC, western blot assays were
performed to validate whether AKT and its downstream targets were
altered in response to T-cadherin overexpression. The results
demonstrated that levels of p-AKT, p-mTOR and p-S6K were
significantly decreased in T-cadherin-overexpressing HGC-27 cells
(P<0.05; Fig. 5A and B),
suggesting that T-cadherin expression may regulate AKT/mTOR
signaling pathway activities. The current study further
investigated whether effects associated with T-cadherin
overexpression may be reversed by administration of insulin-like
growth factor-1 (IGF-1), an AKT-activator. It was observed that
cell viability was partly restored when IGF-1 was added to the
culture medium (Fig. 5C). These
results suggested T-cadherin overexpression suppressed gastric
tumorigenesis potentially through inhibition of the AKT/mTOR
signaling pathway.
Discussion
The current study focused on T-cadherin, the only
cadherin known to be membrane-anchored via a GPI anchor rather than
a transmembrane domain (23).
Previous studies have described the human CDH13 gene to be
an anti-tumor gene, as its expression is suppressed in several
types of cancer (16,27,28).
T-cadherin has been reported to inhibit bladder tumor cell
proliferation, invasion and angiogenesis, whereas reduced
T-cadherin was associated with a poor prognosis among patients with
bladder cancer (29–31). However, few studies have reported
associations between T-cadherin expression and clinicopathological
features in GC.
In a previous study on the biological activity of
T-cadherin in GC, it was reported that mRNA levels and T-cadherin
protein expression were significantly downregulated in GC tissues
compared with adjacent noncancerous tissues (24). Another study observed that
downregulation of T-cadherin in tumor correlated with larger tumor
size (diameter >4 cm), invasiveness, poor differentiation, lymph
node metastasis and higher TNM stage (25). The current study revealed that
T-cadherin expression was associated with overall survival in a
follow-up study of 81 patients. Patients with high T-cadherin
expression levels exhibited a significantly higher postoperative
survival rate compared with patients with low T-cadherin levels,
suggesting that T-cadherin may be useful as a therapeutic target
and indicator of GC prognosis.
Previous studies on the effect of T-cadherin on cell
growth reported cell type-dependent outcomes (32,33).
Small interfering RNA-mediated silencing of T-cadherin expression
had no significant effect on growth of Mahlavu hepatocellular
carcinoma cells (34,35). However, Huang et al (36) demonstrated that T-cadherin inhibited
growth of C6 glioma cells by increasing cell attachments to
fibronectin and decreasing cell mobility. Similar to this, the
current study revealed that T-cadherin overexpression inhibited
growth of HGC-27 cells and induced G2 phase arrest
during cell cycle, with a corresponding increase in the
G0/G1 phase. In addition, T-cadherin
overexpression significantly inhibited MGC8-03 and AGS GC cell
growth, migration and invasion (24), suggesting that T-cadherin exerts
antiproliferation activity in different GC cell lines. HGC-27 was
established through culturing of metastatic lymph node cells from a
patient with GC diagnosed histologically as undifferentiated
carcinoma (37). The current study
suggested T-cadherin downregulation may be a risk factor for lymph
node metastasis in GC.
T-cadherin negatively regulated squamous cell
carcinoma growth by regulating cell adhesion to the extracellular
matrix and β1 integrins and inhibiting epidermal growth factor
receptor phosphorylation to reduce invasiveness (17). Invasiveness and metastasis are
important biological characteristics of malignancies that affect
disease recurrence and influence the prognosis of cancer patients
(38). In the present study, the
results of Transwell assays on migration and invasion revealed that
T-cadherin overexpression significantly decreased both
characteristics in HGC-27 cells. In other words, T-cadherin may
promote overall survival in patients with GC by partial inhibition
of tumor cell invasion and metastasis. These results were
consistent with findings of previous studies. Yan et al
(35) indicated that cell
proliferation decreased in HepG2 cells expressing high levels of
T-cadherin. Philippova et al (17) observed an increase in squamous cell
carcinoma invasion and metastasis in the absence of T-cadherin and
Hebbard et al (39) reported
that loss of T-cadherin promoted tumor angiogenesis and metastasis
in breast cancer.
It is well known that AKT/mTOR signaling serves a
critical role in tumor development and progression (40,41). The
current study determined effects of T-cadherin on AKT/mTOR
signaling in HGC-27 cells. mTOR-dependent regulation of ribosomal
gene transcription requires S6K1 (9). The current study confirmed that
T-cadherin overexpression decreased p-AKT, p-mTOR and p-S6K
expression in HGC-27 cells, when compared with blank and negative
control cells, but did not affect AKT, mTOR and S6K. Additionally,
AKT-activator IGF-1 significantly inhibited the suppressive role of
T-cadherin overexpression in HGC-27 cells, suggesting that AKT/mTOR
may act as downstream signaling mediator of T-cadherin. A previous
study reported that T-cadherin overexpression suppressed GC cell
migration and invasion by upregulating E-cadherin expression and
downregulation of vimentin and matrix metalloproteinase-2
expression (10). The current study
investigated effects of AKT/mTOR signaling in HGC-27 cells
regulated by T-cadherin, however, the mechanisms by which
T-cadherin influences the AKT/mTOR signaling pathway require
further investigation. Luciferase and pull down assays may be
performed to demonstrate whether T-cadherin directly or indirectly
regulates downstream markers.
In conclusion, the present study provided evidence
for the role of T-cadherin in GC tumorigenesis. It demonstrated
that overall survival was associated with T-cadherin
overexpression. Furthermore, T-cadherin overexpression
significantly inhibited HGC-27 cell proliferation and led to cell
cycle arrest in the G0/G1 phase. It was
further demonstrated that T-cadherin-overexpressing HGC-27 cells
exhibited reduced invasiveness and metastatic potential. Studies of
the molecular mechanism suggested that T-cadherin regulated
AKT/mTOR signaling pathway proteins and their downstream mediators.
Administration of AKT-activator IGF-1 in T-cadherin-overexpressing
HGC-27 cells restored the proliferation phenotype. Based on these
results, it is suggested that T-cadherin may be a novel target for
therapeutic intervention of GC.
Acknowledgements
Not applicable.
Funding
The study was supported by the Fujian Natural
Science Foundation (grant no. 2015J01439).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL conceived, designed and performed experiments,
analyzed data and prepared the manuscript. ZC conceived and
designed experiments, analyzed data and prepared the manuscript.
ZH, FC, ZY, SL and WW performed experiments. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of the Second Affiliated Hospital of Fujian Medical
University (Quanzhou, Fujian, China) and all patients agreed to
participate in the study.
Patient consent for publication
All patients provided their informed consent for
publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lin Y, Ueda J, Kikuchi S, Totsuka Y, Wei
WQ, Qiao YL and Inoue M: Comparative epidemiology of gastric cancer
between Japan and China. World J Gastroenterol. 17:4421–4428. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Uemura N, Okamoto S, Yamamoto S, Matsumura
N, Yamaguchi S, Yamakido M, Taniyama K, Sasaki N and Schlemper RJ:
Helicobacter pylori infection and the development of gastric
cancer. N Engl J Med. 345:784–789. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Yang J, Wang J, Guo H and Jing N:
LMO1 is a novel oncogene in lung cancer, and its overexpression is
a new predictive marker for anti-EGFR therapy. Med Oncol.
31:992014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Angst BD, Marcozzi C and Magee AI: The
cadherin superfamily: Diversity in form and function. J Cell Sci.
114:629–641. 2001.PubMed/NCBI
|
|
5
|
Dasen B, Vlajnic T, Mengus C, Ruiz C,
Bubendorf L, Spagnoli G, Wyler S, Erne P, Resink TJ and Philippova
M: T-cadherin in prostate cancer: Relationship with cancer
progression, differentiation and drug resistance. J Pathol Clin
Res. 3:44–57. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Philippova M, Joshi M, Kyriakakis E, Pfaff
D, Erne P and Resink T: A guide and guard: The many faces of
T-cadherin. Cell Signal. 21:1035–1044. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, beta-catenin, and ZEB1 in malignant progression of
cancer. Cancer Metastasis Rev. 28:151–166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fedor-Chaiken M, Hein PW, Stewart JC,
Brackenbury R and Kinch MS: E-cadherin binding modulates EGF
receptor activation. Cell Commun Adhes. 10:105–118. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Suyama K, Shapiro I, Guttman M and Hazan
RB: A signaling pathway leading to metastasis is controlled by
N-cadherin and the FGF receptor. Cancer Cell. 2:301–314. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Derycke LD and Bracke ME: N-cadherin in
the spotlight of cell-cell adhesion, differentiation,
embryogenesis, invasion and signalling. Int J Dev Biol. 48:463–476.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tryndyak VP, Beland FA and Pogribny IP:
E-cadherin transcriptional down-regulation by epigenetic and
microRNA-200 family alterations is related to mesenchymal and
drug-resistant phenotypes in human breast cancer cells. Int J
Cancer. 126:2575–2583. 2010.PubMed/NCBI
|
|
12
|
Ivanov D, Philippova M, Allenspach R, Erne
P and Resink T: T-cadherin upregulation correlates with cell-cycle
progression and promotes proliferation of vascular cells.
Cardiovasc Res. 64:132–143. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang Y, Dai Y and Huo J: Decreased
expression of T-cadherin is associated with gastric cancer
prognosis. Hepatogastroenterology. 59:1294–1298. 2012.PubMed/NCBI
|
|
14
|
Kong DD, Yang J, Li L, Wang W, Chen YN,
Wang SB and Zhou YZ: T-cadherin association with
clinicopathological features and prognosis in axillary lymph
node-positive breast cancer. Breast Cancer Res Treat. 150:119–126.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Z, Wang B, Guo H, Shi G and Hong X:
Clinicopathological significance and potential drug target of
T-cadherin in NSCLC. Drug Des Devel Ther. 9:207–216.
2014.PubMed/NCBI
|
|
16
|
Ren JZ and Huo JR: Correlation between
T-cadherin gene expression and aberrant methylation of T-cadherin
promoter in human colon carcinoma cells. J Med Oncol. 29:915–918.
2012. View Article : Google Scholar
|
|
17
|
Philippova M, Pfaff D, Kyriakakis E,
Buechner SA, Iezzi G, Spagnoli GC, Schoenenberger AW, Erne P and
Resink TJ: T-cadherin loss promotes experimental metastasis of
squamous cell carcinoma. Eur J Cancer. 49:2048–2058. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bosserhoff AK, Ellmann L, Quast AS, Eberle
J, Boyle GM and Kuphal S: Loss of T-cadherin (CDH-13) regulates AKT
signaling and desensitizes cells to apoptosis in melanoma. Mol
Carcinog. 53:635–647. 2014.PubMed/NCBI
|
|
19
|
Toyooka KO, Toyooka S, Virmani AK,
Sathyanarayana UG, Euhus DM, Gilcrease M, Minna JD and Gazdar AF:
Loss of expression and aberrant methylation of the CDH13
(H-cadherin) gene in breast and lung carcinomas. Cancer Res.
61:4556–4560. 2001.PubMed/NCBI
|
|
20
|
Wang XD, Wang BE, Soriano R, Zha J, Zhang
Z, Modrusan Z, Cunha GR and Gao WQ: Expression profiling of the
mouse prostate after castration and hormone replacement:
Implication of H-cadherin in prostate tumorigenesis.
Differentiation. 75:219–234. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takeuchi T, Misaki A, Liang SB, Tachibana
A, Hayashi N, Sonobe H and Ohtsuki Y: Expression of T-cadherin
(CDH13, H-Cadherin) in human brain and its characteristics as a
negative growth regulator of epidermal growth factor in
neuroblastoma cells. J Neurochem. 74:1489–1497. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee SW: H-cadherin, a novel cadherin with
growth inhibitory functions and diminished expression in human
breast cancer. Nat Med. 2:776–782. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takeuchi T, Misaki A, Chen BK and Ohtsuki
Y: H-cadherin expression in breast cancer. Histopathology.
35:87–88. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin J, Chen Z, Huang Z, Chen F, Ye Z, Lin
S and Wang W: Upregulation of T-cadherin suppresses cell
proliferation, migration and invasion of gastric cancer in vitro.
Exp Ther Med. 14:4194–4200. 2017.PubMed/NCBI
|
|
25
|
Wei B, Shi H, Lu X, Shi A, Cheng Y and
Dong L: Association between the expression of T-cadherin and
vascular endothelial growth factor and the prognosis of patients
with gastric cancer. Mol Med Rep. 12:2075–2081. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chan DW, Lee JMF, Chan PC and Ng IO:
Genetic and epigenetic inactivation of T-cadherin in human
hepatocellular carcinoma cells. Int J Cancer. 123:1043–1052. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Adachi Y, Takeuchi T, Nagayama T and
Furihata M: T-cadherin modulates tumor-associated molecules in
gallbladder cancer cells. Cancer Invest. 28:120–126. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin Y, Sun G, Liu X, Chen Y and Zhang C:
Clinical significance of T-cadherin tissue expression in patients
with bladder transitional cell carcinoma. Urol Int. 86:340–345.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin YL, Xie PG and Ma JG: Aberrant
methylation of CDH13 is a potential biomarker for predicting the
recurrence and progression of non-muscle-invasive bladder cancer.
Med Sci Monit. 20:1572–1577. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin YL, He ZK, Li ZG and Guan TY:
Downregulation of CDH13 expression promotes invasiveness of bladder
transitional cell carcinoma. Urol Int. 90:225–232. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fujishima Y, Maeda N, Matsuda K, Masuda S,
Mori T, Fukuda S, Sekimoto R, Yamaoka M, Obata Y, Kita S, et al:
Adiponectin association with T-cadherin protects against neointima
proliferation and atherosclerosis. FASEB J. 31:1571–1583. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kong DD, Wang MH, Yang J, Li L, Wang W,
Wang SB and Zhou YZ: T-cadherin is associated with prognosis in
triple-negative breast cancer. Oncology letters. 14:2975–2981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Riou P, Saffroy R, Chenailler C, Franc B,
Gentile C, Rubinstein E, Resink T, Debuire B, Piatier-Tonneau D and
Lemoine A: Expression of T-cadherin in tumor cells influences
invasive potential of human hepatocellular carcinoma. FASEB J.
20:2291–2301. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yan Q, Zhang ZF, Chen XP, Gutmann DH,
Xiong M, Xiao ZY and Huang ZY: Reduced T-cadherin expression and
promoter methylation are associated with the development and
progression of hepatocellular carcinoma. Int J Oncol. 32:1057–1063.
2008.PubMed/NCBI
|
|
36
|
Huang ZY, Wu Y, Hedrick N and Gutmann DH:
T-cadherin-mediated cell growth regulation involves G2 phase arrest
and requires p21(CIP1/WAF1) expression. Mol Cell Biol. 23:566–578.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akagi T and Kimoto T: Human cell line
(HGC-27) derived from the metastatic lymph node of gastric cancer.
Acta Medica Okayama. 30:215–219. 1976.PubMed/NCBI
|
|
38
|
Brown GT and Murray GI: Current
mechanistic insights into the roles of matrix metalloproteinases in
tumour invasion and metastasis. J Pathol. 237:227–281. 2015.
View Article : Google Scholar
|
|
39
|
Hebbard LW, Garlatti M, Young LJ, Cardiff
RD, Oshima RG and Ranscht B: T-cadherin supports angiogenesis and
adiponectin association with the vasculature in a mouse mammary
tumor model. Cancer Res. 68:1407–1416. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Claudio F: Targeting the PI3K/AKT/mTOR
pathway in prostate cancer development and progression: Insight to
therapy. Clin Cancer Drugs. 20:R83–R99. 2016.
|
|
41
|
Ewald F, Nörz D, Grottke A, Bach J,
Herzberger C, Hofmann BT, Nashan B and Jücker M: Vertical targeting
of AKT and mTOR as well as dual targeting of AKT and MEK signaling
is synergistic in hepatocellular carcinoma. J Cancer. 6:1195–1205.
2015. View Article : Google Scholar : PubMed/NCBI
|