Introduction
Diabetes, also known as diabetes mellitus (DM), has
an increased morbidity and mortality and represents a serious
public health issue worldwide (1,2).
Vascular complications are involved in pathological changes of DM
(3,4)
and are the leading cause of mortality in this population. The
endothelium, covering the luminal surface of all blood vessels, has
a key role in the maintenance of vascular homeostasis (5,6).
Endothelial dysfunction initiates vascular pathogenesis and may
finally lead to diabetic vasculopathy (7). The luminal surface of endothelial cells
is covered by a thick layer of glycocalyx, which is composed of
proteoglycans (PGs) and glycoproteins (8,9).
Therefore, dysfunction of glycocalyx may promote the development of
diabetic vasculopathy.
It has been demonstrated that endothelial glycocalyx
has a significant impact on factors including vascular permeability
(10), inflammation (11), coagulation (12) and mechanotransduction (13). It was observed that the thickness of
the glycocalyx was altered and its integrity was disrupted in a
streptozocin-induced animal model of diabetes (14). Diabetic patients are characterized by
endothelial glycocalyx damage, the severity of which is associated
with vascular damage (15,16). An in vitro study also observed
that hyperglycemia induced glycocalyx dysfunction in either
microvascular or macrovascular endothelial cells (17,18). It
has been reported that the loss of glycocalyx leads to a reduction
in endothelial surface charge and accelerates atherosclerosis in
patients with type 2 diabetes (19).
Taken together, these studies indicate that glycocalyx dyfunction
drives the development of vascular complications in diabetes.
Panax notoginseng is one of the most commonly
used Chinese herbal medicines due to its efficacy in promoting
blood circulation and removing blood stasis (20). It is frequently used in the
management of diabetes in Asian countries (21) and also possesses cardiovascular
protection effects by preserving endothelial cell function and
inhibiting thermogenesis (22).
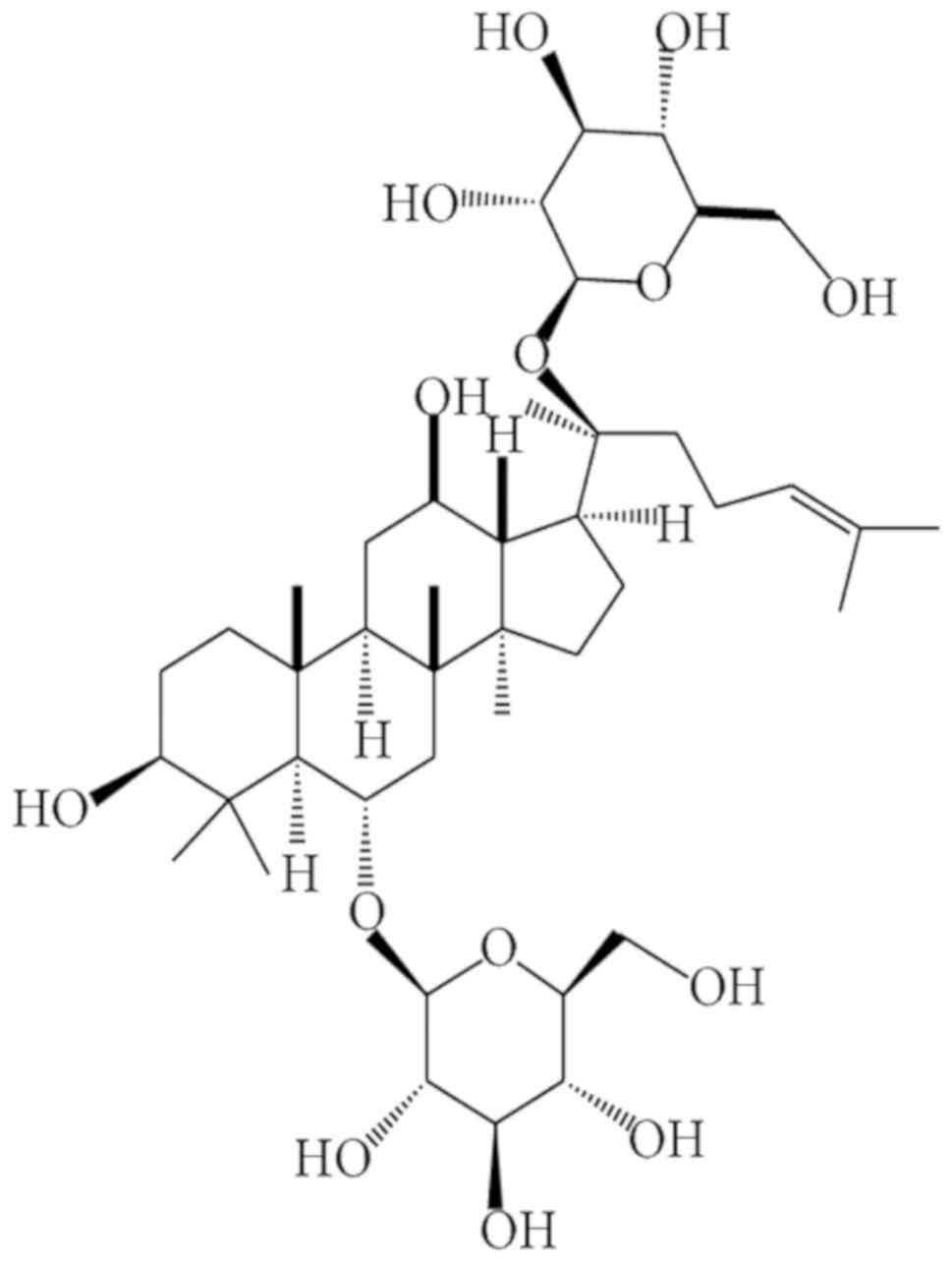
Ginsenoside Rg1 (Rg1; Fig. 1), the
major active component isolated from Panax notoginseng,
appears to be accountable for its extensive pharmacological
actions, including anti-oxidant, anti-inflammatory and anti-cancer
effects (23). Rg1 has been
demonstrated to have cardiovascular protection effects, and may be
of potential preventive and therapeutic value for cardiovascular
injury in diabetic patients (24,25). In
addition, Rg1 has been demonstrated to be potent in improving renal
function and attenuating diabetes-induced renal damage (26,27).
However, the underlying mechanisms remain to be elucidated.
Based on the above, the presents study hypothesized
that Rg1 may attenuate diabetes-induced vascular dysfunction via
restoring the loss of endothelial glycocalyx. To test this
hypothesis, the effect of high glucose on the endothelial
glycocalyx and endothelial barrier function, and the potential
benefits of Rg1 in protecting endothelial barrier function from
high glucose-induced endothelial cell injury were investigated.
Materials and methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
purchased from the American Type Culture Collection (Manassas, VA,
USA). The cells were cultured in low-glucose Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 5.5 mmol/l D-glucose, supplemented
with L-glutamine, 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
at 37°C in an incubator containing 5% CO2. Cells were
seeded into cell culture dishes and cultured until confluent.
Treatment
For control and high-glucose treatment, HUVECs were
cultured with DMEM containing 5.5 and 30 mmol/l D-glucose,
respectively. For drug treatment, the cells were incubated with
high-glucose DMEM, and at the same time, Rg1 (>98% pure; DiDa
Kexiang Biological Co., Ltd., Guizhou, China) was added to the
culture at concentrations ranging from 10−8 to
10−5 mol/l for 1 or 3 days.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HUVECs using TRIzol
reagent (Thermo Fisher Scientific, Inc.). RNA was
reverse-transcribed into complementary (c)DNA using the RevertAid
First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Inc.).
The mRNA levels of heparanase (HPSE) were quantified with an
RT-qPCR system (Mastercycler realplex2; Eppendorf, Hamburg,
Germany), using SYBR Green SuperMix (Roche Diagnostics, Mannheim,
Germany) with appropriate primers pairs. Sequences of primers used
in the present study were as follows: HPSE forward,
5′-CCAAAGTTGCTGCTTGCATC-3′ and reverse, 5′-AGTGTCCCAGTGTCTCTCAA-3′;
GAPDH forward, 5′-CTGGGCTACACTGAGCACC-3′ and reverse,
5′-AAGTGGTCGTTGAGGGCAATG−3′. The reaction was started by
pre-incubation at 95°C for 10 min, followed by 40 cycles of
amplification (95°C for 15 sec, 65°C for 15 sec and 72°C for 20
sec). Gene expression levels of HPSE were normalized to those of
the reference gene GAPDH measured in the same sample and the
results were analyzed by the 2−∆∆Cq method (28).
Western blot analysis
Total proteins were prepared using the lysis buffer
(20 mmol/l Tris, pH 7.4, 150 mmol/l NaCl, 1 mmol/l EDTA, 1
mmol/lEGTA, 1% Triton X-100, 2.5 mmol/l deoxycholic acid, 1 mmol/l
β-glycerophosphate and 1 mmol/l Na3VO4),
supplemented with protease inhibitors. Protein concentration was
determined using a BCA protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China) according to the manufacturer's
protocol. The protein of each sample (25 µg) was separated using
10% SDS-PAGE and then transferred to polyvinylidene difluoride
membranes (EMD Millipore, Billerica, MA, USA). After blocking with
5% fat-free milk for 1 h at room temperature, the membranes were
incubated with the following respective primary antibodies:
Syndecan-1 monoclonal antibody (1:1,000 dilution; cat. no.
ab128936; Abcam, Cambridge, MA, USA), glypican-1 polyclonal
antibody (1:1,000 dilution; cat. no. NBP1-33197; Novus Biologicals
LLC, Littleton, CO, USA) and GAPDH monoclonal antibody (1:1,000
dilution; cat. no. sc32233; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA) at 4°C overnight. The membranes were then gently washed
for three times (5 min each) and incubated with horseradish
peroxide-conjugated goat anti-rabbit secondary antibody (1:3,000
dilution; cat. no. ZB2301; Zhongshan Goldenbridge Bio, Beijing,
China) at room temperature for 1.5 h. After washing, the protein
bands were visualized with chemiluminescent substrate (EMD
Millipore) for 1 min, and capturing of images and densitometric
analysis were performed using an imaging station (Bio-Rad
Laboratoris, Inc., Hercules, CA, USA). The relative protein
expression levels were normalized to GAPDH and the ratio was
compared with that of the control group.
Detection of endothelial surface
glycocalyx
Wheat germ agglutinin (WGA) from Triticum
vulgaris binds to sugar moieties of glycocalyx present on the
cell surface, the majority of which are likely to be PG
constituents of glycocalyx. Therefore, endothelial surface
glycocalyx was labeled by fluorescein isothiocyanate
(FITC)-conjugated WGA lectin as previously reported (29). Cells were grown to confluence on
glass coverslips and fixed in 4% paraformaldehyde for 10 min. After
being washed for three times, the cells were incubated with
FITC-WGA lectin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at
2 µg/ml for 30 min. Coverslips were mounted and visualized using a
fluorescence microscope (Leica Microsystems, Wetzlar, Germany).
Transendothelial electrical resistance
(TEER) measurement
Cells were seeded onto microporous polyester
membranes (0.4-µm pore size) of Transwell filter inserts (Corning
Inc., Corning, NY, USA). The cells were grown onto the upper
chamber of the Transwell until confluent. The HUVECs were
stimulated with vehicle, 30 mmol/l high glucose or Rg1 for 24 h as
indicated. The TEER of the monolayer of HUVECS was measured using a
Millicell ERS-2 Volt-Ohm meter (EMD Millipore) according to the
protocol of a previous study (30).
After subtraction of the value determined using a blank, cell-free
filter, the mean value of the TEER was expressed in common units
(Ωcm2). TEER values of a vehicle-treated monolayer of
endothelial cells were designated as baseline values. The
percentage of TEER relative to baseline value was calculated via
the following formula: TEER%=(TEER of experimental wells/baseline
TEER of experimental wells) ×100%.
Transendothelial albumin passage
The transendothelial passage of albumin was analyzed
by measuring the passage of FITC-labeled bovine serum albumin (BSA;
Sigma-Aldrich; Merck KGaA) across the monolayer as described
previously (31). In brief, cells
were seeded onto the upper chambers of Transwells and allowed to
grow to confluence as described above. The medium in the insert was
replaced with serum-free medium (SFM) containing 0.5 mg/ml
FITC-labelled BSA and the medium in the well was replaced by SFM
only. After 1, 2 and 3 h of incubation, the medium was collected
from each well, and the fluorescence of the aliquots was measured
on a fluorometer with excitation at 495 nm and emission at 520 nm.
The amount of albumin passing the endothelial cell monolayer was
calculated with a standard curve generated from a set of
FITC-labeled BSA dilutions.
Statistical analyses
All data were analyzed using SPSS software (version
17.0; SPSS, Inc., Chicago, IL, USA). Values are expressed as the
mean ± standard deviation. The independent Student's t-test and
one-way analysis of variance (ANOVA) with a least significant
difference (LSD) post hoc test, were used for comparison between
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of Rg1 on the PG core proteins
in high glucose-induced HUVECs
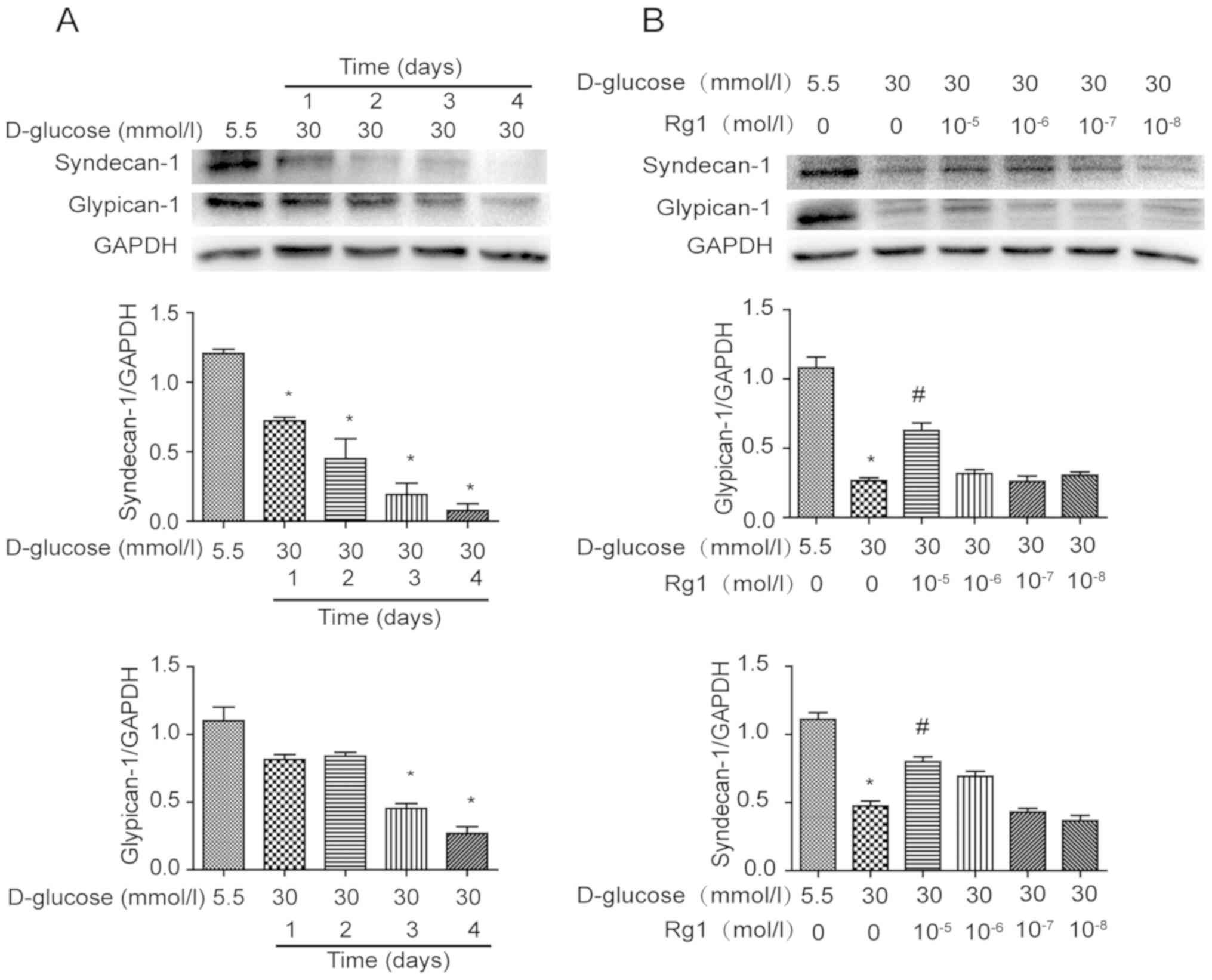
PG core proteins are important constituents of
glycocalyx on the cell surface. Expression of PG core proteins
syndecan-1 and glypican-1 was analyzed by western blot analysis
after exposure to high glucose for different durations (Fig. 2). It was demonstrated that syndecan-1
was gradually decreased in HUVECs incubated with high glucose from
1–4 days. Over the same duration, the expression of glypican-1 was
also decreased by high-glucose stimulation, with the changes being
significant at 3 and 4 days (Fig.
2A). HUVECs under high-glucose stimulation were then treated
with different concentrations of Rg1 for 3 days. It was observed
that treatment with Rg1 increased PG core proteins in HUVECs at
concentrations ranging from 10−8 to 10−5
mol/l and a significant difference was identified at
10−5 mol/l (Fig. 2B).
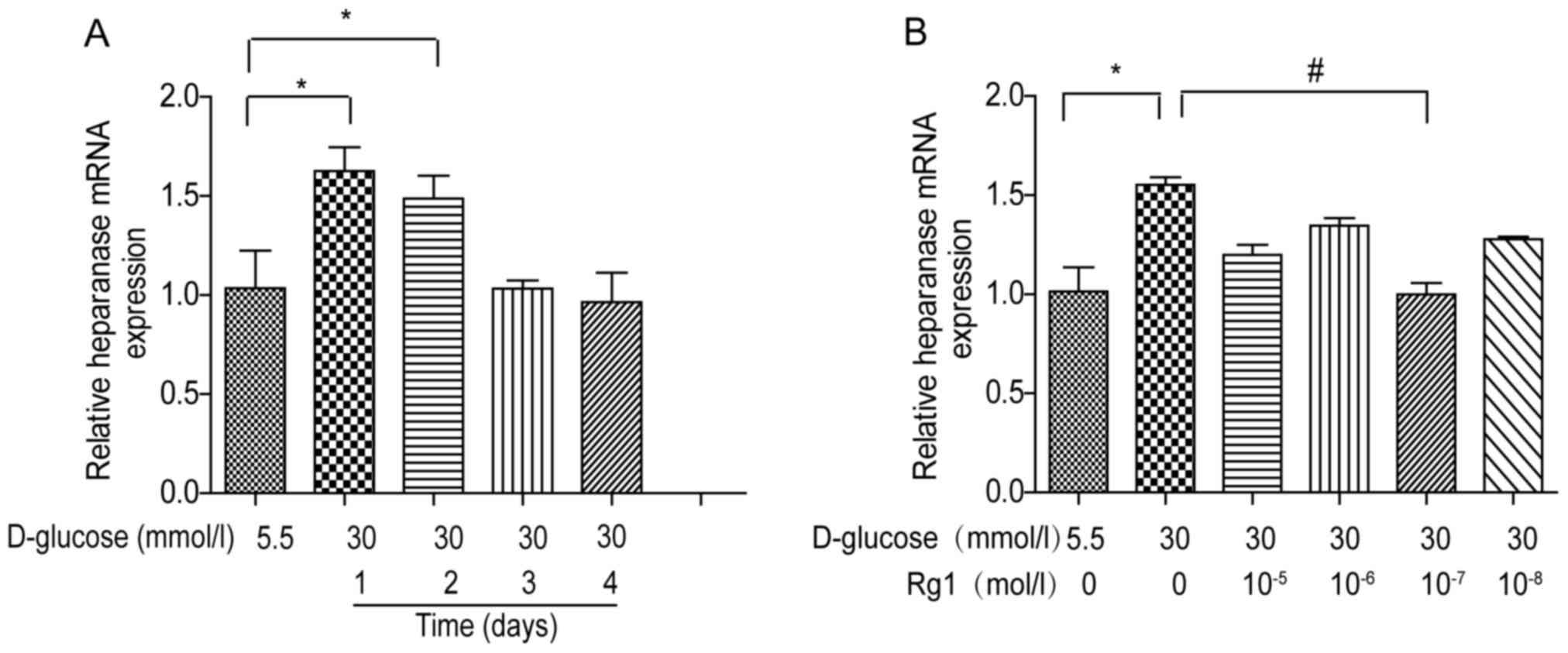
High glucose-induced HPSE expression
is attenuated by Rg1 treatment in HUVECs
To assess the effects of high glucose and Rg1
treatment on HPSE expression, HUVECs were incubated with 30 mmol/l
glucose and different concentrations of Rg1 as indicated. The
expression of HPSE mRNA was detected by RT-qPCR. It was observed
that HPSE mRNA expression in high glucose-treated cells was rapidly
increased and reached a peak at 1 day, and then it returned to
baseline levels at 3 and 4 days (Fig.
3A). To observe the effect of Rg1, HUVECs were first incubated
with high glucose, followed by different concentrations of Rg1 for
1 day. It was indicated that HPSE expression in HUVECs was reduced
by treatment with Rg1 at concentrations ranging from
10−8 to 10−5 mol/l, and a significant
difference was observed at 10−7 mol (Fig. 3B).
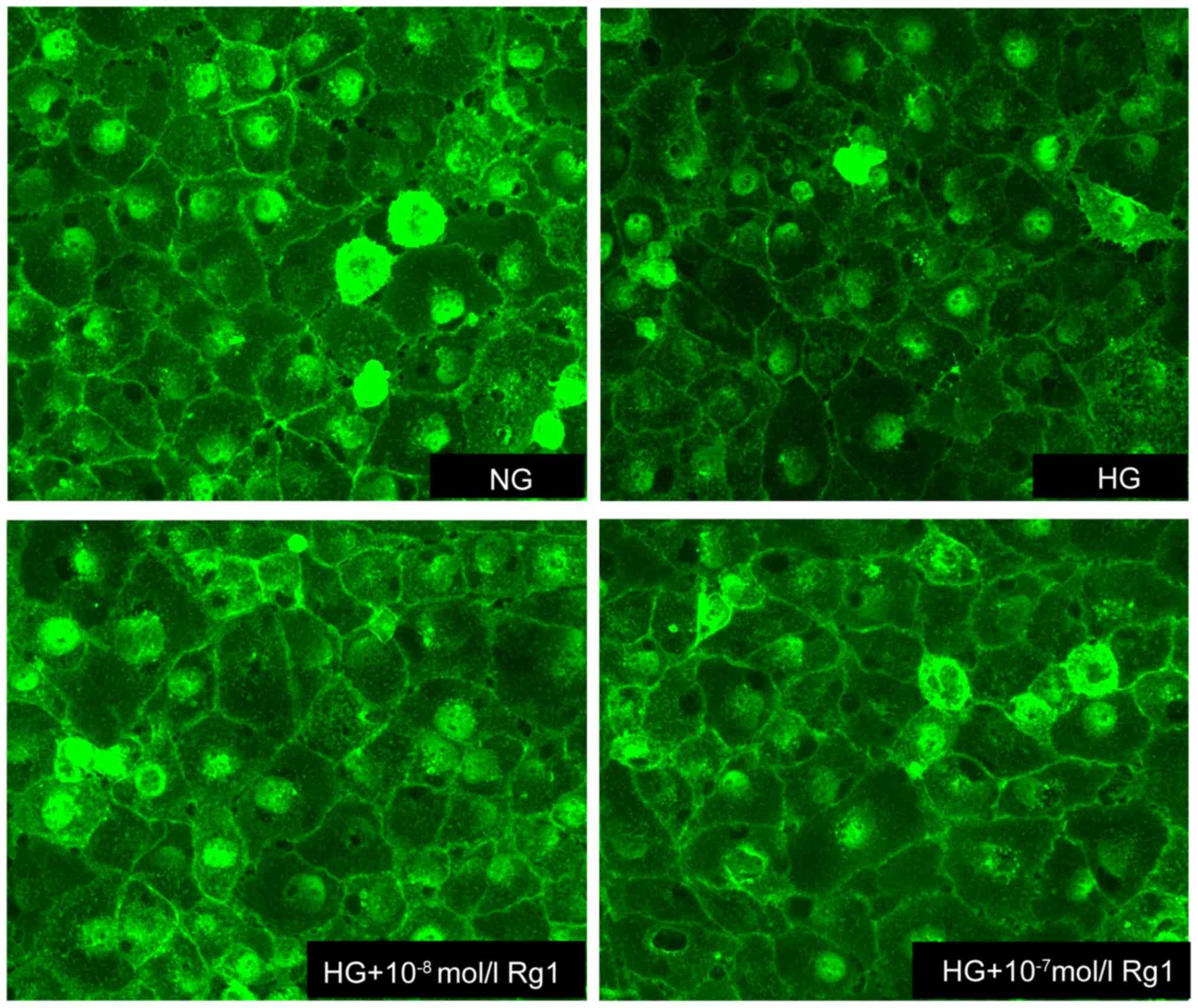
Rg1 prevents the disruption of the
glycocalyx induced by high glucose
WGA-FITC, which binds to sugar residues on cell
surface, was used to quantify the expression of glycocalyx in
HUVECs. A marked reduction in WGA-FITC binding was observed in high
glucose-induced cells compared with untreated controls, suggesting
the disruption of endothelial glycocalyx. As expected, the
expression of glycocalyx was increased by treatment with Rg1,
indicating that Rg1 prevented the loss of glycocalyx and attenuated
the disruption of the glycocalyx in HUVECs incubated with 30 mmol/l
glucose (Fig. 4).
Rg1 treatment increases TEER in the
presence of high glucose
TEER is associated with the integrity of cell
monolayers. Its decline represents an increase in the passage of
water and small molecules across the cell monolayer and an impaired
cell barrier function. To study the effects of Rg1 treatment on
TEER, HUVECs were stimulated with high glucose in the presence or
absence of Rg1. High glucose caused a modest reduction in the mean
TEER by up to 20% relative to that in the controls, which was
inhibited in the presence of Rg1 (Fig.
5A).
High glucose leads to increased
transendothelial albumin passage that is attenuated by treatment
with Rg1
Since 30 mmol/l high glucose leads to decreased
TEER, it was examined whether Rg1 was able to preserve the
permeability of cultured HUVEC monolayers. The permeability was
assessed by measuring the passage of FITC-labeled BSA across the
monolayer, i.e., the transendothelial albumin passage. As presented
in Fig. 5B, stimulation with 30
mmol/l high glucose significantly increased transendothelial
albumin passage across HUVECs, while this effect was inhibited in
the presence of Rg1.
Discussion
A pivotal role of the endothelium is to serve as a
regulated barrier to partially separate the contents of the blood
from the extravascular space (32).
The endothelial glycocalyx covers the luminal surface of the
vascular endothelium (33,34), and is responsible for endothelial
barrier function. Widespread loss of the endothelial surface
glycocalyx leads to damage of vascular function and an elevation in
the microvascular permeability to water as well as albumin, and
results in systemic vascular dysfunction in proteinuric kidney
disease (35). The results of the
present study indicated a marked decrease in the PG core proteins
syndecan-1 and glypican-1 on HUVECs after exposure to high glucose,
while treatment with Rg1 increased the formation of PG core
proteins. This was further confirmed in an assay using WGA-FITC
lectin, in which WGA-FITC lectin binding was markedly reduced after
high-glucose treatment, while this reduction was inhibited by Rg1
treatment. These results suggested that treatment with Rg1
increased the formation of PG and enhanced the integrity of the
endothelial cell monolayer, which is beneficial for preserving
endothelial barrier function under a high-glucose conditions.
In response to persistent activators of the
endothelium, i.e., in diabetes mellitus, hypertension and systemic
inflammation, endothelial cells undergo pathological changes
(36,37), including the induction of the
expression of certain enzymes, including HPSE. HPSE, a degrading
enzyme of the endothelial cell glycocalyx, is the only known
mammalian enzyme to cleave PGs (38). It has been reported that increased
HPSE expression causes damage to the glycocalyx of mouse glomerular
endothelial cells and increases the trans-endothelial albumin
passage of the cell monolayer (39).
The present study indicated that high glucose increased HPSE mRNA
expression on HUVECs at the early stage of high-glucose treatment,
whereas Rg1 reduced the increase of HPSE induced by high glucose.
The results indicated that Rg1 inhibited the production of HPSE to
ameliorate endothelial glycocalyx disorders.
Endothelial glycocalyx is located between the blood
stream and the endothelium, provides a barrier for certain
molecules, and has an important role in endothelial permeability
and endothelial functions (40,41). A
significant degradation of the glomerular glycocalyx has been
reported in the setting of diabetes in that loss of glycocalyx
increases vascular permeability (16). In, TEER is used as an indicator of
the passage of water and small molecules across a cell layer. TEER
is regarded as a measure of the resistance to the passage of ions
across a confluent cell monolayer. In the present study, the
passage of albumin across cell monolayers was tested to evaluate
the permeability of the endothelial cell monolayer, and the results
indicated that Rg1 attenuated the damage of endothelial barrier
function by enhancing TEER and decreasing the cell transendothelial
albumin passage.
In conclusion, the present study demonstrated that
Rg1 inhibited the loss of endothelial glycocalyx and HPSE mRNA
expression, and increased TEER, while decreasing endothelial cell
monolayer permeability and protecting endothelial barrier function.
The present results may provide a novel mechanism of action of Rg1
in the treatment of diabetic vasculopathy.
Acknowledgements
This study was supported by the Science and
Technology Development Fund of Macau (grant no. 071/2014/A), the
Education Department Program of Sichuan Province (grant no.
17TD0046), the Key Project of the Health Department of Sichuan
Province (grant no. 16ZD034) and the Innovative Research Team of
Luzhou (grant no. 2016LZXNYD-T05).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
García-Pérez LE, Alvarez M, Dilla T,
Gil-Guillén V and Orozco-Beltrán D: Adherence to therapies in
patients with type 2 diabetes. Diabetes Ther. 4:175–194. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shaw JE, Sicree RA and Zimmet PZ: Global
estimates of the prevalence of diabetes for 2010 and 2030. Diabetes
Res Clin Pract. 87:4–14. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Potenza MA, Gagliardi S, Nacci C, Carratu'
MR and Montagnani M: Endothelial dysfunction in diabetes: From
mechanisms to therapeutic targets. Curr Med Chem. 16:94–112. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Santi D, Giannetta E, Isidori AM, Vitale
C, Aversa A and Simoni M: Therapy of endocrine disease: Effects of
chronic use of phosphodiesterase inhibitors on endothelial markers
in type 2 diabetes mellitus: A meta-analysis. Eur J Endocrinol.
172:R103–R114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bertoluci MC, Cé GV, da Silva AM,
Wainstein MV, Boff W and Punales M: Endothelial dysfunction as a
predictor of cardiovascular disease in type 1 diabetes. World J
Diabetes. 6:679–692. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Widlansky ME, Gokce N, Keaney JF Jr and
Vita JA: The clinical implications of endothelial dysfunction. J Am
Coll Cardiol. 42:1149–1160. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
De Vriese AS, Verbeuren TJ, Van de Voorde
J, Lameire NH and Vanhoutte PM: Endothelial dysfunction in
diabetes. Br J Pharmacol. 130:963–974. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pries AR, Secomb TW and Gaehtgens P: The
endothelial surface layer. Pflugers Arch. 440:653–666. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reitsma S, Slaaf DW, Vink H, van Zandvoort
MA and oude Egbrink MG: The endothelial glycocalyx: Composition,
functions, and visualization. Pflugers Arch. 454:345–359. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Salmon AH and Satchell SC: Endothelial
glycocalyx dysfunction in disease: Albuminuria and increased
microvascular permeability. J Pathol. 226:562–574. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mulivor AW and Lipowsky HH:
Inflammation-and ischemia-induced shedding of venular glycocalyx.
Am J Physiol Heart Circ Physiol. 286:H1672–H1680. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pearson MJ and Lipowsky HH: Effect of
fibrinogen on leukocyte margination and adhesion in postcapillary
venules. Microcirculation. 11:295–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tarbell JM and Pahakis MY:
Mechanotransduction and the glycocalyx. J Intern Med. 259:339–350.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dogné S, Rath G, Jouret F, Caron N, Dessy
C and Flamion B: Hyaluronidase 1 deficiency preserves endothelial
function and glycocalyx integrity in early streptozotocin-induced
diabetes. Diabetes. 65:2742–2753. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nieuwdorp M, Mooij HL, Kroon J, Atasever
B, Spaan JA, Ince C, Holleman F, Diamant M, Heine RJ, Hoekstra JB,
et al: Endothelial glycocalyx damage coincides with
microalbuminuria in type 1 diabetes. Diabetes. 55:1127–1132. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Broekhuizen LN, Lemkes BA, Mooij HL,
Meuwese MC, Verberne H, Holleman F, Schlingemann RO, Nieuwdorp M,
Stroes ES and Vink H: Effect of sulodexide on endothelial
glycocalyx and vascular permeability in patients with type 2
diabetes mellitus. Diabetologia. 53:2646–2655. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pahwa R, Nallasamy P and Jialal I:
Toll-like receptors 2 and 4 mediate hyperglycemia induced
macrovascular aortic endothelial cell inflammation and perturbation
of the endothelial glycocalyx. J Diabetes Complications.
30:563–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singh A, Fridén V, Dasgupta I, Foster RR,
Welsh GI, Tooke JE, Haraldsson B, Mathieson PW and Satchell SC:
High glucose causes dysfunction of the human glomerular endothelial
glycocalyx. Am J Physiol Renal Physiol. 300:F40–F48. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nassimizadeh M, Ashrafian H, Drury NE,
Howell NJ, Digby J, Pagano D, Frenneaux MP and Born GV: Reduced
negative surface charge on arterial endothelium explains
accelerated atherosclerosis in type 2 diabetic patients. Diab Vasc
Dis Res. 7:213–215. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang X, Xiong X, Wang H and Wang J:
Protective effects of panax notoginseng saponins on
cardiovascular diseases: A comprehensive overview of experimental
studies. Evid Based Complement Alternat Med. 2014:2048402014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uzayisenga R, Ayeka PA and Wang Y:
Anti-diabetic potential of Panax notoginseng saponins (PNS):
A review. Phytother Res. 28:510–516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ling S, Nheu L, Dai A, Guo Z and
Komesaroff P: Effects of four medicinal herbs on human vascular
endothelial cells in culture. Int J Cardiol. 128:350–358. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lü JM, Yao Q and Chen C: Ginseng
compounds: An update on their molecular mechanisms and medical
applications. Curr Vasc Pharmacol. 7:293–302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu HT, Zhen J, Pang B, Gu JN and Wu SS:
Ginsenoside Rg1 ameliorates oxidative stress and myocardial
apoptosis in streptozotocin-induced diabetic rats. J Zhejiang Univ
Sci B. 16:344–354. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu H, Zhen J, Yang Y, Gu J, Wu S and Liu
Q: Ginsenoside Rg1 ameliorates diabetic cardiomyopathy by
inhibiting endoplasmic reticulum stress-induced apoptosis in a
streptozotocin-induced diabetes rat model. J Cell Mol Med.
20:623–631. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang LN, Xie XS, Zuo C and Fan JM: Effect
of ginsenoside Rgl on the expression of TNF-alpha and MCP-1 in rats
with diabetic nephropathy. Sichuan Da Xue Xue Bao Yi Xue Ban.
40:466–471. 2009.(In Chinese). PubMed/NCBI
|
|
27
|
Ma X, Xie X, Zuo C and Fan J: Effects of
ginsenoside Rg1 on streptozocin-induced diabetic nephropathy in
rats. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 27:342–347. 2010.(In
Chinese). PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh A, Satchell SC, Neal CR, McKenzie
EA, Tooke JE and Mathieson PW: Glomerular endothelial glycocalyx
constitutes a barrier to protein permeability. J Am Soc Nephrol.
18:2885–2893. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang W, Xu Q, Wu J, Zhou X, Weng J, Xu J,
Wang W, Huang Q and Guo X: Role of src in vascular
hyperpermeability induced by advanced glycation end products. Sci
Rep. 5:140902015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garsen M, Sonneveld R, Rops AL, Huntink S,
van Kuppevelt TH, Rabelink TJ, Hoenderop JG, Berden JH, Nijenhuis T
and van der Vlag J: Vitamin D attenuates proteinuria by inhibition
of heparanase expression in the podocyte. J Pathol. 237:472–481.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu C, Wu X, Hack BK, Bao L and Cunningham
PN: TNF causes changes in glomerular endothelial permeability and
morphology through a Rho and myosin light chain kinase-dependent
mechanism. Physiol Rep. 3:e126362015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Haraldsson B and Nyström J: The glomerular
endothelium: New insights on function and structure. Curr Opin
Nephrol Hypertens. 21:258–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fridén V, Oveland E, Tenstad O, Ebefors K,
Nyström J, Nilsson UA and Haraldsson B: The glomerular endothelial
cell coat is essential for glomerular filtration. Kidney Int.
79:1322–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salmon AH, Ferguson JK, Burford JL,
Gevorgyan H, Nakano D, Harper SJ, Bates DO and Peti-Peterdi J: Loss
of the endothelial glycocalyx links albuminuria and vascular
dysfunction. J Am Soc Nephrol. 23:1339–1350. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rabelink TJ, de Boer HC and van Zonneveld
AJ: Endothelial activation and circulating markers of endothelial
activation in kidney disease. Nat Rev Nephrol. 6:404–414. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gil N, Goldberg R, Neuman T, Garsen M,
Zcharia E, Rubinstein AM, van Kuppevelt T, Meirovitz A, Pisano C,
Li JP, et al: Heparanase is essential for the development of
diabetic nephropathy in mice. Diabetes. 61:208–216. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van den Hoven MJ, Rops AL, Bakker MA, Aten
J, Rutjes N, Roestenberg P, Goldschmeding R, Zcharia E, Vlodavsky
I, van der Vlag J and Berden JH: Increased expression of heparanase
in overt diabetic nephropathy. Kidney Int. 70:2100–2108. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garsen M, Lenoir O, Rops AL, Dijkman HB,
Willemsen B, van Kuppevelt TH, Rabelink TJ, Berden JH, Tharaux PL
and van der Vlag J: Endothelin-1 induces proteinuria by
heparanase-mediated disruption of the glomerular glycocalyx. J Am
Soc Nephrol. 27:3545–3551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goligorsky MS: Vascular endothelium in
diabetes. Am J Physiol Renal Physiol. 312:F266–F275. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sieve I, Münster-Kühnel AK and
Hilfiker-Kleiner D: Regulation and function of endothelial
glycocalyx layer in vascular diseases. Vascul Pharmacol. 100:26–33.
2018. View Article : Google Scholar : PubMed/NCBI
|