Introduction
Osteosarcoma (OS) is a bone tumor that is primarily
observed in children and adolescents (1), making up 2.4% of all pediatric patient
malignancies and ~20% of all types of primary bone cancer (2–4). OS is
the most common type of primary bone malignancy, and normally
affects the long bones of legs and arms (4). Although the therapy of OS has improved
over the past decade, the prognosis of OS remains poor. The main
cause of the poor prognosis is the occurrence of metastases
following surgical resection and intensive chemotherapy (5,6). OS
pathogenesis, progression and prognosis have been revealed to be
regulated by a number of tumor-associated signaling pathways.
However, the detailed molecular mechanisms underlying OS formation
and development remain poorly understood (7,8). In
order to improve the therapy and develop better prognoses for
patients with OS, the potential underlying molecular mechanisms
must be elucidated and new therapeutic targets must be identified
for OS treatment.
MicroRNAs (miRNAs) have been demonstrated to
regulate gene expression via binding to the 3′-untranslated region
(3′-UTR) of their target mRNAs (9).
Previous studies have demonstrated that miRNAs are important cancer
biomarkers and play a key role in cancer cell growth and metastasis
(10,11). Tumor-associated miRNAs can function
as both or either tumor suppressors and oncogenes, depending on the
biological function of the target genes (12,13),
meaning the function of miRNAs in tumors can be two-sided (14). miRNA-150 (miR-150) is a
tumor-associated miRNA, which has been reported as a biomarker in
several different types of cancer, including osteosarcoma (15), gastric cancer (16), non-small cell lung cancer (17). Colorectal cancer (18) and leiomyoma (19). However, the biological function and
the underlying mechanisms of miR-150 in OS have not yet been
investigated.
The runt-related transcription factor 2 (RUNX2) gene
has been suggested to be a cancer-associated gene that is
implicated in chemotherapeutic resistance. RUNX2 is often amplified
and aberrantly expressed in OS, and is the master regulator of
skeletal development, directly regulating cell proliferation,
apoptosis and differentiation in osteoblasts (20). However, in order to evaluate whether
RUNX2 is a viable biomarker and therapeutic target for OS
treatment, it is necessary to investigate the specific biological
role of RUNX2 in OS. Therefore, the aim of the present study was to
investigate the biological function and associated mechanism of
miR-150 in OS doxorubicin (DOX) sensitivity of OS.
Materials and methods
Tissue samples and cell lines
A total of 26 paired OS and adjacent non-tumor
tissues were collected from patients (sex, 15 males and 11 females;
age, 60±5 years) with OS who underwent curative tumor resection
between September 2010 and August 2014 at the Jinling Hospital
(Nanjing, China). The tissues samples were obtained once the
patients had provided written informed consent, and the process was
approved by the Ethics Committee of Jinling Hospital (Nanjing,
China). SAOS2, MG-63, HOS and U2OS human OS cell lines and hFOB1.19
normal osteoblast cells were purchased from the American Type
Culture Collection.
Cell proliferation and cell viability
assay
All cells were cultured in DMEM (Invitrogen; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Sigma-Aldrich;
Merck KGaA) at 37°C with 5% CO2. For plasmid
transfection, cells were first seeded in six-well plates, and
transfection was performed when 80% confluence was achieved. A
total of two 8 µl (500 ng/µl) plasmids (pcDNA3.1-RUNX2, RUNX2-shRNA
(5′-AAAAGCGCATTCCTCATCCCAGTATTTCGATACTGGGATGAGGAATGCGC-3′, Shanghai
GenePharma Co., Ltd.) or miR-150 mimics and NC (hsa-miR-150 mimics,
5′-UCUCCCAACCCUUGUACCAGUG-3′; hsa-miR-150 mimics NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; Shanghai GenePharma Co., Ltd.) and 8
µl Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
were suspended in 100 µl Opti-MEM (Gibco; Thermo Fisher Scientific,
Inc.) and the final concentration of RUNX2 plasmids or mimics used
was 1 mg/ml. Following incubation at room temperature for 30 min,
the mixture was added into cell culture. After 6 h of culture, the
cell medium was replaced by fresh medium. After another 24 h of
culture, the cells were used to detect the rates of proliferation.
The proliferation rates in the different groups of HOS and U2OS
cells (blank, vector and RUNX2 overexpression groups) were detected
via MTT assay. Cells were suspended in 200 µl culture medium in a
96-well plate at 1×104 cells/well. At every 24 h after
cell seeding for 72 h, the culture medium of the 96-well plate was
replaced with DMEM plus 0.5 mg/ml MTT and incubated at 37°C for 4
h. DMEM medium was then replaced with isopycnic DMSO. Following
mixing by extensive shaking, absorbance at 490 nm was detected for
each well using a SpectraMax 190 (Molecular Devices in Sunnyvale)
to measure the optical density value at 0, 24, 48 and 72 h.
Cell Counting Kit-8 (CCK-8) assay
DOX (Selleck Chemicals) was used to construct an OS
cell chemotherapy resistance model, where cell viability was
measured using CCK8 assay. A dose-dependent assay was performed
using 0, 25, 50, 75 and 100 µM DOX treatment at 37°C for 4 h. Since
cell viability did not reduce further on increasing the
concentration of DOX beyond 50 µM, this concentration was chosen
for subsequent experiments. OS cell lines HOS and U2OS were seeded
in 96-well plates with 5,000 cells/well. Following DOX treatment,
cell proliferation was assessed using a CCK-8 assay (Dojindo
Molecular Technologies). Every 24 h for 72 h after seeding, 10 µl
CCK-8 solution was added to the culture medium, and the cultures
were incubated for 30 min at 37°C in 5% CO2. The
absorbance was measured at 450 nm using a Microplate Reader
(Bio-Rad Laboratories, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from the HOS and U2OS cell
lines and tissues using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Reverse
transcription was performed using a Takara PrimeScript™ RT reagent
kit (Takara Bio Inc.) at 37°C for 1 h and 85°C for 15 sec. qPCR was
performed using SYBR Green (Takara Bio Inc.). The thermocycling
conditions were as follows: Initial denaturation at 90°C for 30
sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 34 sec)
with the ABI Prism 7700 Sequence Detection system (Applied
Biosystems; Thermo Fisher Scientifc, Inc.). RUNX2 or miR-150
expression was quantified using the 2−∆∆Cq method
(21). GAPDH and U6 were used as
internal controls of the mRNA or miRNA, respectively. Sequences of
the primers used in the present study were as follows: miR-150
forward, 5′-TGTCGTGGAGTCGGCAATTCAGTTGAGCACTGG-3′ and reverse,
5′-ACACTCCAGCTGGGTCTCCCAACCCTTGTA-3; GAPDH forward,
5′-TGCACCACCAACTGCTTAGC-3′ and reverse,
5′-GGCATGGACTGTGGTCATGAG-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
Luciferase assay
TargetScan bioinformatics algorithm (version 7.2;
http://www.targetscan.org/vert_72/)
was used to identify the potential targets of miR-150. The 293T
cells (Type Culture Collection of the Chinese Academy of Sciences)
were seeded in 24-well plates and cultured until they reached 60%
confluence. After incubation overnight at 37°C, the cells were
co-transfected pmirGLO plasmids encoding RUNX2 wild-type (WT)
3′-UTR or mutant (Mut) 3′-UTR (Shanghai GenePharma Co., Ltd.) and
miR-150 mimics (Shanghai GenePharma Co., Ltd.) or the control
mimics (Shanghai GenePharma Co., Ltd.) using Lipofectamine 2000.
Firefly and Renilla luciferase activities were detected using
Dual-Luciferase Reporter assay system (Promega Corporation) 48 h
after transfection, according to the manufacturer's protocol. All
transfection assays were performed in triplicate.
Western blot analysis
RIPA lysis buffer (Beyotime Institute of
Biotechnology) with protease inhibitor cocktail (Beyotime Institute
of Biotechnology) was used for cell lysis. A BCA Protein Assay kit
(Pierce; Thermo Fisher Scientific, Inc.) was used to detect the
protein concentration. In total, 4 µg all proteins were resolved by
10% SDS-PAGE and transferred onto PVDF membranes. The membranes
were subsequently incubated with blocking buffers, consisting of
10% skimmed milk powder (dissolved in TBS supplemented with 0.1%
Tween-20) for 2 h at room temperature, prior to incubation with the
respective primary antibodies. The primary antibodies used in the
present study were as follows: Anti-RUNX2 (1:1,000; cat. no. D1H7;
Cell Signaling Technology, Inc.), anti-β-actin (1:2,000; cat. no.
20536-1-AP; Proteintech Group, Inc.), anti-caspase-8 (1:1,000; cat.
no. 9746; Cell Signaling Technology, Inc.), anti-caspase-3
(1:1,000; cat. no. 9662; Cell Signaling Technology, Inc.). The
membranes were incubated with the primary antibodies at 4°C
overnight. Subsequently, the membranes were incubated with
horseradish peroxidase conjugated-goat anti-mouse IgG anti-rabbit
secondary antibodies (1:4,000; cat. no. sc-2030; Santa Cruz
Biotechnology, Inc.) for 2 h at room temperature. The membranes
were exposed using an ECL kit (EMD Millipore) and assessed using
Image Lab™ Software 2.0 (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data are presented as the mean ± standard error of
the mean, and experiments were repeated at least three times.
Significance between groups was analyzed by one-way analysis of
variance followed by Student-Newman-Keuls test. Paired Student's
t-test was used to analyze differences between paired samples. SPSS
software (version 17.0; SPSS, Inc.) was used for the data analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-150 is decreased in OS tissues and
cell lines
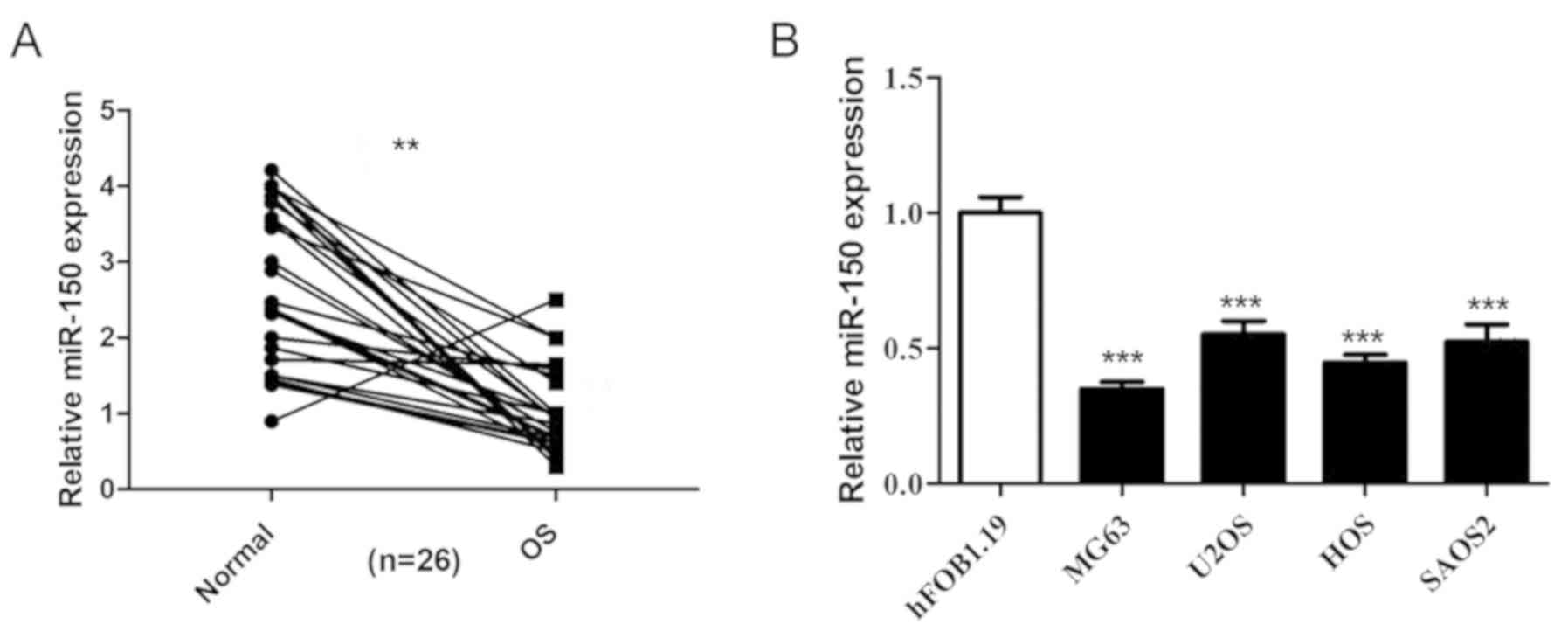
The expression levels of miR-150 in 26 OS tissues
and non-tumor tissues were detected by RT-qPCR, and the results
revealed that miR-150 was significantly decreased in OS tissues
compared with non-tumor tissues (Fig.
1A). The expression levels of miR-150 were also significantly
decreased in the four human OS cell lines (HOS, SAOS2, MG-63 and
U2OS) compared with the normal osteoblast cells (hFOB1.19)
(Fig. 1B), indicating that miR-150
exhibits low expression levels in OS.
miR-150 inhibits OS cell proliferation
and sensitizes OS cells to DOX
In order to investigate the biological role of
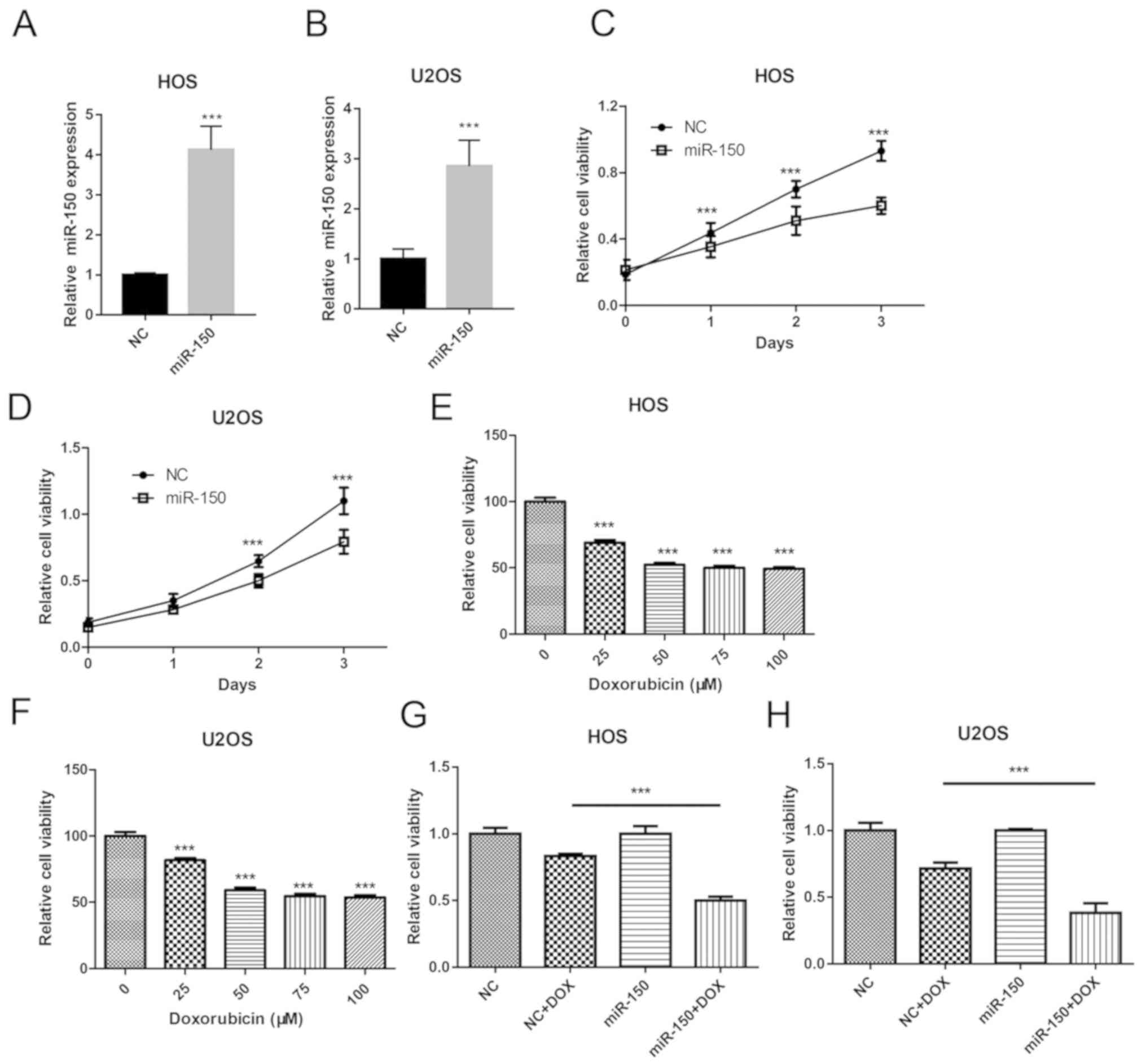
miR-150 in OS tumorigenesis, miR-150 was overexpressed in OS cell
lines HOS and U2OS in the present study (Fig. 2A and B). Cell proliferation was
assessed in order to determine the effects of miR-150 on OS cell
growth using CCK-8. The data revealed that overexpression of
miR-150 significantly inhibited the proliferation of HOS and U2OS
cells (Fig. 2C and D). In order to
investigate the role of miR-150 in OS cell chemotherapy resistance,
an OS cell chemotherapy model was constructed using DOX. Cell
viability was assessed in order to determine the effect of DOX
treatment on OS cell viability. A dose-dependency assay was
performed using 0, 25, 50, 75 and 100 µM DOX treatment. The results
revealed that the cell viability of both HOS (Fig. 2E) and U2OS (Fig. 2F) was significantly decreased with
DOX treatment in a dose-dependent manner. No further reductions in
cell viability was observed when the concentration of DOX exceeded
50 µM. Furthermore, miR-150 overexpression significantly decreased
the cell viability of HOS and U2OS in the DOX treatment group,
compared with the NC mimic + DOX group (Fig. 2G and H). Thus, these data suggested
that miR-150 inhibited OS cell proliferation and sensitized OS
cells to DOX.
RUNX2 is a target gene of miR-150
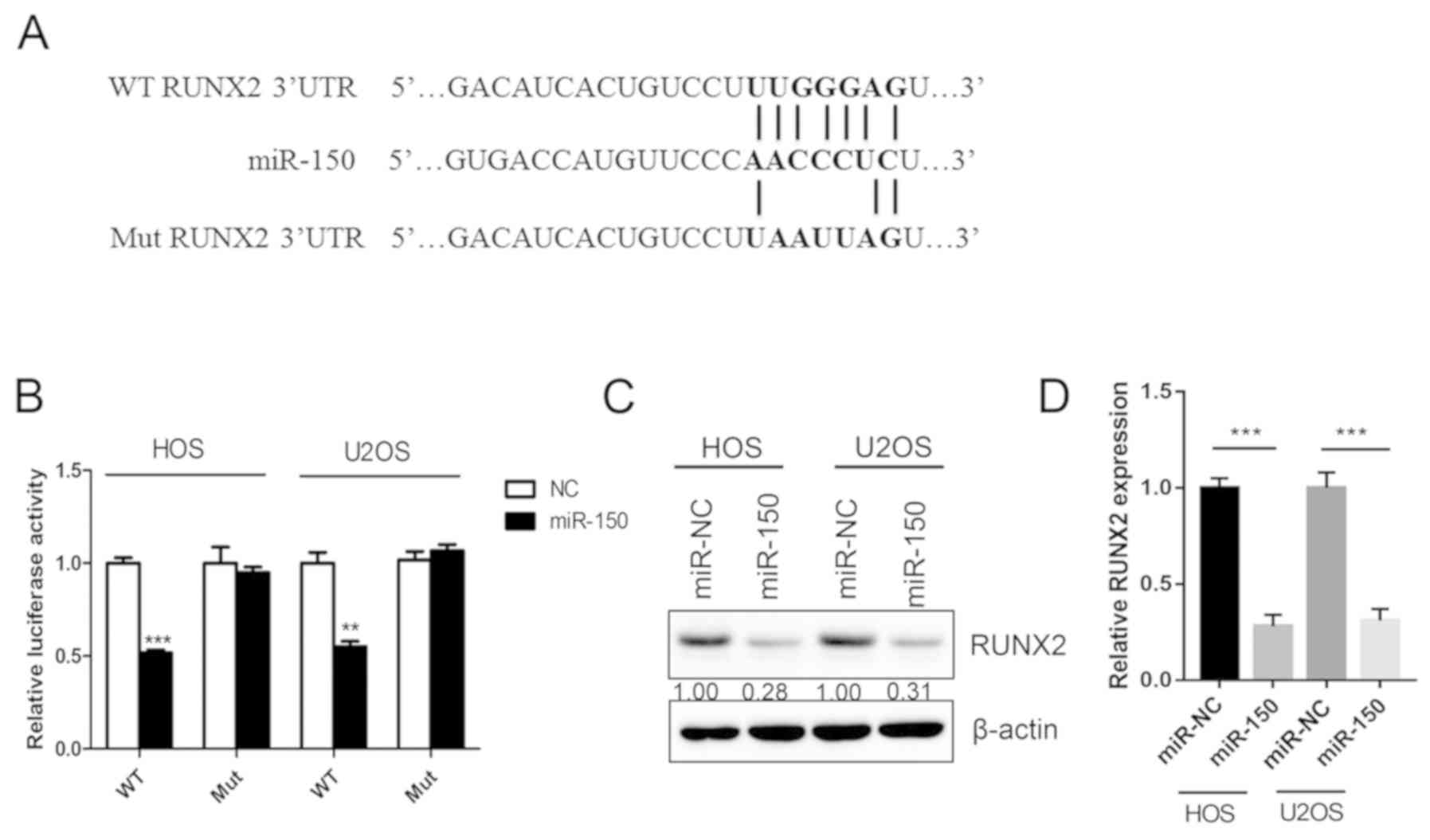
In order to assess the downstream target and
underlying molecular mechanism of miR-150 in OS, a TargetScan
bioinformatics algorithm was used to identify the potential target
gene of miR-150. The results revealed that RUNX2 was a potential
target of miR-150 based on a putative 3′-UTR sequence (Fig. 3A). A luciferase assay was performed
in order to investigate the association between miR-150 and RUNX2.
Luciferase reporter constructs containing the WT or Mut 3′-UTR of
the RUNX2 were constructed. The results revealed that miR-150
significantly decreased the luciferase activity of the WT RUNX2
3′-UTR but not the Mut RUNX2 3′-UTR (Fig. 3B). Overexpression of miR-150
significantly downregulated the expression of RUNX2 both in HOS and
U2OS cells (Fig. 3C and D). The
results indicated that RUNX2 was a target gene of miR-150 in
OS.
RUNX2 is involved in miR-150-induced
OS cell proliferation suppression and chemotherapy
sensitization
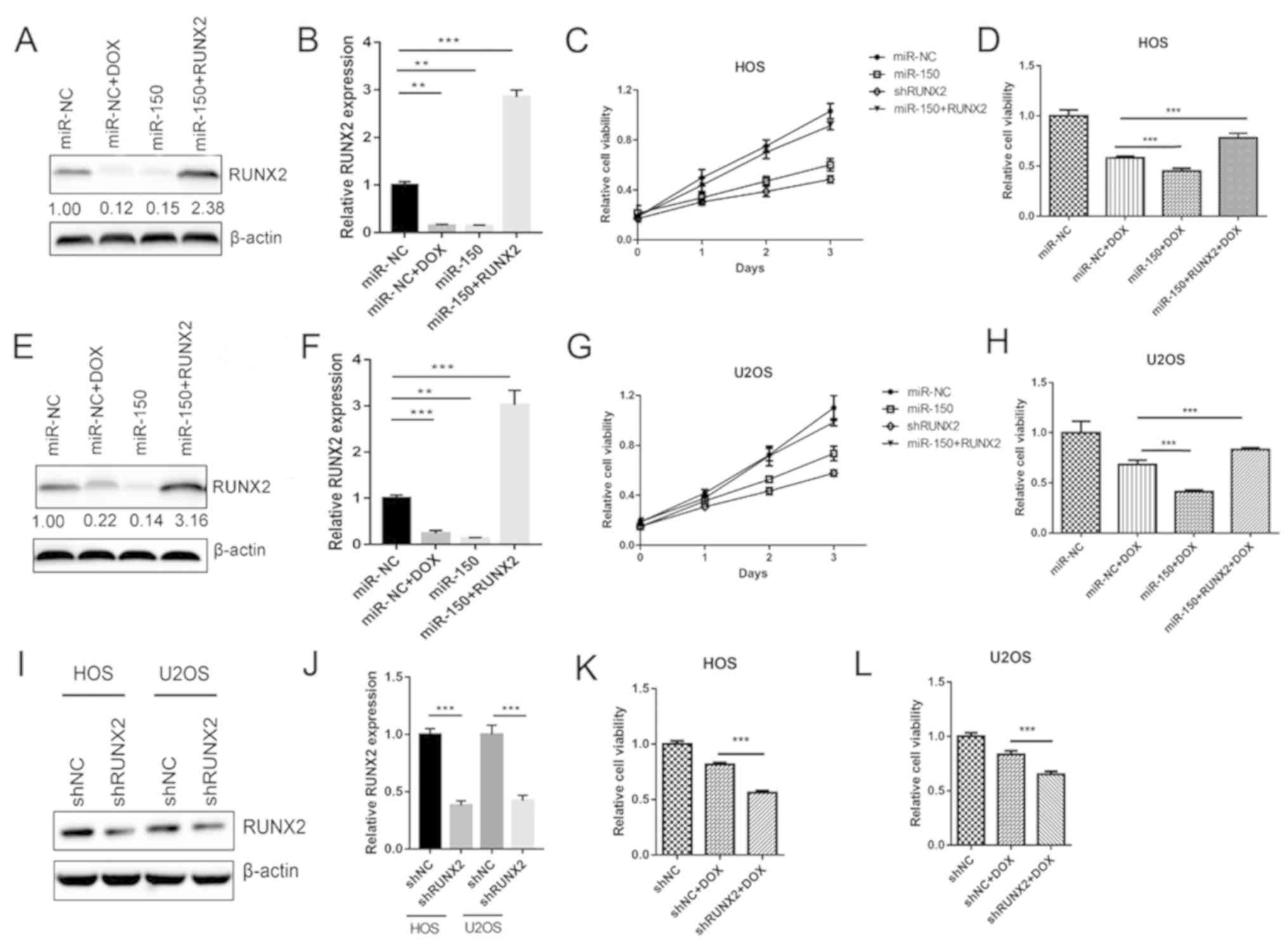
The aforementioned results indicated that miR-150
inhibited OS cell growth by targeting RUNX2. Therefore, the roles
of RUNX2 in OS cell proliferation and chemotherapy resistance were
further detected in the present study. The data revealed that RUNX2
expression was significantly decreased by transfection with miR-150
mimic in both HOS (Fig. 4A and B)
and U2OS (Fig. 4E and F) cells,
compared with the NC mimic group. Treatment of the miR-NC group
with DOX also significantly reduced the expression of RUNX2
(Fig. 4B and F), suggesting that
sensitization of OS to DOX chemotherapy is associated with RUNX2
expression. RUNX2 was then overexpressed using the pcDNA3.1 vector
to rescue RUNX expression. Proliferation assays revealed that the
suppressive effect of miR-150 on HOS (Fig. 4C) and U2OS (Fig. 4G) cells was comparable to that of
shRUNX2. As presented in Fig. 4D and
H, overexpression of RUNX2 reversed the effects of miR-150 on
the DOX sensitization of HOS and U2OS cells, respectively.
Furthermore, following RUNX2 knockdown (Fig. 4I and J), sensitization of HOS and
U2OS cells to DOX was also significantly increased (Fig. 4K and L). Taken together, the results
indicated that RUNX2 was an important functional target of miR-150
involved in the proliferation and chemotherapy sensitization of OS
cells.
miR-150 and RUNX2 affect OS cell
chemosensitivity by regulating apoptosis proteins
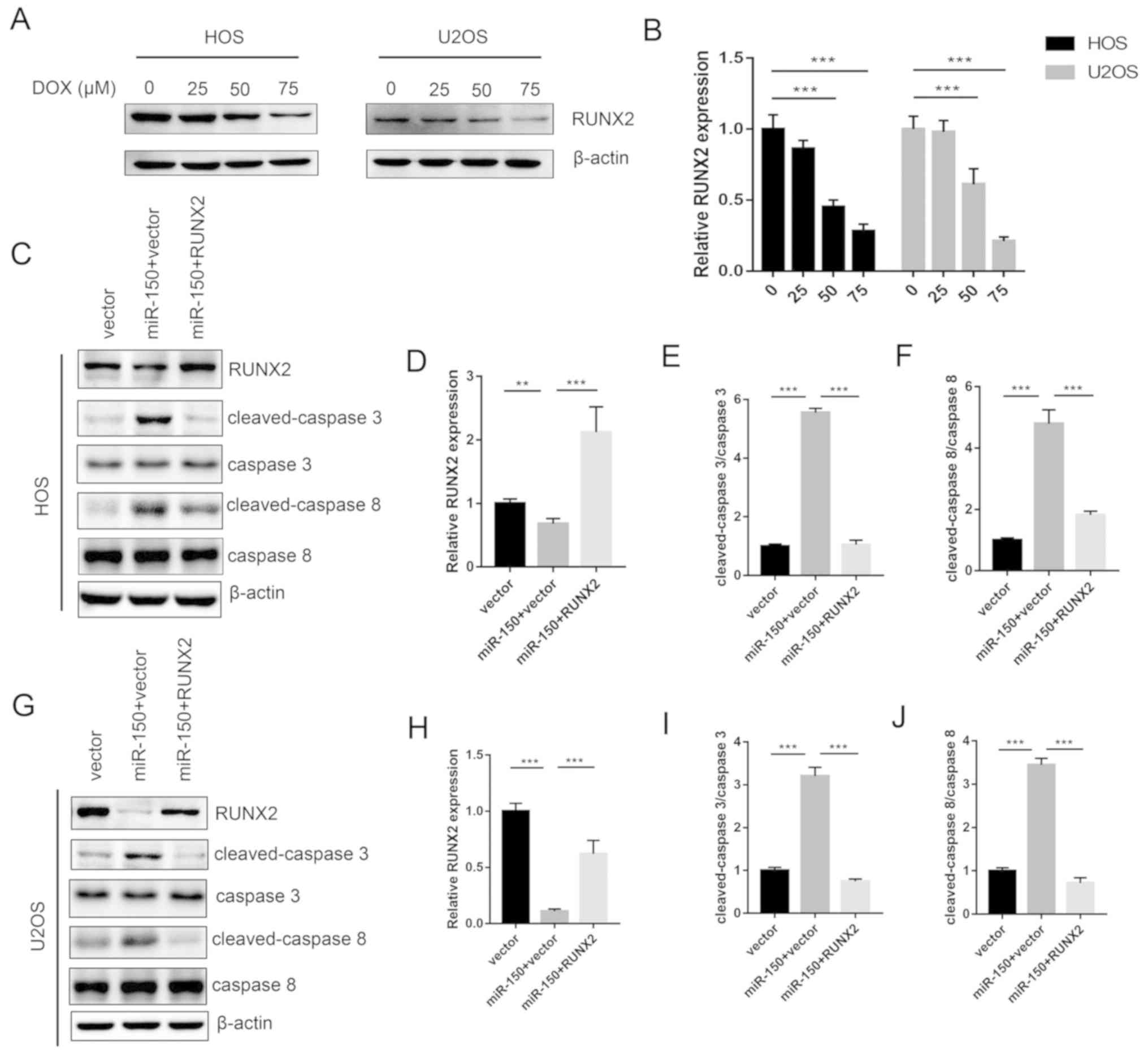
Expression levels of RUNX2 were decreased with
increased concentrations of DOX treatment in HOS and U2OS cells in
a dose-dependent manner (Fig. 5A and
B). The levels of apoptosis proteins, including cleaved
caspase-3 and cleaved caspase-8 were tested. The results revealed
that miR-150 significantly increased the expression of cleaved
caspase-3 and cleaved caspase-8 in both HOS and U2OS cells, while
RUNX2 significantly decreased the expression levels of cleaved
caspase-3 and cleaved caspase-8 (Fig.
5C-J). Therefore, the data from the present study demonstrated
the molecular mechanism underlying the miR-150-RUNX2 axis in OS
resistance to DOX.
Discussion
In the present study, it was revealed that RUNX2
knockdown sensitized OS to the treatment of DOX. In addition, a
novel mechanism for the miR-150-RUNX2 axis was indicated, which
demonstrated that RUNX2 is a target gene of miR-150. miR-150
significantly suppressed cell proliferation and invasion in OS
cells by directly targeting RUNX2. Restoration of RUNX2 in OS cells
reversed the tumor suppressor and chemoprotective roles of miR-150.
In summary, miR-150 functions as a tumor suppressor and sensitizes
OS to chemotherapy-induced apoptosis by targeting RUNX2.
In recent years, miRNAs have been demonstrated as
important cancer biomarkers in a wide range of different types of
cancer (12–17). During cancer progression, miRNAs
regulate the transcription and expression of a large number of
genes, a number of which are vital for the genesis and development
of cancer cells (22,23). There are abundant reports of the
aberrant expression and the function of miRNAs in OS; certain
miRNAs act as oncogenes, while others serve as tumor suppressors
(24,25). miR-150 is also a tumor-associated
miRNA, but the biological function and the underlying molecular
mechanisms of miR-150 in OS have not yet been fully
investigated.
miR-150 has been reported as downregulated in OS
tissues and cells (15). miR-150 has
also been reported as a tumor suppressor by suppressing the
PI3K-AKT pathway (26). miR-150,
insulin like growth factor 2 mRNA binding protein 1 (IGF2BP1) and
combined miR-150/IGF2BP1 expressions were all identified as
independent prognostic factors for overall and disease-free
survival of patients with osteosarcoma (27). In a previous study, exogenous miR-150
expression stimulated cell apoptosis and inhibited proliferation,
invasion and migration (28).
However, to the best of our knowledge, the downstream targets and
details regarding the molecular mechanisms underlying tumor
suppression by miR-150 have not yet been elucidated.
RUNX2 is an important transcription factor, which
regulates the cell fate during normal osteoblast differentiation,
such as cell apoptosis in response to tumor necrosis factor-α
(29,31). Furthermore, RUNX2 has been implicated
as a putative oncogene and a cancer biomarker in OS (32,33).
However, until now, the biological consequences of RUNX2
overexpression and its molecular role in the chemoprotection of OS
had not been clearly defined.
In the present study, a TargetScan bioinformatics
algorithm was used to investigate whether RUNX2 is a target of
miR-150, and it was revealed that miR-150 may bind to RUNX2 3′-UTR.
The subsequent luciferase reporter assay demonstrated that miR-150
could specifically decrease the luciferase activity of the RUNX2
3′-UTR, but not that of the Mut, and that miR-150 could suppress
RUNX2 in OS cell lines, indicating that RUNX2 was a direct target
of miR-150. Proliferation assays revealed that miR-150
overexpression and RUNX2 knockdown exhibited similar inhibiting
effects on OS cell proliferation and chemotherapy resistance.
Furthermore, restoration of RUNX2 reversed the suppressive effects
of miR-150 on OS cell proliferation and chemotherapy treatment.
In the present study, although it was demonstrated
that the miR-150-RUNX2 axis played an important role in OS cell
chemotherapeutic agent resistance, several issues remain to be
investigated. The regulators of the miR-150-RUNX2 axis require
further investigation, and this theory must be verified by
additional clinical experiments. A biological function study of the
network of miR-150-RUNX2 axis would facilitate the pathogenesis
research of human OS. New molecular targeting drugs may be
developed based on this miR-150-RUNX2 axis network.
Taken together, the present study identified miR-150
as a tumor suppressor in OS, and that it inhibits OS growth by
targeting RUNX2. miR-150 can be used as a potential therapy in
future OS treatment.
Acknowledgements
Not applicable.
Funding
This article is supported by the national natural
science foundation of China (grant no. 81802693).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC conceived and designed the present study. YC, JF
and GZ developed the methodology. ZL and GF performed the majority
of the experiments. GF, DY, JZ and YZ performed the data analysis,
ZL wrote the manuscript. All authors approved the final version of
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Jinling Hospital (Nanjing, China). All patients
provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mousa SA, Gallati C, Simone T, Dier E,
Yalcin M, Dyskin E, Thangirala S, Hanko C and Rebbaa A: Dual
targeting of the antagonistic pathways mediated by Sirt1 and TXNIP
as a putative approach to enhance the efficacy of anti-aging
interventions. Aging (Albany NY). 1:412–424. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bielack SS, Marina N, Ferrari S, Helman
LJ, Smeland S, Whelan JS and Reaman GH: Osteosarcoma: The same old
drugs or more? J Clin Oncol. 26:3102–3105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Longhi A, Errani C, De Paolis M, Mercuri M
and Bacci G: Primary bone osteosarcoma in the pediatric age: State
of the art. Cancer Treat Rev. 32:423–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gorlick R and Khanna C: Osteosarcoma. J
Bone Mineral Res. 25:683–691. 2010. View
Article : Google Scholar
|
|
5
|
Bacci G, Briccoli A, Rocca M, Ferrari S,
Donati D, Longhi A, Bertoni F, Bacchini P, Giacomini S, Forni C, et
al: Neoadjuvant chemotherapy for osteosarcoma of the extremities
with metastases at presentation: Recent experience at the Rizzoli
Institute in 57 patients treated with cisplatin, doxorubicin, and a
high dose of methotrexate and ifosfamide. Ann Oncol. 14:1126–1134.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rainusso N, Wang LL and Yustein JT: The
adolescent and young adult with cancer: State of the art e bone
tumors. Curr Oncol Rep. 15:296–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang J and Zhang W: New molecular insights
into osteosarcoma targeted therapy. Curr Opin Oncol. 25:398–406.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang T, Ji F, Dai Z, Xie Y and Yuan D:
Increased expression of microRNA-191 as a potential serum biomarker
for diagnosis and prognosis in human osteosarcoma. Cancer Biomark.
15:543–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Muoio DM: TXNIP links redox circuitry to
glucose control. Cell Metab. 5:412–414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Forrester MT, Seth D, Hausladen A, Eyler
CE, Foster MW, Matsumoto A, Benhar M, Marshall HE and Stamler JS:
Thioredoxin-interacting protein (Txnip) is a feedback regulator of
S-nitrosylation. J Biol Chem. 284:36160–36166. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dreyer F and Baur A: Biogenesis and
functions of exosomes and extracellular vesicles. Methods Mol Biol.
1448:201–216. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang D, Qiu C, Zhang H, Wang J, Cui Q and
Yin Y: Human microRNA oncogenes and tumor suppressors show
significantly different biological patterns: From functions to
targets. PLoS One. 5(pii): e130672010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li CH, Yu TB, Qiu HW, Zhao X, Zhou CL and
Qi C: miR-150 is downregulated in osteosarcoma and suppresses cell
proliferation, migration and invasion by targeting ROCK1. Oncol
Lett. 13:2191–2197. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Quan X, Chen D, Li M, Chen X and Huang M:
MicroRNA-150-5p and SRC kinase signaling inhibitor 1 involvement in
the pathological development of gastric cancer. Exp Ther Med.
18:2667–2674. 2019.PubMed/NCBI
|
|
17
|
Jin M, Shi C, Yang C, Liu J and Huang G:
Upregulated circRNA ARHGAP10 predicts an unfavorable prognosis in
NSCLC through regulation of the miR-150-5p/GLUT-1 axis. Mol Ther
Nucleic Acids. 18:219–231. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Xu X, Pan B, Zeng K, Xu M, Liu X,
He B, Pan Y, Sun H and Wang S: miR-150-5p suppresses tumor
progression by targeting VEGFA in colorectal cancer. Aging (Albany
NY). 10:3421–3437. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JH, Choi YS, Park JH, Kim H, Lee I,
Won YB, Yun BH, Park JH, Seo SK, Lee BS and Cho S: miR-150-5p may
contribute to pathogenesis of human leiomyoma via regulation of the
Akt/p27Kip1 pathway in vitro. Int J Mol Sci. 20(pii): E26842019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sadikovic B, Thorner P, Chilton-Macneill
S, Martin JW, Cervigne NK, Squire J and Zielenska M: Expression
analysis of genes associated with human osteosarcoma tumors shows
correlation of RUNX2 overexpression with poor response to
chemotherapy. BMC Cancer. 10:2022010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Villadsen SB, Bramsen JB, Ostenfeld MS,
Wiklund ED, Fristrup N, Gao S, Hansen TB, Jensen TI, Borre M,
Ørntoft TF, et al: The miR-143/-145 cluster regulates plasminogen
activator inhibitor-1 in bladder cancer. Br J Cancer. 106:366–374.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chiyomaru T, Tatarano S, Kawakami K,
Enokida H, Yoshino H, Nohata N, Fuse M, Seki N and Nakagawa M:
SWAP70, actin-binding protein, function as an oncogene targeting
tumor-suppressive miR-145 in prostate cancer. Prostate.
71:1559–1567. 2011.PubMed/NCBI
|
|
24
|
Takagi T, Iio A, Nakagawa Y, Naoe T,
Tanigawa N and Akao Y: Decreased expression of microRNA-143 and
−145 in human gastric cancers. Oncology. 77:12–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang M, Lin L, Cai H, Tang J and Zhou Z:
MicroRNA-145 downregulation associates with advanced tumor
progression and poor prognosis in patients suffering osteosarcoma.
Onco Targets Ther. 6:833–838. 2013.PubMed/NCBI
|
|
26
|
Watanabe A, Tagawa H, Yamashita J, Teshima
K, Nara M, Iwamoto K, Kume M, Kameoka Y, Takahashi N, Nakagawa T,
et al: The role of microRNA-150 as a tumor suppressor in malignant
lymphoma. Leukemia. 25:1324–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang L, Aireti A, Aihaiti A and Li K:
Expression of microRNA-150 and its Target Gene IGF2BP1 in human
osteosarcoma and their clinical implications. Pathol Oncol Res.
25:527–533. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li X, Chen L, Wang W, Meng FB, Zhao RT and
Chen Y: MicroRNA-150 inhibits cell invasion and migration and is
downregulated in human osteosarcoma. Cytogenet Genome Res.
146:124–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ghali O, Chauveau C, Hardouin P, Broux O
and Devedjian JC: TNF-alpha's effects on proliferation and
apoptosis in human mesenchymal stem cells depend on RUNX2
expression. J Bone Miner Res. 25:1616–1626. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Olfa G, Christophe C, Philippe L, Romain
S, Khaled H, Pierre H, Odile B and Jean-Christophe D: RUNX2
regulates the effects of TNFalpha on proliferation and apoptosis in
SaOs-2 cells. Bone. 46:901–910. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nathan SS, Pereira BP, Zhou YF, Gupta A,
Dombrowski C, Soong R, Pho RW, Stein GS, Salto-Tellez M, Cool SM
and van Wijnen AJ: Elevated expression of Runx2 as a key parameter
in the etiology of osteosarcoma. Mol Biol Rep. 36:153–158. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pereira BP, Zhou Y, Gupta A, Leong DT,
Aung KZ, Ling L, Pho RW, Galindo M, Salto-Tellez M, Stein GS, et
al: Runx2, p53, and pRB status as diagnostic parameters for
deregulation of osteoblast growth and differentiation in a new
pre-chemotherapeutic osteosarcoma cell line (OS1). J Cell Physiol.
221:778–788. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee DH, Qi J, Bradner JE, Said JW, Doan
NB, Forscher C, Yang H and Koeffler HP: Synergistic effect of JQ1
and rapamycin for treatment of human osteosarcoma. Int J Cancer.
136:2055–2064. 2015. View Article : Google Scholar : PubMed/NCBI
|