Introduction
Atherosclerosis (AS) is a chronic inflammatory
disease characterized by the formation of plaques in the walls of
large and medium-sized arteries (1).
Unstable atherosclerotic plaques-especially those with scarce
smooth muscle cells (SMCs) and increased inflammation and
apoptosis-are closely related to numerous diseases, including acute
coronary syndrome, myocardial infarction and stroke (2). Notwithstanding the numerous available
therapeutic alternatives, AS remains the leading cause of mortality
worldwide. Therefore, the development of novel strategies to
prevent the formation and rupture of plaques has become a paramount
area of research.
Macroautophagy (hereafter referred to as autophagy)
is an evolutionarily conserved mechanism which eliminates various
intracellular components such as damaged proteins, lipid droplets
and organelles. Accumulating evidence suggests autophagy has an
important role in the pathogenesis of AS and related disorders
(3–5). Basal autophagy maintains cellular lipid
homeostasis (6–9) and the induction of autophagy has been
observed to prevent AS (10–13). However, previous research shows
conflicting results concerning the impact of the inhibition of
autophagy on AS with some studies indicating impaired autophagy
promotes this process (14–18) and others suggesting otherwise. For
example, Chen et al (19)
found inhibition of autophagy in endothelial cells provided
anti-atherosclerotic effects. Similarly, Dai et al (20) found 3-methyladenine (3-MA), a widely
used inhibitor of autophagy, hindered the formation of
atherosclerotic lesions and increased plaque stabilization. Further
research is needed to elucidate the effects and molecular
mechanisms of autophagy in AS, in order to explore potential
therapeutic targets.
SBI-0206965 (hereafter referred to 6965) is a newly
discovered inhibitor of the uncoordinated (Unc) 51-like kinase 1
(ULK1), which plays a pivotal role in autophagy. As the only
conserved serine/threonine kinase in autophagy, ULK1 has become a
very attractive target for therapeutic development (21). 6965 has also been observed to
suppress autophagy induced by inhibition of mammalian target of
rapamycin (22), inhibit
AMP-activated protein kinase (23)
and induce apoptosis (24). In one
of the authors' previous studies (25), it was reported that 6965 regulated
the functionality of granulocytic myeloid-derived suppressor cells
(MDSCs), which have been linked to the development of AS (26).
The present study systemically administered 6965 to
apolipoprotein E deficient (ApoE−/−) mice fed with a
high-fat diet (HFD) and assessed its effect on AS. The current
study found that 6965 promoted the formation of atherosclerotic
lesions and reduced plaque stability. Interestingly, lower levels
of cluster of differentiation (CD)11b+Gr-1+
MDSCs were also detected following treatment with 6965. Subsequent
in vitro experiments revealed 6965 reduced the viability and
promoted the apoptosis of MDSCs in the presence of oxidized
low-density lipoprotein (oxLDL). Adoptive transfer of MDSCs
impaired the development of atherosclerotic plaques in
ApoE−/− mice. In consonance, these results suggest
inhibition of autophagy by 6965 intervenes in the pathophysiology
of AS by decreasing levels of MDSCs, outlining the mechanisms
underlying the role of autophagy in this process.
Materials and methods
Animals
A total of 40 ApoE−/− mice (19–21.5 g)
were purchased from Beijing Vital River Laboratory Animal
Technology Co., Ltd., and kept in a specific pathogen-free
environment. All experiments were conducted according to the
institutional guidelines for animal care and use, and ethical
approval was obtained prior to the start of the study from an
ethics committee of the Jining Medical University (approval number:
2019-FJ-002). The mice were housed at a constant temperature (22°C)
and relative humidity 40–70% under a 12 h dark/12 h light cycle.
All mice had access to water and food ad libitum. To induce
AS, 8-week old male ApoE−/− mice were fed an HFD (0.25%
cholesterol and 15% cocoa butter) for 6 weeks. After one week of
HFD, the mice were randomly divided into two groups (n=6 per
group): The 6965-treated group, which was injected
intraperitoneally with 6965 (Selleck Chemicals), 150 µg/mouse once
per week for 4 weeks; and a control group of ApoE−/− AS
mice injected with PBS following the same pattern. One week after
the last treatment, the mice were sacrificed and the tissues were
harvested for further use. In the experiment with the adoptive
transferring of MDSCs, mice were fed an HFD for 6 weeks. During
this time, mice (n=6 per group) received an intravenous injection
of 6×106 MDSCs or PBS every 10 days in the experimental
and control groups, respectively.
Serum cholesterol levels
Blood samples (110 µl per mouse) were collected by
retro-orbital bleed following general anesthesia (2–4% ether
inhalation anesthesia) according to ethical approvals (27,28).
After 30 min of incubation at room temperature, samples were
centrifuged at 2,775 × g at room temperature for 10 min to obtain
serum, which was stored at −80°C until analysis. Total plasma
cholesterol and total triglyceride levels were determined with
automated enzymatic techniques (7080; Hitachi, Ltd.).
Histopathology and
immunohistochemistry (IHC)
After sacrificing, mice were perfused with PBS via
the left cardiac ventricle and their hearts and aortic roots were
harvested, fixed in 4% paraformaldehyde at room temperature
overnight and embedded with paraffin. Serial paraffin sections of
4–6 µm were dissected longitudinally. At least three sections were
obtained from each aortic root, with 50 µm of separation between
each other. After staining with hematoxylin and eosin (H&E) for
15 and 5 min, respectively (both at room temperature), sections
were blocked using 5% goat serum (Beijing Solarbio Science &
Technology Co., Ltd.) at room temperature for 30 min. Corresponding
sections on separate sliders were then stained for various
antibodies at 37°C for 1 h. The antibodies used included Anti-F4/80
(1:400; cat. no. ab111101; Abcam), rabbit anti-α smooth muscle
actin (1:200; cat. no. bs-10196R; Biosynthesis Biotechnology Co.,
Ltd.) or anti-mouse Gr-1 (1:50; cat. no. 108436; Biolegend). The
Rabbit two-step test kit (cat. no. PV-9001; Origene Technologies,
Inc.) was used for the secondary antibody binding procedure, and
the staining with secondary antibodies was performed at 37°C for 15
min, according to the manufacturer's protocol. Terminal
deoxynucleotidyl transferase-mediated dUTP nick end labelling
(TUNEL) was performed on paraffin sections with a TUNEL kit (Roche
Diagnostics) following the manufacturer's protocol in order to
assess apoptosis. Images were captured using an Olympus
fluorescence/light microscope (IX71; Olympus Corporation) and
positive staining was quantified by computer-assisted
histomorphometry (Image-Pro Plus 6.0, Media Cybernetics, Inc.). To
evaluate vascular obstruction, the ratio of total plaque area to
aortic root cross sectional area was calculated. For each IHC
analysis, it was calculated as the percentage of immunostained area
in relation to the total area. Mean values were calculated from the
corresponding three consecutive sections from each mouse.
Flow cytometry
After sacrificing the mice, blood, bone marrow and
spleen cells were harvested from each group (n=6 per group). Red
blood cells were removed using erythrocyte lysis buffer [Multi
Sciences (Lianke) Biotech, Co., Ltd.]. Then, the remaining cells
were washed with PBS and resuspended in proportions of
1×106 cells per ml. Fluorescently labeled antibodies
against CD11b (FITC-labeled) and Gr-1 (APC-labeled) were purchased
from Biolegend and used at a 1:100 dilution to detect MDSCs
(CD11b+, Gr-1+). Flow cytometry was also
performed to detect the apoptosis of cells stained with the Annexin
V-PI kit (Annexin V-FITC Apoptosis Detection kit; Biouniquer)
according to the manufacturer's protocol.
Adoptive transfer of MDSCs
Bone marrow-derived MDSCs were obtained as described
previously (25). In brief, bone
marrow cells were isolated from the tibias and femurs of
ApoE−/− mice. Then, these were cultured in complete
DMEM, which was supplemented and stimulated with combinations of
granulocyte macrophage colony stimulating factor (GM-CSF) (40
ng/ml, PeproTech, Inc.) and interleukin (IL)-6 (40 ng/ml,
PeproTech, Inc.). The cultures were maintained during 4 days in an
atmosphere humidified with 5% CO2 at 37°C with the
objective of inducing MDSCs. The experimental group of
ApoE−/− mice which underwent adoptive transfer (n=6)
received MDSCs suspended in PBS via intravenous injection. Each
animal received 6×106 cells every 10 days during 6
weeks. The control group was injected with PBS only following the
same schedule.
En face staining analysis
After the mice were sacrificed, the aortas were
harvested from heart to the iliac arteries. Then, they were fixed
in 4% paraformaldehyde overnight at room temperature and stained
with 0.5% Oil Red O (O0625; Sigma-Aldrich; Merck KGaA) for 12 h at
4°C using a light microscope.
CCK-8 assay
Cell viability was tested using Cell Counting Kit-8
(CCK-8) assays (HY-K0301; MedChemExpress) according to the
manufacturer's protocols. Mouse MDSCs were seeded in 96-well cell
culture plates at 5×103 cells per well and divided into
two groups: One subjected to oxLDL (25 µg/ml) treatment only, and
one preincubated with 6965 (10 µM) for 30 min before treatment with
oxLDL. Next, the cells were cultured at 37°C for 24, 48 and 72 h
respectively, and then incubated with 10 µk CCK-8 solution at 37°C
for 1 h. Well absorbance was set to 450 nm. Each treatment was
performed in triplicate.
Statistical analysis
Data were expressed as the mean ± standard
deviation. Unpaired Student's t-test was used to compare data
between different groups. Results were considered significant when
P<0.05. Statistical analysis was performed with GraphPad Prism
5.0 software (GraphPad Software, Inc.).
Results
6965 increases the size of
atherosclerotic plaques independent of lipid levels
To investigate the effects of 6965 on AS, eight-week
old male ApoE−/− mice were fed an HFD for 6 weeks total
in order to induce AS. After one week of HFD, the 6965-treated
group was injected intraperitoneally with 6965 for 4 weeks. One
week after the last treatment, the mice were sacrificed and
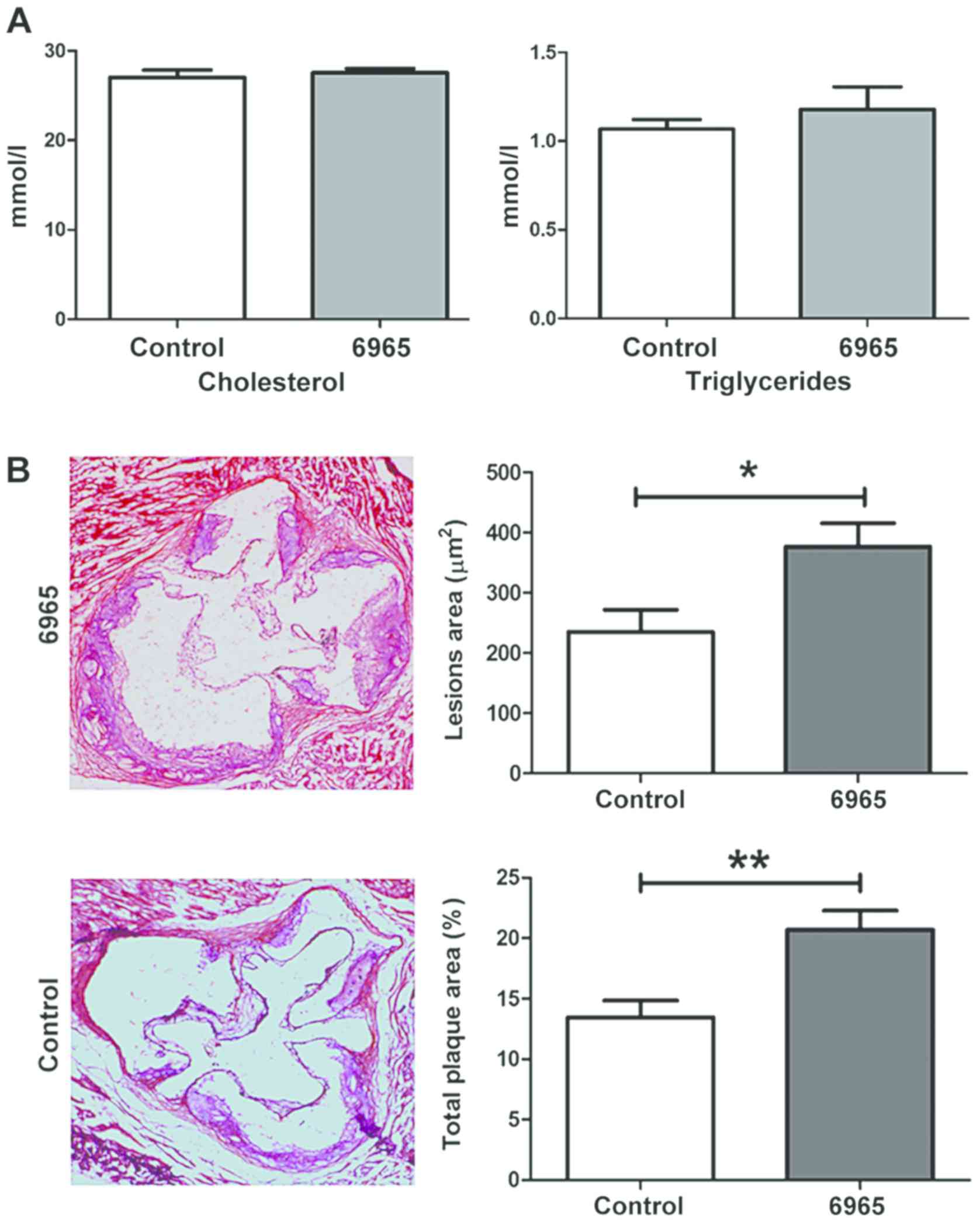
analyzed. As shown in Fig. 1A, 6965
did not affect the lipid profile, including total cholesterol and
triglyceride levels. H&E staining revealed 6965 administration
was associated with significantly greater plaque size on aortic
roots and obvious damage to the adventitia (P<0.05; Fig. 1B). To assess vascular obstruction,
the ratio of total plaque area to aortic root cross-sectional area
was measured, and it was found that treatment with 6965 was related
to significant increases in this parameter in comparison with the
controls. Therefore, the present study surmised that 6965 increased
aortic obstruction and aggravated AS independently of cholesterol
and triglyceride levels.
6965 decreases the stability of
atherosclerotic plaques
Plaque stability rather than size may be more
important in the development of complications associated with AS.
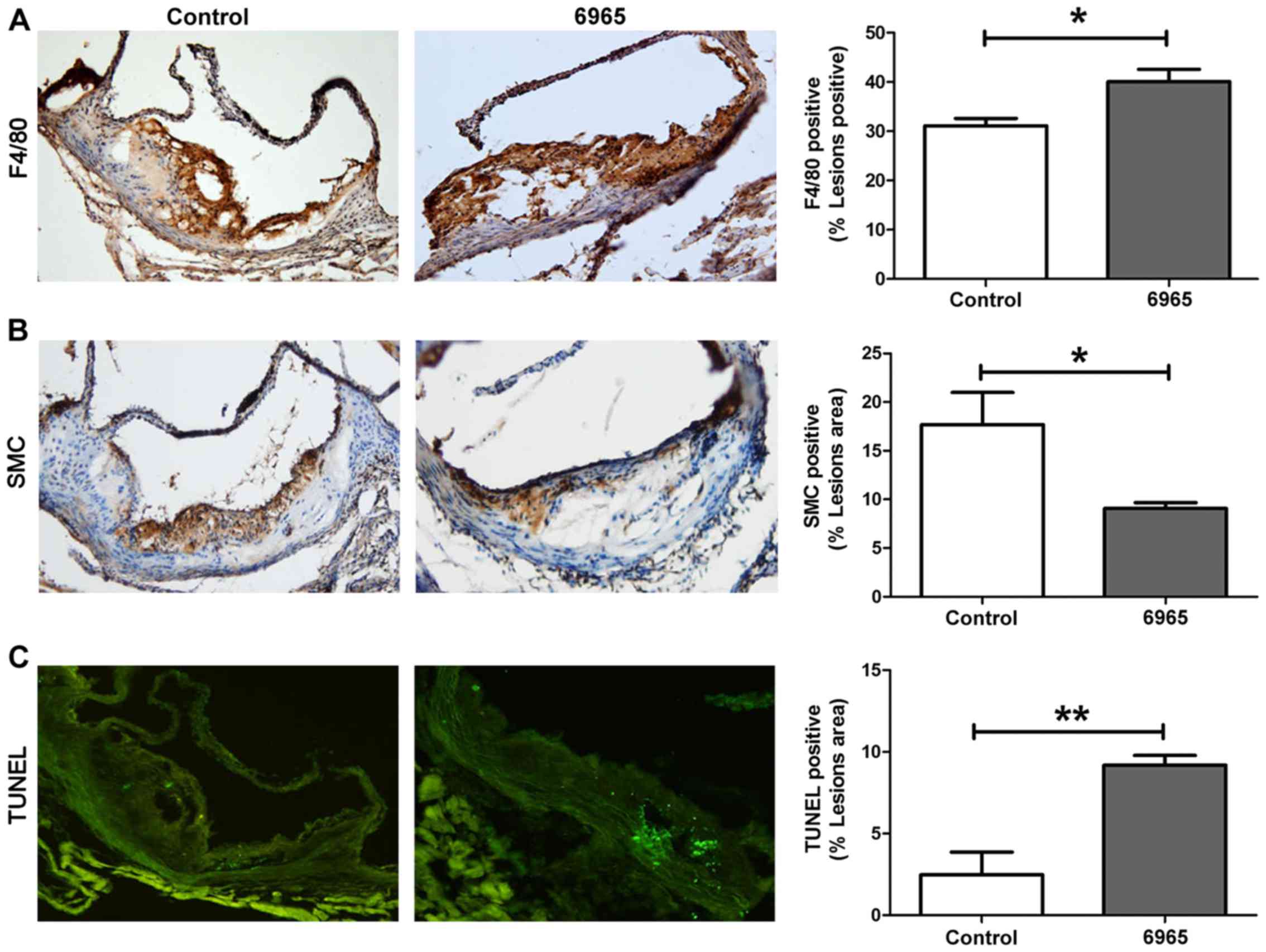
In order to evaluate the effects of 6965 on plaque stability, IHC
staining was implemented to observe the proportions of SMC and
infiltrated macrophages in local lesions; whereas the TUNEL method
was used to assess levels of apoptosis. As shown in Fig. 2, 6965 significantly increased the
proportions of macrophages that presented as F4/80 and decreased
the proportion of SMC in plaques (P<0.05). In addition, 6965
significantly increased the number of TUNEL-positive apoptotic
cells (P<0.01). These results indicate treatment with 6965
reduced plaque stability.
6965 decreases the levels of MDSCs in
atherosclerosis models with ApoE−/− mice
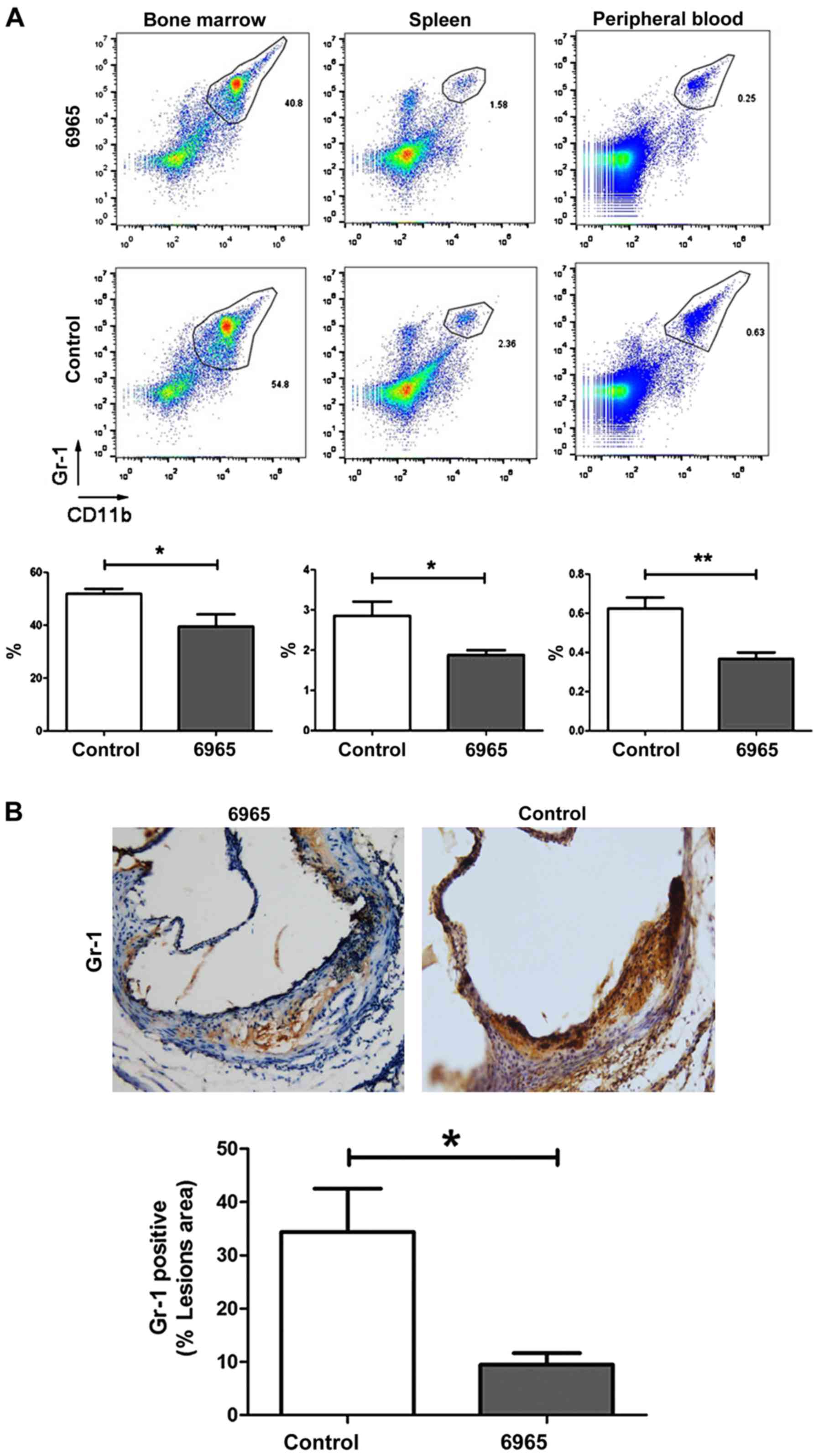
A recent study reported that CD11b+
Gr-1+ MDSCs reduce atherosclerotic plaques in
LDLr−/− mice (26),
suggesting these cells play an important role in the
pathophysiology of AS. In this study, the proportions of
CD11b+ Gr-1+ MDSCs in peripheral blood,
spleen and bone marrow were determined by flow cytometry. The
present study found treatment with 6965 significantly decreased the
proportion of CD11b+ Gr-1+ MDSCs in
comparison with controls. This was the case in the bone marrow, the
spleen and the peripheral blood (P<0.05; Fig. 3A). IHC staining of the lesions showed
Gr-1+ levels were also significantly decreased in
6965-treated mice (P<0.05; Fig.
3B). This suggests 6965 intervenes in the pathogenesis of AS by
lowering the levels of CD11b+ Gr-1+
MDSCs.
Adoptive transfer of MDSCs impairs the
development of atherosclerotic plaques in ApoE−/−
mice
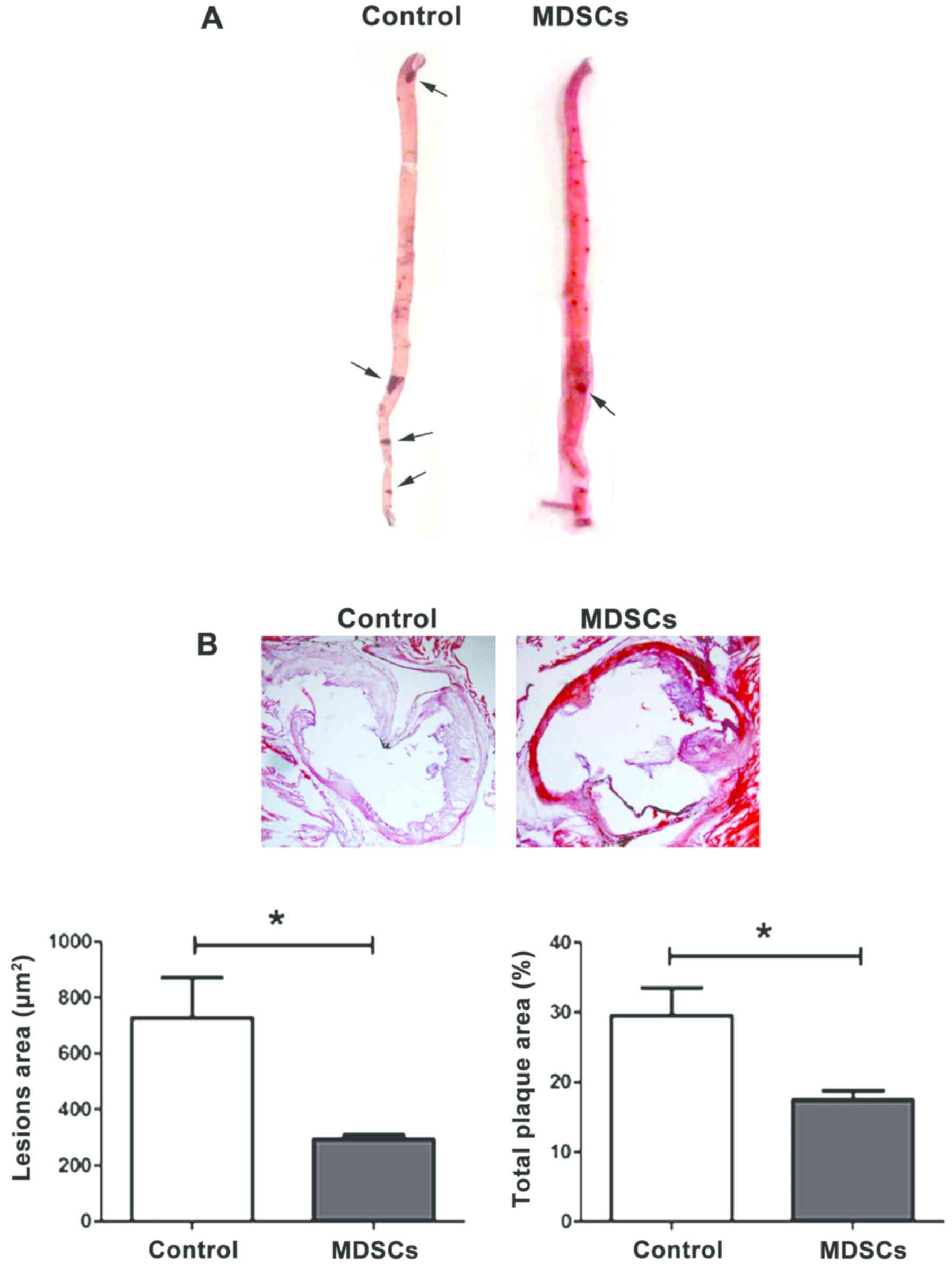
To assess the role of MDSCs in AS, an adoptive
transfer of MDSCs into ApoE−/− mice was performed. MDSCs
were isolated from the bone marrow of ApoE−/− mice
without HFD as described previously (25), and then injected intravenously in
ApoE−/− atherosclerotic models. As expected, a
significant reduction of atherosclerotic lesions in MDSCs-treated
mice in both the aorta (P<0.05; Fig.
4A) and aortic root (P<0.05; Fig.
4B) in comparison with the controls was observed, which did not
undergo adoptive transfer. This confirms the protective effect of
MDSCs on AS in ApoE−/− mice.
6965 reduces the viability and
increases the apoptosis of oxLDL-stimulated MDSCs
In order to investigate the impact of 6965 on MDSCs
in AS, 6965-treated MDSCs from C57 mice were exposed to oxLDL in
vitro. Assessment of CCK-8 levels revealed 6965 reduced MDSCs
viability in the presence of oxLDL (Fig.
5A). Cell Annexin V-PI staining showed 6965 significantly
increased the proportion of apoptosis in the presence of oxLDL in
comparison with exposure to oxLDL alone (P<0.001; Fig. 5B).
Discussion
Modulation of autophagy is an attractive potential
alternative for the treatment of AS; therefore, it is a priority to
focus research on the molecular mechanisms underlying this
relationship in order to assess the feasibility and clinical
applicability of autophagy-regulating agents. In this study, 6965,
a novel inhibitor of ULK1, was found to enhance the formation of
atherosclerotic lesions and reduced plaque stability; reflected in
increased plaque size, reduced SMC content, and increased
proportion of macrophage infiltration and apoptosis. Although a few
studies have associated inhibition of autophagy with
anti-atherosclerotic capacity (19,20),
most studies harmonize with the present conclusions, where
inhibition of autophagy results in aggravation of AS. The
conflicting results may be due to the heterogeneous modes of action
of the various molecules implemented to inhibit autophagy in
previous studies. In this study, 6965, a highly selective inhibitor
of ULK1 was used. This kinase is one of the most upstream
components and the only serine/threonine kinase in the core
autophagy pathway, representing an excellent drug target.
Notably, the proportion of
CD11b+Gr-1+ MDSCs were also found to be
decreased after treatment with 6965. MDSCs are a heterogeneous
population of cells, composed of early myeloid progenitor cells and
immature myeloid cells. MDSCs can block hematoprogenitor cell
differentiation and play a critical role in suppressing immune
responses. Previous research has demonstrated that MDSCs are
important immunosuppressive mediators in cancer and inflammatory
disorders (29–34), including endotoxin shock,
experimental autoimmune encephalomyelitis, graft-vs.-host disease,
rheumatoid arthritis and diabetes. Recently, Foks et al
(26) showed MDSCs reduced AS via
suppression of pro-inflammatory immune responses in LDLr-deficient
mice. To verify the effect of MDSCs on ApoE−/− AS
models, MDSCs were transplanted into ApoE−/− mice, which
resulted in amelioration of the atherosclerotic plaques. In the
present study, it was found that treatment with 6965 was associated
with a decrease in MDSCs levels. Considering MDSCs mainly migrate
to inflamed tissue, Gr1+ proportions were detected in
atherosclerotic lesions and confirmed the decrease of MDSCs in
local plaques in 6965-treated mice. Thus, the decrease of MDSCs may
be a key mechanism underlying the of pro-atherosclerotic effects of
6965.
Numerous studies have identified autophagy as a
major regulator of MDSC viability and function. Wu et al
(35) demonstrated that the
expression of autophagy marker LC3B and p62 was positively
associated with MDSCs number. Parker et al (36) found that autophagy inhibitor
chloroquine or bafilomycin reduced MDSCs viability. However, in
another research, 3-MA did not induce MDSC-like cell model J774M
cell death (37). In the authors'
previous publication (25),
inhibition of autophagy was found to contribute to the accumulation
of granulocytic MDSCs in the context of endotoxin shock. In the
present study, the influence of 6965 on MDSCs in the presence of
oxLDL was appraised. The present results showed inhibition of
autophagy by 6965 decreased MSDC viability and increased their
apoptosis when exposed to oxLDL. It suggested that autophagy might
regulate cell survival depending on the cellular context. Various
conditions and effector molecules linking to autophagy are known to
facilitate MDSC survival. For example, endoplasmic reticulum stress
(ER stress) regulates MDSC half-life by controlling TRAIL receptor
mediated apoptosis (38) and a novel
study found that autophagy protected human umbilical vein
endothelial cells against ER stress-mediated apoptosis (39). It is suggested that inhibition of
autophagy might induce MDSC apoptosis through interruption of ER
stress pathways. Moreover, increased apoptosis and impaired
degradation capacity of lysosomes was observed in
tumor-infiltrating autophagy deficient monocytic-MDSCs (40) which indicates that interruption of
autophagy flux at the late stage induces cell death. Cytokines and
chemokines, such as GM-CSF and IL-6, contributed to the expansion
and accumulation of MDSCs (41). In
the authors' previous study, 6965 treatment reduced the levels of
IL-6, IL-12, tumor necrosis factor-α and CXCL9 in mice, which
suggests that the influence of cytokines of 6965 might also affect
the viability of MDSCs. Although there are numerous possible
mechanisms, an accurate regulatory pathway remains to be
determined.
In conclusion, 6965 may aggravate AS by reducing
MDSCs functionality with a decrease in cell viability and the
promotion of apoptosis being key related mechanisms. The present
findings confirm the importance of autophagy in the pathophysiology
of AS and highlight MDSCs as potential therapeutic targets in this
condition.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Doctoral Fund
of Jining Medical University (grant no. 600353002) and the Teacher
Research Support Fund of Jining Medical University (grant no.
JYFC2018KJ058).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BW and HX designed the experiments. BW, QZ, HZ, ZL
and CL performed animal experiments. GD performed flow cytometry.
QZ and FY performed animal feeding, weighed mice and injected
drugs. JD and BW performed cell culture and CCK-8 assays. CS and QM
analyzed the data. BW wrote the manuscript. FY performed animal
feeding and en face staining. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
The research program was approved by the Ethics
Committee of the Jining Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu Y, Xian X, Wang Z, Bi Y, Chen Q, Han
X, Tang D and Chen R: Research progress on the relationship between
atherosclerosis and inflammation. Biomolecules. 8(pii): E802018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang Q, Hao R, Wang W, Gao H and Wang C:
SIRT1/Atg5/autophagy are involved in the antiatherosclerosis
effects of ursolic acid. Mol Cell Biochem. 420:171–184. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mialet-Perez J and Vindis C: Autophagy in
health and disease: Focus on the cardiovascular system. Essays
Biochem. 61:721–732. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bravo-San Pedro JM, Kroemer G and Galluzzi
L: Autophagy and mitophagy in cardiovascular disease. Circ Res.
120:1812–1824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sasaki Y, Ikeda Y, Iwabayashi M, Akasaki Y
and Ohishi M: The impact of autophagy on cardiovascular senescence
and diseases. Int Heart J. 58:666–673. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grootaert MOJ, Roth L, Schrijvers DM, De
Meyer GRY and Martinet W: Defective autophagy in atherosclerosis:
To die or to senesce? Oxid Med Cell Longev. 2018:76870832018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vion AC, Kheloufi M, Hammoutene A, Poisson
J, Lasselin J, Devue C, Pic I, Dupont N, Busse J, Stark K, et al:
Autophagy is required for endothelial cell alignment and
atheroprotection under physiological blood flow. Proc Natl Acad Sci
USA. 114:E8675–E8684. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sergin I, Evans TD, Zhang X, Bhattacharya
S, Stokes CJ, Song E, Ali S, Dehestani B, Holloway KB, Micevych PS,
et al: Exploiting macrophage autophagy-lysosomal biogenesis as a
therapy for atherosclerosis. Nat Commun. 8:157502017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Evans TD, Jeong SJ, Zhang X, Sergin I and
Razani B: TFEB and trehalose drive the macrophage
autophagy-lysosome system to protect against atherosclerosis.
Autophagy. 14:724–726. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lazaro I, Lopez-Sanz L, Bernal S, Oguiza
A, Recio C, Melgar A, Jimenez-Castilla L, Egido J, Madrigal-Matute
J and Gomez-Guerrero C: Nrf2 activation provides atheroprotection
in diabetic mice through concerted upregulation of antioxidant,
anti-inflammatory, and autophagy Mechanisms. Front Pharmacol.
9:8192018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Zhou Y, Zhang X, Cao X, Wu C and Guo
P: Cordycepin stimulates autophagy in macrophages and prevents
atherosclerotic plaque formation in ApoE−/− mice.
Oncotarget. 8:94726–94737. 2017.PubMed/NCBI
|
|
13
|
Tang F and Yang TL: MicroRNA-126
alleviates endothelial cells injury in atherosclerosis by restoring
autophagic flux via inhibiting of PI3K/Akt/mTOR pathway. Biochem
Biophys Res Commun. 495:1482–1489. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang X, Wei J, He Y, Jing T, Li Y, Xiao Y,
Wang B, Wang W, Zhang J and Lin R: SIRT1 inhibition promotes
atherosclerosis through impaired autophagy. Oncotarget.
8:51447–51461. 2017.PubMed/NCBI
|
|
15
|
Li W, Sultana N, Siraj N, Ward LJ, Pawlik
M, Levy E, Bengtsson E and Yuan XM: Autophagy dysfunction and
regulatory cystatin C in macrophage death of atherosclerosis. J
Cell Mol Med. 20:1664–1672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grootaert MO, da Costa Martins PA, Bitsch
N, Pintelon I, De Meyer GR, Martinet W and Schrijvers DM: Defective
autophagy in vascular smooth muscle cells accelerates senescence
and promotes neointima formation and atherogenesis. Autophagy.
11:2014–2032. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Osonoi Y, Mita T, Azuma K, Nakajima K,
Masuyama A, Goto H, Nishida Y, Miyatsuka T, Fujitani Y, Koike M, et
al: Defective autophagy in vascular smooth muscle cells enhances
cell death and atherosclerosis. Autophagy. 14:1991–2006. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ouimet M, Ediriweera H, Afonso MS,
Ramkhelawon B, Singaravelu R, Liao X, Bandler RC, Rahman K, Fisher
EA, Rayner KJ, et al: microRNA-33 regulates macrophage autophagy in
atherosclerosis. Arterioscler Thromb Vasc Biol. 37:1058–1067. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen PS, Wang KC, Chao TH, Chung HC, Tseng
SY, Luo CY, Shi GY, Wu HL and Li YH: Recombinant thrombomodulin
exerts anti-autophagic action in endothelial cells and provides
anti-atherosclerosis effect in apolipoprotein E deficient mice. Sci
Rep. 7:32842017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dai S, Wang B, Li W, Wang L, Song X, Guo
C, Li Y, Liu F, Zhu F, Wang Q, et al: Systemic application of
3-methyladenine markedly inhibited atherosclerotic lesion in
ApoE−/− mice by modulating autophagy, foam cell
formation and immune-negative molecules. Cell Death Dis.
7:e24982016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zachari M and Ganley IG: The mammalian
ULK1 complex and autophagy initiation. Essays Biochem. 61:585–596.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Egan DF, Chun MG, Vamos M, Zou H, Rong J,
Miller CJ, Lou HJ, Raveendra-Panickar D, Yang CC, Sheffler DJ, et
al: Small molecule inhibition of the autophagy kinase ULK1 and
identification of ULK1 substrates. Mol Cell. 59:285–297. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dite TA, Langendorf CG, Hoque A, Galic S,
Rebello RJ, Ovens AJ, Lindqvist LM, Ngoei KRW, Ling NXY, Furic L,
et al: AMP-activated protein kinase selectively inhibited by the
type II inhibitor SBI-0206965. J Biol Chem. 293:8874–8885. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang F, Hu P, Yang Z, Xue C, Gong J, Sun
S, Shi L, Zhang S, Li Z, Yang C, et al: SBI0206965, a novel
inhibitor of Ulk1, suppresses non-small cell lung cancer cell
growth by modulating both autophagy and apoptosis pathways. Oncol
Rep. 37:3449–3458. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dong G, Si C, Zhang Q, Yan F, Li C, Zhang
H, Ma Q, Dai J, Li Z, Shi H, et al: Autophagy regulates
accumulation and functional activity of granulocytic
myeloid-derived suppressor cells via STAT3 signaling in endotoxin
shock. Biochim Biophys Acta. 1863:2796–2807. 2017. View Article : Google Scholar
|
|
26
|
Foks AC, Van Puijvelde GH, Wolbert J,
Kröner MJ, Frodermann V, Van Der Heijden T, Van Santbrink PJ, Boon
L, Bot I and Kuiper J: CD11b+Gr-1+
myeloid-derived suppressor cells reduce atherosclerotic lesion
development in LDLr deficient mice. Cardiovasc Res. 111:252–261.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakatsu N, Igarashi Y, Aoshi T, Hamaguchi
I, Saito M, Mizukami T, Momose H, Ishii KJ and Yamada H: Isoflurane
is a suitable alternative to ether for anesthetizing rats prior to
euthanasia for gene expression analysis. J Toxicol Sci. 42:491–497.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Akeju O, Hamilos AE, Song AH, Pavone KJ,
Purdon PL and Brown EN: GABAA circuit mechanisms are associated
with ether anesthesia-induced unconsciousness. Clin Neurophysiol.
127:2472–2481. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shaikh F, He J, Bhadra P, Chen X and Siu
SWI: TNF receptor type II as an emerging drug target for the
treatment of cancer, autoimmune diseases, and graft-vs.-host
disease: Current perspectives and in silico search for small
molecule binders. Front Immunol. 9:13822018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Boros P, Ochando J and Zeher M: Myeloid
derived suppressor cells and autoimmunity. Hum Immunol. 77:631–636.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi H, Dong G, Yan F, Zhang H, Li C, Ma Q,
Zhang J, Ning Z, Li Z, Dai J, et al: Arctigenin Ameliorates
inflammation by regulating accumulation and functional activity of
MDSCs in endotoxin shock. Inflammation. 41:2090–2100. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Elliott DM, Singh N, Nagarkatti M and
Nagarkatti PS: Cannabidiol attenuates experimental autoimmune
encephalomyelitis model of multiple sclerosis through induction of
myeloid-derived suppressor cells. Front Immunol. 9:17822018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Casacuberta-Serra S, Costa C, Eixarch H,
Mansilla MJ, López-Estévez S, Martorell L, Parés M, Montalban X,
Espejo C and Barquinero J: Myeloid-derived suppressor cells
expressing a self-antigen ameliorate experimental autoimmune
encephalomyelitis. Exp Neurol. 286:50–60. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hsieh CC, Lin CL, He JT, Chiang M, Wang Y,
Tsai YC, Hung CH and Chang PJ: Administration of cytokine-induced
myeloid-derived suppressor cells ameliorates renal fibrosis in
diabetic mice. Stem Cell Res Ther. 9:1832018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu JS, Li L, Wang SS, Pang X, Wu JB, Sheng
SR, Tang YJ, Tang YL, Zheng M and Liang XH: Autophagy is positively
associated with the accumulation of myeloid-derived suppressor
cells in 4-nitroquinoline-1-oxide-induced oral cancer. Oncol Rep.
40:3381–3391. 2018.PubMed/NCBI
|
|
36
|
Parker KH, Horn LA and Ostrand-Rosenberg
S: High-mobility group box protein 1 promotes the survival of
myeloid-derived suppressor cells by inducing autophagy. J Leukoc
Biol. 100:463–470. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu F, Li X, Lu C, Bai A, Bielawski J,
Bielawska A, Marshall B, Schoenlein PV, Lebedyeva IO and Liu K:
Ceramide activates lysosomal cathepsin B and cathepsin D to
attenuate autophagy and induces ER stress to suppress
myeloid-derived suppressor cells. Oncotarget. 7:83907–83925. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Condamine T, Kumar V, Ramachandran IR,
Youn JI, Celis E, Finnberg N, El-Deiry WS, Winograd R, Vonderheide
RH, English NR, et al: ER stress regulates myeloid-derived
suppressor cell fate through TRAIL-R-mediated apoptosis. J Clin
Invest. 124:2626–2639. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li CF, Pan YK, Gao Y, Shi F, Wang YC and
Sun XQ: Autophagy protects HUVECs against ER stress-mediated
apoptosis under simulated microgravity. Apoptosis. 24:812–825.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Alissafi T, Hatzioannou A, Mintzas K,
Barouni RM, Banos A, Sormendi S, Polyzos A, Xilouri M, Wielockx B,
Gogas H and Verginis P: Autophagy orchestrates the regulatory
program of tumor-associated myeloid-derived suppressor cells. J
Clin Invest. 128:3840–3852. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tamadaho RSE, Hoerauf A and Layland LE:
Immunomodulatory effects of myeloid-derived suppressor cells in
diseases: Role in cancer and infections. Immunobiology.
223:432–442. 2018. View Article : Google Scholar : PubMed/NCBI
|