Introduction
Knee osteoarthritis is a degenerative joint disease
induced by articular cartilage degeneration. Its main clinical
features are arthralgia and joint deformity and dysfunction
(1). It is characterized by a high
incidence and recurrent seizures of the joint and worsens with
time. As the condition advances, prosthetic replacement surgery is
usually required. Over a million people worldwide receive new
diagnoses of articular cartilage disease annually (2) and the condition lowers the quality of
life of these patients (3).
Articular cartilage, an avascular tissue, can be
classified into hyaline cartilage, fibrocartilage and elastic
cartilage. Hyaline cartilage is located on the surface of the knee
joint and has critical functions that differ from the other two
types of cartilage, such as the secretion of lubricating substances
and friction reduction. Hyaline cartilage plays a vital role in
maintaining the normal functions of the knee joint (4).
It is well accepted that knee joint cartilage has a
poor blood supply. Moreover, it lacks stromal stem cells and has an
extremely poor ability for self-repair (5). When cartilage is repaired, it is
usually spontaneously repaired into fibrocartilage, not hyaline
cartilage, therefore losing the extra function of this cartilage
form (6). There are a number of
differences between fibrocartilage and hyaline cartilage in terms
of tissue structure and mechanical properties (7). Articular cartilage defects, the
predominant factor in the pathological process of osteoarthritis,
lead to degeneration of the joint (8).
Much attention has been paid to natural small
molecule compounds based on plant derivatives for the potential
treatment of osteoarticular disease, attributed to the feasibility
of their collection and their bioactivity and therapeutic effects
(9). A previous study indicated
that these compounds may be beneficial in the treatment of
osteoarticular diseases through the regulation of signaling
transduction pathways (10).
In traditional Chinese medicine (TCM), the
pathogenesis of osteoarthritis is thought to involve liver and
kidney deficiencies, as well as malnutrition of the muscles and
bones (11-13).
Drynaria fortunei is a commonly used treatment in TCM which
is thought to benefit the liver and kidneys, strengthen tendons and
bones and target the key treatment points of osteoarthritis.
Drynaria fortunei is thought to be therapeutically
beneficial in the treatment of knee osteoarthritis (14). The present study was designed to
investigate the specific mechanism of action of Drynaria
fortunei, as current knowledge is lacking. Naringin is a plant
derivative and also an active ingredient of Drynaria
fortunei and is thought to have the same effect as the
corresponding dosage of the crude Drynaria fortune (15).
Mesenchymal stem cells (MSCs) are cells with marked
proliferative abilities and the ability to differentiate into
cartilage cells (16). Myeloid
tissue is one of the main sources of MSCs. MSCs that exist in the
bone marrow are termed bone marrow mesenchymal stem cells (BMSCs).
They are one of the most commonly used cells in research into
articular cartilage repair (17). A
number of compounds are thought to exhibit potential for promotion
of the differentiation of BMSCs, such as insulin-like growth factor
I and vascular endothelial growth factor (18).
The transforming growth factor (TGF)-β superfamily
includes TGF-βs and bone morphogenetic proteins. The TGF-β
signaling pathway plays a vital role in cartilage cell
proliferation and differentiation. SOX-9, a downstream factor in
this pathway, plays an indispensable function in articular
cartilage differentiation and is a transcriptional regulator of
chondrogenesis (19). SOX-9 is
necessary to support a normal cartilage cell phenotype (20). Overexpression of SOX-9 results in
increased synthesis of aggrecan, which can provide a suitable
living environment for articular cartilage (21).
Naringin, the active ingredient of Drynaria
fortunei, can be taken orally or injected directly into a
cartilage defect site in combination with BMSCs and has previously
been suggested to stimulate the repair of articular cartilage
defects in rabbit knees (22). The
present study aimed to conduct an in-depth investigation into the
repair if joint structure and cartilage and assess the role of the
TGF-β superfamily signaling pathway in this process. A scoring
system, histological staining and immunohistochemical staining were
used to quantify the efficacy of naringin induced repair and the
potential mechanisms of naringin action were explored (Fig. 1).
Materials and methods
Extraction and separation of rabbit
BMSCs
Following a previously published protocol (23), a 2-week-old New Zealand rabbit was
used for the collection of BMSCs. Rabbits were anesthetized by
intravenous injection of 30 mg/kg pentobarbital sodium into the ear
margin. A bone marrow puncture needle was then inserted into the
posterior superior iliac spine of the rabbit. A total of 20 ml of
low molecular weight heparin calcium (Tianjin Chase Sun
Pharmaceutical Co., Ltd.) was injected to extract the bone marrow.
The inner wall of the syringe was moistened with low molecular
weight heparin 5 min before extraction. The extracted bone marrow
was then injected into a sterile centrifuge tube and was diluted
1:5 with L-DMEM (Beijing Solarbio Science & Technology Co.,
Ltd.). The bone marrow-DMEM mixture was centrifuged at 1,200 x g
for 5 min at 25˚C. The supernatant was discarded and subsequently
cells were resuspended in 20 ml DMEM containing 10% fetal calf
serum (Gibco; Thermo Fisher Scientific, Inc.). The extracted cells
were added to a culture flask and placed in an incubator at 37˚C
and 5% CO2. The medium was replaced with fresh medium 48
h later. The full-marrow adherent method (24) was employed for separation and
purification of rabbit BMSCs. The culture medium was replaced every
3 days until the cells reached 80% confluency, when they were
passaged for further experiments.
Preparation of tested animals
In total, 20 New Zealand rabbits (3-month-old, male)
were selected, with a weight of ~2.5 kg prior to surgery. They were
placed in separate cages and allowed to freely move within the
cage. The animals had access to food and water ad libitum. A
12-h light-dark cycle was maintained. The temperature was
maintained at 21-25˚C and the relative humidity was maintained at
40-70%.
The animals were acclimatized for ≥7 days before
subsequent experiments. The present study followed the National
Institutes of Health Guidelines Regarding the Care and Use of
Laboratory Animals. Animal treatment in the experimental process
conformed to the relevant regulations on ethical standards of
animals specified in the Guiding Proposal for Being Kind to
Experimental Animals issued by the Ministry of Science and
Technology and in Ethical Issues in Animal Experimentation
(11,25). The animal experiments were approved
by the Science and Technology Department of Beijing University of
Chinese Medicine (Beijing, China) and the animal ethics review
committee of the Institute of Basic Theories of Chinese medicine,
Chinese Academy of Chinese Medical Sciences, approval no. 201706058
(Beijing, China).
Animal models
Animal models for articular cartilage defects were
prepared according to a previously described method (22,23).
New Zealand rabbits were purchased from Beijing Changyang Xishan
Experimental Animal Farm. Rabbits were anesthetized using
intravenous administration of 3% sodium pentobarbital (30 mg/kg) in
the ear vein. Knee tissues were cut layer by layer under aseptic
conditions after anesthesia (skin, subcutaneous tissue, muscle,
joint capsule). A corneal trephine (Beijing North Sanyou Medical
Devices Co., Ltd.) was used to drill a hole on the articular
surface of the trochlea from the femoral condyle to the subchondral
bone, with a diameter of 5 mm and a depth of 3 mm. The defect site
was filled according to the experimental scheme. The patella was
reset after completion. The knee joint capsule and skin were
sutured. Penicillin (10x104 U) was injected
intramuscularly postoperatively each day for 3 days. All rabbits
were allowed to move their knee joints freely in their cages
without restriction and clinical signs were observed daily. No
animal was excluded owing to abnormal clinical findings. The
rabbits were anesthetized using intravenous administration of 3%
sodium pentobarbital (30 mg/kg) in the ear vein. Measurement of
anesthetic depth, respiratory rate and pattern and assessment of
mucous membrane color and heart rate monitoring was performed
throughout anesthesia. The animals were sacrificed by an
intravenous air bolus injection into the ear vein containing 30 ml
air, which was injected into the ear margin. Animal death was
confirmed by respiratory arrest, cardiac arrest and changes in
mucosal color. Rabbits in different groups were sacrificed at the
12th week after surgery. There were no statistically significant
differences in body weight among the different groups (Table I).
 | Table INew Zealand white rabbit baseline
information. |
Table I
New Zealand white rabbit baseline
information.
| Parameter | Sham operation
group | Model group | Naringin group | BMSCs group | Naringin+BMSCs
group |
|---|
| Weight, kg | 2.80±0.41 | 2.78±0.17 | 2.83±0.22 | 3.10±0.24 | 3.20±0.34 |
| Sex,
male/female | 4/0 | 4/0 | 4/0 | 4/0 | 4/0 |
| Age, month | 6 | 6 | 6 | 6 | 6 |
Preparation of naringin solution
A naringin solution was prepared with 10 g naringin
and 100 ml deionized water. The gavage concentration (calculated
according to the weight of naringin) was 0.0084 g/kg daily
(26,27).
Laboratory animal grouping and
intervention method
All 20 New Zealand rabbits were randomly distributed
into 5 groups (4 rabbits and 8 knee joints in each group). In the
sham operation group (sham), the joint capsule was cut and seamed,
while the knee joint was not treated. In the model group (Mod),
deionized water was administered via gavage every day after
successful establishment of the articular cartilage defect model.
In the naringin group (Nar), naringin solution was administered via
gavage every day after successful establishment of the articular
cartilage defect model. In the BMSCs group (BMSCs), rabbit BMSCs
were immediately implanted at the joint defect site after
successful establishment of the articular cartilage defect model.
Moreover, deionized water was administered via gavage every day. In
the naringin + BMSCs group (Nar/BMSCs), after successful
establishment of the articular cartilage defect model, BMSCs were
implanted at the defect site immediately and the naringin solution
was administered to the rabbits every day. The daily volume of
deionized water in the Mod and BMSC groups was 0.25 ml. The volume
of gavage in Nar and Nar/BMSCs groups was 0.25 ml.
Macroevaluation
In order to reexamine the intra-articular structure
and to cut out the complete double knee joint of rabbits,
arthrotomy was carried out using the same approach as that used
during modeling. The International Cartilage Repair Society (ICRS)
macroscopic evaluation of cartilage repair (Table II) was adopted to systematically
evaluate the degree of articular cartilage repair in the different
groups (28). The main parameters
monitored were the degree of defect repair, integration to the
border zone and macroscopic appearance. All specimens were
independently evaluated by two professionals with intermediate
professional titles. The investigators who performed the
evaluations were blind to the treatment.
| Table IIInternational Cartilage Repair
Society macroscopic evaluation of cartilage repair. |
Table II
International Cartilage Repair
Society macroscopic evaluation of cartilage repair.
| Category | Score |
|---|
| Degree of defect
repair | |
|
In level
with surrounding cartilage | 4 |
|
75% repair
of defect depth | 3 |
|
50% repair
of defect depth | 2 |
|
25% repair
of defect depth | 1 |
|
No repair of
defect depth | 0 |
| Integration to
border zone | |
|
Complete
integration with surrounding cartilage | 4 |
|
Demarcating
border-1 mm | 3 |
|
Three-quarters
of graft integrated, one-quarter with a notable border-1 mm in
width | 2 |
|
One-half of
graft integrated with surrounding cartilage, one-half with a
notable border-1 mm | 1 |
|
From no
contact, to one-quarter of graft integrated with surrounding
cartilage | 0 |
| Macroscopic
appearance | |
|
Intact
smooth surface | 4 |
|
Fibrillated
surface | 3 |
|
Small,
scattered fissures or cracks | 2 |
|
Several
small or few large fissures | 1 |
|
Total
degeneration of grafted area | 0 |
| Overall repair
assessment | |
|
Grade I,
normal | 12 |
|
Grade II,
nearly normal | 8-11 |
|
Grade III,
abnormal | 4-7 |
|
Grade IV,
severely abnormal | 1-3 |
Histological and immunohistochemical
evaluation
The specimens were fixed in 10% neutral formalin at
room temperature for 24 h. For decalcification the specimens were
placed in EDTA-buffered solution (Beijing Zhongshan Jinqiao
Biotechnology Co. Ltd.) for 4 weeks. The decalcifying solution was
changed twice per week. The articular cartilage of the repaired
area was cut out after obtaining satisfactory decalcification
(Achieved when the syringe needle punctured the tissue without
resistance and the tissue was soft and elastic.), so as to be
trimmed into a sheet with a thickness of 4 mm. Freshly prepared 4%
methanal was used for fixation for 48 h at room temperature,
followed by paraffin embedding and slicing. The specimens were cut
into 4-µm thick slices for histological and immunohistochemical
staining, including hematoxylin and eosin (H&E) staining
(H&E staining kit; Beijing Solarbio Science & Technology
Co., Ltd.) to observe morphological changes in cartilage cells;
toluidine blue staining (Toluidine Blue O cartilage stain solution;
Beijing Solarbio Science & Technology Co., Ltd.) to observe the
secretion of acid mucopolysaccharide and indirectly evaluate the
cartilage matrix; safranin O staining (Safranine O-Fast Green FCF
cartilage stain kit; Beijing Solarbio Science & Technology Co.,
Ltd.) to display the conditions of osteogenesis and chondrogenesis;
type-II collagen immunostaining (rabbit anti-collagen II antibody;
Beijing Bioss Biotechnology Co., Ltd.) to detect specific
indicators of cartilage cells and exhibit the content and quality
of hyaline cartilage. Slices were subsequently blocked in 3%
H2O2 (Beijing Zhongshan Jinqiao Biotechnology
Co. Ltd.) for 10 min at room temperature before staining. Slices
were incubated with rabbit anti-collagen II antibody (Beijing Bioss
Biotechnology Co., Ltd.; cat. no. bs-10589R; 1:200) at 37˚C for 2
h, followed by incubation with horseradish peroxidase conjugated
goat anti-rabbit/mouse secondary antibody (1:1; cat. no. PV6000;
Beijing Zhongshan Jinqiao Biotechnology Co., Ltd) at room
temperature for 30 min. TGF-β3 immunostaining was performed to
evaluate the expression and secretion of TGF-β3. Slices were
incubated with rabbit anti-TGF-β3 antibody (Rabbit anti-TGF-β3
antibody; Beijing Bioss Biotechnology Co., Ltd.; cat. no. bs-1910R;
1:300) at 37˚C for 2 h, followed by incubation with horseradish
peroxidase conjugated goat anti-rabbit secondary antibody (1:1;
cat. no. PV6000; Beijing Zhongshan Jinqiao Biotechnology Co., Ltd)
at room temperature for 30 min. SOX-9 immunostaining was used to
evaluate the expression and secretion of SOX-9. Slices were
incubated rabbit anti-SOX9 antibody (rabbit anti-SOX9 antibody;
Beijing Bioss Biotechnology Co., Ltd.; cat. no. bs-10725R; 1:200)
at 37˚C for 2 h, followed by incubation with horseradish peroxidase
conjugated goat anti-rabbit secondary antibody (1:1; PV6000;
Beijing Zhongshan Jinqiao Biotechnology Co., Ltd.) at room
temperature for 20 min. After staining, the slices were
photographed and examined using an optical microscope (Nikon
Eclipse 600; Nikon Corporation) equipped with a digital camera
(Nikon DXM1200F; Nikon Corporation) to record images of the stained
slices.
A total of 40 slices were prepared from each rabbit
and from each slice three images were captured. All staining
procedures were conducted according to the manufacturers'
instructions. The ICRS Visual Histological Assessment Scale
(Table III) and the modified
O'Driscoll grading system (Table
IV) were utilized to evaluate articular cartilage repair level
in the different groups (29-31).
The main parameters included surface, matrix and cell distribution,
cell population viability, nature of the predominant tissue,
structural characteristics and absence of cellular changes due to
degeneration. All specimens were independently evaluated by two
professionals.
| Table IIIInternational Cartilage Repair
Society visual histological assessment scale. |
Table III
International Cartilage Repair
Society visual histological assessment scale.
| Feature | Score |
|---|
| Surface | |
|
Smooth/continuous | 3 |
|
Discontinuities/irregularities | 0 |
| Matrix | |
|
Hyaline | 3 |
|
Mixture,
hyaline/fibrocartilage | 2 |
|
Fibrocartilage | 1 |
|
Fibrous
tissue | 0 |
| Cell
distribution | |
|
Columnar | 3 |
|
Mixed/columnar-clusters | 2 |
|
Clusters | 1 |
|
Individual
cells/disorganized | 0 |
| Cell population
viability | |
|
Predominantly
viable | 2 |
|
Partially
viable | 1 |
|
<10%
viable | 0 |
| Subchondral
bone | |
|
Normal | 3 |
|
Increased
remodeling | 2 |
|
Bone
necrosis/granulation tissue | 1 |
|
Detached/fracture/callus
at base | 0 |
| Cartilage
mineralization salcified cartilage) | |
|
Normal | 2 |
|
Abnormal/inappropriate
location | 0 |
| Toluidine blue
stain | |
|
Normal | 4 |
|
Slight
reduction | 3 |
|
Moderate
reduction | 2 |
|
Severe
reduction | 1 |
|
No
staining | 0 |
| Percent toluidine
blue in defect | |
|
75-100% | 4 |
|
50-75% | 3 |
|
25-50% | 2 |
|
0-25% | 1 |
|
No toluidine
blue staining | 0 |
| Table IVModified O'Driscoll grading
system. |
Table IV
Modified O'Driscoll grading
system.
| A, Nature of the
predominant tissue |
|---|
| Parameter | Score |
|---|
| Cellular
morphology | |
|
Hyaline
articular cartilage | 4 |
|
Incompletely
differentiated mesenchyme | 2 |
|
Fibrous
tissue or bone | 0 |
| Safranin-O staining
of the matrix | |
|
Normal or
nearly normal | 3 |
|
Moderate | 2 |
|
Slight | 1 |
|
None | 0 |
| B, Structural
characteristics |
| Parameter | Score |
| Surface
regularity | |
|
Smooth and
intact | 3 |
|
Superficial
horizontal lamination | 2 |
|
Fissures,
25-100% of the thickness | 1 |
|
Severe
disruption. including fibrillation | 0 |
| Structural
integrity | |
|
Normal | 2 |
|
Slight
disruption. including cysts | 1 |
|
Severe
disintegration | 0 |
| Thickness | |
|
100 per cent
of normal adjacent cartilage | 2 |
|
50-100 per
cent of normal cartilage | 1 |
|
0-50 per
cent of normal cartilage | 0 |
| Bonding to the
adjacent cartilage | |
|
Bonded at
both ends of graft | 2 |
|
Bonded at
one end, or partially at both ends | 1 |
|
Not
bonded | 0 |
| C, Freedom from
cellular changes of degeneration |
| Parameter | Score |
|
Hypocellularity | |
|
Normal
cellularity | 3 |
|
Slight
hypocellularity | 2 |
|
Moderate
hypocellularity | 1 |
|
Severe
hypocellularity | 0 |
| Chondrocyte
clustering | |
|
No
clusters | 2 |
|
<25 per
cent of the cells | 1 |
|
25-100 per
cent of the cells | 0 |
| Freedom from
degenerative changes in adjacent cartilage | |
|
Normal
cellularity. no clusters, normal staining | 3 |
|
Normal
cellularity, mild clusters, moderate staining | 2 |
|
Mild or
moderate hypocellularity, slight staining | 1 |
|
Severe
hypocellularity. poor or no staining | 0 |
Statistical analysis
SPSS 19.0 software (IBM Corp.) and Prism 7 (GraphPad
Software, Inc.) were used for statistical analysis. Data are
presented as the mean value ± SD. All statistical inferences were
subjected to two-sided tests. P<0.05 was considered to indicate
a statistically significant difference. The confidence interval of
parameter estimation was 95%. Statistical differences were analyzed
using Kruskal-Wallis tests followed by post-hoc testing using
Dunn's multiple comparison tests.
Results
Morphological observation of tissue
repair tissue on the joint defect site
The effect of each treatment on the repair of the
surface structure of the articular cartilage defects was studied on
the basis of morphological observations. ‘Degree of defect repair’,
‘In level with the surrounding cartilage’, ‘Integration to border
zone’, ‘Macroscopic appearance’ markers.
The main findings were as follows: i) Sham group:
There were no surface defects on the knee joint; however, there was
a slight abrasion at the edge (Fig.
2A). ii) Mod group: The defect site displayed some tissue
granulation with an obvious sunken defect area. There were obvious
differences between the areas of normal cartilage tissue and the
areas of regenerated tissue. Moreover, an obvious boundary existed
between the areas of normal tissue and the areas of defective
tissue (Fig. 2B). iii) Naringin
group: At the defect site, tissue was semi-transparent and
granulated. The defect was not completely filled with regenerated
tissue. Compared with the Mod group, the defect area was smaller,
with a reduced depth. The boundary between the defect and the
surrounding cartilage tissue was fuzzy but distinguishable
(Fig. 2C). iv) BMSCs group: Tissue
at the defect appeared transparent, in a similar manner to the
cartilage tissue. When compared with the Mod group, the defect area
was smaller, the depth was shallow, having clearly been reduced and
the boundary appeared fuzzy. Although the thickness of the
regenerated cartilage was similar to that of the surrounding normal
cartilage, the area at the edge of the defect was not as smooth as
that observed in normal cartilage (Fig.
2D). v) Nar/BMSCs group: The repaired defect site was
integrated with the surrounding tissue. Repairing tissue at the
defect site had become milk white, semi-transparent and smooth. It
was hard to distinguish repaired tissue at the defect site from the
surrounding normal cartilage tissue. The defect site had also
developed a smooth surface (Fig.
2E). The characteristic manifestations of the above groups can
also be found in the corresponding images before the index
detection (Fig. 2F-J). In summary
morphological observations in the Naringin group and in the BMSCs
group suggested better tissue healing than in the Mod group.
However, the Nar/BMSCs group showed the most signs of healing from
the defect model animals (Fig.
2).
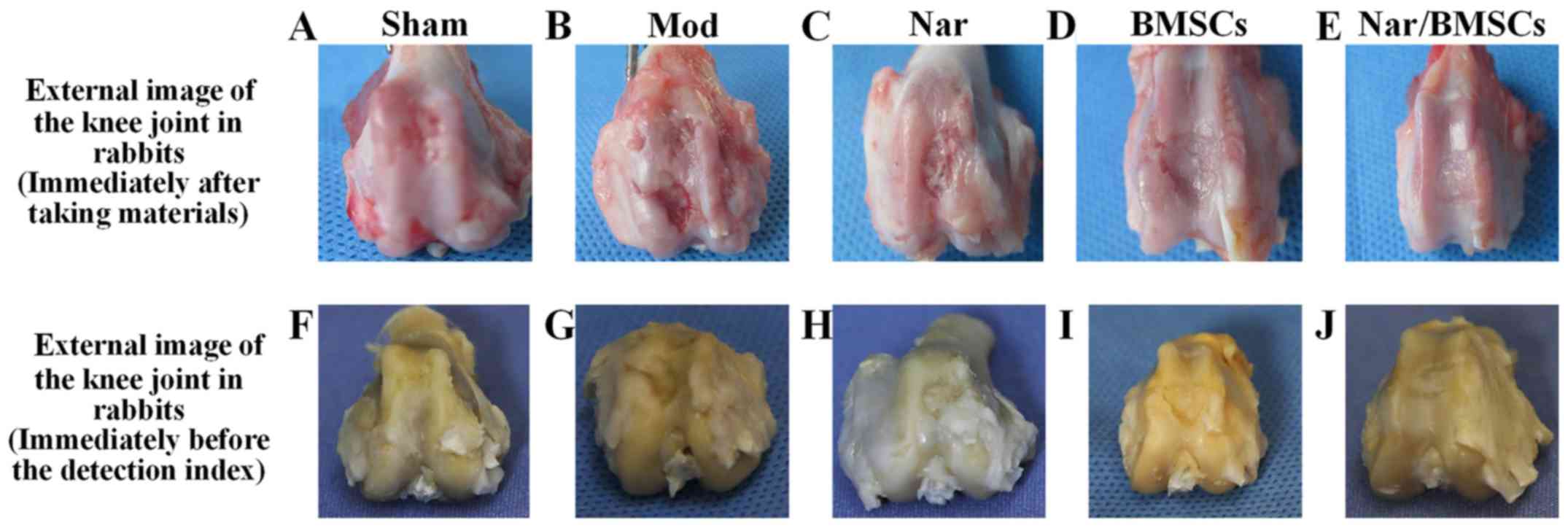 | Figure 2Gross appearance and assessment
results of articular cartilage defects in the rabbit models at 12
weeks. General appearance of rabbit knee joints at 12 weeks
post-operation, including the (A) Sham group, (B) Mod group, (C)
Nar group, (D) BMSCs group and (E) Nar/BMSCs group. General
appearance of rabbit knee joints before the histological and
immunohistochemical assessment, including the (F) Sham group, (G)
Mod group, (H) Nar group, (I) BMSCs group and (J) Nar/BMSCs group.
BMSC, bone marrow mesenchymal stem cells; Mod, model; Nar,
naringin. |
Pathological staining and observation
of the location of repaired tissues at the joint defect site
The repair of joint tissues was observed. The
structural repairs to the articular cartilage defects in the
treatment groups administered BMSCs were semi-transparent and there
were no clear signs of inflammation, however, in the groups that
were not administered BMSCs, articular cartilage defects were not
smooth and the boundaries between new tissue and surrounding
undamaged tissues were readily observed. The treatment groups
administered BMSCs appeared to display improved structural repair
to those that were not administered BMSCs. In terms of the quality
of the repair of articular cartilage defects, in the groups that
were not administered naringin, cartilage damage was obvious, the
hyaline cartilage was severely worn, the thickness of the cartilage
layer was reduced and the tide mark was blurred. However, in the
groups administered naringin, hyaline cartilage appeared at the
site of the cartilage damage, and the tide line was clear, which
was close to the nature of surrounding tissue. The Nar/BMSCs group
was observed to have the most efficient repair regarding structural
and quality repairs of the articular cartilage defects, as shown in
Fig. 3.
 | Figure 3Histological findings of the repair
tissue at articular cartilage defect sites (n=8 knees/group). Scale
bars represent 200 µm. Representative H&E staining at 12 weeks
in the (A) Sham group, (B) Mod group, (C) Nar group, (D) BMSCs
group and (E) Nar/BMSCs group. Toluidine blue staining at 12 weeks
in the (F) Sham group, (G) Mod group, (H) Nar group, (I) BMSCs
group and (J) Nar/BMSCs group. Safranin-O staining at 12 weeks in
the (K) Sham group, (L) Mod group, (M) Nar group, (N) BMSCs group
and (O) Nar/BMSCs group. H&E: Continuous pink on the joint
surface illustrates that the cytoplasm of the cartilage is stained
pink; Toluidine Blue, continuous dark blue on the joint surface
indicates the presence of chondrocytes and cartilage matrix;
Safranin O, continuous red on the joint surface indicates the
cartilage matrix. BMSC, bone marrow mesenchymal stem cells;
H&E, hematoxylin and eosin; Mod, model; Nar, naringin. |
H&E staining was used to observe the cartilage
morphology. Cells in the repaired tissues of the Mod group were
arranged irregularly. Moreover, an obvious gap could be observed
(Fig. 3B). The Nar group was
observed to have poor structural repair at the defect site.
Although the number of repairing cells was limited, the structures
and arrangements of the repairing cells generated at the defect
site were similar to those of the surrounding normal cartilage
cells (Fig. 3C). The BMSCs group
was observed to have an efficient overall contour restoration;
however, the edge of the repairing tissue was irregular, with
obvious gaps (Fig. 3D). The
regenerative tissue of the Nar/BMSCs group appeared smooth.
Differences between regenerative tissues and adjacent normal
cartilage were not clear (Fig. 3E).
The cartilage and cartilage matrix were evaluated using toluidine
blue staining. Cells in the repair tissues of the Mod group were
irregularly arranged and full of fibroblast-like cells, without
signs of repaired deep tissue or the generation of
glycosaminoglycan (GAG; Fig. 3G).
The staining of regenerative tissue in the Nar group was clear, but
repaired tissues were not completely blended with the adjacent
normal cartilage. Although the repair quantity was limited, the
repair quality appeared satisfactory (nuclear staining was clear,
the cartilage matrix was uniformly distributed; Fig. 3H). The thickness of the stained
tissue containing regenerative cells in the BMSCs group was similar
to that of the surrounding cartilage. However, the junction of
repaired tissue and normal cartilage was still distinguishable in
this group (Fig. 3I). The
regenerative tissue in the Nar/BMSCs group stained positive and
appeared homogeneous, similar to that of the adjacent normal
cartilage. The boundary between the normal cartilage and
regenerative cartilage was not clear and the cellular morphology
and distribution in the new cartilage were almost identical to that
of the normal cartilage (Fig.
3J).
Safranin O staining was used to evaluate articular
cartilage repair. Joint cartilage was stained in red. The
structural repair of the defect site of the BMSCs group was
apparent in that Safranin O staining showed a clear structure and
the articular surface was almost smooth. However, the cartilage
matrix was red and discontinuous and the cartilage repair quality
was not complete as the nuclear structure was not clear (Fig. 3N). The articular cartilage
regeneration conditions of the groups administered naringin were
apparent, the collagen content of the cartilage matrix was clear
and there was red staining. The Nar/BMSCs treatment appeared to
demonstrate the best therapeutic effect. The cartilage matrix was a
uniform red and the color of the tissue structure was clear and
uniform in the Nar/BMSCs group. The defect site was occupied by
cartilage cells, which were highly consistent with the surrounding
normal cartilage. Compared with the corresponding position of the
defect in Mod group, the appearance of those areas stained in red
was similar to that of adjacent normal cartilage (Fig. 3O).
Immunohistochemical staining of the
repair tissue on the joint defect site
Type-II collagen immunostaining was used to evaluate
the quality of the cartilage repair in the articular cartilage
defects. The extracellular matrix of hyaline cartilage cells is
mainly composed of type-II collagen and proteoglycan. TGF-β3 and
SOX-9 are key factors in the TGF-β superfamily signaling pathway.
TGF-β3 is crucial in cartilage cell proliferation, differentiation
and other processes. The expression profile of these markers was
evaluated in Fig. 4.
 | Figure 4Immunohistological findings of the
repair tissue at articular cartilage defect sites (n=8
knees/group). Scale bars represent 600 µm. Type II collagen
immunostaining at 12 weeks in the (A) Sham group, (B) Mod group,
(C) Nar group, (D) BMSCs group and (E) Nar/BMSCs group. TGF-β3
immunostaining at 12 weeks in the (F) Sham group, (G) Mod group,
(H) Nar group, (I) BMSCs group and (J) Nar/BMSCs group. SOX-9
immunostaining at 12 weeks in the (K) Sham group, (L) Mod group,
(M) Nar group, (N) BMSCs group and (O) Nar/BMSCs group. Type II
collagen, TGF-β3 and SOX-9 staining: Brown on the joint surface was
positive. BMSC, bone marrow mesenchymal stem cells; Mod, model;
Nar, naringin; TGF, transforming growth factor β. |
In the Sham group, the expression of type-II
collagen was uniform and smooth (Fig.
4A). In the Mod group, there was no significant expression of
type-II collagen, with the defect having a sunken surface and
uneven tissue (Fig. 4B). In
comparison with the Mod group, the BMSCs group presented higher
levels of type-II collagen. The Nar and Nar/BMSCs groups, which
both received naringin treatment, showed more abundant type-II
collagen expression than the group without naringin. The structural
repair of joint defect in the Nar group appeared unsatisfactory,
some of the joint defects were not repaired, but the content of
type-II collagen in the repaired tissue was high, with strong
positive staining. Despite a limited number of cells in the area of
damage, the cells that were present were mainly regenerative cells
of hyaline cartilage (Fig. 4C).
Although a clear structural repair of the joint defect was
exhibited in the BMSCs group, the type-II collagen content of the
repaired tissue was high, with weak positive staining and a limited
quantity of regenerative hyaline cartilage (Fig. 4D). The Nar/BMSCs group displayed the
best therapeutic effect, the expression of type-II collagen was
brownish yellow, and it was superior to other groups in terms of
the uniform distribution of lines on the articular surface.
Meanwhile, the expression of type-II collagen in Nar/BMSCs group
was indicated by strong positive staining and the staining
intensity appeared greater than in other groups. The density of
chondrocytes and cartilage matrix in the Nar/BMSCs group was
similar to that of the normal cartilage. The original defects were
filled with novel hyaline cartilage tissue. A possible reason for
this is that the implanted BMSCs differentiated into chondrocytes
(Fig. 4E).
TGF-β3 and SOX-9 are the main indicators of
activation of the TGF-β superfamily of signaling pathways (32). SOX-9 is regulated and controlled by
the TGF-β superfamily signaling pathway and serves as the main
indicator of downstream signal transmission (33). Areas of positive staining for TGF-β3
in the groups without oral administration of naringin were
relatively weak (Fig. 4G and I),
while they were stronger in the groups with naringin treatment
(Fig. 4J). SOX-9 was uniformly
expressed on the surface of the joint in the Sham group (Fig. 4K). Areas of positive staining for
SOX-9 in the groups without oral administration of naringin were
relatively weak (Fig. 4L). The Nar
group showed more SOX-9 expression than the Mod group (Fig. 4M). Although BMSCs group was
implanted with bone marrow mesenchymal stem cells, the expression
of SOX-9 was lower than in Nar group (Fig. 4N). While they were stronger in the
groups with naringin treatment (Fig.
4O). Notably, the TGF-β3 and SOX9 immunohistochemical staining
reaction in the Nar/BMSCs group was the most obvious in comparison
with that of other groups. The staining was positive in the
Nar/BMSCs group, and distributed evenly in the articular surface.
The repaired articular defect structure was clear. The regenerative
cell matrix of the regenerated hyaline cartilage in the defect
repair site was rich in GAG. The arrangement of chondrocytes tended
to be regular, and the surface of the cartilage was largely smooth.
A satisfactory repair effect can be found from the staining of the
above Nar/BMSCs group. (Fig.
4O).
Two professionals conducted quantitative assessment
of each sample according to: (i) The International Cartilage Repair
Society (ICRS) macroscopic evaluation of cartilage repair (Table II), (ii) The ICRS Visual
Histological Assessment Scale (Table
III) and (iii) the modified O'Driscoll grading system (Table IV). The main parameters assessed
were the degree of defect repair, integration to the border zone
and macroscopic appearance, surface, matrix and cell distribution,
cell population viability, nature of the predominant tissue,
structural characteristics and absence of cellular changes due to
degeneration. Statistical analysis of the scores of the above items
showed that the extent and quality of repair was increased in the
Nar group in comparison to the Mod group; in the Nar/BMSCs group in
comparison to the BMSCs group; in the BMSCs group in comparison to
the Mod group and in the Nar/BMSCs in comparison to the Nar group.
In comparison with the other four groups, the Nar/BMSCs group
exhibited the best quality of repair, as indicated in Fig. 5.
Discussion
Knee osteoarthritis is a common clinical disease
(34). Articular cartilage injury
is the main pathological factor resulting in knee osteoarthritis.
Pathological changes in the morphology of the knee cartilage are
thought to be the most prominent features during osteoarthritis.
Knee cartilage is avascular, aneural and alymphatic, and lacks the
ability to self-repair, thus the repair of articular cartilage
damage is a challenge for clinicians (35). Cartilage cells serve as the main
cellular components of articular cartilage and the articular
cartilage surface is mainly formed by hyaline cartilage, which
helps to maintain the cartilage cells in an undifferentiated state
and to help the cells secrete extracellular matrix (36). The extracellular matrix mainly
consists of two types of macromolecules, type-II collagen and
proteoglycan.
The structure of normal cartilage when visualized
with H&E staining presents a smooth cartilage surface, regular
arrangement and almost round chondrocytes. The cartilage matrix
under toluidine blue staining appears bluish-purple. The cartilage
matrix under safranin O staining appears dark red. The collagen in
cartilage tissue is mainly type II collagen. The brownish yellow
part of type II collagen staining can be used as a specific locator
of cartilage tissue, which indicates a satisfactory repair of
cartilage following injury (37,38).
During the observation of osteoarthritis in the
present study, H&E staining showed unevenly distributed cells,
a decrease in the total cell number, apoptosis of the cells and
uneven staining. The density of toluidine blue staining was uneven,
the staining color was uneven and discoloration was apparent. The
phenomenon of light staining or loss of staining appeared in
safranin O staining and the dyeing area decreased. Type-II collagen
showed a decreased collagen distribution and superficial nuclear
staining.
The BMSCs group displayed typical features of stem
cells, which are undifferentiated cells in their original state.
They possess a self-replicative ability and a multidirectional
differentiation potential, existing in the bone marrow, as well as
in other tissues and organs throughout life, and are responsible
for the repair and updating of the tissue (39). They can be induced to differentiate
into a variety of cells. As the ideal seed cells for cartilage
tissue engineering, these cells been studied from numerous
perspectives (40,41). The BMSCs in this present study
displayed excellent proliferative and differentiation capabilities,
as well as a strong viability. This is consistent with the
characteristics of BMSCs in previous experiments. BMSCs can be
cultured and expanded in order to obtain seed cells, which have
been used to treat cartilage defects in numerous animal models
(42,43). The use of tissue engineering
techniques to study the repair process following cartilage damage
is considered a promising approach (2).
Naringin is a component of the TCM treatment,
Drynaria fortunei, and is thought to be beneficial for the
treatment of osteoarthritis, especially in slowing the occurrence
of inflammation (44). Oral
administration of naringin has been indicate to have the potential
for the treatment of cartilage destruction in osteoarthritis models
(45,46). As a multi-functional substance
(47), naringin has been suggested
to have multiple therapeutic effects, such as the capacity to
inhibit the expression of enzymes which are key mediators in the
development of osteoarthritis, regulate inflammatory protein
secretion and promote cartilage cell infiltration (48,49).
These studies highlight the potential of naringin in the treatment
of osteoarticular degenerative diseases, which may help to restore
the microenvironment of the cartilage defects and improve the state
of the tissue. However, studies investigating the function of
naringin on signaling pathways in osteoarthritis are rare.
Previous studies have clearly indicated that rabbit
BMSCs and naringin solution can be applied to repair rabbit knee
articular cartilage defects. The combination of both can help to
achieve a better repair (20).
However, the specific mechanisms of action remain unknown;
therefore, further investigation into the repair mechanisms would
be of great clinical significance.
Biological processes such as the formation,
metabolism and apoptosis of articular cartilage are regulated by
multiple signaling pathways, among which, the TGF-β superfamily
signaling pathway is prominent in maintenance of the articular
cartilage, through regulating mesenchymal cell migration,
aggregation and differentiation into cartilage cells; and through
regulating the formation of cartilage matrix (50). There are three types of mammalian
TGF-β, namely β1, β2 and β3(51).
The TGF-β superfamily signaling pathway is important in the
development of the growth plate and the formation of permanent
articular cartilage. Previous studies have indicated that TGF-β
plays an important regulatory role in the maintenance of the normal
cartilage cell phenotype, as well as in the promotion of
proliferating cells to differentiate into mature cartilage cells
(52-54).
Similarly, the TGF-β signaling pathway helps to maintain the
cartilage cell phenotype and the stability of the cartilage tissue
environment (55). Not only in
mature cartilage, but also in the process of cartilage formation,
blocking the TGF-β superfamily signaling pathway has been shown to
have harmful effects (56). TGF-β3
has a stronger ability to induce MSCs to differentiate into
cartilage than that of TGF-β1 (57,58).
Therefore, TGF-β3 was selected as one of the main evaluation
indexes of pathway activation in the present study.
SOX-9 can be regulated by the TGF superfamily
signalling pathway. TGF-β can promote the proliferation and
differentiation of cartilage cells by maintaining SOX-9 expression
levels (59). SOX-9 is thought to
be one of the effective transcription factors that are regulated by
type-II collagen and is deemed a modular switch of cartilage
differentiation (60). SOX9 is a
key transcription factor in development and is active during
chondrogenic differentiation, it directly binds to enhancers in the
chondrocyte genome by homodimerizing on pairs of 7-bp SOX-domain
DNA recognition sites (61).
Moreover, it is a key transcription factor of signaling pathways
which induce cartilage differentiation of BMSCs (62). During the cartilage differentiation
process, SOX-9 can be combined with the enhancer element of the
type-II collagen gene to increase the expression of type-II
collagen (63) and to promote the
differentiation of MSCs into chondrocytes (64). The absence of SOX-9 results in the
failure of MSCs to differentiate into cartilage cells, leading to
damage to the cartilage primordium (65). Sufficient SOX-9 expression levels
can not only facilitate the enrichment of MSCs but can also help to
inhibit the conversion of proliferated cartilage cells into a mast
cell phenotype. The maintenance of SOX-9 expression levels highly
depends on the continuous signal stimulation of the TGF-β
superfamily signaling pathway. In summary, Sox-9 has a positive
regulating function on the differentiation and maturation of the
cartilage. Therefore, SOX-9 was selected as one of the downstream
evaluation indexes of pathway activation in the present study
(32,66).
The BMSCs group in the present study was found to
repair the articular cartilage defect of the rabbit knee and the
therapeutic effect on articulation structure repair was deemed
satisfactory; however, the curative effect on the cartilage quality
repair was poor, which was related to the inactivation of the TGF-β
superfamily signaling pathway. Compared with that of the Mod group,
the BMSC group prominently improved the degree of defect repair and
integration to the border zone, the defective joint structure was
restored to a certain degree. In the BMSCs group, BMSCs implanted
at articular cartilage defects may repair the structures of the
joint. However, restoring the structures did not appear to be
sufficient and the quality of the repair was important. The quality
of repair was significantly different between the regenerated
tissue of the BMSCs group and the normal cartilage. The content of
hyaline cartilage in the regenerated tissue was lower than that in
the repaired tissue, which indicated that the BMSCs were not
satisfactory in terms of the repair quality of the articular
cartilage defect.
The expression levels of TGF-β3 and SOX-9 in the Mod
and BMSCs groups were relatively low, which indicated that the
BMSCs did not activate the TGF-β superfamily signaling pathway
during the repair of the articular cartilage defects in rabbit
knees. Naringin, the active ingredient of the TCM treatment
Drynaria fortune (67), was
effective at improving the quality of the repair of the articular
cartilage defects in the rabbit knees, which may be related to the
activation of the TGF-β superfamily signaling pathway. However, its
effect on repairing the defected joint structure was not deemed
satisfactory, which may be attributed to the lack of adequate
repair cells.
Although the Nar group's contour repair for joint
defects was stronger than that of the Mod group, the joint defects
did not demonstrate a satisfactory level of repair to the
structure. This could be attributed to an insufficient number of
repairing seed cells in the articular cartilage defect and could
also be related to a poor intrinsic healing potential (68,69).
Drug treatment alone appears to be unable to significantly promote
the recovery of the joint defect structure. Toluidine blue and
safranin O staining revealed that, compared with that of the Mod
group, the Nar group had significantly improved cartilage quality
in the newly repaired tissue at the defect site. According to the
results of type-II collagen staining, the newly repaired tissue was
composed of predominantly hyaline cartilage. These findings
demonstrated that naringin helped to promote an improved quality of
cartilage regeneration and repair in articular cartilage defects of
the rabbit knees.
The expression levels of TGF-β3 and SOX-9 were
higher in the Nar group compared with those in the Mod group, which
indicated that naringin stimulated the secretion of these two
specific factors. TGF-β3 can act as the chemotactic molecule that
induces chondrogenesis (70). A
study into Sox9 post-translational regulation found that the Sox9
protein levels were closely related to the formation of normal
cartilage (62). Moreover, SOX-9
can be used as a marker of cartilage formation in degenerated
cartilage cells. These data demonstrate that naringin, the
effective ingredient of the TCM treatment Drynaria fortunei,
may be effective at promoting repair of the articular cartilage
defects in rabbit knees through activation of the TGF-β superfamily
signaling pathway.
BMSCs together with naringin, can promote the repair
of articular cartilage defects in rabbit knees. The repair of the
joint structure defects and the quality of the repair of the
articular cartilage were both deemed satisfactory and were found to
be enhanced compared to that of the individual application of BMSCs
or naringin. This may be associated with the simultaneous
activation of the TGF-β superfamily signaling pathway in the
presence of sufficient seed cells.
Both the BMSCs and the Nar/BMSCs groups and
exhibited satisfactory effects on the structural repair of the
joint defects. Regarding the degree of the defect repair and the
association with the surrounding tissues, the Nar/BMSCs group
presented greater improvements than the BMSCs group. The Nar/BMSCs
group displayed better repair effects in articular cartilage
regeneration and cartilage matrix repair as a result of the
treatment with naringin. Regarding type-II collagen, the Nar/BMSCs
group appeared identical to the surrounding normal articular
cartilage tissue and the repaired tissue consisted of a
considerable quantity of type-II collagen. The repaired tissue
maintained a fairly smooth surface profile and its hyaline
cartilage content was higher than that of the BMSCs group. This
indicated that naringin may increase the directional
differentiation of BMSCs into cartilage tissue in the articular
cartilage defect. Furthermore, naringin improved the quality of
cartilage regeneration and repair in articular cartilage defects of
the rabbit knees.
The expression levels of TGF-β3 and SOX-9 were
significantly higher in the Nar/BMSCs group compared to that in the
BMSCs group. Particularly, compared to all other groups in the
present study, the expression levels of TGF-β3 and SOX-9 were
highest in the Nar/BMSCs group. This is consistent with the
conclusion that Sox-9 has the potential to restore degenerated
cartilage structure and functions (71).
The BMSCs implanted in the articular cartilage
defects in the rabbit knees appeared to provide sufficient seed
cells for repair to occur. Oral administration of naringin exerted
a continuous regulatory function on the TGF-β superfamily signaling
pathway, which promoted the directional differentiation of BMSCs
into cartilage tissue at articular cartilage defects in rabbit
knees. Notably, the majority of the differentiated cartilage cells
expressed a hyaline cartilage phenotype.
The present study demonstrated that BMSCs had a
satisfactory effect on the repair of the joint structure of
articular cartilage defects in rabbit knees. Naringin helped to
improve the quality of cartilage repair in articular cartilage
defects in rabbit knees. BMSCs combined with naringin, appeared to
further improve the repair of articular cartilage defects in rabbit
knees, thus delivering satisfactory results for joint structure and
cartilage quality. It appears that the repair mechanism of action
of naringin on articular cartilage defects may be through the
continuous regulation of the TGF-β superfamily signaling pathway
and that combined application with BMSCs may produce better results
than the use of naringin alone. These findings may be attributed to
the directional differentiation of BMSCs into cartilage cells after
regulation of the TGF-β superfamily signaling pathway.
The present study is a preliminary study focused on
articular cartilage defect repair of rabbit knees using a
combination of naringin and BMSCs. Future research will investigate
the effects of different concentrations of naringin on articular
cartilage defect repair in rabbit knees, the target tissue of
naringin (such as chondrocytes or cartilage matrix) and the direct
effects of naringin on cartilage cells. Furthermore, the direct
functional mechanism of action of naringin on the TGF-β superfamily
signaling pathway and the mechanism of naringin's influence on
BMSCs should be further investigated.
In conclusion, naringin, an effective ingredient of
the TCM Drynaria fortunei, and BMSCs were able to stimulate
the repair of articular cartilage defects in rabbit knees. The
combined application of these treatments on joint structural repair
and articular cartilage repair quality delivered improved results,
which may be closely associated with sustained positive regulation
of the TGF-β superfamily signaling pathway and with the directional
differentiation of BMSCs into cartilage cells.
Acknowledgements
The authors would like to thank Professor Mingjing
Zhao (Key Laboratory of Chinese Internal Medicine of Ministry of
Education and Beijing, Dongzhimen Hospital, Beijing University of
Chinese Medicine, Beijing, China) for technical assistance with
this study.
Funding
This study was supported by the Fundamental Research
Funds for the Central Universities (grant nos. 2015-JYB-JSMS058,
2018-JYBZZ-JS091).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PL conceived and designed the study. CY, JC, HL, JY
and FW performed the animal surgeries. YL, CY and PL performed the
histological and immunohistochemical staining. CY, JC, HL, JY, ZY
and YQ acquired the data. CY, JC, ZY and PL analyzed and
interpreted data. CY and JC drafted the manuscript. PL and ZY
critically revised the manuscript for important intellectual
content. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Science
and Technology Department of Beijing University of Chinese Medicine
(Beijing, China) and the Animal Ethics Review Committee of the
Institute of Basic Theories of Chinese Medicine, Chinese Academy of
Chinese Medical Sciences (Beijing, China) approval no.
201706058.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhu Y, Yuan M, Meng HY, Wang AY, Guo QY,
Wang Y and Peng J: Basic science and clinical application of
platelet-rich plasma for cartilage defects and osteoarthritis: A
review. Osteoarthritis Cartilage. 21:1627–1637. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Browne JE and Branch TP: Surgical
alternatives for the treatment of articular cartilage lesions. J Am
Acad Orthop Sur. 8:180–189. 2000.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Katagiri H, Mendes LF and Luyten FP:
Definition of a critical size osteochondral knee defect and its
negative effect on the surrounding articular cartilage in the rat.
Osteoarthritis Cartilage. 25:1531–1540. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang AT, Feng Y, Jia HH, Zhao M and Yu H:
Application of mesenchymal stem cell therapy for the treatment of
osteoarthritis of the knee: A concise review. World J Stem Cells.
11:14–27. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ebihara G, Sato M, Yamato M, Mitani G,
Kutsuna T, Nagai T, Ito S, Ukai T, Kobayashi M, Kokubo M, et al:
Cartilage repair in transplanted scaffold-free chondrocyte sheets
using a minipig model. Biomaterials. 33:3846–3851. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Armiento AR, Alini M and Stoddart MJ:
Articular fibrocartilage-Why does hyaline cartilage fail to repair?
Adv Drug Deliv Rev. 146:289–305. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bae DK, Yoon KH and Sang JS: Cartilage
healing after microfracture in osteoarthritic knees. Arthroscopy.
22:367–374. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wang Y, Yuan M, Guo QY, Lu SB and Peng J:
Mesenchymal stem cells for treating articular cartilage defects and
osteoarthritis. Cell Transplant. 24:1661–1678. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Che CT, Wong MS and Lam CW: Natural
products from chinese medicines with potential benefits to bone
health. Molecules. 21(239)2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
An J, Yang H, Zhang Q, Liu C, Zhao J,
Zhang L and Chen B: Natural products for treatment of osteoporosis:
The effects and mechanisms on promoting osteoblast-mediated bone
formation. Life Sci. 147:46–58. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Su YX, Yan H, Chen BJ, Zahn Q, Wang YR, Lu
ML, Wang WT, He Z and Sheng L: Effect of naringin of Drynaria
Rhizome, a Chinese medical component of Zhuanggu Jianxi recipe
containing serum on Caveolin-p38MAPK signal pathway in IL-1β
induced rabbit degenerated chondrocytes. Zhongguo Zhong Xi Yi Jie
He Za Zhi. 34:1492–1498. 2014.PubMed/NCBI(In Chinese).
|
|
12
|
Lo PC, Lin FC, Tsai YC and Lin SK:
Traditional Chinese medicine therapy reduces the risk of total knee
replacement in patients with knee osteoarthritis. Medicine
(Baltimore). 98(e15964)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Teixeira J, Santos MJ, Matos LC and
Machado JP: Evaluation of the effectiveness of acupuncture in the
treatment of knee osteoarthritis: A case study. Medicines (Basel).
5(18)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
An J, Hao D, Zhang Q, Chen B, Zhang R,
Wang Y and Yang H: Natural products for treatment of bone erosive
diseases: The effects and mechanisms on inhibiting
osteoclastogenesis and bone resorption. Int Immunopharmacol.
36:118–131. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen R, Qi QL, Wang MT and Li QY:
Therapeutic potential of Naringin: An overview. Pharm Biol.
54:3203–3210. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Farrell MJ, Shin JI, Smith LJ and Mauck
RL: Functional consequences of glucose and oxygen deprivation on
engineered mesenchymal stem cell-based cartilage constructs.
Osteoarthritis Cartilage. 23:134–142. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wolf D and Wolf AM: Mesenchymal stem cells
as cellular immunosuppressants. Lancet. 371:1553–1554.
2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Santo VE, Gomes ME, Mano JF and Reis RL:
Controlled release strategies for bone, cartilage, and
osteochondral engineering-Part II: Challenges on the evolution from
single to multiple bioactive factor delivery. Tissue Eng Part B
Rev. 19:327–352. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Cha BH, Kim JH, Kang SW, Do HJ, Jang JW,
Choi YR, Park H, Kim BS and Lee SH: Cartilage tissue formation from
dedifferentiated chondrocytes by codelivery of BMP-2 and SOX-9
genes encoding bicistronic vector. Cell Transplant. 22:1519–1528.
2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Paul R, Haydon RC, Cheng H, Ishikawa A,
Nenadovich N, Jiang W, Zhou L, Breyer B, Feng T, Gupta P, et al:
Potential use of Sox9 gene therapy for intervertebral degenerative
disc disease. Spine (Phila Pa 1976). 28:755–763. 2003.PubMed/NCBI
|
|
21
|
Tew SR, Pothacharoen P, Katopodi T and
Hardingham TE: SOX9 transduction increases chondroitin sulfate
synthesis in cultured human articular chondrocytes without altering
glycosyltransferase and sulfotransferase transcription. Biochem J.
414:231–236. 2008.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Li PY, Li CG, Ye C, Qu Y, Chen J, Zhu GQ
and Zhao H: Experimental study on Naringin decoction combined with
bone marrow mesenchymal stromal stem cells in repair of articular
cartilage defects in rabbits. J Liaoning Univ Tradit Chinese Med.
16:34–38. 2014.
|
|
23
|
Li CG, Qu Y, Ye C, Chen J, Wang FX, Li PY,
Li SH, Ren JP and Qi J: Histology research on repairing of rabbit
articular cartilage defects with Naringin and tissue engineering
cartilage. Chin J Tissue Eng Res. 18:3165–3171. 2014.
|
|
24
|
Cai YT, Liu TS, Fang F, Xiong CL and Shen
SL: Comparisons of mouse mesenchymal stem cells in primary adherent
culture of compact bone fragments and whole bone marrow. Stem Cells
Int. 2015(708906)2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Parodi AL: Ethical issue in animal
experimentation. Bull Acad Natl Med. 193:1737–1745. 2009.PubMed/NCBI(In French).
|
|
26
|
Wei W, Wu XM and Li YJ: Pharmacological
Experimental Methodology. 4th edition. People's Medical Publishing
House, pp69-73, 2010.
|
|
27
|
Gao XM: Science of Chinese Materia Medica.
2nd edition. China Press of Traditional Chinese Medicine,
pp335-336, 2007.
|
|
28
|
Smith GD, Taylor J, Almqvist KF, Erggelet
C, Knutsen G, Garcia Portabella M, Smith T and Richardson JB:
Arthroscopic assessment of cartilage repair: A validation study of
2 scoring systems. Arthroscopy. 21:1462–1467. 2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
van den Borne MP, Raijmakers NJ, Vanlauwe
J, Victor J, de Jong SN, Bellemans J and Saris DB: International
Cartilage Repair Society. International Cartilage Repair Society
(ICRS) and Oswestry macroscopic cartilage evaluation scores
validated for use in Autologous Chondrocyte Implantation (ACI) and
microfracture. Osteoarthritis Cartilage. 15:1397–1402.
2007.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Mainil-Varlet P, Aigner T, Brittberg M,
Bullough P, Hollander A, Hunziker E, Kandel R, Nehrer S, Pritzker
K, Roberts S, et al: Histological assessment of cartilage repair: A
report by the histology endpoint committee of the international
car-tilage repair society (ICRS). J Bone Joint Surg Am. 85A (Suppl
2):S45–S57. 2003.PubMed/NCBI
|
|
31
|
O'Driscoll SW, Keeley FW and Salter RB:
The chondrogenic potential of free autogenous periostal grafts for
biological resurfacing of major full-thickness defects in joint
surfaces under the influence of continuous passive motion. An
experimental investigation in the rabbit. J Bone Joint Surg Am.
68:1017–1035. 1986.PubMed/NCBI
|
|
32
|
Coricor G and Serra R: TGF-β regulates
phosphorylation and stabilization of Sox9 protein in chondrocytes
through p38 and Smad dependent mechanisms. Sci Rep.
6(38616)2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ni Q, Lu K, Li J, Tan Y, Qin J, Magdalou
J, Chen L and Wang H: Role of TGFβ signaling in maternal
ethanol-induced fetal articular cartilage dysplasia and adult onset
of osteoarthritis in male rats. Toxicol Sci. 164:179–190.
2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Johnson K, Zhu S, Tremblay MS, Payette JN,
Wang J, Bouchez LC, Meeusen S, Althage A, Cho CY, Wu X and Schultz
PG: A stem cell-based approach to cartilage repair. Science.
336:717–721. 2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Bekkers JE, Creemers LB, Tsuchida AI, van
Rijen MH, Custers RJ, Dhert WJ and Saris DB: One-stage focal
cartilage defect treatment with bone marrow mononuclear cells and
chondrocytes leads to better macroscopic cartilage regeneration
compared to microfracture in goats. Osteoarthritis Cartilage.
21:950–956. 2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Caron MM, Emans PJ, Coolsen MM, Voss L,
Surtel DA, Cremers A, van Rhijn LW and Welting TJ:
Redifferentiation of dedifferentiated human articular chondrocytes:
Comparison of 2D and 3D cultures. Osteoarthritis Cartilage.
20:1170–1178. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Yu F, Zeng H, Yu HY, Lei M, Yuan H and
Xiao DM: Comparison of different special staining techniques of
chondrocytes and their application values. Chinese J Comp Med.
8:58–61. 2015.
|
|
38
|
Yu F, Lei M, Zeng H, Cao JF and Xiao DM:
Different special staining methods on cartilage tissue of
osteoarthritis morphology: A comparison study. Orthop J China.
19:79–85. 2015.
|
|
39
|
Chamberlain G, Fox J, Ashton B and
Middleton J: Concise review: Mesenchymal stem cells: Their
phenotype, differentiation capacity, immunological features, and
potential for homing. Stem Cells. 25:2739–2749. 2010.
|
|
40
|
Kock L, Donkelaar CCV and Ito K: Tissue
engineering of functional articular cartilage: The current status.
Cell Tissue Res. 347:613–627. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Caplan AI: Review: Mesenchymal stem cells:
Cell-based reconstructive therapy in orthopedics. Tissue Eng.
11:1198–1211. 2005.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Anraku Y, Mizuta H, Sei A, Kudo S,
Nakamura E, Senba K, Takagi K and Hiraki Y: The chondrogenic repair
response of undifferentiated mesenchymal cells in rat
full-thickness articular cartilage defects. Osteoarthritis
Cartilage. 16:961–964. 2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Kurth TB, Dell'Accio F, Crouch V, Augello
A, Sharpe PT and De Bari C: Functional mesenchymal stem cell niches
in adult mouse knee joint synovium in vivo. Arthritis Rheumatol.
63:1289–1300. 2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Manna K, Das U, Das D, Kesh SB, Khan A,
Chakraborty A and Dey S: Naringin inhibits gamma radiation-induced
oxidative DNA damage and inflammation, by modulating p53 and NF-κB
signaling pathways in murine splenocytes. Free Radical Res.
49:422–439. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Zhao Y, Zhong L, Wang W, Zhang H, Chen J,
Su P, Liu L and Li W: Naringin protects against cartilage
destruction in osteoarthritis through repression of NF-κB signaling
pathway. Inflammation. 39:385–392. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Xu Q, Zhang ZF and Sun WX: Effect of
naringin on monosodium iodoacetate-induced osteoarthritis pain in
rats. Med Sci Monitor. 23:3746–3751. 2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Cao X, Lin W, Liang C, Zhang D, Yang F,
Zhang Y, Zhang X, Feng J and Chen C: Naringin rescued the
TNF-α-induced inhibition of osteogenesis of bone marrow-derived
mesenchymal stem cells by depressing the activation of NF-кB
signaling pathway. Immunol Res. 62:357–367. 2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Kawaguchi K, Maruyama H, Hasunuma R and
Kumazawa Y: Suppression of inflammatory responses after onset of
collagen-induced arthritis in mice by oral administration of the
Citrus flavanone naringin. Immunopharm Immunot. 33:723–729.
2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Ahmad SF, Zoheir KM, Abdelhamied HE,
Ashour AE, Bakheet SA, Attia SM and Abd-Allah AR: Amelioration of
autoimmune arthritis by Naringin through modulation of T regulatory
cells and Th1/Th2 cytokines. Cell Immunol. 287:112–120.
2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ashraf S, Cha BH, Kim JS, Ahn J, Han I,
Park H and Lee SH: Regulation of senescence associated signaling
mechanisms in chondrocytes for cartilage tissue regeneration.
Osteoarthritis Cartilage. 24:196–205. 2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Bi. 21:659–693. 2005.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Dong R, Ying J, Xu TT, Hu SF, Zhang P, Xia
CJ, Fang L, Jin HT and Wang PG: Bushenhuoxue formula facilitates
articular cartilage repair and attenuates matrix degradation by
activation of TGF-β signaling pathway. Evid Based Complement
Alternat Med. 2018(2734581)2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Patil AS, Sable RB and Kothari RM: An
update on transforming growth factor-β (TGF-β): Sources, types,
functions and clinical applicability for cartilage/bone healing. J
Cell Physiol. 226:3094–3103. 2011.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Grimaud E, Heymann D and Rédini F: Recent
advances in TGF-beta effects on chondrocyte metabolism. Potential
therapeutic roles of TGF-beta in cartilage disorders. Cytokine
Growth Factor Rev. 13:241–257. 2002.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Blaney Davidson EN, van der Kraan PM and
van den Berg WB: TGF-beta and osteoarthritis. Osteoarthritis
Cartilage. 15:597–604. 2007.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Li YP, Wei XC, Zhou JM and Wei L: The
age-related changes in cartilage and osteoarthritis. Biomed Res
Int. 2013(916530)2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Mackay AM, Beck SC, Murphy JM, Barry FP,
Chichester CO and Pittenger MF: Chondrogenic differentiation of
cultured human mesenchymal stem cells from marrow. Tissue Eng.
4:415–428. 1998.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Lu CH, Lin KJ, Chiu HY, Chen CY, Yen TC,
Hwang SM, Chang YH and Hu YC: Improved chondrogenesis and
engineered cartilage formation from TGF-β3-expressing
adipose-derived stem cells cultured in the rotating-shaft
bioreactor. Tissue Eng Part A. 18:2114–2124. 2012.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Furumatsu T, Tsuda M, Taniguchi N, Tajima
Y and Asahara H: Smad3 induces chondrogenesis through the
activation of SOX9 via CREB-binding protein/p300 recruitment. J
Biol Chem. 280:8343–8350. 2005.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Sha'ban M, Cassim SO, Yahya NM, Saim A and
Idrus R: Sox-9 Transient transfection enhances chondrogenic
expression of osteoarthritic human articular chondrocytes in vitro:
Preliminary analysis. J Tissue Eng Regen. 8:32–41. 2011.
|
|
61
|
Liu CF and Lefebvre V: The transcription
factors SOX9 and SOX5/SOX6 cooperate genome-wide through
super-enhancers to drive chondrogenesis. Nucleic Acids Res.
43:8183–8203. 2015.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Kawakami Y, Rodriguez-León J and Izpisúa
Belmonte JC: The role of TGFbetas and Sox9 during limb
chondrogenesis. Curr Opin Cell Biol. 18:723–729. 2006.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Lefebvr V, Huang W, Harley VR, Goodfellow
PN and De CB: SOX9 is a potent activator of the
chondrocyte-specific enhancer of the pro alpha1(II) collagen gene.
Mol Cell Biol. 17:2336–2346. 1997.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Wang Z, Li K, Sun H, Wang J, Fu Z and Liu
M: Icariin promotes stable chondrogenic differentiation of bone
marrow mesenchymal stem cells in self-assembling peptide nanofiber
hydrogel scaffolds. Mol Med Rep. 17:8237–8243. 2018.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Yoon BS, Ovchinnikov DA, Yoshii I, Mishina
Y, Behringer RR and Lyons KM: Bmpr1a and Bmpr1b have overlapping
functions and are essential for chondrogenesis in vivo. Proc Natl
Acad Sci USA. 102:5062–5067. 2005.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Lefebvre V and Dvir-Ginzberg M: SOX9 and
the many facets of its regulation in the chondrocyte lineage.
Connect Tissue Res. 58:2–14. 2017.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Lee YE, Liu HC, Lin YL, Liu SH, Yang RS
and Chen RM: Drynaria fortune J. Sm. improves the bone mass
of ovariectomized rats through osteocalcin-involved endochondral
ossification. J Ethnopharmacol. 158:94–101. 2014.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Hunziker EB, Lippuner K, Keel MJ and
Shintani N: An educational review of cartilage repair: Precepts and
practice - myths and misconceptions - progress and prospects.
Osteoarthritis Cartilage. 23:334–350. 2015.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Huey DJ, Hu JC and Athanasiou KA: Unlike
bone, cartilage regeneration remains elusive. Science. 338:917–921.
2012.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Hara ES, Ono M, Hai PT, Sonoyama W, Kubota
S, Takigawa M, Matsumoto T, Young MF, Olsen BR and Kuboki T:
Fluocinolone acetonide is a potent synergistic factor of
TGF-β3-associated chondrogenesis of bone marrow-derived mesenchymal
stem cells for articular surface regeneration. J Bone Miner Res.
30:1585–1596. 2015.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Tew SR, Li Y, Pothacharoen P, Tweats LM,
Hawkins RE and Hardingham TE: Retroviral transduction with SOX9
enhances re-expression of the chondrocyte phenotype in passaged
osteoarthritic human articular chondrocytes. Osteoarthritis
Cartilage. 13:80–89. 2005.PubMed/NCBI View Article : Google Scholar
|
















