1. Introduction
According to the general logic of tissue
architecture and dynamics, a tissue that expands needs to build a
vascular network, an interstitium and it needs the help of other
supporting cells in order to survive and to grow. This is the case
of the epithelia that regenerate, of the embryo that develops, of
wounds that heal and, finally, of benign and malignant tumors
(1).
In the meantime, even if it has its independent
growth, the tumoral tissue is connected with the rest of the body;
different cells infiltrate it, either as homeostatic elements, or
as an attempt to fight against it, or even recruited by the tumor
to help it, contributing in these different ways to the shaping of
the tumoral growth (2).
These interactions proved to be important in the
development of tumors and are the object of the present review.
2. Cellular participants to the tumor
microenvironment
Cells of the innate immune system and
other stromal cells Macrophages
They have important functions as the first line of
defense against pathogens and tissue damage: phagocytic,
antigen-presenting, inflammatory cytokines and chemokine secretion
(3).
According to these roles, macrophages can be found
in different activation states, M1 state for inflammation and
immune defense, and M2 for tissue homeostasis and regeneration. In
fact, the spectrum of macrophage activation is much more complex
and intermediate forms between these states can be found (4). The macrophage is a versatile cell and
transitions between states can occur, under the influence of
external conditions and cytokine milieu.
In cancer, macrophages are thought to be recruited
by local conditions-hypoxia and necrosis- and by cytokines and
chemokines from the tumor cells, such as colony stimulating
factor-1 (CSF-1), interleukin 34 (IL34), as well as IL6, C-C motif
chemokine ligand 2 (CCL2) or CXCL10. They are mainly in the M2
state, produce IL10, transforming growth factor β (TGFβ), growth
and angiogenic factors like epidermal growth factor (EGF),
fibroblast growth factor (FGF), vascular endothelial growth factor
(VEGF), matrix metalloproteinases (MMPs), chemokines (CCL2, 5, 3,
8, 22), they do not secrete IL12 and do not present tumor antigens.
They are called tumor associated macrophages (TAMs) and they
contribute to the immune suppression and angiogenesis, migration
and invasion and recruit other immune cells (reviewed in 5).
There is also a M1-Th1 component in tumors,
triggered by tumor antigens, mainly in the initial stages, which
contributes to the antitumor defense (6).
What seems to determine a protumoral profile in
macrophages is the exposure to factors from the tumor (ILs 4, 6,
10, TGFβ, exosomes), while Toll-like receptors (TLR) ligation
through damage-associated molecular patterns (DAMPs) or agonists or
exposure to interferon-γ (IFNγ) will turn them into antitumoral
macrophages (5).
The presence of tumoral macrophages is generally
linked to an unfavorable prognosis (7). By consequence, tumoral macrophages are
considered for inhibition (Table
I). Given the versatility of these cells, ‘educating’ them is
also taken into account (8).
 | Table ICells that compose the tumor
microenvironment. |
Table I
Cells that compose the tumor
microenvironment.
| Type of cell | Specific
markers | Recruitment in
tumors | Protumoral
actions | Determinants of the
pro/antitumoral role | Antitumoral
actions | Presence in
tumors/prognostic associations | Targeting | (Refs.) |
|---|
| Macrophage
(TAM) | CD68,
CD11b+ HLADR+, -M1-CD86+
CD80+ INOS+ -M2-CD 163+ or
CD206+ | CCL2, 5 CXCL12
CSF1, VEGF (from the tumor cells) |
M2-like:-angiogenesis (Vegf, IL8, Ang-2,
FGF, MMP9) -EMC remodeling (MMPs 9, 12) -Growth factors (EGF, FGF,
PDGF) immunosuppression (Arg, PDL1, Fasl, IL10, TGF) adhesion to
the tumor cells, co-migration (ICAM1, VCAM, PECAM) -Recruitment of
other cells (CXCL17, 22, 24). -M1-contribution to oncogenesis
through inflammation | -For the protumoral
role-GCSF, L10, TGFβ, IL4, 13, tumor exosomes -For the antitumoral
role: IFNγ IL12, GM-CSF, DAMP, TLR, NLR agonists, apoptotic
cells | M1-Supports Th1, NK
LTs -Recruitment of defensive cells (CXCL9, 10) -Directly
tumoricidal (ROS, phagocytosis) -ADCC -Th1 effector | -Frequent (10-50%
of the tumor mass) -Mainly M2 Associated with poor prognosis in:
breast, gastric, pancreatic, oral cancer, lymphoma -Good prognosis
in melanoma, cervical, esophageal, colorectal cancer | Anti CCL2/CCR2,
-Anti CSF1, CSF1R -Anti IL6R, PIK3CA, STAT3 -Anti CXCL12, CXCR4
-IFNγ, IL12 (aiming at educating or eliminating TAMs) | (3,4,6) |
| Fibroblast
(CAF) | SMA-α FAP
Vimentin | FGF, PDGF SDF (from
the tumor cells) | Secretion of growth
factors (PDGF, TGF) angiogenesis (VEGF, CXCL12) myeloid cells
recruitment (CCL7) adhesion to the tumor cells (ICAM, Itg a11, IGF
KGF, HGF) invasion ECM secretion, remodeling (MMPs)
-Immunosuppression (TGFβ) -Inflammation (IL6) -Resistance to
therapies | -For the protumoral
role: -Mainly TGFβ -IL6 -Exosomes from tumor cells | Normal fibroblasts
restricts tumor initiation, while CAFs promote tumor growth | Main component of
the tumor stroma, in some situations (pancreas) being up to 80% of
the tumor mass Associated with poor prognosis | Normalization or
elimination of CAFs -Anti FAP, TGFβ, FGF, FGFR PDGFR, CXCL12,
CXCR4, IL6, PGE2 | (7,9,10) |
| Endotheliocyte | CD31 | -Angiogens: -VEGF,
PDGF, FGF, Ang2, IL8, CXCL1, 2, 3, 5, 12 -ILs 1, 6, 23 (from tumor
and associated cells | -Nutritive support
of tumors -Dysregulated network, maximally dilated→hypoxia
-Increase in NCAM→cell influx, angiogenesis -Increases resistance
to therapy -Adhesion to the tumor cells, interendothelial adhesion→
invasion -VEGF→decreased ICAM-1, VCAM→decreased influx of
immunocytes | -For the protumoral
role, angiogens, decreased ICAM-1 -For the antitumoral role:
Proinflammatory ILs (IL1, 6, 36, TNFα) →increased cells influx | Release of
proinflammatory ILs, chemokines→increased ICAM-1, VCAM- → influx of
defensive cells (LTs, monocytes) | In all tumors;
presence is necessary for tumor survival | Anti-VEGF
(bevacizumab) -Anti-PDGFR, Ang2, Tie2, ανδ3 Itg. -Depletion of
M2.N2 -IL1 should precede adoptive therapy (to increase cell
adhesion to the endothelium) | (12,13,97) |
| Neutrophil
(TAN) | CD16+
CD66+ CD15+ | CXCL1, 2, 3, 5,
IL8, GMCSF, G-CSF (from the tumor cells, macrophages) | Angiogenesis
(through VEGF), -EMC remodeling (MMPs 8, 9) -Tregs
recruitment-(CCL17) -Myeloid cell recruitment (IL8) -Adhesion to
the tumor cells, migration -Arginase and i-NOS are
immunosuppressive -Inflammation in tumors -NETs are
thrombogenic | N2 polarization
(protumoral)-mainly TGFβ N1 polarization-IFNγ, TNFα |
Tumoricidal-directly through
degranulation, phagocytosis -Through ADCC -Th17 effector Chemokines
secretion- -CCL19, 20-for DCs -CXCL9, 10-for LTs Stimulation of
CD8+, NK (through TNFα) Antigen presentation,
stimulation of LB in lymph nodes | TANs accumulate in
correlation with tumor stage -poor prognosis in: melanoma, renal,
hepatocellular, neck, lung, pancreatic cancer -NLR associated with
poor prognostic in many tumors | -Anti-G-CSF -IFNγ,
anti-TGF -Anti-IL8, anti- CXCR1, 2; -Anti-IL17 -MABs targeting
markers -NET targeting | (15,17,18) |
| Eosinophil | CD193+
Siglec-8+ CD15+ | CCL11, 24, 26, IL8,
CXCL1 -DAMP -IL5;-PGE2 ICAM-1, V-CAM | Angiogenesis
secretion of Th2 cytokines | For the antitumoral
role: DAMP, necrosis -A context of Th1 response favors the
antitumoral actions -Contributes to immune surveillance | Tumoricidal
-Directly through degranulation independent of Th2 -Th2 effector
-Through ADCC -Stimulation of CD8+ LTs -Antigen
presentation | Associated with
favorable prognosis in gastric, ovarian, nasopharyngeal,
colorectal, lung, esophageal, lung, prostate cancer
-Unfavorable-Hodgkin lymphoma, cervical cancer -TABE
(Tumor-associated blood eosinophilia) unfavorable prognostic in
renal, gallbladder, breast, pancreatic tumors | -Adoptive therapy
-Combined therapy eosinophil-lymphocyte increases the inhibition of
the tumor growth on models | (20,22,24) |
| Mast cell | FCεRα1+
CD117+ Triptase+ -Activated-
CD203c+ | SCF VEGF IL8 CCL2
CXCL1, 10, 15 (from the tumor cells) | Angiogenesis (they
accumulate near CD31): -Through histamin, PDGF, IL8 -EMC remodeling
-Proteases, MMPs, heparin -Immunosuppresion-IL10, TGFβ, adenosine
-Secretion of Th2 ILs→ accumulation of M2 macrophages; these ILs
have protumoral roles -Genotoxic through ROS and inflammation | For the antitumoral
role-TLR2 activation -For the protumoral role-exposure to the
TME | -Activation of
CD8+ LTs -Recruitment of defensive cells -Antigen
presentation -It is not directly tumoricidal -IL9-stimulates CD8,
CD4+ LTs -Through IFNγ, TNFα they stimulate the
defense | -Negative role in:
Hodgkin lymphoma, CLL bladder, thyroid, esophageal, breast,
prostatic, pancreatic colorectal cancer -Positive role in: breast,
lung, ovarian cancer -No association: Renal, lung cancer | Targeting where it
has a negative role -c-Kit inhibitors -Sylimarin inhibits Mc
recruitment, MMPs 2, 9 -Cromolyn | (26,27) |
| Myeloid-derived
suppressor cell (MDSC) | -Monocytic-
CD11b+CD14+ CD15- HLA-DR-
-Polinuclear CD66b+CD15+ CD14-HLA-DR- | CCL2, 5, CXCL5, 8,
12, GMCSF, VEGF, from tumor cells-soluble or exosomal | -Immune suppression
through Arg, ROS, galectin (TCR nitration) -ADAM17-↓ E-selectin
-Angiogenesis -At the ingestion of tumoral exosomes, MDSC express
IL6, VEGF -MDSCs (in ovarian tumors) have decreased
miR101→stemness | STAT3, NFκB;
tumoral exosomes + TGFβ, PGE2α | | -Present in most
tumors, correlated with stage -Associated with poor prognosis in:
breast, lung, pancreatic, uterine, prostatic tumors, HCC,
glioma | -Inhibition of CSFR
-Inhibition of CCL2 -Sildenafil (↓NO) -Inhibition of exosomes
release (dimethyl amiloride) -Education through IFNγ, IL1β, IL4,
TNFα, TLR-ligands | (71,98) |
| Dendritic cell
(DC) | Conventional-
CD11c+ HLADR+ CD1c+/-,
CD141+/- -Plasmacytoid- CD123, 303+
-Activated- CD83+ | CXCL9, 10, 12, 14,
CCL19, 20, 21 | -Dysfunctional DCs
in antigen presentation, maturation, infiltration -Tolerogenic
DCs-expressing PDL-1, IDO, Arg→ stimulation of LTreg, inhibition of
CD8+, CD4+ LTs, macrophages, neutrophils
-Immature DCs (MDSC) -DC deficiency→Th1 deficiency, bias towards
Th2 in tumors; also CD8+ deficiency | Protumoral role:
-DCreg profile due to TGFβ, IL10, tumoral exosomes -Inhibition
through PDL, Gal. IDO, Arg -Antitumoral-action on TLRs (DAMP,
apoptotic cells), IFNγ, TNFα | Presents antigen to
CD8, CD4+ LTs -Through IL12, IL15 -Stimulation of Th1,
CD8+, NK LTs -Role in the Th1, 2.9.17, Tfh, Treg
differentiation -Direct cytolitic action -Stimulation of memory
LTs | -Mature DCs-good
prognostic in melanoma, head-neck, colorectal. bladder, oral,
gastric, uterine cancer -Plasmacytoid DCs-associated with poor
prognostic in melanoma, glioma, breast, ovarian, oral, gastric,
renal and lung cancer | -DC-based vaccines
-Anti-PDL.TIM-3, -Anti-IL10, TGFβ, miR155 -CD-40 agonists -SiRNA
anti STAT3 | (39,40) |
| CD8+
lymphocyte | CD3+
CD8+ Gzm, Perf. -activated: CD69+,
CD25+ -exhausted TIM-3+,
LAG3+ | CXCL9, 10, CCL5
(from DCs, M1, some tumor cells) | -In oncogenesis
(for example in HCC) -Self-inhibition through the upregulation of
PDL-1 by IFNγ | -Inhibited through
signals from the tumor, from CAFs, TAMs, Treg (PDL-, TIM3-L Fasl,
B7-H3) -It depends on antigen presentation, co-stimulation and ILs
from DCs -Support from eosinophils, Th1, Th9, NK cells | -Cytotoxic through
direct contact (Granzyme, perforins) -Recruitment, stimulation of
other cells | -Infiltrating
CD8+LTs associated with good prognosis in many cancers
like breast, colorectal, renal, prostatic, bladder, ovarian
cancers | Adoptive therapies
bispecific antibodies Anti PDL-1, anti- CTLA4 -ILs 2, 9, 12, 15, 18
21, 27 -CXCL9, 10 | (43-45) |
| CD8+
memory lymphocyte | CD3+
CD45RO+ CCR7 (CD197)+ (central)
CD127+ | CXCL9,10 CCL3.4,5,
8,14 | In some tumors
(lung, ovarian)-exhausted | -Differentiation
through IL15, IL33, TGFβ -CD1c | -Cytotoxicity
(through granzyme, perforins) -Secretion of IFNγ, TNFα -Chemokines
for other cells (CCL3, 4, 5) -Through CD103 they inhibit tumors
expressing E-cadherin -Response to vaccines requires
CD8+ mem | -Associated with
good prognosis in: ovarian, lung, urothelial, uterine, breast,
bladder tumors, glioma (cd103+) -Exception: colorectal
cancer -Marker of immunogenic tumors | -Vaccines -Adoptive
therapies -Anti-PDL-1 | |
| NK lymphocyte | CD56+
CD3- CD16+ | CX3CL1 | -Inhibited in many
tumors due to the immunosuppressive milieu | Stimulation through
ILs 2, 12, 15, 21-antitumoral | -Immune
surveillance -Cytotoxic on cells with low MHC, MICA+
cells -Through ADCC, CDC, -Stimulation of CD8+LT, DCs
-IL12, 2, 15, IFNγ secretion | Associated with
good prognosis in gastric, colorectal, liver, lung, renal cancer
and others | -Adoptive therapy,
CAR-NK -IL-IL2, 12, 15 -Anti-PDL, anti-KIR anti-NGG2A -MAB, BsAB
for CD16 or NCR-tumor antigens | (34,35) |
| γδ T
lymphocyte | CD3+
γδTCR+ | CCL5 CXCL9, 10,
LFA, VLA1-4 L-selectin, Integrin ανβ7 | γδ-17
subset-production of IL17 in colorectal tumors→chronic
inflammation, angiogenesis, recruitment of myeloid cells
-Suppressive subset | -Tumoral
phosphoantigens or injection of phosphoantigens stimulates the
cytolitic activity -The immune suppression from tumors-they become
suppressive | -Presents antigens
to CD4, 8+ -Non-peptidic antigens -Through NKG2D, TRAIL
perforins, granzyme -ADCC -Secretion of IFNγ, TNFα, IL12, 15,
36-stimulation of DCs -Rapid response -Cooperation with NK, LB
(γδTfh subset) | The strongest
association with favorable prognostic in solid cancers, leukemias,
lymphomas -Negative association through the γδT17 subset in
ovarian, bladder, colorectal cancer | -Phosphoantigens
-Bisphosphonates -Adoptive therapy -Multi-immunocyte- γδT+αβT or
CIK-with IL17 measurement -Modifying the IL balance-giving ILs 21,
15, 12, 36 | (28-30) |
| NKT lymphocyte | CD56+
CD3+ | | NKT II
subset-secretion of ILs 4, 13→↑ M2, MDSC Inhibits LT
CD8+; stimulates LB, LTh2 | -Sulfatides from
the tumor→activation of NKTII-protumoral -ILs polarize NKT LTs pro
or anti-tumoral -α-Gal-cer promotes antitumor immunity | Glycolipidic
antigens presented on CD1d -in CMH-deficient tumors -Secretion of
IFN, activation of NK -Rapid response to IL, DAMP-TLR -Adaptive
response regulation -IL12, CD40 → DCs, NK, CD8 | -Associated with
good prognosis in myeloma, lung cancer -They protect from some
tumors, being necessary for this effect | -Adoptive
therapy-CIK cells -The agonist α-Gal-cer. -Anti IL4, 13, TGFβ
-α-Gal-cer.-adjuvant for vaccines | (31-33) |
| Th1 lymphocyte | CD3+
CD4+ T-bet+ | CXCL9,10. CCL5 from
APCs, certain tumor cells | Inhibited in many
tumors through Th2, M2, Treg, TGF, IL10, IL4 -Exhausted by chronic
exposure to tumoral antigens -Self-inhibition through the
upregulation of PDL-1 by IFNγ | For the antitumoral
role-IL12, 18, 27, TNFα | -Tumoricidal
through M1 Mϕ -stimulates CD8+, NK LTs -DC licensing
-Necessary for an efficient antitumoral response or response to
vaccines especially with exhausted CD8+ LTs or tumors
CD8+ resistant or expressing Fasl -Recruitment of
CD3+ cells -CD4+ CTLs | Associated with
favorable prognosis in many tumors, from which melanoma,
colorectal, ovarian, breast, cancer, multiple myeloma | -Stimulation with
IL2, IL12, IL18, IL27 -In adoptive therapy -CXCL 9, 10 | (46,47) |
| Th2 lymphocyte | CD3+,
CD4+ GATA3+ | -CCL 17, 22 (tumor
cells, M2 macrophages) or locally polarized | -Inhibition of Th1
-Through the Th2 cytokines which are protumoral -Non-stimulation of
CD8+ LTs -Stimulation of M2 macrophages -Stimulation of
B lymphocytes- sometimes protumoral | -DC
dysfunction→↓IL12 -TSLP (from CAFs) →bias towards Th2 in
tumors | -Together with Th1
they contribute to a complete response including to vaccines
with/without the intervention of CD8+ LTs -Through the
tumor cell necrosis -Through eosinophils -In adoptive therapy they
eradicate Th1 or CD8+-resistant tumors -IL4 necessary
for the development of CD8+ LTs | Associated with
good prognostic in some tumors like breast cancer and lymphoma
-Poor prognostic in gastric, pancreatic, ovarian cancer -No
association in colorectal cancer | -IL12, IFNγ -Anti
IL4 (models) -For the positive role: -Adoptive therapies -Vaccines
(when both Th1 and Th2 ILs increase) | (22,49,50) |
| Th9 lymphocyte | CD3+
CD4+ IL9+IFNγ- | | IL9 stimulates mast
cells that can be protumoral | Th9 polarization:
IL4+ TGFβ | -IL9-stimulates
CD8+LTs, mast cells (CCL20) -IL21-stimulation of NK
cytolysis, IFNγ secretion of CD8+LTs -Direct cytolysis
through Granzyme | -Presence in tumors
has been reported | -Adoptive therapy-
sometimes more effective than Th1 -The effect is mainly through
CD8+ | (55) |
| Th 17
lymphocyte | CD3+
CD4+ IL17+ RORΥt+ | CCL18, 20, 22, CCL
4, 5 | -Th17 cytokines
(IL17, 22, 26) are protumoral, proangiogenic -Through the LTh17-reg
subset -Through the inflammation which is protumoral | -For the
anti-tumoral role IL12 through stimulation of Th17-1 subset -For
the protumoral role- TGFβ-stimulation of Th17-reg subset | -Cytotoxic through
neutrophils -In a Th1 milieu they contribute to the defense
-Stimulation of B lymphocytes through IL21 -Through Mφ and DC
-Through the Th17-1 subset- secretion of TNFα, IFNγ -Recruitment of
CD8+LTs, neutrophils, LB, DCs in tumors (CCL20, 2, 7,
CXCL9, 10) | -Adoptive
transfer-good effect -Natural Th17LTs-associated with poor
prognosis -Better prognosis in ovarian, prostatic, lung cancer,
seminoma -Worse prognosis in pancreatic, colorectal tumors,
HCC | -Adoptive therapies
with LTh17 -Inhibition of IL17 IL22, 23, 26 | (42,51-54) |
| T regulatory
LT | CD3+
CD4+ Foxp3+ CD25 (IL2R)+ | -CCL22, 28 from
tumor cells -Activation of RAS→ infiltration with Tregs | -Immunosuppression
-Angiogenesis -Inhibition of immunotherapy -Inhibitory potential of
Tregs increases much in cancer | -TGFβ, IL10, IL35,
IL2 -Other cells stimulate Tregs (MDSC, M2, Breg, CAF) -Adenosine
from tumor cells | -Positive role
where the protumoral inflammation is dominant -Versatility under
the influence of LTh2; (TGF+IL4) LTreg become LTh9,
antitumoral | -Present in cancer,
associated with worse prognostic in many cancers like breast,
ovarian, gastric cancer -Associated with good prognosis in
colorectal, breast cancer and lymphoma | -Anti TCR, Foxp3,
-Anti CD25 (IL2R) -Anti PDL1, CTLA4 -IFNγ -GITR, TNFR inhibition
-LTreg education | (57,58,60) |
| T-follicular
lymphocyte (Tfh) | -CXCR5 (homing in
follicles) -Bcl6 | | Tfh2, 17
subsets-protumoral action | -Antigen
presentation by DCs, LBs, IL6, ICOS, TGF→Tfh differentiation
-Stimulation by DCs, Mφ through IL6, 21, 27 | -Secretion of ILs
21, 4, 12 -CXCL13→influx of LBs, generation of tertiary lymphoid
organs (TLO) -Affinity maturation of ABs -Differentiation of LB mem
-Rescuing LTs from anergy in the TLO -Associated with increase of
CD8+, Th1 LTs (IL21) -Inhibition of LTreg | -Associated with
favorable prognosis in breast, colonic cancer (presence of TLOs)
-Unfavorable prognosis in gastric cancer | -Adoptive therapy
-CD19-directed CAR-T-cells | (28,57) |
| B lymphocyte
(LB) | -CD19+
-Plasma cells CD138+ | CXCL 13 | -They inhibit the
antitumor response -Immune complexes occupy FcR of neutrophils,
mast cells, macrophages →angiogenesis -The Breg subset inhibits
CD8+, NK, Th1, NKT, LTs and stimulates LTregs through
IL10, TGFβ -Lymphotoxin-lyphangiogenesis | -The activation
state seems to determine the pro-or antitumoral profile
-Activated-antitumoral, resting-protumoral -Breg-protumoral -Plasma
cells-antitumoral | -Presents antigen
to CD4+LTs -ADCC -Complement activation -Some LBs are
cytotoxic -IL2 secretion -In some models LBs were necessary for the
Th1, CD8+ response -Resistant to immuno-suppression | -Associated with
good prognosis in breast, hepatocellular, biliary, gastric cancer
-Negative role in melanoma pancreatic, lung, oral cancer | -Adoptive
therapies-successful -Antibody usage according to the B lymphocyte
model (MABs) -Inhibition where they have a proven negative role
(rituximab) -Association of adoptive B and T LTs superior to single
cell therapy | (61,62) |
| B-memory
lymphocyte | CD19+
IGD- CD27+ | | | | -Tumor
antigens-weakly immunogenic, sometimes tolerogenic- →Bmem LTs are
important for vaccines -Bmem LTs present antigens to
CD8+LTs, costimulation (CD40-CD27) | | -Searching epitopes
with effect on Bmem is important | |
Fibroblasts
They are the main component of the connective tissue
and also of the tumor stroma; they secrete the intercellular matrix
and fibrils, sustain tissues, contribute to the tissue and wound
healing, to fibrosis and, when activated, to inflammation. Specific
markers are vimentin, smooth muscle actin-α (SMAα), fibroblast
activation protein (FAP) (9).
According to these roles, there are sub-populations
of fibroblasts: tissue resident fibroblasts, which realize tissue
turnover and sustaining, fibroblasts with regeneration function
that migrate to injured tissues and contribute to healing,
inflammatory fibroblasts, activated by immune cells; there are also
sub-populations of fibroblasts specific for each body region, with
specific HOX gene codes (10).
In cancer, fibroblasts are induced by tumor cells,
together with blood vessels, through factors such as FGF or PDGF
(platelet-derived growth factor), as well as by hypoxia in tumors.
They have a predominantly activated state, due to the action of
IL1, TGFβ, PDGF, stromal cell-derived factor (SDF) and reactive
oxygen species (ROS) (11) and are
known as cancer-associated fibroblasts (CAFs). They are the
majority of the tumor stromal cells.
CAFs secrete the extracellular matrix (ECM) and also
active substances such as cytokines, growth factors (GFs) like
TGFβ, HGF, SDF and MMPs, through which they shape the
microenvironment of tumors, they are angiogenic through VEGF and
PDGF and support the tumor growth and invasion. They usually
negatively modulate the antitumor immunity through chemokines (CCL2
and 5) and cytokines (TGFβ), attracting T-regulatory lymphocytes (T
regs), myeloid-derived suppressor cells (MDSCs) and also helping
tumor cells to migrate (through CXCL12) (11,12).
Fibroblasts are also subjects of immunotherapy
(Table I) (reviewed in 12).
Endothelial cells
Together with pericytes, they form the boundaries of
capillaries and regulate the flow of substances and cells from
within the vessel out and backwards. They are dynamic structures,
responding to cytokines, growth factors and other active substances
like IL1, tumor necrosis factor-α (TNFα), IFNγ, IL4,
PGI2-prostaglandin I2, NO-nitric oxide, VEGF, FGF and secreting
active substances (IL1, TNFα, IL6, IL8, IL20, IL33,
LPS-lipopolysaccharide), through which they participate in the
inflammatory processes and augments them when needed (13).
The endothelial cell also participates in the
regeneration and healing processes, responding to angiogenic
factors (VEGF, FGF, Ang-angiopoietin) and secreting them (13).
In tumors, due to the multiplication of cancer
cells, there is hypoxia, which leads to the hypoxia-inducible
factor-1 (Hif-1)-dependent augmentation of VEGF and other
angiogenic factors (14); this
causes angiogenesis. The vessels that form are different from
normal vessels, being tortuous, disorganized, with few or no
pericytes and to some of their length without walls (15).
The result is a modified metabolism in tumors,
changes in the physical qualities of the ECM, metastasization,
abnormal distribution of drugs and abnormal trafficking of immune
cells to and from the tumor.
Neutrophils in tumors
Their normal function is to respond to pathogens at
the beginning of the immune response, through phagocytosis and
extracellular traps (NETs). Through the cytokines they produce,
they amplify the response of other cells in inflammation. In
addition, they present antigens and contribute to the end of the
inflammatory process, by phagocytizing dead cells (16).
There are fewer neutrophils than macrophages in
tumors, attracted by chemokines such as IL8 from the tumor cells or
by inflammation and necrosis. They are short-lived, but they have a
turnover and contribute to the process in two opposite ways: they
can be antitumoral (N1 neutrophils), especially in a milieu with
IL12 and TNF, and this effect requires the presence of
CD8+ cells (17).
They can be protumoral through the secretion of
MMP9, HGF and VEGF, where the neutrophil depletion leads to the
cancelling of the angiogenic switch (18). This is the natural state of
neutrophils in tumors and this is one of the reasons for which
blocking IL8 reduces tumor growth (17). TGFβ is believed to be the main cause
for this protumoral profile, called N2, while IFNγ turns the
neutrophils into N1 tumoricidal cells (17).
As a rule, neutrophilic infiltration in solid tumors
is associated with worse prognosis (19). A greater number of circulating
neutrophils, reflected in the neutrophil/lymphocyte ratio, is
associated with worse prognosis in many tumors (20).
Eosinophils in tumors
The eosinophils have a role in clearing parasitic
and some bacterial infection. They participate in the immune
response, especially in the Th2 type. They are activated by
IL5(3) and secrete more than 30
cytokines, such as IL1, 2, 3, 4, 5, 9, 10, 12, 13, 17, 25, IFN,
TNF, chemokines such as CCL11, MIP, MCP, CCL5 and growth factors
NGF, PDGF, EGF, TGFα and β (21).
Eosinophils were shown to be tumoricidal in some
tumors, where the presence of CD8+ cells was needed
(22). However, they are an
important source of IL4, which is Th2 polarizing and protumoral
(23). Nonetheless, eosinophils are
effective in the CTL (cytotoxic T lymphocyte)-resistant tumors on
models (24) and in tumors with
cells engineered to express IL4 there was a rich infiltration with
eosinophils and macrophages, which had tumoricidal effect (25).
The eosinophilic infiltration in tumors (except
Hodgkin lymphoma) is associated with good prognosis (26).
Mast cells in tumors
Mast cells are cells with an important role in the
innate and adaptive immunity. They are involved in the immune
defense of the mucosal barriers and express TLRs 1-7, 9 and Fcε
receptors (FcεR). They recognize DAMPs and release inflammatory
mediators contained in their granules or cytokines (IL1, 6, TNF)
and recruit other cells such as neutrophils, eosinophils,
CD8+ and natural-killer lymphocytes (NK LTs). The mast
cells present antigens via MHCI or II (major histocompatibility
complex), they stimulate DCs (dendritic cells) and contribute to
angiogenesis (27).
In tumors, mast cells can play either pro or
antitumoral role (Table I)
(reviewed in 28). Mast cells exposed to the tumor microenvironment
are mostly protumoral, while the action on their TLR-2 receptors
has been shown to stimulate an antitumoral profile in them
(29).
Lymphocytes of the innate immune
system γδ T-cells
They form 0.5-5% of the lymphocyte population, being
present mostly in the gut and in the skin. γδ T-cells act on
phosphoantigens, but they also present antigens to CD8+
and CD4+ LTs and cooperate with NKs (3). Of all immune cells in tumors, it is
the subset with the strongest association with good prognosis in
cancer (30).
However, there are also protumoral subsets: γδ-17
LTs, which secrete IL17, and a suppressive subset (31). The suppressive TME can turn γδ LTs
into suppressive LTs. They are extensively studied for adoptive and
other therapies (Table I) (reviewed
in 32).
NKT cells
They respond to hydrophobic antigens,
presented by CD1d-type MHC. There are two main subsets of NKT, type
I with an invariant T-cell receptor (TCR), which is antitumoral by
stimulating DCs, CD8+ and NK LTs, and type II, which is
mostly protumoral (33).
There are also NKT-17, Tfh (T follicular)-like and
Treg-like subsets, with dominant protumoral activity (34).
NKT cells are also intensively studied for adoptive
therapies (CIK cells), for selective stimulation of NKT1 with
α-galactosyl-ceramide (α-Gal-Cer), for interleukin therapy and
others (reviewed in 35) (Table
I).
NK lymphocytes
They are main antitumor defenders acting on cells
with low level of self-proteins (MHC-I) like some tumor cells. They
also possess NKG2D receptors for atypical MHC, such as MICA and Fcγ
receptors, through which they perform antibody-mediated cell
destruction (ADCC). They stimulate CD8+ LTs and DCs
through the IFNγ they secrete, receiving in turn support from Th1
(T-helper 1 lymphocyte), Th9, CD8+ LTs and M1
macrophages through ILs 9, 12, 15, 21 and type I and II IFNs
(3,36). NK cells often become exhausted and
suppressed in the inhibitory TME.
They are also investigated for adoptive cell
therapy, CAR-NK therapy and others (reviewed in 37) (Table I).
Innate lymphoid cells (ILCs)
ILCs possess CD127 (IL7R) and have three main
subsets: ILC1 that secrete IFNγ, having some antitumoral activity
(38), ILC2, with pattern of
secretion such as Th2 LTs and ILC3, with secretion pattern like
Th17 LTs. Intervention of these subsets in cancer resembles that of
their adaptive counterparts (36).
An antitumoral role has been found for ILC3 in some models
(39).
Dendritic cells (DCs)
DCs are a heterogeneous cell population present in
every tissue and they are professional antigen-presenting cells to
LTs. They also secrete cytokines (ILs 1, 2, 6, 12, 15, 18, 37, 23,
27, 7, 37, 31, 10, IFNs) and chemokines (IL8, IL16, CCL9). They
express MHC II type proteins. DCs have myeloid or plasmacytoid
origin. Depending on the type of antigen exposure and cytokines
they secrete or are exposed to, DCs contribute to the Th1, Th2,
Th17 polarization or to immune tolerance (40).
In cancer, dendritic cells present tumor antigens to
LTs, their number being linked to good prognosis (41). Their depletion leads to the
depletion of LTh1 cells, which is detrimental. Tumor cells
influence infiltrating DCs, lowering their number and antigen
presenting capabilities. They become tolerogenic DCs, which
contribute to the immune suppression in the TME and accumulation of
Treg cells. Cytokines from the tumor (IL10, TGFβ, VEGF, low IL12)
and activation of PD receptors contribute to this (reviewed in 42).
Certain miRNAs (micro-RNAs) also influence the behavior of DCs
(43).
Cells of the adaptive immune system
CD8+ T lymphocytes (CTLs)
CTLs represent the main antitumor defenders in the
human organism; they act after tumor antigen presentation by DCs
(1), also receiving support from
Th1, Th9 and M1 macrophages (reviewed in 44). CD8+ LTs
are active especially in the earlier stages of the tumor
development. Later, they develop exhaustion and apoptosis due to
the activation of programmed death (PD) receptors and to the
immunosuppression of the TME (45).
The presence of CD8+ lymphocytes is
associated with good prognosis in many cancers (46). Given its position as key player in
the antitumor immune defense, this subset is the most extensively
used in different immunotherapeutic approaches, immune checkpoint
blockade, adoptive therapies, bispecific antibodies, interleukin
therapy, chimeric antigen receptors (CAR)-engineered cells and
others (reviewed in 47) (Table
I).
CD4+ T lymphocytes
They are in different states of polarization,
depending on which cytokine combinations act on them (1):
The Th1 (T-helper-1) subset polarizes in the
presence of IL12 from M1 macrophages and IFN, to which also IL18,
IL27, IL1α contribute; they are also contributors to the antitumor
defense (48) (Table I). It has been shown that an intact
CD4+ component is necessary for an efficient antitumoral
response (49).
The Th2 subset is polarized by ILs 4, 13, 19, 25 and
33 from mast cells, NK and CD4+ mem
(CD4+-memory) cells (1,22) and
secrete Th2 cytokines such as ILs 4, 5, 10, 13, 25, 31, 33. In
tumors, they are not favorable to the defense process, because of
the secretion of IL4 and 13 that inhibit the Th1 response, and of
the cytokines mentioned above, which have mainly a pro-tumoral
effect (50). Until recently, the
Th2 subset was considered almost entirely protumoral, but in the
light of recent data, it was shown that Th2 lymphocytes can also
contribute to the antitumor defense, being necessary for some of
its observed effects (25,51) (Table
I). This is the reason why Th2 LTs were studied for adoptive
therapies, with good effect (52).
The Th17 subset is polarized in the presence of
IL1β, IL6, IL23 and TGFβ and they secrete ILs 17, 22 and 26,
through which they sustain the tumor growth (53). There is a plasticity of the Th17
cells, which can be reprogrammed in Th1 or Treg lymphocytes
(54). As in the case of the Th2
subset, the initial view that Th17 is protumoral was reconsidered
in the light of some data that showed a beneficial role, especially
in adoptive therapy, with success even in cases that were
CD8+ and Th1-resistant (44,55,56)
(Table I).
A particularity of the antitumoral immune response
is just this combination of subsets that are usually mutually
exclusive in the immune response, such as Th1 and Th2; in tumors
they seem to coexist and even to cooperate, building an immune
defense that is unique to tumors (44,51).
The Th9 subset, through the ILs 9 and 21, which have
a stimulatory action on the CD8+ cells, are contributors
to the antitumor defense. Th9 lymphocytes also have a direct
tumoricidal action (57).
The Th22 subset secretes IL22, whose action is
protumoral (58).
T follicular LTs (Tfh) are associated with good
prognosis in many tumors (30).
They support the antitumor defense by building the tertiary
lymphoid organs in tumors, which are associated with good prognosis
(59).
The Treg (T-regulatory) subset polarizes in the
presence of TGFβ from the tumor and associated cells and has an
immunosuppressive and by consequence protumoral action through TGFβ
and ILs 10 and 35 that it secretes, and through other inhibitory
mechanisms (reviewed in 60) (Table
I).
Their presence is associated with poor prognosis
(61). An exception is represented
by tumors in which there is a strong inflammatory component, where
the presence of the Tregs is beneficial (62). They are considered for inhibition
through different strategies (60,62)
(Table I).
B lymphocytes
They are antitumoral in some tumors such as breast,
hepatocellular, gastric and biliary tumors, while being protumoral
in others (melanoma, pancreatic, lung, oral cancers) (reviewed in
63) through different mechanisms (64) (Table
I).
A more detailed review of the pro and antitumoral
role of the mentioned cell types of in tumors, along with the
factors that drive their pro or antitumoral state, can be found in
Table I and in some other recent
works (30).
3. The tumor microenvironment, from
biogenesis to complex relations
Complicated networks are generated through the
interaction of these cells, having as result either the promotion
or the inhibition of the tumoral growth. In a previous work, we
have reviewed these pro and antitumoral networks, the way in which
they interact and the therapeutic opportunities that the
understanding of these complex relations may open to immunotherapy
(65). In the present study, these
networks are analyzed in their dynamics, starting from the
biogenesis of the TME, with the same objective of exploring the
ways in which the knowledge of this network structure may help to
improve therapy.
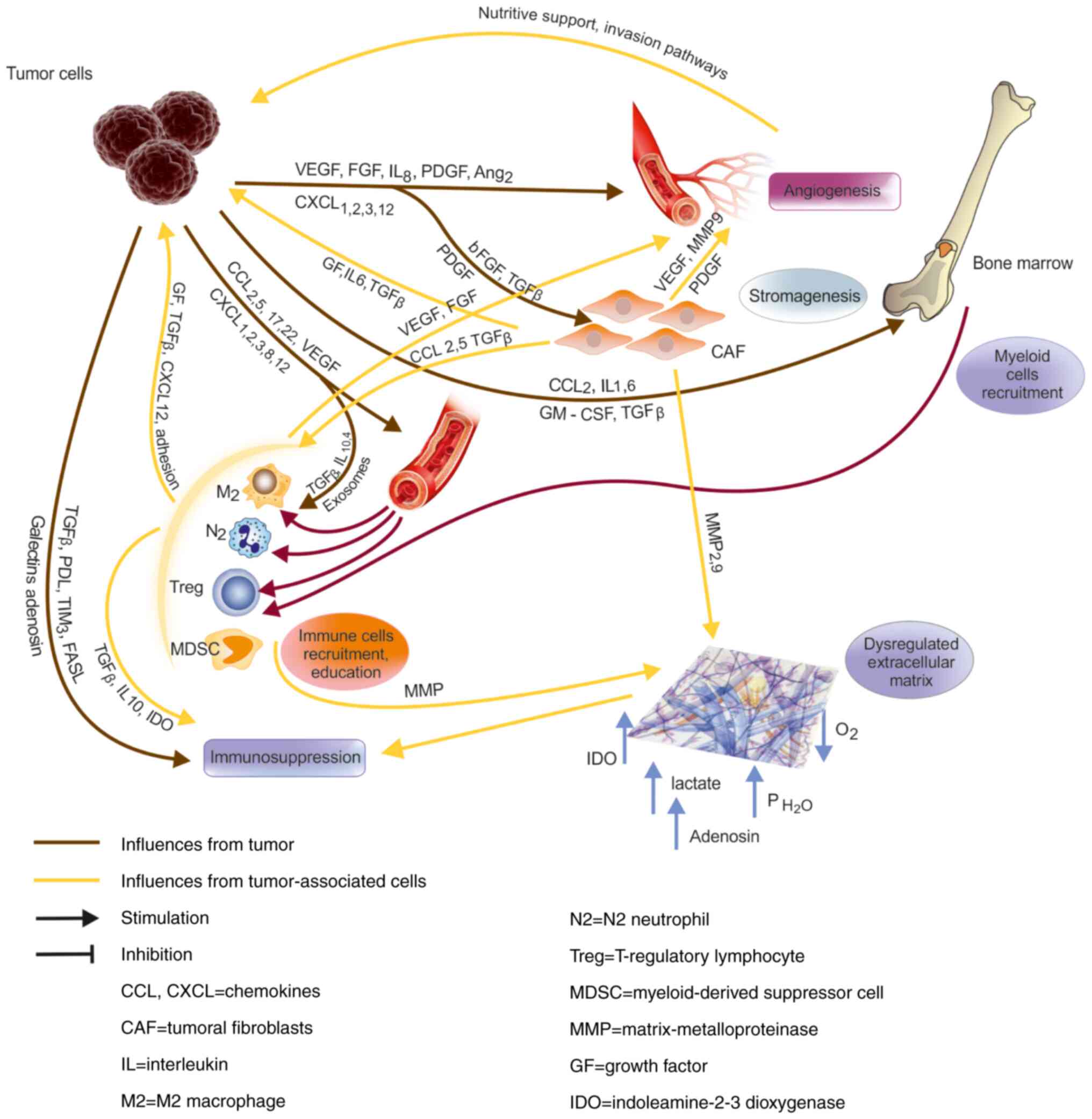
The biogenesis of the tumor
microenvironment Angiogenesis and stromagenesis
In their continuous expansion and proliferation,
tumor cells arrive at a certain point where they become hypoxic and
they need to build a vascular network.
Some studies show that hypoxia does not have any
role, the activation of the oncogene being enough to cause the
overexpression of angiogens (66);
other studies also reveal a hypoxia- and Hif-1-dependent mechanism
(67). In fact, to the embryo, a
main regulator of angiogenesis is the partial pressure of
O2 (PpO2).
It has been shown that the angiogenic switch takes
place early in the evolution of tumors, even in the premalignant
stages (18).
Stromagenesis
The tumor cells secrete FGF, through which they
recruit fibroblasts from the surrounding tissues; other sources of
fibroblasts are thought to be the endothelial and the tumor cells,
through metaplasia. The fibroblasts secrete the ECM, which is
dysregulated in tumors, with a different collagen content and
structurally altered, compared to normal ECM (11).
Recruitment of cells in tumors
The tumors secrete chemokines: CCL2, IL8, CXCL12,
CXCL1, 2, 3, GM-CSF, CCL 5, CCL17, sometimes also anti-tumoral
chemokines CXCL9 and 10. This leads to the accumulation of
monocytes, neutrophils, regulatory lymphocytes (68).
The cells arrive in tumors for several reasons: they
enter, as in any tissue, due to a basal secretion of chemokines
(but dysregulated in cancer); they also enter, in an increased
number, in the case of the existence of distress signals from
within the tumor. This leads to the overexpression of inflammatory
chemokines (IL8, CCL2, 5, CXCL9, 10) and infiltration of the tumor
with defensive cells.
It has been shown, under experimental conditions,
that the activation of an oncogene leads to the overexpression of
chemokines such as IL8, CCL2, CCL17, leading to the infiltration
with myeloid or lymphoid cells (69).
Some studies report on auto-inflammation in the
tumor because the activation of the EGFR or other oncogenic
pathways would also lead to the activation of the NFκB pathway with
consecutive secretion of IL1, 6, TNF and inflammation of the tumor
environment. The NFκB pathway can carry oncogenic mutations itself
(reviewed in 70).
This cell infiltration in tumors occurs from early
stages of the tumor development, even from preneoplastic stages
(71).
Under the influence of tumor-generated factors (IL4,
10, TGFβ, exosomes), these cells acquire a protumoral profile, of
tissue reconstruction and immune suppression (M2, N2, Treg)
(72).
Immature myeloid cell recruitment
Tumor-secreted factors (IL1, IL6, GM-CSF, TGFβ,
CXCL12) act at the level of the bone marrow and trigger an
accelerated myelopoiesis, having as result immature myeloid cells
(MDSCs), which accumulate in the tumor, leading to
immunosuppression and favoring tumor growth (73). This phenomenon is more advanced as
the tumor progresses.
Immune suppression in tumors
The tumor microenvironment is immunosuppressive.
This immunosuppression takes place through a few mechanisms, from
which: the overexpression of inhibitory molecules by the tumor and
by the tumor-associated cells (PDL-1, B7-H3, TIM-3 ligands, CD47)
and of death factors (FasL); cytokines such as TGFβ, ILs 4, 10, 35;
secretion of exosomes with immunosuppressive content; recruitment
of suppressive cells (Treg, M2, MDSCs); metabolites (lactate,
adenosine), hypoxia, increased hydrostatic pressure in tumors
(Fig. 1) (reviewed in 74). The
nature of this immunosuppression becomes more clear from some
experiments with conditionally activated oncogenes; it has been
shown that following the oncogene inactivation, the TME was
infiltrated by immune cells (CD8+, NK LTs), that
destroyed tumors (75). Insight
into the mechanisms of this relation between oncogenes and immunity
showed that activation of Myc led to the upregulation of both CD47
and PDL-1 on the tumor cells (76).
The immunosuppression was an effect of the oncogene
activation, along with the others mentioned above (angiogenesis,
cell recruitment).
The level of immunosuppression is directly
correlated with the tumor progression (74). Other characteristics complete the
picture of the TME:
Physico-chemical qualities of the ECM: high
hydrostatic pressure, hypoxia, high lactate and adenosine content,
IDO-Indoleamine 2, 3-dioxygenase-from MDSCs (77).
Inflammation: In tumors, inflammation can be
both pro or antitumoral. It has been shown that inflammation exerts
a tumor-promoting role in cancer, through multiple mechanisms such
as angiogenesis, release of genotoxic product like ROS, enhanced
survival, stimulation of proliferation, stemness or invasion of
tumor cells (reviewed in 70); this is true especially with regard
to chronic inflammation. On the contrary, experimental data also
support a positive role of inflammation in cancer (78). This happens especially in the
presence of a strong Th1 component and in the acute phase of
inflammation (79).
Proliferation, angiogenesis, cell
recruitment and immune suppression - a coordinated program?
The events that occur in the TME (proliferation,
angiogenesis, cell recruitment and immune suppression) are
recognized hallmarks of cancer (80). Considering them as a whole made some
researchers compare tumors to a tissue regeneration process, like
the one that occurs after a tissue destruction or after the
resolution of an infection (81).
This program is triggered by stimuli such as growth
factors or signals of termination of the immune response, and has
the same components: cell multiplication, angiogenesis, recruitment
of cells with regeneration potential (macrophages, neutrophils,
fibroblasts, Tregs, epitheliocytes) and immunosuppression.
The program is stopped when signals of tissue
integrity and completion (integrin or cadherin signaling, hippo
pathway signals) are received.
By contrast, in cancer the program is started
aberrantly by the activation of oncogenes, is dysregulated and does
not respond to stop signals.
There is some experimental evidence that supports
this analogy (82). The comparison
is not 100% accurate, but it can serve as starting point for some
therapeutic considerations (83).
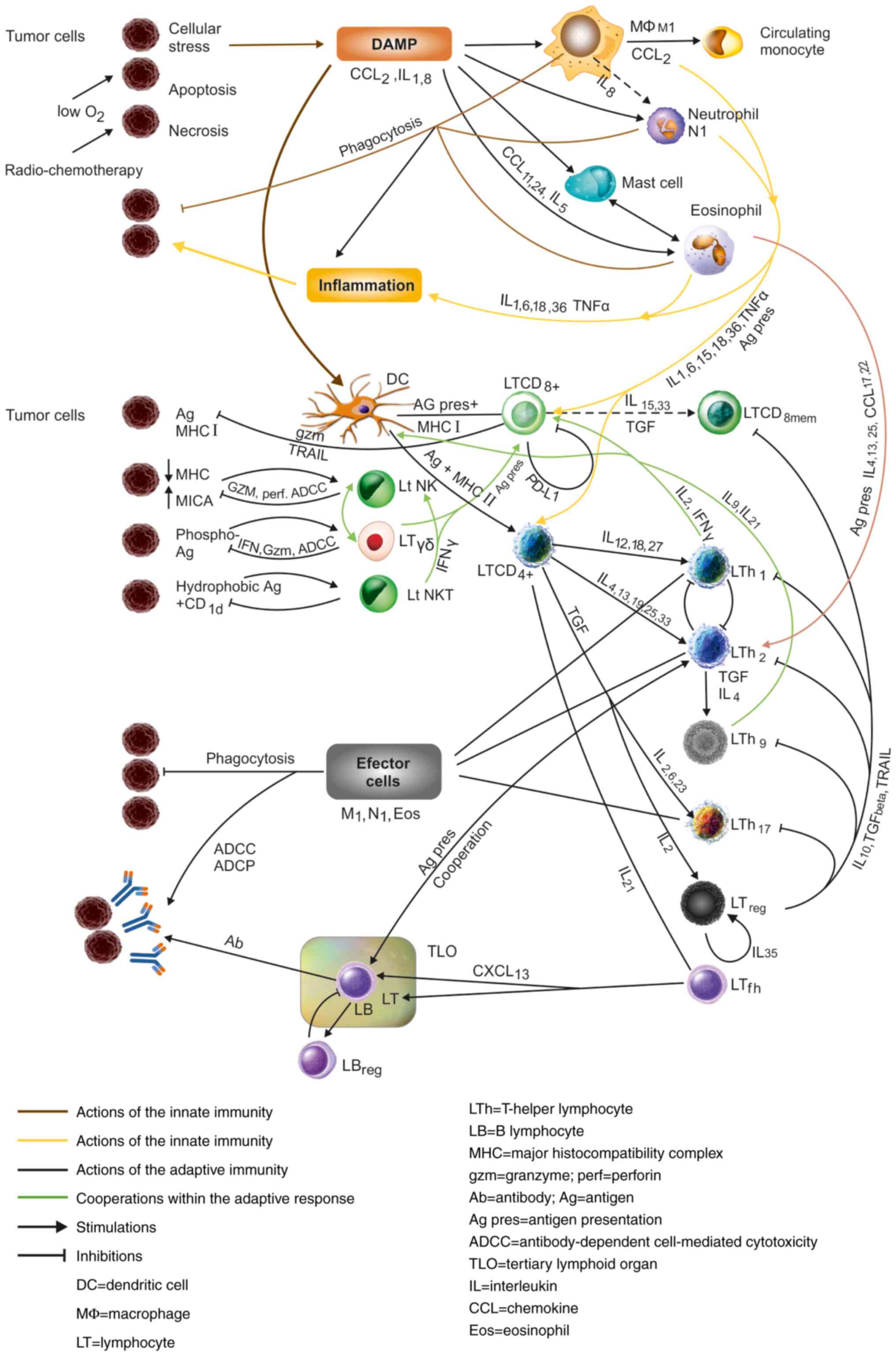
Immune response in tumors
The presence of the danger signals from the hypoxic
or necrotic tumor cells triggers an innate response from
macrophages, neutrophils, mast cells and eosinophils, directed
against the tumor.
On the other hand, dysregulation of MHC provokes a
tumoricidal response from the NK LTs, while particular antigens,
phosphoantigens or lipids, will awake the response of cells like γδ
or NKT-1 cells.
The antigen presentation from antigen-presenting
cells (APCs) such as DCs, but also B-lymphocytes (LBs), macrophages
and even neutrophils, mast cells or eosinophils opens the way for
the intervention of CD8+CTLs, which is completed by the
activation of CD4 T-helper lymphocytes, with their effector
mechanisms (effector cells of Th subsets, M1 macrophages,
neutrophils and eosinophils) (Fig.
2) (3,84).
There is an adequacy of the immune response to a
large array of stimuli. The diversity of signals and antigens in
tumors requires the deployment of such a great diversity of cells,
as mentioned above. The immune response is multimodal and adequate
to all types of antigens and stimuli.
There is also a cooperativity in the immune
response, as indicated in the first section and in Table I. This cooperation occurs between
the cells of the innate immunity, between the innate and adaptive
immunity and, finally, within the adaptive response (Fig. 2).
The immune system, with a complexity far beyond what
could be described in this review, is still working as a unit, but
a unit with adaptable modules (85). This unity is achieved through a
network of signals between cells, both soluble (interleukins,
chemokines), exosomal and through direct contact through cell
adhesion, co-stimulation and co-inhibition.
The immune system is extremely efficient. A great
body of experimental evidence shows that innate cells such as
macrophages, neutrophils, eosinophils, NK, NKT or γδ cells can be
strong tumoricidal elements, sometimes completely eradicating
tumors (86). Clinical evidence
from the above-mentioned prognostic association (30) and from new treatments such as
bispecific antibodies also shows that there is an extreme
efficiency of the lymphocyte when it faces the tumor cells
(87).
Unfortunately, this is not always the situation in
tumors. They are not eradicated, but continue to grow in spite of
such a powerful system that is directed against them.
What are the reasons for this situation? The answer
resides in the way in which the immune system and the tumors
interact.
Interaction between the immune system
and tumors
The first problem that the immune system encounters
is the nature of the tumor antigens: they are not true non-self
elements, but rather an altered self. There are also many stop
signals for immunocytes in tumors, such as CD-47 or PD-L1. The
tumor antigens are changing through mutations of an unstable
genome, another hallmark of cancer (80).
Another problem is represented by the
physico-chemical qualities of the environment in which these cells
work; in tumors there is acidosis, lactate, hypoxia, IDO and an
increased hydrostatic pressure; the blood vessels are modified and
do not offer enough cell adhesion molecules (CAMs) for
extravasation of leukocytes. By consequence, the immune response is
weakened and less efficient (77).
Furthermore, there is immune suppression in tumors;
the lymphocytes have to overcome this barrier as well, which they
do, but at the expense of losing much of the efficiency of their
response. They finally become exhausted and ineffective against
tumors (74).
A major problem is that innate immune cells, that
prove so tumoricidal in experiments, are subjects of tumor-secreted
factors that transform them into cells supporting the tumor. The
lymphocytes of the adaptive response lose the important support of
these cells, weakening once more in their capacity of response
(88).
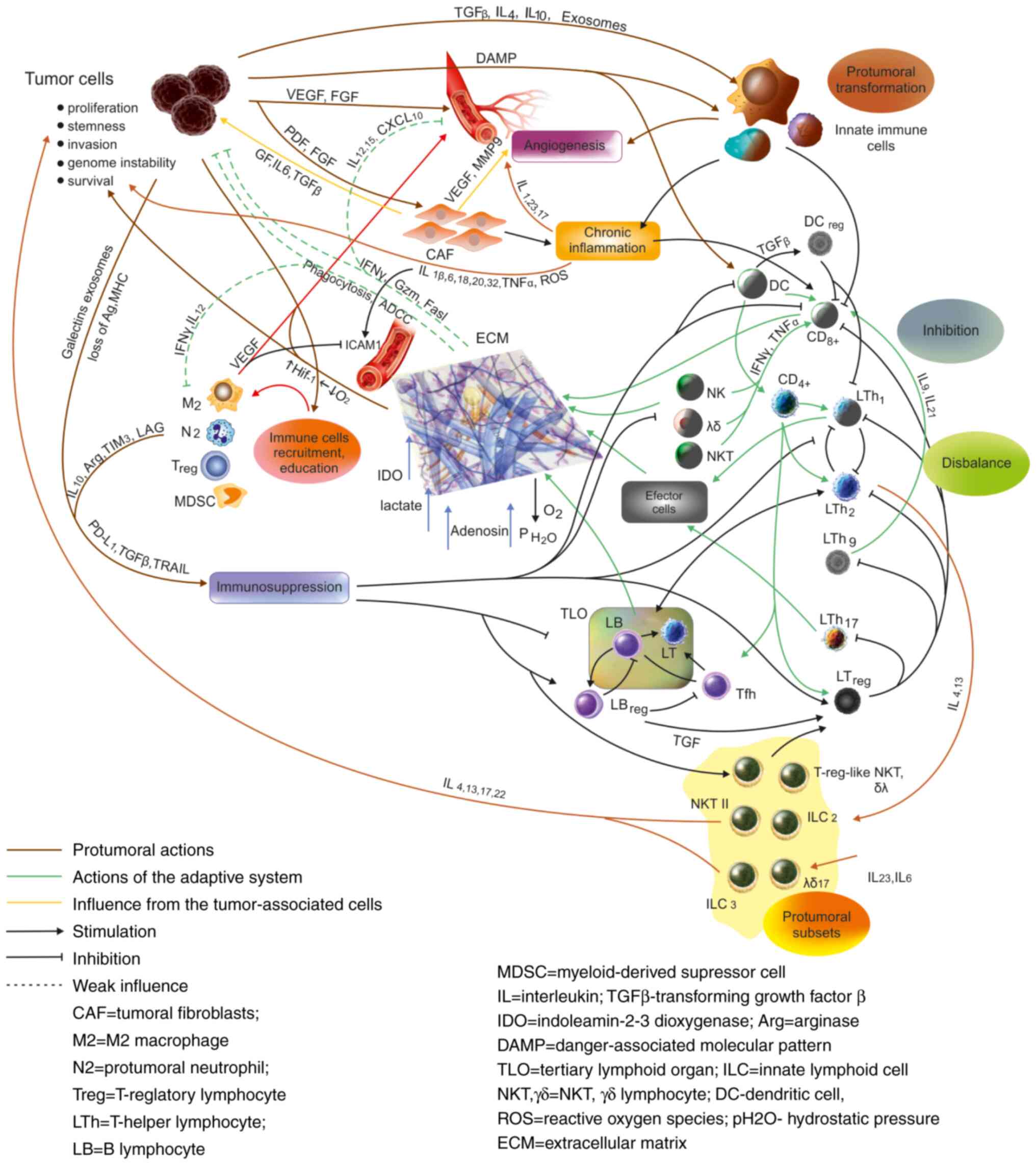
A seemingly paradoxical situation is that some of
the components of the immune response in tumors have protumoral
effects themselves; this is the case of the chronic, smoldering
inflammation that accompanies tumors, and also of some types of
adaptive response, the Th2, Th17 response, some γδ or NKT-cell
subsets, as mentioned earlier (Table
I and Fig. 3). This situation
is caused by the fact that interleukins can act as growth factors,
can promote angiogenesis especially in a situation of chronic
inflammation and, acting on epithelia, including tumor cells, they
can promote proliferation and cell survival (reviewed in 65).
Finally, the dysfunction of the dendritic cell in
tumors leads to a misbalance between lymphocyte subsets, with a
bias towards Th2 and regulatory subsets and a decrease of the
Th1-M1 subsets. As shown earlier, it is this misbalance that is
harmful to the antitumor defense, and in this situation, Th2 and
Th17 lose most of their antitumoral activity and become mainly
protumoral (42).
As a result, the immune system becomes gradually
inefficient and the tumors continue to grow.
Varieties of tumor
microenvironment
Tumors are heterogeneous structures and their TME
differs from one tumor to another. A recent study showed that there
are at least six types of TME in tumors, based on which of these
networks predominate, the subtype with angiogenesis, with
inflammation, interferon-dominant, TGFβ-dominant,
lymphocyte-depleted and immunologically quiet tumors. The authors
suggest that these data should be incorporated in the future
strategies of cancer therapy (89).
There are also differences between the different
locations of tumors (90), between
stages and even between patients. The type of carcinogen may also
cause differences concerning not only the genomic alterations, but
also the profile of the immune response that follows (91,92).
These differences between tumors should prompt the development of
more personalized approaches in immunotherapy (93). Personalized approaches are a
developing field, and they involve the use of biomarkers such as
tissue expression of checkpoint molecules, serum cytokines profile
or proteomic approaches to direct precision targeted therapy of
tumors (93,94).
The role of neuroendocrine
factors
To complete the picture of networks in tumors, the
role of the neuro-endocrine factors has to be mentioned; indeed,
there is experimental data that demonstrate an influence of the
nervous and endocrine system on the immune response (95). This is also true for the tumor
immunology, since both immunocytes (96) and cancer cells (97) possess receptors for catecholamines,
cortisol or different neuropeptides. This fact can be
therapeutically exploited, where there are receptors in the tumor.
It has been shown that through these receptors, the nervous and the
endocrine systems can modulate the tumoral growth and invasion
(98). The neuroendocrine factor,
and through it the psychological factor, proved to be not
neglectable factors in influencing the prognosis of patients with
tumors.
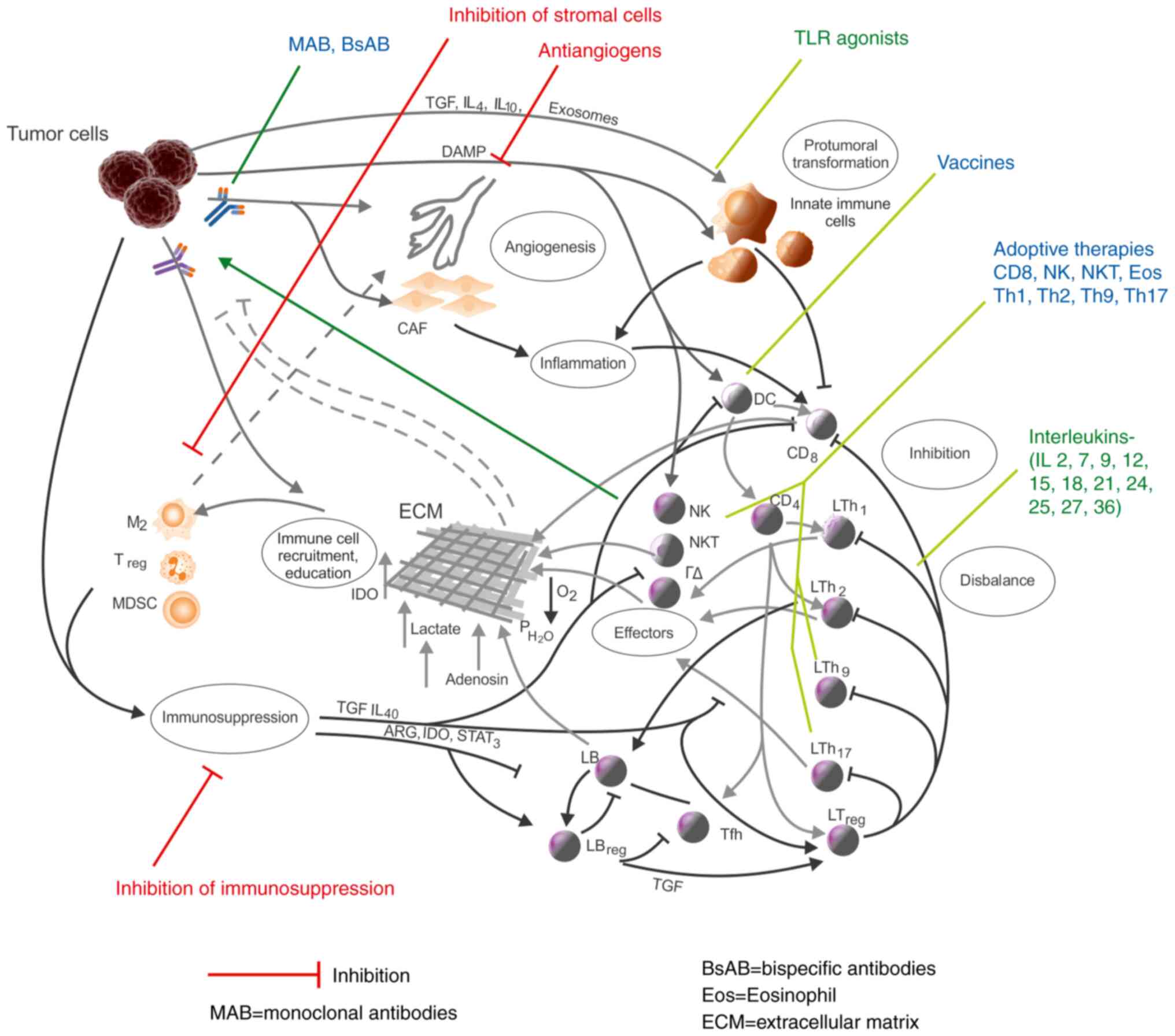
Networks and immunotherapy
A new factor, immunotherapy, has recently entered
this dynamic relation between tumors and the immune system.
One side of the immunotherapy is an inhibitory one,
which addresses the tumoral side of the environment; its targets
are angiogenesis (99), tumor
associated cells (100),
immuno-suppression (101) and
inflammation (70).
Another side is the positive immunotherapy, which
uses parts of the immune response to attack tumors. Antibodies
(monoclonal or bispecific) are used to direct immune cells against
the tumor cells, immune cells themselves are used in adoptive
therapy, vaccines are used to strengthen the antigen-specific
response, TLR agonists are used to stimulate innate cells, and
interleukins are used to stimulate the defense (101,102) (Fig.
4).
Immunotherapies must be considered in the larger
frame of immune networks in tumors; such a perspective opens the
way of new strategies, which result from the network structure of
the TME (103) and completes
intelligent approaches like bispecific antibodies, CAR-engineered
lymphocytes or the attempt to modify the antigenic interface of
tumors (104). Undoubtedly these
are efficient therapeutic means and have proven results, but they
also have limitations, which may, at least partially, be due to the
existence of these inhibitory loops that work in the TME.
Subsequently, the analysis of the immune networks in
tumors is an area of increasing interest, because it is expected to
offer solutions based on the understanding of the structure of
these networks in the TME.
4. Conclusions and perspectives
The present study is an attempt to decipher the
complex pro and antitumoral networks that form and interact in the
tumor microenvironment.
The study underlines the great potential of
immunotherapies; however, based on the existence of this network
structure of the TME, it suggests that therapeutic approaches
should be network-based and should take into account all these
complex interactions within the microenvironment of tumors.
At present, performant imaging and computational
approaches going as far as the single-cell level begin to enter
clinic (105), computer-based
learning is used to project anticancer molecules (106), but also network medicine begins to
enter all fields of pathology (107), including tumor immunology and
immunotherapy.
Immunotherapy is at its beginnings, but much
progress has been done in recent years, based on the growing
knowledge of immunobiology and genomics of malignant tumors.
Apart from the progress in the molecular and
cellular biology, the knowledge of the tumor microenvironment as a
whole, with its complex network of cells and cytokines, will
contribute to the development of the immune therapy in the years to
come.
Acknowledgements
Professional editing, linguistic and technical
assistance performed by Irina Radu, Individual Service Provider,
certified translator in Medicine and Pharmacy (certificate
credentials: Series E no. 0048).
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
OF contributed in the conception of the study and
wrote the manuscript. VC contributed in the conception of the study
and revised it critically for important intellectual content. Both
authors read and approved the final manuscript and agreed to be
accountable for all aspects of the work.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Egeblad M, Nakasone ES and Werb Z: Tumors
as organs: Complex tissues that interface with the entire organism.
Dev Cell. 18:884–901. 2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
de Visser KE, Eichten A and Coussens LM:
Paradoxical roles of the immune system during cancer development.
Nat Rev Cancer. 6:24–37. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Murphy K, Weaver C and Janeway C:
Janeway's immunobiology. 9th edition. New York, Garland Science,
2017.
|
|
4
|
Xue J, Schmidt SV, Sander J, Draffehn A,
Krebs W, Quester I, De Nardo D, Gohel TD, Emde M, Schmidleithner L,
et al: Transcriptome-based network analysis reveals a spectrum
model of human macrophage activation. Immunity. 40:274–288.
2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Aras S and Zaidi MR: TAMeless traitors:
Macrophages in cancer progression and metastasis. Br J Cancer.
117:1583–1591. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kawai O, Ishii G, Kubota K, Murata Y,
Naito Y, Mizuno T, Aokaje K, Saijo N, Nishiwaki Y, Gemma A, et al:
Predominant infiltration of macrophages and CD8(+) T cells in
cancer nests is a significant predictor of survival in stage IV
nonsmall cell lung cancer. Cancer. 113:1387–1395. 2008.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ryder M, Ghossein RA, Ricarte-Filho JC,
Knauf JA and Fagin JA: Increased density of tumor-associated
macrophages is associated with decreased survival in advanced
thyroid cancer. Endocr Relat Cancer. 15:1069–1074. 2008.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Georgoudaki AM, Prokopec KE, Boura VF,
Hellqvist E, Sohn S, Östling J, Dahan R, Harris RA, Rantalainen M,
Klevebring D, et al: Reprogramming tumor-associated macrophages by
antibody targeting inhibits cancer progression and metastasis. Cell
Rep. 15:2000–2011. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Phan SH: Biology of fibroblasts and
myofibroblasts. Proc Am Thorac Soc. 5:334–337. 2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chang HY, Chi JT, Dudoit S, Bondre C, van
de Rijn M, Botstein D and Brown PO: Diversity, topographic
differentiation, and positional memory in human fibroblasts. Proc
Nat Acad Sci USA. 99:12877–12882. 2002.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Dumont N, Liu B, Defilippis RA, Chang H,
Rabban JT, Karnezis AN, Tjoe JA, Marx J, Parvin B and Tlsty TD:
Breast fibroblasts modulate early dissemination, tumorigenesis, and
metastasis through alteration of extracellular matrix
characteristics. Neoplasia. 15:249–262. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Gascard P and Tlsty TD:
Carcinoma-associated fibroblasts: Orchestrating the composition of
malignancy. Genes Dev. 30:1002–1019. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Michiels C: Endothelial cell functions. J
Cell Physiol. 196:430–443. 2003.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang L, Yang N, Park JW, Katsaros D,
Franchiolli S, Cao G, O'Brien-Jenkins A, Randall TC, Rubin SC and
Coukos G: Tumor-derived vascular endothelial growth factor
upregulates angiopoietin-2 in host endothelium and destabilizes
host vasculature, supporting angiogenesis in ovarian cancer. Cancer
Res. 63:3403–3412. 2003.PubMed/NCBI
|
|
15
|
Konerding MA, Malkusch W, Klapthor B, van
Ackern C, Fait E, Hill SA, Parkins C, Chaplin DJ, Presta M and
Denekamp J: Evidence for characteristic vascular patterns in solid
tumours: Quantitative studies using corrosion casts. Br J Cancer.
80:724–732. 1999.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mócsai A: Diverse novel functions of
neutrophils in immunity, inflammation, and beyond. J Exp Med.
210:1283–1299. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Fridlender ZG, Sun J, Kim S, Kapoor V,
Cheng G, Ling L, Worthen GS and Albelda SM: Polarization of
tumor-associated neutrophil phenotype by TGF-beta: ‘N1’ versus ‘N2’
TAN. Cancer Cell. 16:183–194. 2009.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Nozawa H, Chiu C and Hanahan D:
Infiltrating neutrophils mediate the initial angiogenic switch in a
mouse model of multistage carcinogenesis. Proc Natl Acad Sci USA.
103:12493–12498. 2006.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jensen HK, Donskov F, Marcussen N,
Nordsmark M, Lundbeck F and von der Maase H: Presence of
intratumoral neutrophils is an independent prognostic factor in
localized renal cell carcinoma. J Clin Oncol. 27:4709–4717.
2009.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Walsh SR, Cook EJ, Goulder F, Justin TA
and Keeling NJ: Neutrophil-lymphocyte and ratio as a prognostic
factor in colorectal cancer. J Surg Oncol. 91:181–184.
2005.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Shamri R, Xenakis JJ and Spencer LA:
Eosinophils in innate immunity: An evolving story. Cell Tissue Res.
343:57–83. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Carretero R, Sektioglu IM, Garbi N,
Salgado OC, Beckhove P and Hämmerling GJ: Eosinophils orchestrate
cancer rejection by normalizin tumor vessels and enhancing
infiltration of CD8(+) T cells. Nat Immunol. 16:609–617.
2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Akdis M, Aab A, Altunbulakli C, Azkur K,
Costa RA, Crameri R, Duan S, Eiweger T, Eljaszewicz A, Ferstl R, et
al: Interleukins (from IL-1 to IL-38), interferons, transforming
growth factor β and TNF-α: Receptors, functions, and roles in
diseases. J Allergy Clin Immunol. 138:984–1010. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Mattes J, Hulett M, Xie W, Hogan S,
Rottenberg ME, Foster P and Parish C: Immunotherapy of cytotoxic T
cell-resistant tumors by T helper 2 cells: An eotaxin and
STAT6-dependent process. J Exp Med. 197:387–393. 2003.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tepper RI, Pattengale PK and Leder P:
Murine interleukin-4 displays potent anti-tumor activity in
vivo. Cell. 57:503–512. 1989.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Fernández-Aceñero MJ, Galindo-Gallego M,
Sanz J and Aljama A: Prognostic influence of tumor-associated
eosinophilic infiltrate in colorectal carcinoma. Cancer.
88:1544–1548. 2000.PubMed/NCBI
|
|
27
|
Krystel-Whittemore M, Dileepan KN and Wood
JG: Mast cell: A multi-functional master cell. Front Immunol.
6(620)2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Varricchi G, Galdiero MR, Loffredo S,
Marone G, Iannone R, Marone G and Granata F: Are mast cells MASTers
in cancer? Front Immunol. 8(424)2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Oldford SA, Haidl ID, Howatt MA, Leiva CA,
Johnston B and Marshall JS: A critical role for mast cells and mast
cell-derived IL-6 in TLR2-mediated inhibition of tumor growth. J
Immunol. 185:7067–7076. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Gentles AJ, Newman AM, Liu CL, Bratman SV,
Feng W, Kim W, Nair SW, Xu H, Khuong A, Hoang CD, et al: The
prognostic landscape of genes and infiltrating immune cells across
human cancers. Nat Med. 21:938–945. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Patil RS, Shah SU, Shrikhande SV, Goel M,
Dikshit RP and Chiplunkar SV: IL17 producing γδT cells induce
angiogenesis and are associated with poor survival in gallbladder
cancer patients. Int J Cancer. 139:869–881. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhao Y, Niu C and Cui J: Gamma-delta (γδ)
T cells: Friend or foe in cancer development? J Transl Med.
16(3)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Dhodapkar MV and Kumar V: Type II NKT
cells and their emerging role in health and disease. J Immunol.
198:1015–1021. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Terabe M, Matsui S, Noben-Trauth N, Chen
H, Watson C, Donaldson DD, Carbone DP, Paul WE and Berzofsky JA:
NKT cell-mediated repression of tumor immunosurveillance by IL-13
and the IL-4R-STAT6 pathway. Nat Immunol. 1:515–520.
2000.PubMed/NCBI View
Article : Google Scholar
|
|
35
|
Terabe M and Berzofsky JA: The role of NKT
cells in tumor immunity. Adv Cancer Res. 101:277–348.
2008.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yang Q, Goding SR, Hokland ME and Basse
PH: Antitumor activity of NK cells. Immunol Res. 36:13–25.
2006.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Minetto P, Guolo F, Pesce S, Greppi M,
Obino V, Ferretti E, Sivori S, Genova C, Lemoli RM and Marcenaro E:
Harnessing NK cells for cancer treatment. Front Immunol.
10(2836)2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
van Beek JJP, Martens AWJ, Bakdash G and
de Vries IJM: Innate lymphoid cells in tumor immunity.
Biomedicines. 4(7)2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Eisenring M, vom Berg J, Kristiansen G,
Saller E and Becker B: IL-12 initiates tumor rejection via lymphoid
tissue-inducer cells bearing the natural cytotoxicity receptor
NKp46. Nat Immunol. 11:1030–1038. 2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Mellman I: Dendritic cells: Master
regulators of the immune response. Cancer Immunol Res. 1:145–149.
2013.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lijun Z, Xin Z, Danhua S, Xiaoping L,
Jianliu W, Huilan W and Lihui W: Tumor-infiltrating dendritic cells
may be used as clinicopathologic prognostic factors in endometrial
carcinoma. Int J Gynecol Cancer. 22:836–841. 2012.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Ma Y, Shurin GV, Peiyuan Z and Shurin MR:
Dendritic cells in the cancer microenvironment. J Cancer. 4:36–44.
2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhan Y and Wu L: Functional regulation of
monocyte-derived dendritic cells by microRNAs. Protein Cell.
3:497–507. 2012.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Ostroumov D, Fekete-Drimusz N, Saborowski
M, Kühnel F and Woller N: CD4 and CD8 T lymphocyte interplay in
controlling tumor growth. Cell Mol Life Sci. 75:689–713.
2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Woo SR, Turnis ME, Goldberg MV, Bankoti J,
Selby M, Nirschl CJ, Bettini ML, Gravano DM, Vogel P, Liu CL, et
al: Immune inhibitory molecules LAG-3 and PD-1 synergistically
regulate T-cell function to promote tumoral immune escape. Cancer
Res. 72:917–927. 2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ziai J, Gilbert HN, Foreman O,
Eastham-Anderson J, Chu F, Huseni M and Kim JM: CD8+ T
cell infiltration in breast and colon cancer: A histologic and
statistical analysis. PLoS One. 13(e0190158)2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Durgeau A, Virk Y, Corgnac S and
Mami-Chouaib F: Recent advances in targeting CD8 T-cell immunity
for more effective cancer immunotherapy. Front Immunol.
9(14)2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Akbulut G, Özkazanç D and Esendağlı G: Th1
cells in cancer-associated inflammation. Turk J Biol. 41:20–30.
2017.
|
|
49
|
Bos R and Sherman LA: CD4+
T-cell help in the tumor milieu is required for recruitment and
cytolytic function of CD8+ T lymphocytes. Cancer Res.
70:8368–8377. 2010.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Fujisawa T, Joshi BH and Puri RK: IL-13
regulates cancer invasion and metastasis through IL-13Rα2 via
ERK/AP-1 pathway in mouse model of human ovarian cancer. Int J
Cancer. 131:344–356. 2012.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Nishimura T, Iwakabe K, Sekimoto M, Ohmi
Y, Yahata T, Nakui M, Sato T, Habu S, Tashiro H, Sato M and Ohta A:
Distinct role of antigen-specific t helper type 1 (Th1) and Th2
cells in tumor eradication in vivo. J Exp Med. 190:617–627.
1999.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Lorvik KB, Hammarström C, Fauskanger M,
Haabeth OA, Zangani M, Haraldsen G, Bogen B and Corthay A: Adoptive
transfer of tumor-specific Th2 cells eradicates tumors by
triggering an in situ inflammatory immune response. Cancer
Res. 76:6864–6876. 2016.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Amicarella F, Muraro MG, Hirt C, Cremonesi
E, Padovan E, Mele V, Governa V, Han J, Huber X, Droeseret RA, et
al: Dual role of tumour-infiltrating T helper 17 cells in human
colorectal cancer. Gut. 66:692–704. 2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Lee YK, Turner H, Maynard CL, Oliver JR,
Chen D, Elson CO and Weaver CT: Late developmental plasticity in
the T helper 17 lineage. Immunity. 30:92–107. 2009.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Kryczek I, Banerjee M, Cheng P, Vatan L,
Szeliga W, Wei S, Huang E, Finlayson E, Simeone D, Welling TH, et
al: Phenotype, distribution, generation, and functional and
clinical relevance of Th17 cells in the human tumor environments.
Blood. 114:1141–1149. 2009.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Martin-Orozco N, Muranski P, Chung Y, Yang
XO, Yamazaki T, Lu S, Hwu P, Restifo NP, Overwijk WW and Dong C: T
helper 17 cells promote cytotoxic T cell activation in tumor
immunity. Immunity. 31:787–798. 2009.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Lu Y, Hong S, Li H, Park J, Hong B, Wang
L, Zheng Y, Liu Z, Xu J, He J, et al: Th9 cells promote antitumor
immune responses in vivo. J Clin Invest. 122:4160–4171.
2012.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kim K, Kim G, Kim JY, Yun HJ, Lim SC and
Choi HS: Interleukin-22 promotes epithelial cell transformation and
breast tumorigenesis via MAP3K8 activation. Carcinogenesis.
35:1352–1361. 2014.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Gu-Trantien C, Loi S, Garaud S, Equeter C,
Libin M, de Wind A, Ravoet M, Le Buanec H, Sibille C,
Manfouo-Foutsop G, et al: CD4' follicular helper T cell
infiltration predicts breast cancer survival. J Clin Invest.
123:2873–2892. 2013.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Verma A, Mathur R, Farooque A, Kaul V,
Gupta S and Dwarakanath BS: T-regulatory cells in tumor progression
and therapy. Cancer Manag Res. 11:10731–10747. 2019.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Facciabene A, Motz GT and Coukos G:
T-regulatory cells: Key players in tumor immune escape and
angiogenesis. Cancer Res. 72:2162–2171. 2012.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Correale P, Rotundo MS, Del Vecchio MT,
Remondo C, Migali C, Ginnaneschi C, Tsang KY, Lichetta A, Manucci
S, Loiacono L, et al: Regulatory (FoxP3+) T-cell tumor
infiltration is a favorable prognostic factor in advanced colon
cancer patients undergoing chemo or chemoimmunotherapy. J
Immunother. 33:435–441. 2010.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Wouters MCA and Nelson BH: Prognostic
significance of tumor-infiltrating B cells and plasma cells in
human cancer. Clin Cancer Res. 24:6125–6135. 2018.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Yuen GJ, Demissie E and Pillai S: B
lymphocytes and cancer: A love-hate relationship. Trends Cancer.
2:747–757. 2016.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Farc O and Cristea V: Pro-and antitumor
role of interleukins 1 to 41. Roum Arch Microbiol Immunol.
78:149–162. 2019.
|
|
66
|
Figueras A, Arbos MA, Quiles MT, Viñals F,
Germà JR and Capellà G: The impact of KRAS mutations on VEGF-A
production and tumour vascular network. BMC Cancer.
13(125)2013.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Talks KL, Turley H, Gatter KC, Maxwell PH,
Pugh CW, Ratcliffe PJ and Harris AL: The expression and
distribution of the hypoxia-inducible factors HIF-1alpha and
HIF-2alpha in normal human tissues, cancers, and tumor-associated
macrophages. Am J Pathol. 157:411–421. 2000.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Hill BS, Sarnella A, D'Avino G and
Zannetti A: Recruitment of stromal cells into tumour
microenvironment promote the metastatic spread of breast cancer.
Semin Cancer Biol. 60:202–213. 2020.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Austenaa L and Natoli G: A shortcut for
early macrophage recruitment into tumors by activated oncogenes.
Genes Dev. 31:223–225. 2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444.
2008.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Wörmann SM, Diakopoulos KN, Lesina M and
Algül H: The immune network in pancreatic cancer development and
progression. Oncogene. 33:2956–2967. 2014.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Le Naour A, Prat M, Thibault B, Mevel R,
Lemaître L, Leray H, Joubert MV, Coulson K, Golzio M, Lefevre L, et
al: Tumor cells educate mesenchymal stromal cells to release
chemoprotective and immunomodulatory factors. J Mol Cell Biol.
12:202–215. 2020.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Bosiljcic M, Cederberg RA, Hamilton MJ,
LePard NE, Harbourne BT, Collier JL, Halvorsen EC, Shi R, Franks
SE, Kim AY, et al: Targeting myeloid-derived suppressor cells in
combination with primary mammary tumor resection reduces metastatic
growth in the lungs. Breast Cancer Res. 21(103)2019.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Dunn GP, Old LJ and Schreiber RD: The
three Es of cancer immunoediting. Annu Rev Immunol. 22:329–360.
2004.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Casey SC, Li Y and Felsher DW: An
essential role for the immune system in the mechanism of tumor
regression following targeted oncogene inactivation. Immunol Res.
58:282–291. 2014.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Casey SC, Tong L, Li Y, Do R, Walz S,
Fitzgerald KN, Gouw AM, Baylot V, Gütgemann I, Eilers M and Felsher
DW: MYC regulates the antitumor immune response through CD47 and
PD-L1. Science. 352:227–231. 2016.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Xiong GF and Xu R: Function of cancer
cell-derived extracellular matrix in tumor progression. J Cancer
Metastasis Treat. 2:357–364. 2016.
|
|
78
|
Agrawal N, Bettegowda C, Cheong I,
Geschwind JF, Drake CG, Hipkiss EL, Tatsumi M, Dang LH, Diaz LA Jr,
Pomper M, et al: Bacteriolytic therapy can generate a potent immune
response against experimental tumors. Proc Natl Acad Sci.
101:15172–15177. 2004.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Haabeth OA, Bogen B and Corthay A: A model
for cancer - suppressive inflammation. Oncoimmunology. 1:1146–1155.
2012.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Schäfer M and Werner S: Cancer as an
overhealing wound: An old hypothesis revisited. Nat Rev Mol Cell
Biol. 9:628–63879. 2008.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Riss J, Khanna C, Koo S, Chandramoulli DV,
Yang HH, Hu Y, Kleiner DE, Rosenward A, Schaeffer CF, Ben-Sasson
SA, et al: Cancers as wounds that do not heal: Differences and
similarities between renal regeneration/repair and renal cell
carcinoma. Cancer Res. 66:7216–7224. 2006.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Pesic M and Greten FR: Inflammation and
cancer: Tissue regeneration gone awry. Curr Opin Cell Biol.
43:55–61. 2016.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Naga A, Siddiqui A and Bindu H: Immuno
defense mechanism against tumors. J Cancer Sci Ther. 17(2)2011.
|
|
85
|
Kondratova M, Czerwinska U, Sompairac N,
Amigorena SD, Soumelis V, Barillot E, Zinovyev A and Kuperstein I:
A multiscale signalling network map of innate immune response in
cancer reveals cell heterogeneity signatures. Nat Commun.
10(4808)2019.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Liu Y and Zeng G: Cancer and innate immune
system interactions: Translational potentials for cancer
immunotherapy. J Immunother. 35:299–308. 2012.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Wong R, Pepper C, Brennan P, Nagorsen D,
Man S and Fegan C: Blinatumomab induces autologous T-cell killing
of chronic lymphocytic leukemia cells. Haematologica. 98:1930–1938.
2013.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Burkholder B, Huang RY, Burgess R, Luo S,
Jones VS, Zhang W, Lu ZQ, Gao CY, Wang BL, Zhang YM and Huang RP:
Tumor-induced perturbations of cytokines and immune cell networks.
Biochim Biophys Acta. 1845:182–201. 2014.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Thorsson V, Gibbs DL, Brown SD, Wolf D,
Bortone DS, Ou Yang TH, Porta-Pardo E, Gao GF, Plaisier CL, Eddy
JA, et al: The immune landscape of cancer. Immunity.
48:812–830.e14. 2018.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Ringelhan M, Pfister D, O'Connor T,
Pikarsky E and Heikenwalder M: The immunology of hepatocellular
carcinoma. Nat Immunol. 19:222–232. 2018.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Neagu M, Caruntu C, Constantin C, Boda D,
Zurac S, Spandidos DA and Tsatsakis AM: Chemically induced skin
carcinogenesis: Updates in experimental models (Review). Oncol Rep.
35:2516–2528. 2016.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Boda D, Docea AO, Calina D, Ilie MA,
Caruntu C, Zurac S, Neagu M, Constantin C, Branisteanu DE,
Voiculescu V, et al: Human papilloma virus: Apprehending the link
with carcinogenesis and unveiling new research avenues. Int J
Oncol. 52:637–655. 2018.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Palucka AK and Coussens LM: The basis of
oncoimmunology. Cell. 164:1233–1247. 2016.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Ion A, Popa IM, Papagheorghe LM, Lisievici
C, Lupu M, Voiculescu V, Caruntu C and Boda D: Proteomic approaches
to biomarker discovery in cutaneous T-cell lymphoma. Dis Markers.
2016(9602472)2016.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Grigore O, Mihailescu AI, Solomon I, Boda
D and Caruntu C: Role of stress in modulation of skin neurogenic
inflammation. Exp Ther Med. 17:997–1003. 2019.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Solomon I, Voiculescu VM, Caruntu C, Lupu
M, Popa A, Ilie MA, Albulescu R, Caruntu A, Tanase C, Constantin C,
et al: Neuroendocrine factors and head and neck squamous cell
carcinoma: An affair to remember. Dis Markers.
2018(9787831)2018.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Caruntu C, Boda D, Constantin C, Caruntu A
and Neagu M: Catecholamines increase in vitro proliferation of
murine B16F10 melanoma cells. Acta Endocrinol. 10:545–558.
2014.
|
|
98
|
Lupu M, Caruntu A, Caruntu C, Papagheorghe
LML, Ilie MA, Voiculescu V, Boda D, Constantin C, Tanase C, Sifaki
M, et al: Neuroendocrine factors: The missing link in non melanoma
skin cancer (Review). Oncol Rep. 38:1327–1340. 2017.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Maj E, Papiernik D and Wietrzyk J:
Antiangiogenic cancer treatment: The great discovery and greater
complexity (Review). Int J Oncol. 49:1773–1784. 2016.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Cotechini T, Medler TR and Coussens LM:
Myeloid cells as targets for therapy in solid tumors. Cancer J.
21:343–350. 2015.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Kruger S, Ilmer M, Kobold S, Cadilha BL,
Endres S, Ormanns S, Schuebe G, Renz BW, D'Haese JG, Schloesser H,
et al: Advances in cancer immunotherapy 2019 - latest trends. J Exp
Clin Cancer Res. 38(268)2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Farkona S, Diamandis EP and Blasutig IM:
Cancer immunotherapy: The beginning of the end of cancer? BMC Med.
14(73)2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Clancy T and Hovig E: Profiling networks
of distinct immune-cells in tumors. BMC Bioinformatics.
17(263)2016.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Munks M, Levitsky V, Hill A and Knoetgen
H: Cytomegalovirus-specific CD8 T cells kill B16 melanoma cells in
vivo whe activated by bifunctional major histocompatibility class
I-antibody fusion molecules (pMHCI-IgGs). J Immunotherapy Cancer.
3(237)2015.
|
|
105
|
Boda D: Cellomics as integrative omics for
cancer. Curr Proteomics. 10:237–245. 2013.
|
|
106
|
Ancuceanu R, Dinu M, Neaga I, Laszlo FG
and Boda D: Development of QSAR machine learning-based models to
forecast the effect of substances on malignant melanoma cells.
Oncol Lett. 17:4188–4196. 2019.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Robin X, Creixell P, Radetskaya O, Santini
CC, Longden J and Linding R: Personalized network-based treatments
in oncology. Clin Pharmacol Ther. 94:646–650. 2013.PubMed/NCBI View Article : Google Scholar
|