Introduction
Ethylparaben is a widely used preservative in
Chinese eye drops (1,2). Subconjunctival fibrosis (SCF) leads to
failure of glaucoma filtration surgery, inducing conjunctival
scarring during wound healing (3-5).
Nevertheless, the action of ethylparaben in SCF remains to be
elucidated.
The Wnt/β-catenin axis participates in development
and in tissue homeostasis, and is divided into canonical and
non-canonical signaling (6,7). Abnormal stimulation of the canonical
Wnt pathway stimulates various pathological states including
malignancies (8). The transcription
factor complex, consisting of β-catenin, T cell factor and lymphoid
enhancer factor, is widely recognized as the main modulator of the
Wnt/β-catenin axis (9,10). When the Wnt ligand is not activated,
a destroying complex consisting of the tumor suppressor adenomatous
polyposis coli, axin and glycogen synthase kinase-3 (GSK3β) anchors
cytoplasmic β-catenin (11-13).
GSK3β and casein kinase 1 participate in the phosphorylation of
cytoplasmic β-catenin, bringing about ubiquitination and
degeneration of proteasome (14,15).
When the Wnt ligand is activated, it links with the Frizzled
receptor and its co-receptor, low-density lipoprotein, by a
receptor-related protein 5/6 on the surface of the cell. This
propels dishevelled to attract GSK3β and Axin, consequently
separating β-catenin from the destroying complex (16,17).
Degeneration and phosphorylation of β-catenin is subsequently
suppressed prior to aggregation of cytoplasmic β-catenin and
nuclear translocation (18). This
association induces the expression of target genes in the
Wnt/β-catenin axis. Previous studies have demonstrated that the
Wnt/β-catenin axis participates in corneal fibrosis (19,20).
Nevertheless, the current understanding of the influence of
stimulation of the Wnt/β-catenin axis on SCF triggered by
ethylparaben is insufficient.
The present study explored the etiology of SCF
triggered by ethylparaben through examination of the concentration
of the extracellular matrix (ECM) and agents linked with the
Wnt/β-catenin axis in vivo and in vitro. It was
discovered that deposition of collagen in murine conjunctival
subepithelium was triggered by ethylparaben supplemented for one
month, and that the Wnt/β-catenin axis was crucial to this
reaction.
Materials and methods
Materials
Dulbecco's Modified Eagle Medium (DMEM) and fetal
bovine serum (FBS) were purchased from Invitrogen; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Ethylparaben was provided by
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). XAV-939 was
obtained from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA) and
used at 1 µM.
Animals and ethylparaben
treatment
Experiments involving animal subjects in the present
study were conducted in accordance with the internationally
accepted principles for laboratory animal use and care. The present
study was approved by the Committee on the Ethics of Animal
Experiments of Zoucheng People's Hospital (Zoucheng, China).
Sprague-Dawley rats (age, 6 weeks; weight, 215±30 g) were obtained
from the Beijing Vital River Laboratory Animal Technology Co., Ltd.
(Beijing, China) and used in the present study. Rats were housed in
microisolator cages in a room illuminated from 0700-1900 h (12-h
light/dark cycle; humidity, 55±5%; 25±2˚C), with ad libitum
access to water and chow. Animal procedures were carried out in
conformity to guidelines of the ARVO Statement for the Use of
Animals in Ophthalmic and Vision Research (21). Each rat was verified for ocular
surface illness prior to experimentation. A total of 18 rats were
randomized into 3 groups (n=6/group). In the first group, 0.01%
ethylparaben in PBS was dropped locally into the left eye (twice
daily, 10 µl each). In the second group (control group), PBS was
supplemented the same way (twice daily, 10 µl each). All rats were
executed 1 month following supplementation. Bulbar conjunctival
tissues (BCTs) were excised to separate RNA and protein. Rats in
the third group received no treatment prior to sacrifice and their
left eyes were enucleated under aseptic circumstances for
separation of bulbar subconjunctival tissues (BSTs).
Cultivation of primary conjunctival
fibroblasts (CFs)
Primary CFs were separated from BSTs and cultivated.
BSTs separated under aseptic circumstances were washed thrice with
PBS. BSTs were subsequently processed into 2x2-mm explants, which
were planted in cultivating plates at a density of 5x103
cells/well filled with DMEM and 10% FBS and were incubated in an
environment with 5% CO2, 95% humidity, and 37˚C. The
medium was replaced every three days. CFs were subcultivated with
trypsin for procedures between passages five and seven.
In order to examine the influence of low levels of
ethylparaben on CF survival and to explore the optimal ethylparaben
level for examining the etiology of ECM gene expression of CFs
activated by ethylparaben, CFs were planted in plates with 96 wells
(3x103 cells/well) and received media with either
ethylparaben (0.000005%, 0.00001%, 0.00003%, 0.00005%, 0.0001%,
0.001%) or PBS at 37˚C.
Analysis of β-catenin nuclear
translocation
CFs were treated with ethylparaben for 4 h. Nuclear
fractionation was used to analyze β-catenin nuclear translocation.
Briefly, nuclear extracts were isolated from CFs treated in
75-cm2 flasks using the NEPER Nuclear/Cytoplasmic
Extraction Kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions, and analyzed by β-catenin WB
analysis.
Western blotting (WB)
Western blotting was performed as previously
detailed (22-24).
The total proteins were extracted from BCTs, and the CFs were
extracted using a cold radioimmunoprecipitation assay buffer that
contains a proteinase inhibitor cocktail (Roche Diagnostics, Basel,
Switzerland). Protein concentration was determined using a Piece
BCA Protein Assay kit (Thermo Fisher Scientific, Inc.). Equivalent
quantities of proteins (50 µg) harvested from CFs and conjunctival
tissues were subjected to electrophoresis with 10% SDS-PAGE.
Proteins were transferred onto polyviylidene difluoride membranes.
Subsequently, the membranes were blocked at room temperature for 1
h using 5% not-fat milk in tris-buffered saline and incubated with
primary antibodies against: α-smooth muscle actin (SMA; cat. no.
ab5694), collagen 1 (cat. no. ab138492), fibronectin (FN)-1 (cat.
no. ab2413; all Abcam, Cambridge, UK), β-actin (cat. no. 3700),
collagen type 1 (Col1)α1 (cat. no. 84336), phosphorylated
(p)-β-catenin (cat. no. 4176), β-catenin (cat. no. 8480), Lamin A/C
(cat. no. 4777; all Cell Signaling Technology, Danvers, MA, USA) at
1:1,000 dilution and 4˚C overnight. The membranes were washed and
then incubated with horseradish peroxidase-conjugated secondary
antibodies (1:10,000; cat. no. ab6721; Abcam,) for 1 h at room
temperature. Finally, an enhanced chemiluminescence ECL Detection
kit (Thermo Fisher Scientific, Inc.) was utilized to evaluate the
bands, and a transilluminator (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was used to examine the intensity of the
image.
Cell survival assessment
A cell counting kit-8 (CCK-8; Sigma-Aldrich; Merck
KGaA) was used to assess survival. The media was replaced with
fresh media with CCK-8 solution. Following a 2-h period of
incubation at 37˚C, the microplate reader was used to
spectrophotometrically assess absorbance at 450 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to isolate total RNA from BCTs, which
then underwent reverse transcription to cDNA using a First Stand
cDNA synthesis kit (Sigma-Aldrich; Merck KGaA). qPCR was performed
in triplicate using SYBR Premix Ex Taq (Takara Biotechnology Co.,
Ltd., Dalian, China) and SsoFast™ Probes Supermix
(Bio-Rad Laboratories, Inc.). The following thermocycling
conditions were used for the RT-qPCR: Initial denaturation at 95˚C
for 1 min; 38 cycles for 95˚C for 1 min, 58˚C for 1 min, 72˚C for 1
min; and a terminal extension at 72˚C for 10 min. The outcome of
RT-qPCR was evaluated using the 2-ΔΔCq approach
(25) and was normalized in
conformity with β-actin. The primers used were as follows: Col1α1,
forward 5'-CCTCCCAGAACATTACATA-3' and reverse
5'-GACTGTCTTGCTCCATTCACCA-3'; FN-1, forward
5'-GACTCGTTTGACTTTGACTTCACCAC-3' and reverse
5'-GCTGAGACCCAGGAGACCAC-3'; collagen 1, forward
5'-TACAGCACGCTTGTGGATG-3' and reverse 5'-CAGATTGGGATGGAGGGAGTT-3';
α-SMA, forward 5'-GAGGCACCACTGAACCCTAA-3' and reverse
5'-CATCTCCAGAGTCCAGCACA-3'; β-catenin, forward
5'-GCTGACCTGATGGAGTTGGA-3' and reverse 5'-GCTACTTGCTCTTGCGTGAA-3';
and β-actin, forward 5'-GACGTTGACATCCGTAAAGACC-3' and reverse
5'-CTAGGAGCCAGGGCAGTAATCT-3'.
Hematoxylin and eosin (H&E)
staining
H&E staining was carried out in 5-µm-thick
paraffin-embedded sections of murine eye specimens, which were
fixed with 4% paraformaldehyde at 25˚C for 30 min. The slices
underwent deparaffinization (xylene, 3 min) and rehydration (100%
ethanol, 3 min; 95% ethanol, 3 min; 70% ethanol, 3 min; 50%
ethanol, 3 min) prior to H&E staining (1 min/each) for
histopathological observation at 25˚C. A Nikon Diaphot 200 inverted
microscope (Nikon Corporation, Tokyo, Japan) was used to evaluate
images.
Statistical analysis
Data are presented as the mean ± standard deviation.
Student's t-test was used to identify the significance of
differences between two groups, and one-way analysis of variance
followed by Tukey's test was used for analysis of multiple groups.
P<0.05 was considered to indicate a statistically significant
difference. GraphPad Prism IV software (GraphPad Software, Inc., La
Jolla, CA, USA) was used for analyses.
Results
Chronic supplement of 0.01%
ethylparaben induces murine bulbar SCF
In order to examine the influence of chronic
supplementation of 0.01% ethylparaben on SCF in murine bulbar
conjunctivas (BCs), the present study investigated slices of murine
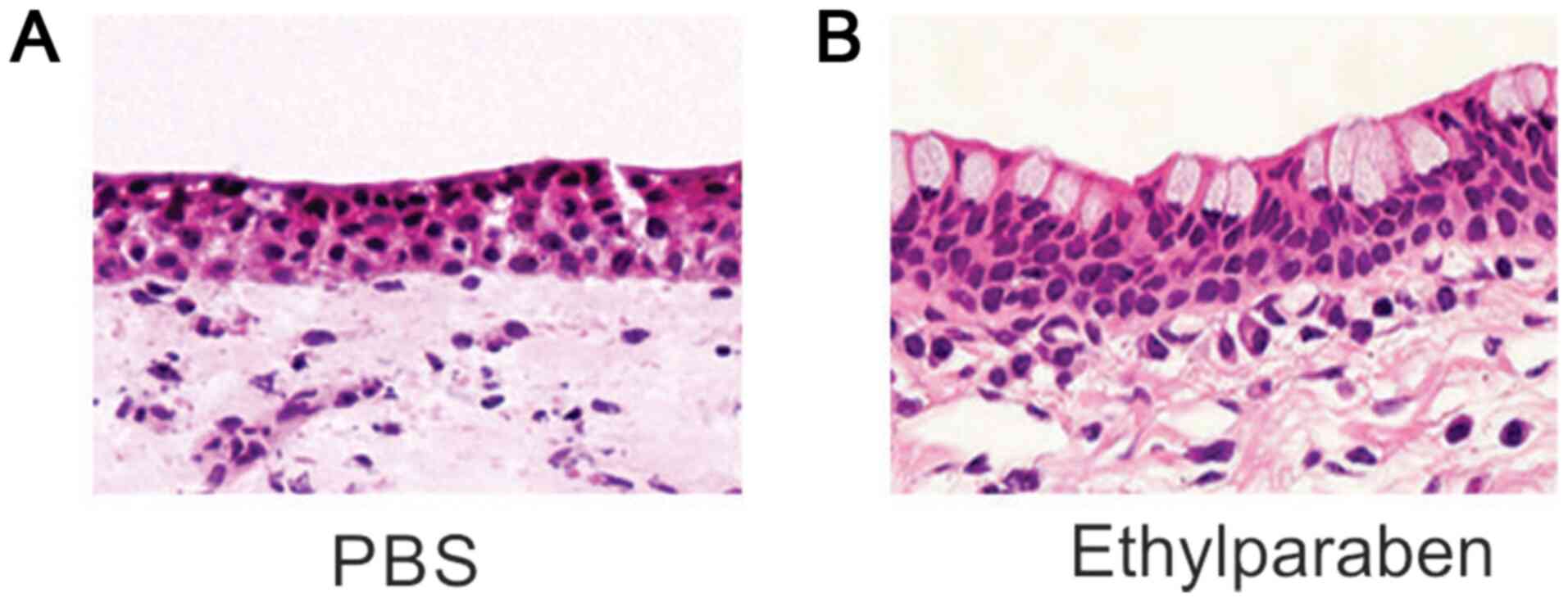
eye specimens that underwent H&E staining. H&E staining
demonstrated that the density of fibroblasts was mildly promoted
and the deposition of collagen was tighter in the subepithelium of
BCs in the experimental group than the control group (Fig. 1). These findings suggested that
chronic supplementation of ethylparaben triggered bulbar
subconjunctival fibrosis (BSF).
Chronic supplementation of 0.01%
ethylparaben enhances the expression of ECM in BCT
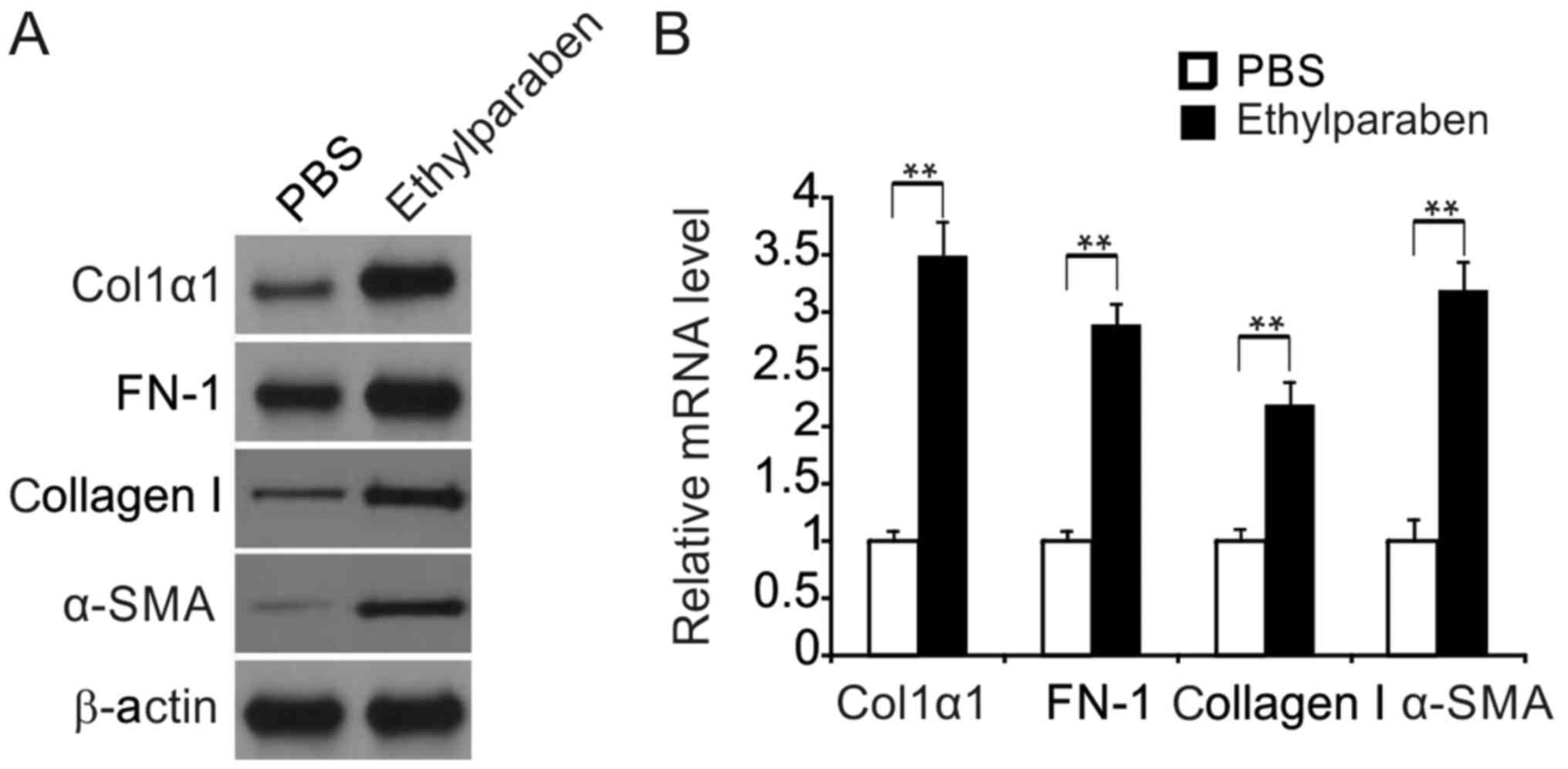
In order to verify that BSF triggered by chronic
supplementation of 0.01% ethylparaben resulted from increased
collagen production and other ECM material in BCTs, FN-1 and Col1α1
expression was evaluated in murine BCTs using WB. Expression of
FN-1, Col1α1, collagen I and α-SMA were markedly promoted in the
experimental group compared with the control group (Fig. 2A). The mRNA level of these genes was
also promoted in the experimental group according to RT-qPCR
(Fig. 2B).
Chronic supplementation of 0.01%
ethylparaben activates the expression of agents linked with the
Wnt/β-catenin axis in BCTs
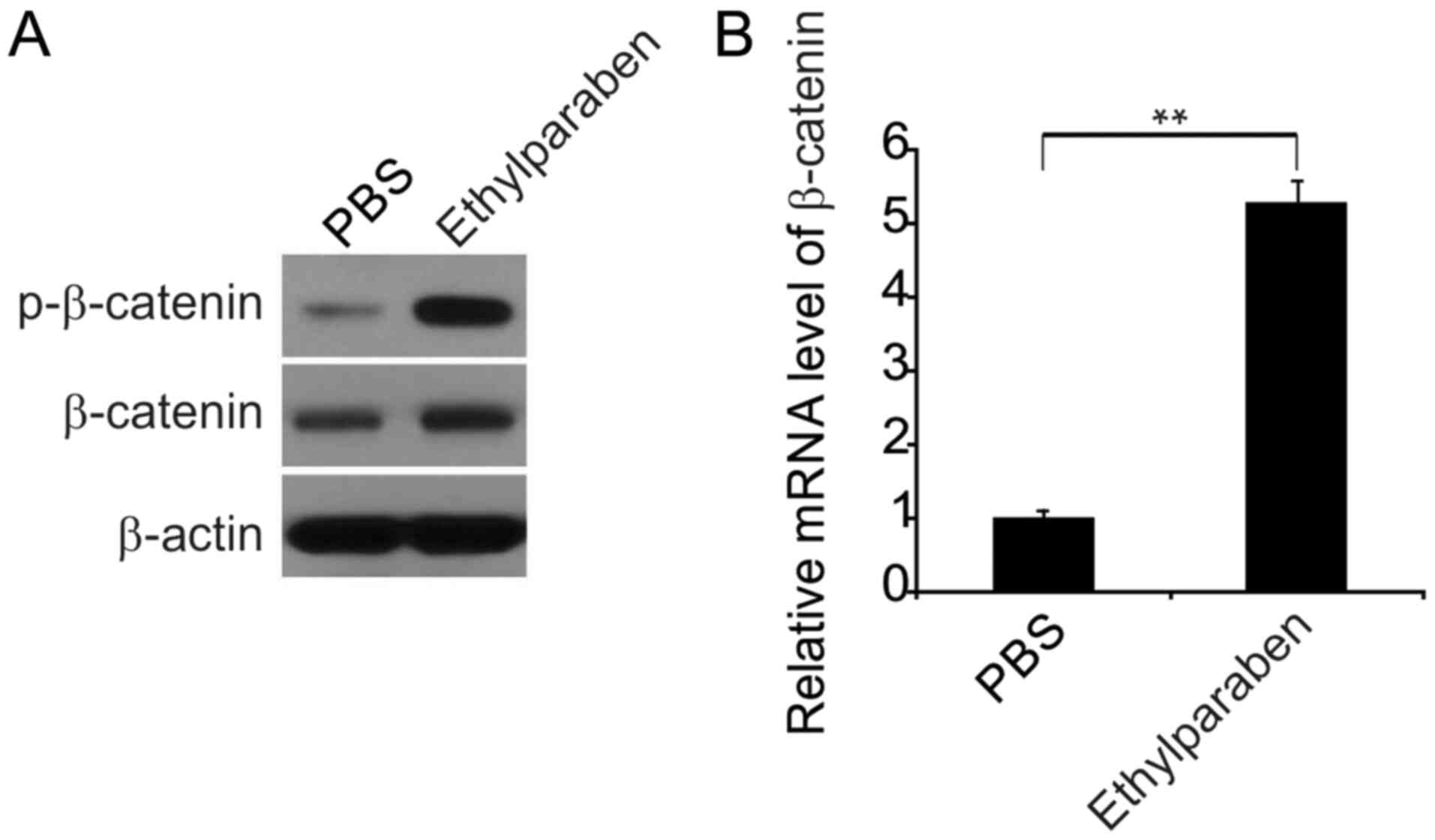
It has been demonstrated previously that the
Wnt/β-catenin axis participates in fibrosis (26,27).
To explore the stimulation of the Wnt/β-catenin axis by chronic
treatment with 0.01% ethylparaben, the expression of agents linked
with the β-catenin pathway was explored in murine BCTs using WB. It
was observed that both p- and total β-catenin levels were markedly
upregulated in the ethylparaben-treated group compared with the
control group (Fig. 3A), and that
the β-catenin transcription was noticeably promoted in the
experimental group according to RT-qPCR (Fig. 3B). These findings suggested that
chronic supplementation of 0.01% ethylparaben was able to trigger
stimulation of the Wnt/β-catenin axis, ECM overproduction and
bringing about BSF.
Influence of ethylparaben on the
expression of ECM and agents is associated with the Wnt/β-catenin
axis in primary CFs
In order to reveal the etiology of SCF triggered by
0.01% ethylparaben, the influence of ethylparaben on the expression
of ECM and agents linked with the Wnt/β-catenin axis in cultured
murine primary CFs was explored.
In order to explore the influence of ethylparaben at
low levels on CF survival, and to elucidate the optimal level of
ethylparaben to reveal the etiology of the expression of ECM and
agents linked with the Wnt/β-catenin axis, CFs were supplemented
with media containing either ethylparaben or PBS for 24 h. The
CCK-8 assay was used to evaluate survival. The cut-off
concentration, 0.0001% is presented in Fig. 4A. Ethylparaben above this level was
deleterious to CFs and suppressed CF survival. However,
ethylparaben below this level did no significant damage to CFs.
Consequently, 0.00005% ethylparaben was recognized as eligible for
exploration of the etiology of expression of ECM and agents linked
with the Wnt/β-catenin axis triggered by ethylparaben in CFs. It
was demonstrated that the expression of FN-1, Col1α1, collagen I,
α-SMA, total and p-β-catenin was markedly promoted 24 h following
supplementation of CFs with 0.00005% ethylparaben (Fig. 4B). In addition, ethyparaben
treatment promoted β-catenin translocation (Fig. 4C). These findings verified that
ethylparaben at a low level did no damage to CFs and activated CF
expression of ECM and agents linked with the Wnt/β-catenin
axis.
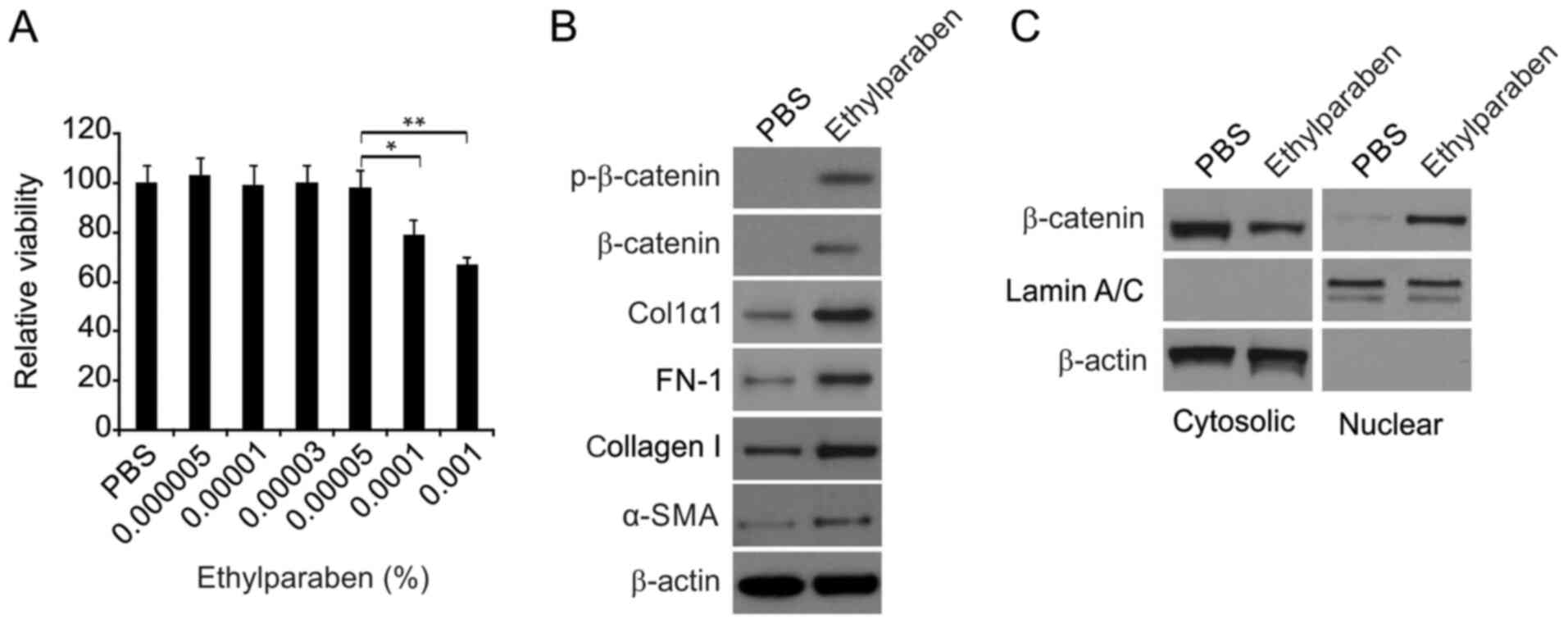 | Figure 4Effects of ethylparaben on primary
CFs. (A) The effect of ethylparaben on CFs was determined via cell
counting kit-8 assay. The data are representative examples of three
independent experiments. *P<0.05 and
**P<0.01. (B) Western blotting assay of expression of
FN-1, Col1α1, collagen I and α-SMA, total and p-β-catenin in the
PBS and ethylparaben-treated groups. (C) Nuclear fractions were
isolated from CFs and analyzed for β-catenin by western blotting.
The expression of FN-1, Col1α1, collagen I, α-SMA, total and
phosphorylated β-catenin was markedly promoted at 24 h following
supplementation of CFs with 0.00005% ethylparaben. CFs,
conjunctival fibroblasts; FN, fibronectin; Col1, collagen type 1;
SMA, smooth muscle actin; p, phosphorylated. |
Ethylparaben at a low level promotes
the CF expression of ECM by stimulating the Wnt/β-catenin axis
To evaluate whether ECM gene expression was
triggered by Wnt/β-catenin stimulation via low levels of
ethylparaben in CFs, CFs were supplemented with ethylparaben,
ethylparaben+XAV-939, or PBS. WB revealed that p-β-catenin
expression was markedly promoted in the experimental group compared
with the control group. However, p-β-catenin concentration was
markedly downregulated in CFs that received ethylparaben+XAV-939
compared with those that received ethylparaben (Fig. 5A). Similarly, FN-1, Col1α1, collagen
I and α-SMA expression were markedly suppressed in CFs that
received ethylparaben+XAV-939 compared with those that received
ethylparaben (Fig. 5B). These
findings suggested that ethylparaben at low levels triggered the
expression of ECM by stimulation of the Wnt/β-catenin axis in
CFs.
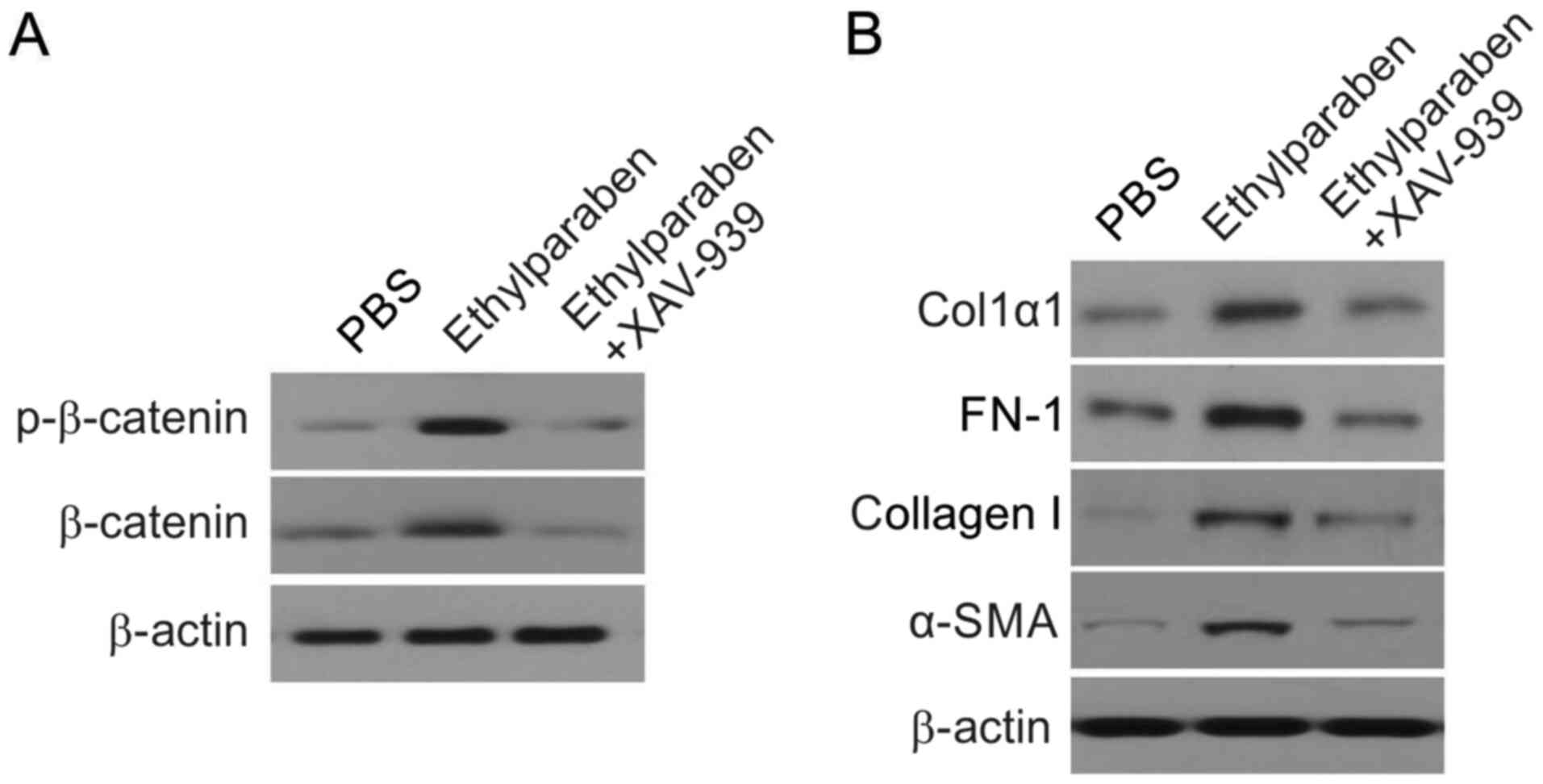 | Figure 5Effect of β-catenin inhibitor XAV-939
on the expression of p-β-catenin, Col1α1, and FN-1. (A) Western
blot assay of expression of p-β-catenin in CFs of ethylparaben and
ethylparaben+XAV-939 (1 µM)-treated groups. p-β-catenin
concentration was markedly suppressed in CFs that received
ethylparaben+XAV-939 (1 µM) compared with those that received
ethylparaben (B) Western blotting assay of expression of FN-1,
Col1α1, collagen I and α-SMA in CFs in ethylparaben and
ethylparaben+XAV-939 (1 µM)-treated groups. FN-1, Col1α1, collagen
I and α-SMA expression was markedly suppressed in CFs that received
ethylparaben+XAV-939 (1 µM) compared with those that received
ethylparaben. P, phosphorylated; CFs, conjunctival fibroblasts; FN,
fibronectin; Col1, collagen type 1; SMA, smooth muscle actin. |
Discussion
In the present study, it was demonstrated for the
first time that ethylparaben-induced SCF was caused by the
Wnt/β-catenin signaling pathway, to the best of our knowledge.
Increased density of fibroblasts and deposition of collagen in
subconjunctival tissue likely accounts for SCF (28,29).
The present study initially revealed that chronic ethylparaben
supplementation mildly promoted fibroblast density and induced
tighter deposition of collagen in murine BST. Chronic ethylparaben
supplementation markedly promoted ECM gene expression and
stimulated the Wnt/β-catenin axis in murine BCT. Low levels of
ethylparaben did no marked harm to cultivated CFs, but promoted ECM
gene expression and triggered the Wnt/β-catenin axis. The ECM gene
expression triggered via ethylparaben was impaired by the
Wnt/β-catenin inhibitor, XAV-939. These findings elucidate the
etiology of SCF triggered by ethylparaben.
Ethylparaben at a low level potentially serves as a
xenobiotic to promote ECM gene expression in CFs by stimulation of
the Wnt/β-catenin axis, which may be an extra side effect of
ethylparaben on the conjunctiva. In investigation of ethylparaben
toxicity, cell cultivation in vitro supplements animal
experiments in vivo, despite the fact that cell monolayer
fails to simulate the more complicated tissue structure. The
cultivated cells are widely used to explore the influence of
ethylparaben on the conjunctiva (30). Several objective standards are
utilized to assess ethylparaben toxicity on cultivated cells such
as survival, condensation of chromatin, ROS generation, and
transmembrane mitochondrial potential (31-33).
In order to determine the nontoxic influence of ethylparaben on CTs
below the cutoff level, primary cultivated CFs were supplemented
with ethylparaben for 24 h. It was demonstrated that ethylparaben
did no harm to CFs, but mildly promoted CF proliferation.
Supplementation of ethylparaben at a low level in the long-term
likely accounts for CFs. Animal experiments were repeated, and the
same results were obtained. Future studies will focus on how
ethylparaben regulates the Wnt/β-catenin signaling pathway, and the
protective effect of Wnt/β-catenin signaling pathway inhibition on
ethylparaben-induced subconjunctival fibrosis.
In summary, the present study revealed that SCF
triggered by ethylparaben results from stimulation of the
Wnt/β-catenin axis in CFs. These findings also suggest that the
development of ethylparaben-free eye drops may be necessary for
patients and will also contribute to the development of novel
therapeutic agents for ethylparaben-related ocular diseases.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and material
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FGL and XFK conceived the study and designed the
experiments, XFK and HK contributed to the data collection,
performed the data analysis and interpreted the results. FGL wrote
the manuscript; XFK contributed to the critical revision of
article. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Committee on
the Ethics of Animal Experiments at Zoucheng People's Hospital
(Zoucheng, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Final amended report on the safety
assessment of Methylparaben, Ethylparaben, Propylparaben,
Isopropylparaben, Butylparaben, Isobutylparaben and Benzylparaben
as used in cosmetic products. Int J Toxicol. 27 (Suppl 4):S1–S82.
2008.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kolatorova L, Vitku J, Hampl R, Adamcova
K, Skodova T, Simkova M, Parizek A, Starka L and Duskova M:
Exposure to bisphenols and parabens during pregnancy and relations
to steroid changes. Environ Res. 163:115–122. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Huang C, Wang H, Pan J, Zhou D, Chen W, Li
W, Chen Y and Liu Z: Benzalkonium chloride induces subconjunctival
fibrosis through the COX-2-modulated activation of a TGF-β1/Smad3
signaling pathway. Invest Ophthalmol Vis Sci. 55:8111–8122.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yalniz-Akkaya Z, Simsek GG, Uney GO, Burcu
A and Ornek F: Effect of cauterization on the subconjunctival
fibrovascular reaction in rabbit eyes. Semin Ophthalmol.
30:202–205. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Paula JS, Ribeiro VR, Chahud F, Cannellini
R, Monteiro TC, Gomes EC, Reinach PS, Rodrigues Mde L and
Silva-Cunha A: Bevacizumab-loaded polyurethane subconjunctival
implants: Effects on experimental glaucoma filtration surgery. J
Ocul Pharmacol Ther. 29:566–573. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhan T, Rindtorff N and Boutros M: Wnt
signaling in cancer. Oncogene. 36:1461–1473. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Krishnamurthy N and Kurzrock R: Targeting
the Wnt/beta-catenin pathway in cancer: Update on effectors and
inhibitors. Cancer Treat Rev. 62:50–60. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhu X, Yuan C, Tian C, Li C, Nie F, Song
X, Zeng R, Wu D, Hao X and Li L: The plant sesquiterpene lactone
parthenolide inhibits Wnt/β-catenin signaling by blocking synthesis
of the transcriptional regulators TCF4/LEF1. J Biol Chem.
293:5335–5344. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Feng XY, Wu XS, Wang JS, Zhang CM and Wang
SL: Homeobox protein MSX-1 inhibits expression of bone
morphogenetic protein 2, bone morphogenetic protein 4, and lymphoid
enhancer-binding factor 1 via Wnt/β-catenin signaling to prevent
differentiation of dental mesenchymal cells during the late bell
stage. Eur J Oral Sci. 126:1–12. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Tacchelly-Benites O, Wang Z, Yang E,
Benchabane H, Tian A, Randall MP and Ahmed Y: Axin phosphorylation
in both Wnt-off and Wnt-on states requires the tumor suppressor
APC. PLoS Genet. 14(e1007178)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Emerick B, Schleiniger G and Boman BM: A
kinetic model to study the regulation of beta-catenin, APC, and
Axin in the human colonic crypt. J Math Biol. 75:1171–1202.
2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Pecina-Slaus N, Kafka A, Vladusic T,
Pecina HI and Hrascan R: AXIN1 expression and localization in
meningiomas and association to changes of APC and E-cadherin.
Anticancer Res. 36:4583–4594. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Niu C, Yin L and Aisa HA: Novel
furocoumarin derivatives stimulate melanogenesis in B16 melanoma
cells by up-regulation of MITF and TYR family via
Akt/GSK3β/β-catenin signaling pathways. Int J Mol Sci.
19(E746)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mbom BC, Siemers KA, Ostrowski MA, Nelson
WJ and Barth AI: Nek2 phosphorylates and stabilizes β-catenin at
mitotic centrosomes downstream of Plk1. Mol Biol Cell. 25:977–991.
2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Dijksterhuis JP, Baljinnyam B, Stanger K,
Sercan HO, Ji Y, Andres O, Rubin JS, Hannoush RN and Schulte G:
Systematic mapping of WNT-FZD protein interactions reveals
functional selectivity by distinct WNT-FZD pairs. J Biol Chem.
290:6789–6798. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Endo Y, Beauchamp E, Woods D, Taylor WG,
Toretsky JA, Uren A and Rubin JS: Wnt-3a and Dickkopf-1 stimulate
neurite outgrowth in ewing tumor cells via a Frizzled3- and c-Jun
N-terminal kinase-dependent mechanism. Mol Cell Biol. 28:2368–2379.
2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lietman C, Wu B, Lechner S, Shinar A,
Sehgal M, Rossomacha E, Datta P, Sharma A, Gandhi R, Kapoor M and
Young PP: Inhibition of Wnt/β-catenin signaling ameliorates
osteoarthritis in a murine model of experimental osteoarthritis.
JCI Insight. 3(96308)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jeon KI, Phipps RP, Sime PJ and Huxlin KR:
Antifibrotic actions of peroxisome proliferator-activated receptor
gamma ligands in corneal fibroblasts are mediated by
β-catenin-regulated pathways. Am J Pathol. 187:1660–1669.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kato N, Shimmura S, Kawakita T, Miyashita
H, Ogawa Y, Yoshida S, Higa K, Okano H and Tsubota K: Beta-catenin
activation and epithelial-mesenchymal transition in the
pathogenesis of pterygium. Invest Ophthalmol Vis Sci. 48:1511–1517.
2007.PubMed/NCBI View Article : Google Scholar
|
|
21
|
ARVO statement on registering clinical
trials. Invest Ophthalmol Vis Sci. 47:1–2. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Tong J, Zheng X, Tan X, Fletcher R,
Nikolovska-Coleska Z, Yu J and Zhang L: Mcl-1 phosphorylation
without degradation mediates sensitivity to HDAC inhibitors by
liberating BH3-only proteins. Cancer Res. 78:4704–4715.
2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tong J, Wang P, Tan S, Chen D,
Nikolovska-Coleska Z, Zou F, Yu J and Zhang L: Mcl-1 degradation is
required for targeted therapeutics to eradicate colon cancer cells.
Cancer Res. 77:2512–2521. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Brown KD, Shah MH, Liu GS, Chan EC,
Crowston JG and Peshavariya HM: Transforming growth factor
β1-induced NADPH oxidase-4 expression and fibrotic response in
conjunctival fibroblasts. Invest Ophthalmol Vis Sci. 58:3011–3017.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen L, Tang RZ, Ruan J, Zhu XB and Yang
Y: Up-regulation of THY1 attenuates interstitial pulmonary fibrosis
and promotes lung fibroblast apoptosis during acute interstitial
pneumonia by blockade of the WNT signaling pathway. Cell Cycle.
4:1–12. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Shao X and Wei X: FOXP1 enhances fibrosis
via activating Wnt/β-catenin signaling pathway in endometriosis. Am
J Transl Res. 10:3610–3618. 2018.PubMed/NCBI
|
|
28
|
Garcia-Posadas L, Soriano-Romani L,
Lopez-Garcia A and Diebold Y: An engineered human conjunctival-like
tissue to study ocular surface inflammatory diseases. PLoS One.
12(e0171099)2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ekinci M, Cagatay HH, Ceylan E, Keles S,
Koban Y, Gokce G, Huseyinoğlu U, Ozcan E and Oba ME: Reduction of
conjunctival fibrosis after trabeculectomy using topical α-lipoic
acid in rabbit eyes. J Glaucoma. 23:372–379. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Takahashi N: Quantitative cytotoxicity of
preservatives evaluated in cell culture with Chang's human
conjunctival cells-effect of temperature on cytotoxicity. Jpn J
Ophthalmol. 26:234–238. 1982.PubMed/NCBI
|
|
31
|
Martins RC, Gmurek M, Rossi AF, Corceiro
V, Costa R, Quinta-Ferreira ME, Ledakowicz S and Quinta-Ferreira
RM: Application of fenton oxidation to reduce the toxicity of mixed
parabens. Water Sci Technol. 74:1867–1875. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Liu T, Li Y, Zhao X, Zhang M and Gu W:
Ethylparaben affects lifespan, fecundity, and the expression levels
of ERR, EcR and YPR in drosophila melanogaster. J Insect Physiol.
71:1–7. 2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Vad NM, Shaik IH, Mehvar R and Moridani
MY: Metabolic bioactivation and toxicity of ethyl 4-hydroxybenzoate
in human SK-MEL-28 melanoma cells. J Pharm Sci. 97:1934–1945.
2008.PubMed/NCBI View Article : Google Scholar
|