Introduction
Gastric mucosal injury is considered a significant
contributor to gastric cancer, with high incidence and recurrence
rates (1,2). Gastric acid acts as a chemical barrier
in the stomach that kills foreign bacteria and protects from
potential infections caused by pathogenic bacteria, which helps
maintain homeostasis in gut microbiota. However, excessive gastric
acid is a major risk factor for gastric mucosal injury (3,4). Since
proton pump inhibitors (PPIs) remarkably inhibit gastric acid
secretion, their wide application is an important and plausible
therapeutic approach for gastric mucosa lesions (5-7).
However, the use of PPIs did not result in the expected fall in
incidence rates of gastric cancer, and gastric cancer remains a
great threat to humans worldwide (8-10).
A study reported that the damaged gastric mucosa did not fully
recover during PPIs treatment and was prone to recurrent gastric
mucosal injury, which could lead to gastric cancer (11). Therefore, new therapies for gastric
mucosal repair need to be urgently identified.
Butyrate is one of the most abundant short-chain
fatty acids (SCFAs), produced by bacterial fermentation of dietary
fibres in the colon, that are essential to maintain an intact
digestive tract mucosa and prevent inflammatory bowel disease (IBD)
development and carcinogenesis (12,13).
Butyrate serves as a critical energy source and is estimated to
provide 70% of the total energy supply to intestinal epithelial
cells. Butyrate combines with GPR109A, GPR43 and GPR41 and
activates signalling pathways (13,14).
Butyrate reportedly attenuated inflammation in murine IBD models,
and its effect could be dismissed in GPR109A−/− mice
(15,16). GPR43−/− (17,18)
and GPR41−/− animals (19,20)
exhibited similar outcomes, showing that mucosal repairs were
counteracted and inflammatory response was not decreased.
Most studies on butyrate were dedicated to its
effects on the intestine, whereas few paid attention to its effects
in the stomach. The response of gastric mucosa to butyrate is
possibly similar to that of the intestine, considering the high
embryonic homologies between the two organs. In previous
experiments, mice that received butyrate administration beforehand
were less likely to be afflicted with severe ethanol-induced acute
gastric mucosal lesions (21).
Nevertheless, whether butyrate has a healing effect on chronic
gastric mucosal lesions and the precise pathways mediating this
effect remain unclear.
In a previous study, gastric mucosal damage with
ulcer formation induced by a local injection of acid was similar to
human chronic gastric ulcer (GU) lesions in terms of morphological,
histological and clinical characteristics (22). This classic chronic GU model
(23) was used in the present study
to determine the therapeutic effects of butyrate on chronic gastric
mucosal injury and the mechanisms thus involved.
Materials and methods
Reagents
Butyrate sodium (cat. no. B5887) was obtained from
Sigma-Aldrich (Merck KGaA). Detection kits for hematoxylin and
eosin (HE; cat. no. D006-1-1) staining, superoxide dismutase (SOD;
cat. no. A001-3-1) and catalase (CAT; cat. no. A007-1-1) were
purchased from Nanjing Jiancheng Bioengineering Institute. Kits for
malondialdehyde (MDA; cat. no. S0131S) and bicinchoninic acid (BCA;
cat. no. P0011) protein assays were purchased from Beyotime
Institute of Biotechnology. Enzyme-linked immunosorbent assay
(ELISA) kits for mouse cytokines including interleukin-1β (IL-1β;
cat. no. F10770), tumour necrosis factor-α (TNF-α; cat. no.
F11630), leukotriene B4 (LTB4; cat. no. F10961) and
6-keto-PGF-1α (cat. no. F11422) were obtained from Westang Biotech
Co., Ltd. Primary antibodies against BAX (cat. no. bs-0127R) and
GPR43 (cat. no. bs-23785R) were purchased from Beijing Biosynthesis
Biotechnology Co., Ltd., and those for GPR41 (cat. no. BS5750) and
GPR109A (cat. no. BS72723) were purchased from Bioworld Technology,
Inc. PrimeScript™ RT reagent kit (cat. no. RR047Q) for reverse
transcription and TB Green® Premix Ex Taq™ (cat. no.
SR4110) were purchased from Takara Bio, Inc.
Animals
Male ICR mice were obtained from the Experimental
Animal Center of Wenzhou Medical University (Wenzhou, China) and
housed in specific pathogen-free animal quarters under a 12-h
light-dark cycle. The mice were given free access to tap water and
food. All animal procedures were performed in accordance with the
guidelines of the Animal Ethics Committee of Wenzhou Medical
University.
GU model and grouping
GU was induced experimentally in mice according to a
method described by Mizuno et al (22), with modifications. The mice were
placed supine on rat plates after overnight fasting and
anesthetized by intraperitoneal injection of 50 mg/kg
pentobarbital. The abdomen was sterilised with alcohol. A median
incision of ~2-3 cm was made along the midline. Diluted
hydrochloric acid (pH=1; 10 µl) was then injected into the
subserosa of the anterior wall at the lesser curvature of the
stomach using a syringe. Later, the stomach was gently placed back
into the abdomen, followed by suturing of the abdominal wall. The
incision was sterilised again with alcohol, and the mice were
placed back in cages after awakening. In control mice, the abdomen
was opened and sutured without any injection.
Laparotomy was performed again 3 days after the
operation, and mice with typical local ulcers were regarded as
successful models. They were randomly divided into the sham, model
and butyrate groups, with 10 mice in each group. The butyrate group
mice were administered with 400 mg/kg butyrate intragastrically for
7 days. No mice was euthanized due to a 20-% weight loss or
debilitating signs, including reduced mobility, ruffled fur,
hunched gait, inactivity or difficulty with eating and drinking.
After 7 days, all mice were euthanized using sodium pentobarbital
(150 mg/kg, intraperitoneal). Gastric tissues were subsequently
obtained.
Histopathological observation of
GU
Gastric tissues were collected, fixed in 10%
buffered formalin at 4˚C for 48 h, dehydrated in an ascending
ethanol followed by two xylene treatments. The samples were
embedded in paraffin and the tissues were cut into 5-µm thick
sections. The sections were subsequently deparaffinized in xylene
and rehydrated with a descending ethanol gradient. The sections
were and stained with haematoxylin for 5 min at 37˚C and 1% eosin
for 3 min at 37˚C. All gastric sections were observed under a light
microscope (magnification, x200).
Immunohistochemical (IHC)
analysis
From gastric tissue blocks, 5-µm sections were
deparaffinised, rehydrated and washed in distilled water three
times. Antigen retrieval was achieved by using high pressure in
citrate buffer (pH=6.0; cat. no. C1010-2L; Beijing Solarbio Science
& Technology Co., Ltd.) at 110˚C for 2 min. They were then
blocked with 5% goat serum (Zhongshan Jinqiao Biotechnology Co.,
Ltd., OriGene Technologies, Inc.) for 30 min at 37˚C. Primary
antibodies against BAX, GPR109A, GPR43 and GPR41 were diluted with
PBST to a dilution of 1:200. Sections were covered with primary
antibodies at 4˚C overnight. The bound antibody was developed with
diaminobenzidine using a Dako REAL Envision staining kit (cat. no.
K5007; Agilent Technologies, Inc.) according to the manufacturer's
instructions. The sections were soaked with haematoxylin for 5 min
at room temperature, hydrochloric alcohol for 5 sec, PBS (pH=7.4)
for 10 sec. Stained sections were examined under a light microscope
(magnification, x200) by ZY and JXW.
Determination of MDA levels, and SOD
and CAT activities
MDA levels and total SOD and CAT activities in
gastric tissues were determined using thiobarbituric acid, xanthine
oxidase and ammonium molybdate, respectively. Absorbance was
detected using a 722N spectrophotometer (Scientific Instrument Co.,
Ltd). Procedures were performed according to the instructions
provided for each kit.
ELISA for cytokines
IL-1β, TNF-α, LTB4 and 6-keto-PGF-1α
levels in GU tissues were determined using commercially available
ELISA kits as per the corresponding manufacturer's protocol.
Briefly, GU tissue homogenates were pipetted into 96-well plates
coated with the primary antibodies and incubated for 3 h at 37˚C.
Biotin conjugate was then added for incubation for 45 min at room
temperature after washing. Streptavidin-horseradish peroxidase was
incubated for 45 min at 37˚C after washing nonspecific binding.
Then, a chromogen was pipetted into each well for absorbance at 450
nm. IL-1β, TNF-α, LTB4 and 6-keto-PGF-1α levels were
calculated using standard curves according to the ELISA kit
instructions.
Reverse transcription-quantitative
polymerase PCR analysis
Total RNA was isolated from cultured cells using
TRIzon reagent (cat. no. CW0580S; Beijing ComWin Biotech Co., Ltd.)
as per the manufacturer's protocols. Reverse transcription was
performed using the PrimeScript™ RT reagent kit. The thermocycling
condition was as follows: 37˚C for 15 min; followed by 85˚C for 5
sec and then 4˚C for storage. qPCRs were prepared using SYBR Green
on a Prism 7500 Sequence Detector (Thermo Fisher Scientific, Inc.).
The thermocycling condition was as follows: Initial denaturation at
95˚C for 10 min; followed by 40 cycles of 95˚C for 15 sec, 60˚C for
45 sec and then 95˚C for 15 sec. mRNA expression levels of BAX,
Caspase-3, trefoil factors (TFFs) 1-3, MUC5AC, fibroblast growth
factor-7 (FGF7), GPR109A, GPR43 and GPR41 were normalised to levels
of GAPDH. The 2-∆∆Cq method was used for relative
quantification (24). Sequences for
qPCR primers for all targeted genes are listed in Table I.
 | Table ISpecific primers used for
amplification of targeted genes. |
Table I
Specific primers used for
amplification of targeted genes.
| Gene name | Forward primer | Reverse primer |
|---|
| BAX |
5'-ACCAAGAAGCTGAGCGAGTG-3' |
5'-CCCAGTTGAAGTTGCCATCA-3' |
| Caspase 3 |
5'-ATGGGAGCAAGTCAGTGGAC-3' |
5'-GTCCACATCCGTACCAGAGC-3' |
| TFF1 |
5'-AAGGTGATCTGTGTCCTCGC-3' |
5'-AAACAGCAACCTCTCTCCGT-3' |
| TFF2 |
5'-CGGAGCAGTGTGTCATGGAA-3' |
5'-AAGAAACACCAGGGCACTTCA-3' |
| TFF3 |
5'-GCCCTCTGGCTAATGCTGTT-3' |
5'-CGGTTGTTACACTGCTCCGA-3' |
| MUC5AC |
5'-GTTCACTCTACCACTCCCTGC-3' |
5'-CAATCCTGGCTACACATCGC-3' |
| FGF7 |
5'-CGTGGCAGTTGGAATTGTGG-3' |
5'-AGGCAACGAACATTTCCCCT-3' |
| GPR109A |
5'-TACCACCCTTAGCTTTACCT-3' |
5'-CCTGGAATACTTCTGGTTGT-3' |
| GPR43 |
5'-GGTGTGCTTTGGACCCTACA-3' |
5'-CTGTCTCTTTGGCTCCCCTG-3' |
| GPR41 |
5'-TGAGCATCGAACGTTTTCTG-3' |
5'-CCAGGTAGCAGGTTCCATTG-3' |
| GAPDH |
5'-AGGTCGGTGTGAACGGATTTG-3' |
5'-GGGGTCGTTGATGGCAACA-3' |
Statistical analysis
All experimental data are expressed as mean ± SD.
Mean differences were compared with one-way ANOVA followed by
multiple comparison by Bonferroni test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Butyrate alleviates pathological
damage to gastric mucosa in mice
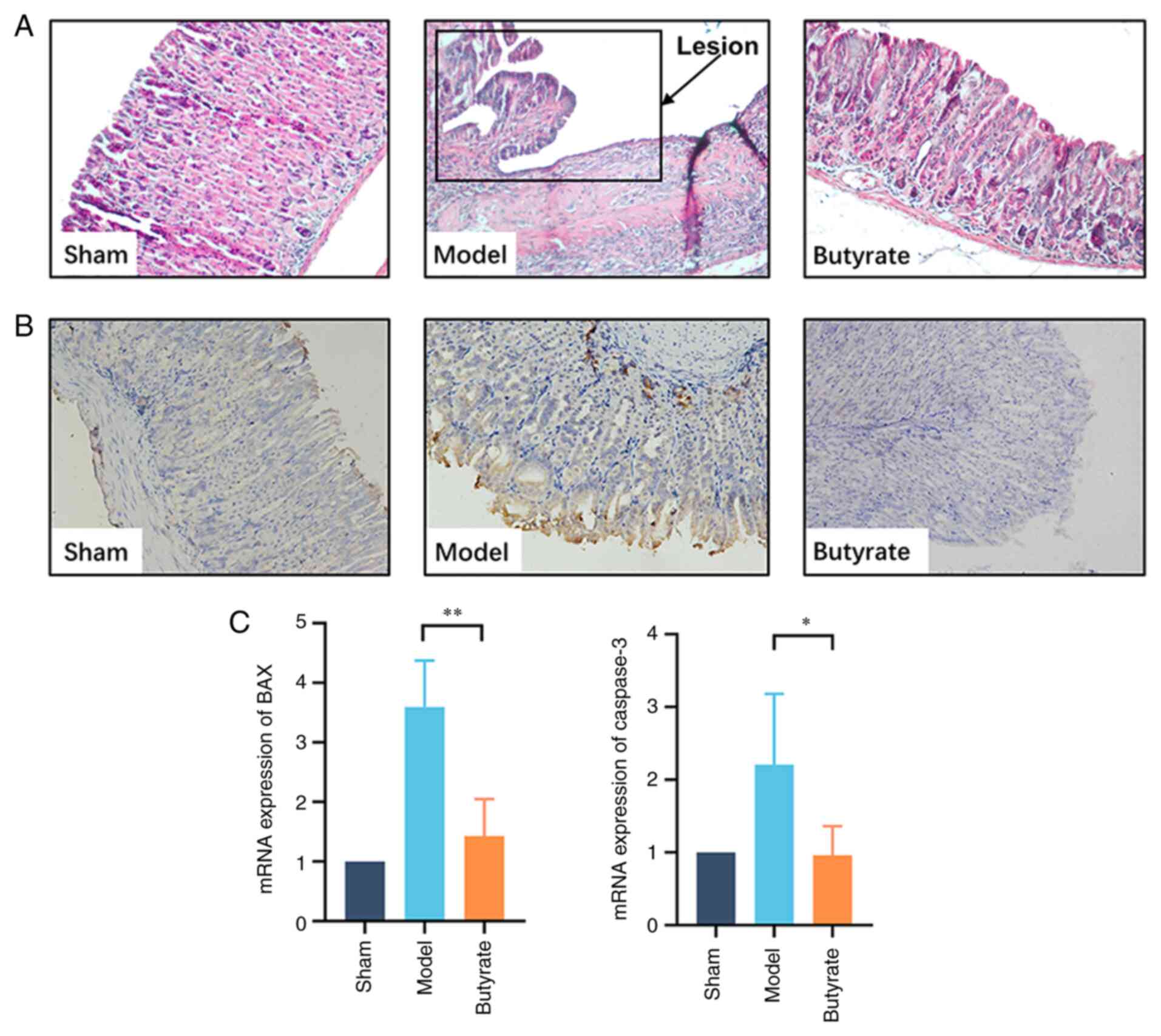
Conventional HE staining was used to examine the
pathological changes in different groups. Histological examination
in the sham group demonstrated that the epithelial lining was
intact and glandular cavities were clear; furthermore, there was no
inflammatory cell infiltration in the gastric mucosa. Necrotic
tissue was observed on the mucosal ulcer surface in the model
group. There was granulation hyperplasia with inflammatory cell
infiltration; furthermore, hyperaemia, oedema and
neovascularisation were observed surrounding the ulcer. Butyrate
treatment significantly attenuated the gastric mucosal damage
induced by acid injection. The gastric mucosal structure appeared
intact with regular hyperplastic glands, and inflammatory cells
were occasionally found under the mucosa (Fig. 1A).
IHC results suggested that BAX was strongly and
positively expressed in the GU group compared with that in control
animals. Butyrate treatment significantly counteracted such changes
(Fig. 1B). qPCR results suggested
that butyrate treatment decreased BAX and caspase-3 expression
levels (Fig. 1C), and this finding
was consistent with the IHC analysis results.
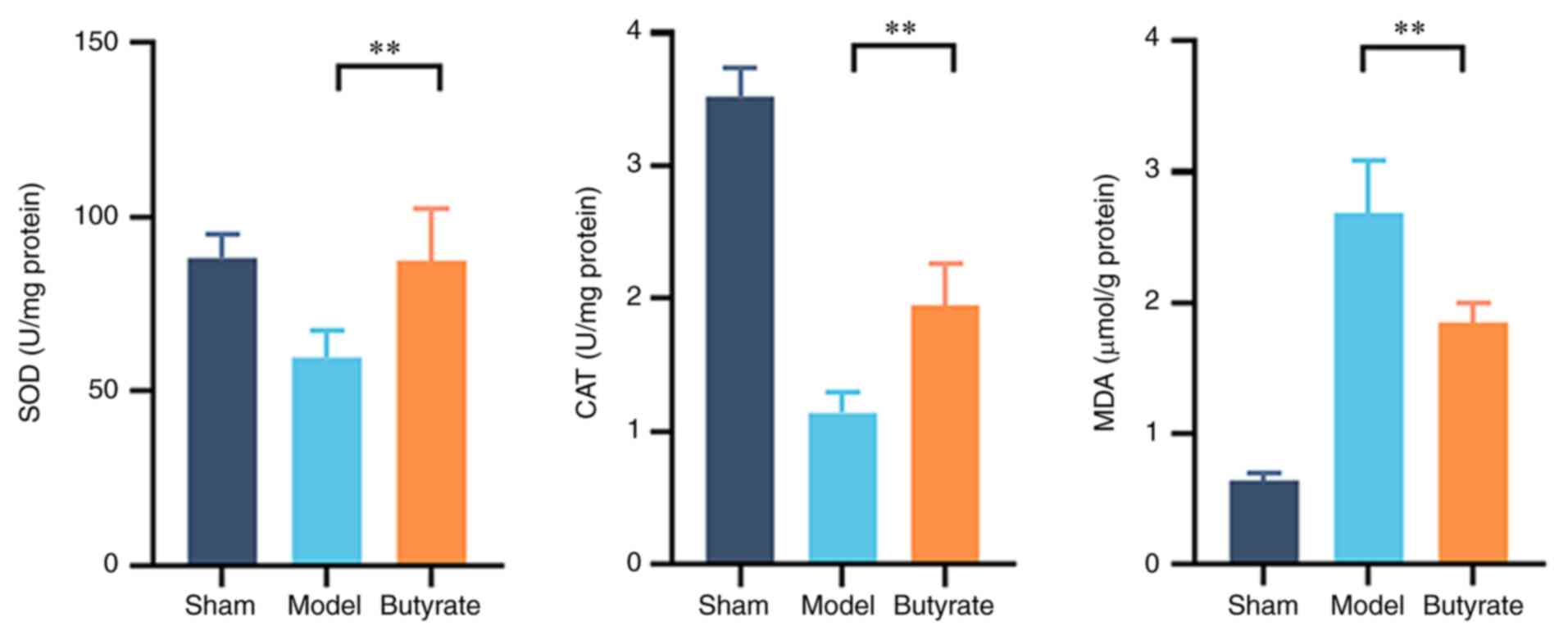
Butyrate ameliorated oxidative stress
in GU
It is well established that oxidative stress plays
an important role in GU formation. SOD and CAT activities, as well
as MDA levels in GU tissues, were used to measure oxidative stress
levels in the various groups (Fig.
2). SOD and CAT activities were significantly lower and MDA
levels were significantly higher (P<0.01) in the model group
compared with other groups. Butyrate treatment significantly
attenuated these changes (Fig. 2;
P<0.01), suggesting the mitigating effect of butyrate on
oxidative stress in GU.
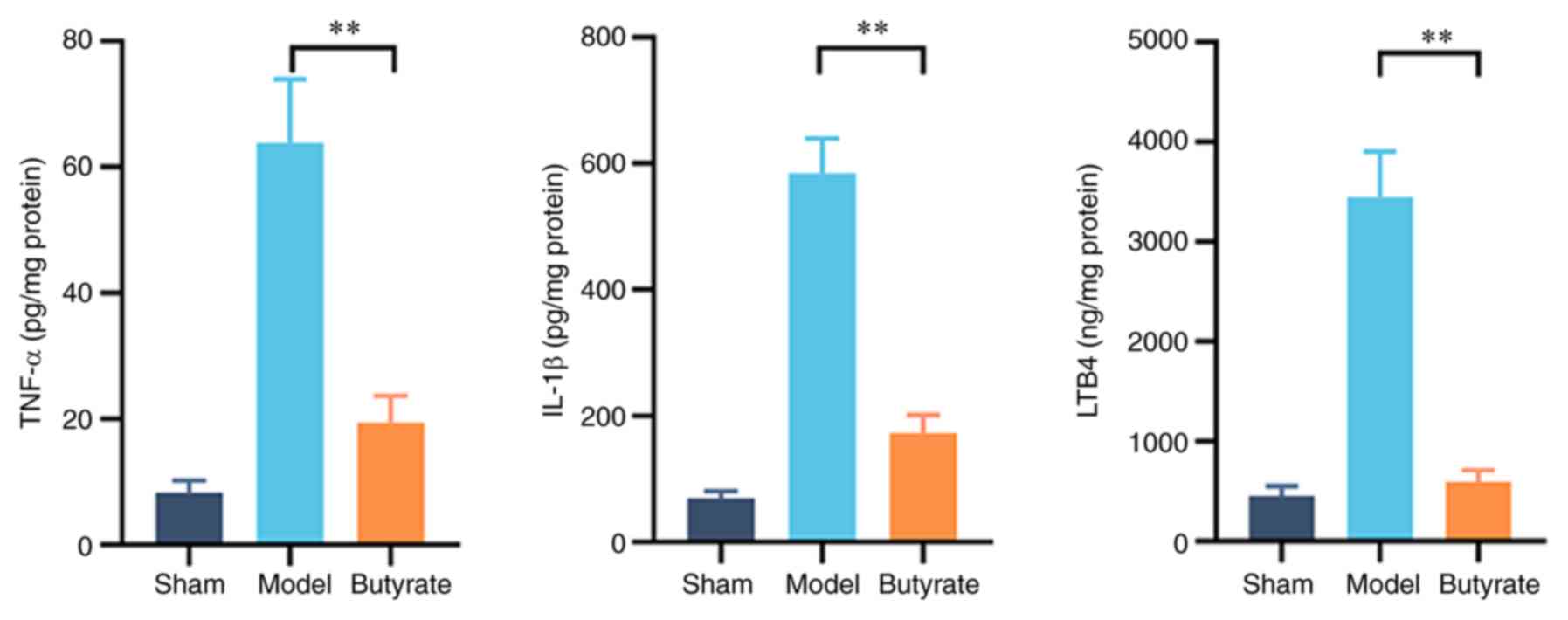
Butyrate attenuates inflammation in
GU
Cytokine levels were measured to investigate the
underlying mechanisms of the therapeutic effects of butyrate on
gastric mucosal lesions because of the involvement of excessive
inflammatory responses in GU pathogenesis. Changes in TNF-α, IL-1β
and LTB4 levels in gastric tissues of the various groups
are shown in Fig. 3; the levels of
these cytokines were significantly higher in the model group
compared with the other groups. Butyrate treatment decreased the
elevation in the levels of these pro-inflammatory cytokines
(P<0.01; Fig. 3). This suggests
that butyrate attenuates the excessive inflammation involved in the
development of gastric mucosal ulcers induced by erosion due to
acid in mice.
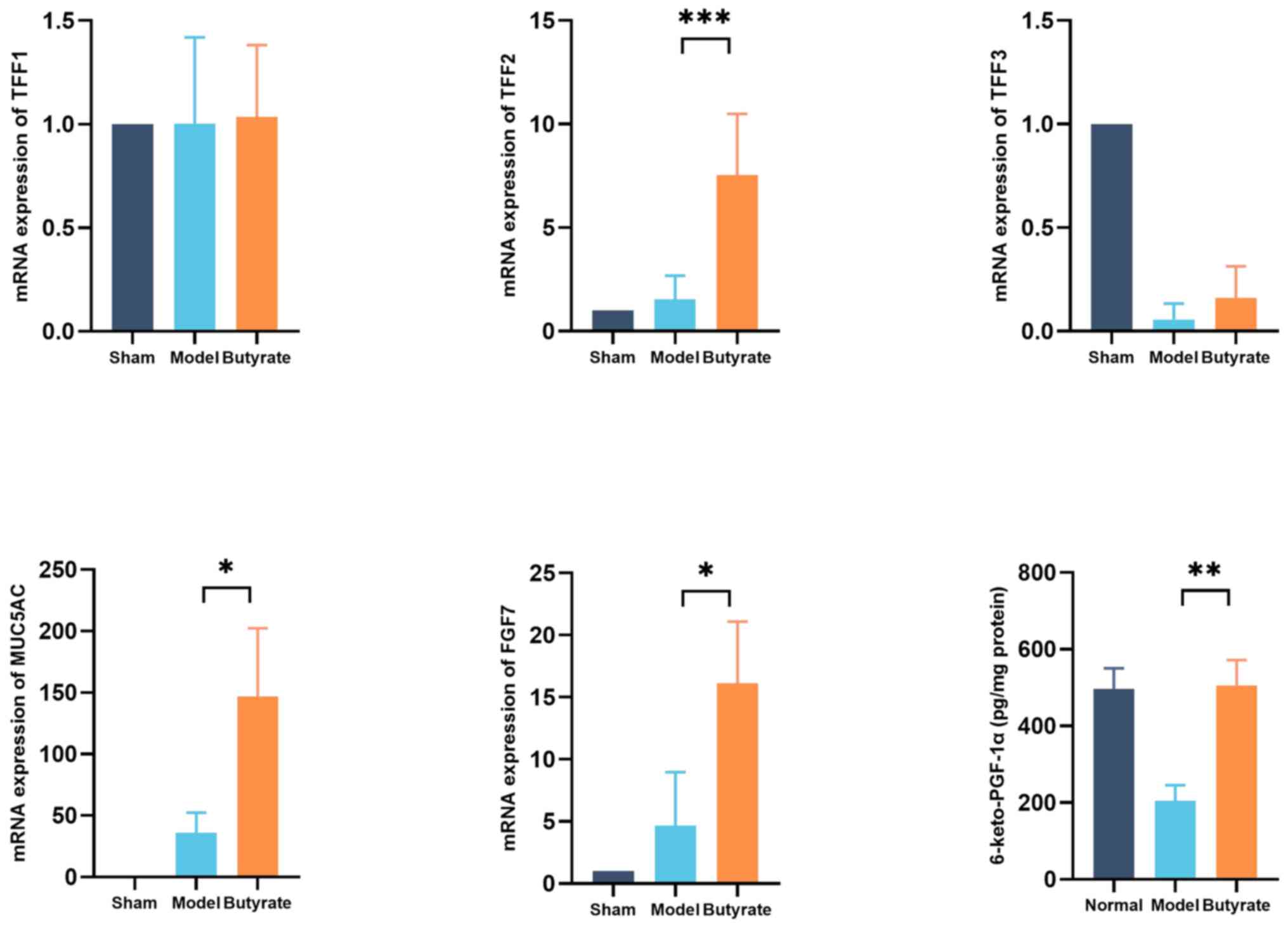
Butyrate promotes gastric mucosal
repair
TFFs1-3, MUC5AC and FGF7 are crucial for gastric
mucosal repair. qPCR was performed to determine the changes in
expression levels of these proteins in the various groups. Butyrate
significantly increased the expression levels of genes encoding
MUC5AC, FGF7 and TFF2 proteins (P<0.05), but not of those
encoding TFF1 or TFF3 proteins, in gastric tissues (Fig. 4). Prostacyclin (PGI2) is important
for maintaining gastric mucosal defences. The levels of
6-keto-PGF-1α (a PGI2 metabolite) were measured, which reflect
gastric mucosal repair; were significantly higher in
butyrate-treated mice compared with those of the model mice
(P<0.01; Fig. 4).
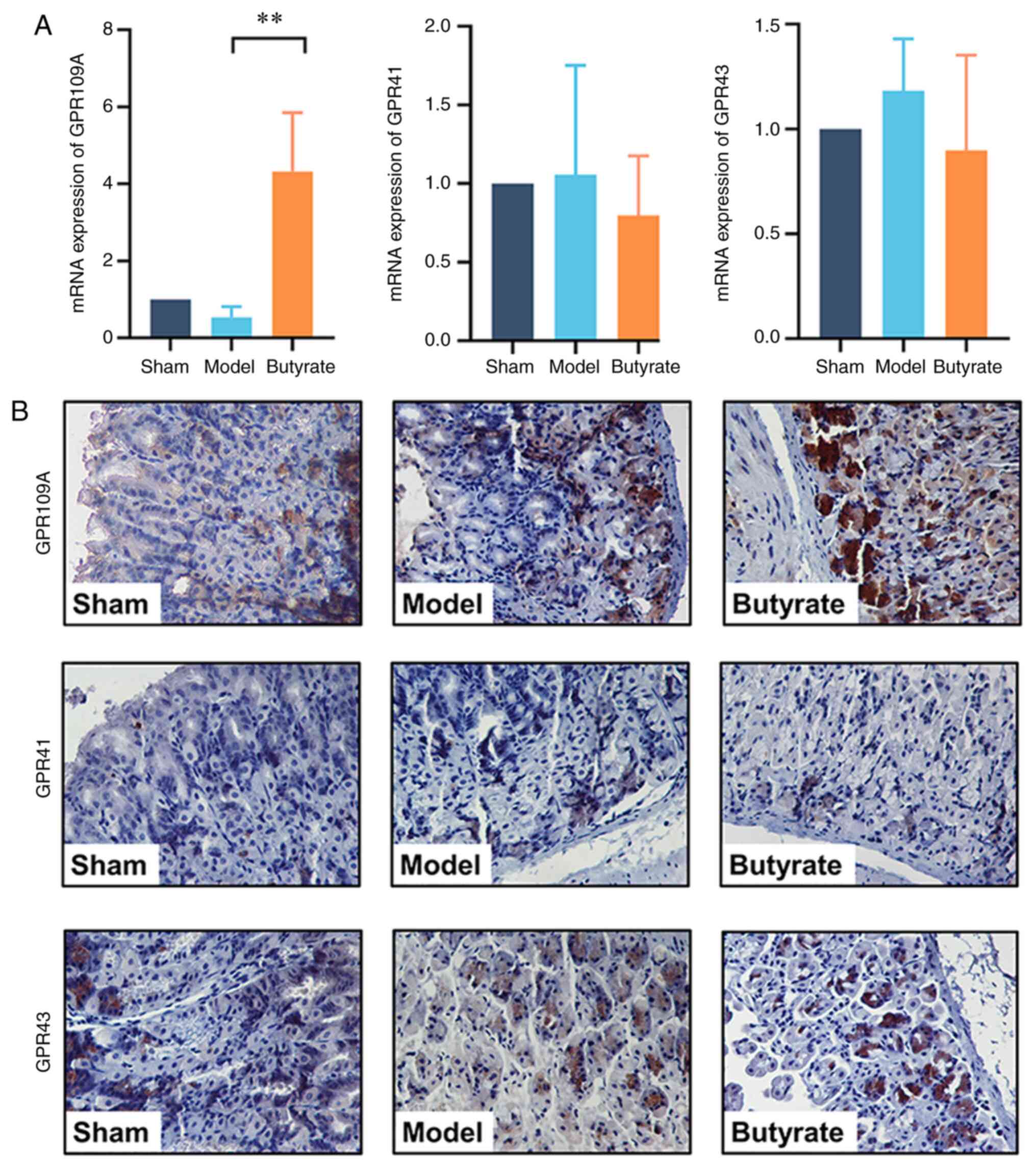
Butyrate upregulates GPR109A
expression
Given that SCFA receptors, including GPR109A, GPR43
and GPR41, are important for the biological function of SCFAs, the
expression levels of these receptors were tested to determine the
signalling mechanisms underlying the therapeutic effects of
butyrate. qPCR results showed that the expression of GPR109A, but
not of GPR43 or GPR41, was upregulated by butyrate treatment
(P<0.01; Fig. 5A). Consistent
with these findings, IHC analysis showed that GPR109A was strongly
positively expressed in the butyrate group compared with that in
the GU mice. However, butyrate treatment did not change GPR43 or
GPR41 expression levels (Fig.
5B).
Discussion
Nonsteroidal anti-inflammatory drugs and
Helicobacter pylori infection are the leading factors that
impair gastric mucosa defences and give rise to GUs; however, the
acid corrosive effect on gastric mucosa is the basic factor
contributing to GU pathogenesis (1). For this reason, the chronic GU model
is traditionally established using a local injection of 20% acetic
acid under the mucosal lumen to imitate acid corrosion-induced GU
(22). Due to the protective effect
of SCFA acetate on gastric mucosa observed in a previous study
(25), diluted hydrochloric acid
(pH=1), the main component of gastric acid, was used instead of
acetic acid, to build the GU model in the present study. The
diameter of a GU usually ranges from 0.4 to 0.8 cm, depending
directly on the concentration and dosage of the acid used in this
model. Based on a preliminary experiment, 10 µl hydrochloric acid
(pH=1) was chosen as suitable for the chronic GU establishment.
It was difficult to maintain an accurate depth of
acid injection for GU formation during the animal operation.
Therefore, the abdomen was opened to select the mice with GU 3 days
after the acid injection for the experiment. In the model group,
pathological examination showed that acid injection induced a
series of typical changes that occur during ulcer development,
including local mucosal necrosis and submucosal granulation tissue
formation, similar to human chronic ulcer. This suggests that the
GU model was successfully constructed.
Since excessive gastric acid secretion contributes
to mucosal injury, its proper inhibition lays a foundation on
current GU treatment. However, despite the extensive PPIs usage
(26,27), the recurrence of GU remains high.
Therefore, experimental agents with mucosal repairing ability were
investigated. Butyrate is an important metabolite from indigestive
fibers fermented by gut microbiota. When orally administrated,
butyrate is mainly absorbed by digestive mucosa and thereby
elevates the expression of tight junction proteins MUC3, MUC5B,
ZO-1 and Occludin to protect the mucosa (28-31).
It is noteworthy that butyrate prompts the repair and recovery in
intestinal mucosa (13,21,30),
while its effect on gastric acid secretion has not been reported
yet. Hence, corresponding experiments were conducted to uncover
that butyrate pretreatment protected gastric mucosa from
ethanol-induced lesions. Therefore, it is postulated that butyrate
prompts recovery in gastric mucosa similar to that in intestine on
a distinct basis of its remarkable ability of mucosal repair,
rather than that of acid secretion inhibition.
Butyrate is a preferable energy source for human
intestinal epithelial cells, and has been widely used as a food
additive (32). Egorin et al
(33) reported the maximum
tolerated dose of sodium butyrate in mice as 1.25 g/kg in a study
of sodium butyrate plasma pharmacokinetics, which was over triple
times higher than 400 mg/kg used in the present study. Liang et
al (34) found that 400 mg/kg
sodium butyrate, equivalent to 640 mg/kg for mice, could defend
severe burn-induced remote acute lung injury in rats. Therefore,
the dose of butyrate used in the present study's experimental mice
(400 mg/kg), equal to 44 mg/kg for human, was of easy access and
less severe dose-dependent side effects.
In the present study, it was found that oral
butyrate significantly alleviated the pathological damage to
gastric mucosa in mice. The butyrate group exhibited less necrotic
tissues and more proliferative glands with regular arrangement. The
physical condition of the mice improved following treatment with
butyrate. The mice showed inactivity, food intake reduction,
fluffy-coat formation and decreased body weight and passed
shapeless faeces after modelling; however, these symptoms were
significantly alleviated following butyrate treatment. Mice
feeding, activity levels and stool appearance were normal, and
their weight also increased on the seventh day. Given that butyrate
is produced by probiotic microbiota, most of the previous studies
focused mainly on intestinal diseases (35,36).
The present study findings suggest that butyrate heals chronic
gastric mucosal lesions.
Oxidative stress is one of the most important
factors contributing to the pathological development of GU. The
anti-oxidants SOD and CAT attenuate oxidative stress. In the
studies on GU, it was found that SOD and CAT activities were
significantly decreased after modelling (25,37).
Butyrate was shown to increase the anti-oxidative capacity in rats
(38) and HepG2 cells (39). The present data suggest that the
levels of oxidative product MDA in gastric tissues were increased
and activities of SOD and CAT were decreased in the model group,
whereas butyrate treatment attenuated the changes in these
parameters, thereby protecting against oxidative damage.
Excessive inflammation is a crucial factor in GU
formation. Levels of pro-inflammatory factors IL-1β, TNF-α and
LTB4 were elevated in the model mice, whereas this
elevation in levels was substantially decreased by butyrate
treatment. The anti-inflammatory effect of butyrate has been
extensively studied in various fields (16,17,36).
Some studies reported that butyrate inhibited the nuclear factor κB
signal pathway to prevent the intestinal epithelial injury caused
by inflammation (40-42).
SCFA-specific receptors GPR43, GPR41 and particularly GPR109A,
mediate the anti-inflammatory effects of SCFAs (15,18,20).
The present data showed that the expression of GPR109A, but not of
GPR43 or GPR41, was significantly upregulated by butyrate
treatment, suggesting GPR109A to be the key receptor of butyrate
for mediating its gastric protective effect. Nevertheless, the
underlying mechanisms remain to be investigated in future
studies.
In the present study, TFFs1-3, MUC5AC and FGF7
levels were determined, which reflect gastric mucosal repair.
Previously, TFFs1-3 (43-45)
and FGF7(46) were shown to promote
gastric mucosal repair. It was found that butyrate treatment
significantly upregulated TFF2, MUC5AC and FGF7 expression levels.
Butyrate is necessary for the nutrition of the intestinal
epithelium to maintain mucosal integrity (13). In the present study, the therapeutic
effect of butyrate on GU was investigated. Combined with the
analysis for the special receptors of SCFAs, the data suggest that
butyrate regulates gene expression via the GPR109A signalling
pathway. Nevertheless, it has been well documented that butyrate
acts as a histone deacetylase inhibitor (HDACi) to regulate gene
expression by increasing acetylation modification of histones
(47-49).
It is noteworthy that because of the Warburg effect, butyrate
concentration increased so that it acts as an HDACi in cancerous
cells, but it entered the tricarboxylic acid cycle at physiological
levels to act as a substrate for energy metabolism (50). In future experiments, the detailed
mechanism underlying the therapeutic effects of butyrate will be
validated.
It was reported that GPR109A mediates cyclooxygenase
(COX) activation to increase prostanoid formation (51,52).
COXs are important for maintaining the gastric mucosal barrier
(53). The present results suggest
that butyrate promotes PGI2 production, reflected by 6-keto-PGF-1α
levels, and significantly decreased LTB4 levels. Thus,
butyrate may exert its therapeutic effects on GUs via arachidonic
acid metabolic pathway regulation.
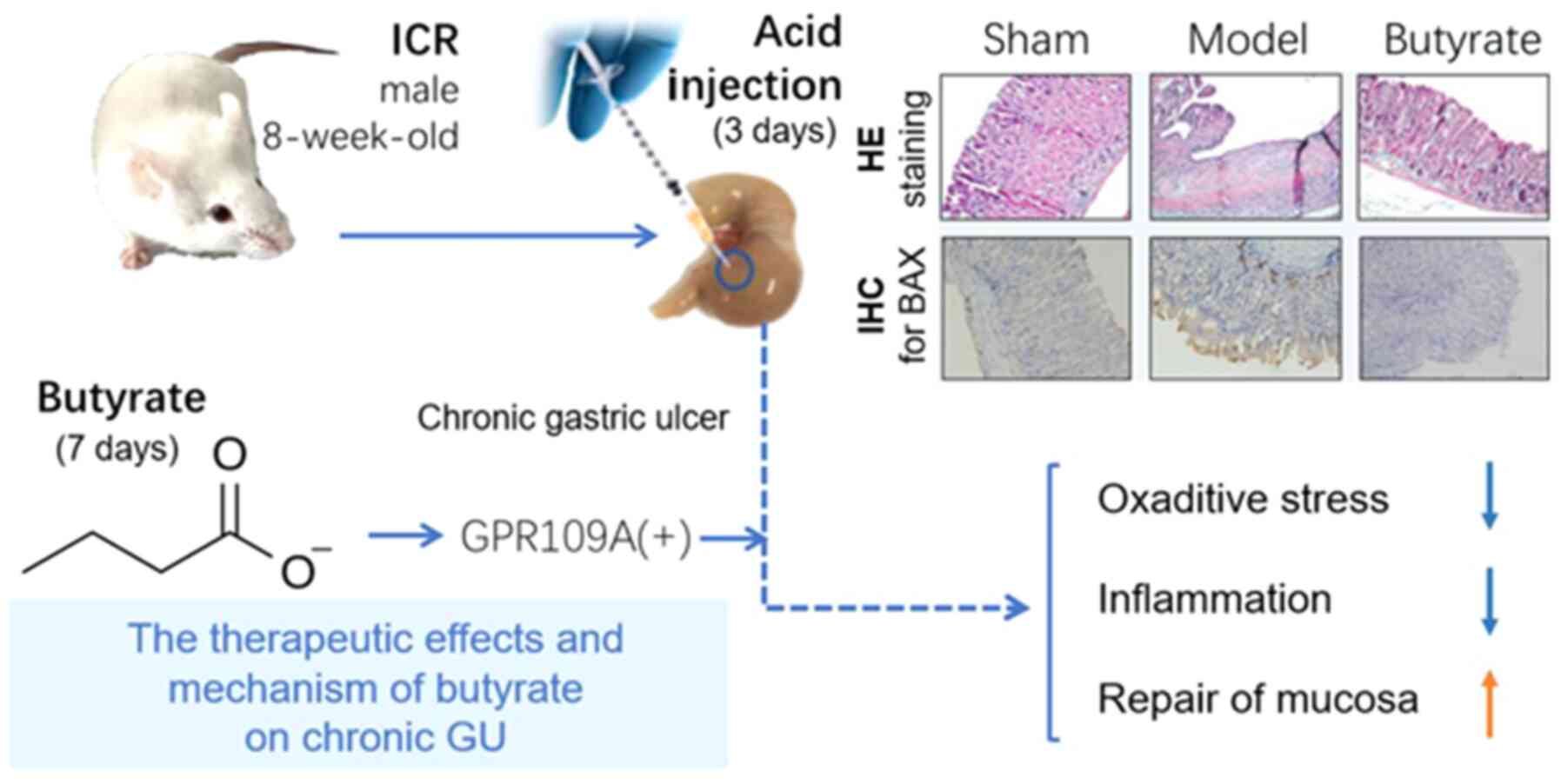
In conclusion, the therapeutic effect of butyrate on
chronic GUs was identified via anti-oxidation, anti-inflammation
and promotion of gastric mucosal repair; the effect may be mediated
by the GPR109A receptor. The hypothesis regarding the therapeutic
mechanism of butyrate is presented in Fig. 6.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by grants from the
Science and Technology Department of Zhejiang (grant nos.
2017C33068, LY20H180010 and LGF20H070003), the Medical and Health
Research Project Grant of Zhejiang Province of China (grant no.
Y2019317606) and the Technology Bureau of Wenzhou (grant no.
Y20180142 and Y20190060). The funders had no role in the study
design, data collection and analysis, decision to publish, or
preparation of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available on reasonable request from the corresponding
author.
Authors' contributions
FYW and CLX conceived and designed the study. YZ,
XWJ, JJC, JKC, YYF, JWH, RG, JHZ, QHZ and AC performed the
experiments. YZ and XWJ authenticated the raw data in this study.
YPH participated in drafting the manuscript together with YZ and
XWJ, and performed data analysis with YZ, XWJ and JKC. YZ and XWJ
wrote up the manuscript. FYW, CLX and YPH revised the critically
manuscript for important intellectual content and finally approved
the version to be published. All authors have read and agreed to
the published version of the manuscript.
Ethics approval and consent to
participate
Experimental procedures and the Animal Use and Care
protocols were approved by the Ethics Committee of Wenzhou Medical
University (approval no. wydw2012-0109; Wenzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Graham DY: History of Helicobacter pylori,
duodenal ulcer, gastric ulcer and gastric cancer. World J
Gastroenterol. 20:5191–5204. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Waldum HL, Kleveland PM and Sordal OF:
Helicobacter pylori and gastric acid: An intimate and reciprocal
relationship. Therap Adv Gastroenterol. 9:836–844. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Adeniyi OS, Emikpe BO and Olaleye SB:
Accelerated gastric ulcer healing in thyroxine-treated rats: Roles
of gastric acid, mucus, and inflammatory response. Can J Physiol
Pharmacol. 96:597–602. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Brzozowski T, Magierowska K, Magierowski
M, Ptak-Belowska A, Pajdo R, Kwiecien S, Olszanecki R and Korbut R:
Recent advances in the gastric mucosal protection against
stress-induced gastric lesions. Importance of renin-angiotensin
vasoactive metabolites, gaseous mediators and appetite peptides.
Curr Pharm Des. 23:3910–3922. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yu LY, Sun LN, Zhang XH, Li YQ, Yu L, Yuan
ZQ, Meng L, Zhang HW and Wang YQ: A review of the novel application
and potential adverse effects of proton pump inhibitors. Adv Ther.
34:1070–1086. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yang X, Li Y, Sun Y, Zhang M, Guo C, Mirza
IA and Li YQ: Vonoprazan: A novel and potent alternative in the
treatment of acid-related diseases. Dig Dis Sci. 63:302–311.
2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yamamoto O, Okada Y and Okabe S: Effects
of a proton pump inhibitor, omeprazole, on gastric secretion and
gastric and duodenal ulcers or erosions in rats. Dig Dis Sci.
29:394–401. 1984.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yoon H and Kim N: Diagnosis and management
of high risk group for gastric cancer. Gut Liver. 9:5–17.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Ang TL and Fock KM: Clinical epidemiology
of gastric cancer. Singapore Med J. 55:621–628. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yan S, Li B, Bai ZZ, Wu JQ, Xie DW, Ma YC,
Ma XX, Zhao JH and Guo XJ: Clinical epidemiology of gastric cancer
in hehuang valley of China: A 10-year epidemiological study of
gastric cancer. World J Gastroenterol. 20:10486–10494.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hansson LE, Nyren O, Hsing AW, Bergström
R, Josefsson S, Chow WH, Fraumeni JF Jr and Adami HO: The risk of
stomach cancer in patients with gastric or duodenal ulcer disease.
N Engl J Med. 335:242–249. 1996.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tedelind S, Westberg F, Kjerrulf M and
Vidal A: Anti-inflammatory properties of the short-chain fatty
acids acetate and propionate: A study with relevance to
inflammatory bowel disease. World J Gastroenterol. 13:2826–2832.
2007.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ploger S, Stumpff F, Penner GB, Schulzke
JD, Gäbel G, Martens H, Shen Z, Günzel D and Aschenbach JR:
Microbial butyrate and its role for barrier function in the
gastrointestinal tract. Ann N Y Acad Sci. 1258:52–59.
2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kimura I, Ichimura A, Ohue-Kitano R and
Igarashi M: Free fatty acid receptors in health and disease.
Physiol Rev. 100:171–210. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Singh N, Gurav A, Sivaprakasam S, Brady E,
Padia R, Shi H, Thangaraju M, Prasad PD, Manicassamy S, Munn DH, et
al: Activation of Gpr109a, receptor for niacin and the commensal
metabolite butyrate, suppresses colonic inflammation and
carcinogenesis. Immunity. 40:128–139. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chen G, Ran X, Li B, Li Y, He D, Huang B,
Fu S, Liu J and Wang W: Sodium butyrate inhibits inflammation and
maintains epithelium barrier integrity in a TNBS-induced
inflammatory bowel disease mice model. EBioMedicine. 30:317–325.
2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Pirozzi C, Francisco V, Guida FD, Gómez R,
Lago F, Pino J, Meli R and Gualillo O: Butyrate modulates
inflammation in chondrocytes via GPR43 receptor. Cell Physiol
Biochem. 51:228–243. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Nakajima A, Nakatani A, Hasegawa S, Irie
J, Ozawa K, Tsujimoto G, Suganami T, Itoh H and Kimura I: The short
chain fatty acid receptor GPR43 regulates inflammatory signals in
adipose tissue M2-type macrophages. PLoS One.
12(e0179696)2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lin HV, Frassetto A, Kowalik EJ Jr,
Nawrocki AR, Lu MM, Kosinski JR, Hubert JA, Szeto D, Yao X, Forrest
G and Marsh DJ: Butyrate and propionate protect against
diet-induced obesity and regulate gut hormones via free fatty acid
receptor 3-independent mechanisms. PLoS One.
7(e35240)2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kim MH, Kang SG, Park JH, Yanagisawa M and
Kim CH: Short-chain fatty acids activate GPR41 and GPR43 on
intestinal epithelial cells to promote inflammatory responses in
mice. Gastroenterology. 145:396–406, e391-310. 2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Liu J, Wang F, Luo H, Liu A, Li K, Li C
and Jiang Y: Protective effect of butyrate against ethanol-induced
gastric ulcers in mice by promoting the anti-inflammatory,
anti-oxidant and mucosal defense mechanisms. Int Immunopharmacol.
30:179–187. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Mizuno H, Sakamoto C, Matsuda K, Wada K,
Uchida T, Noguchi H, Akamatsu T and Kasuga M: Induction of
cyclooxygenase 2 in gastric mucosal lesions and its inhibition by
the specific antagonist delays healing in mice. Gastroenterology.
112:387–397. 1997.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Toma W, Hiruma-Lima CA, Guerrero RO and
Brito AR: Preliminary studies of mammea americana L. (Guttiferae)
bark/latex extract point to an effective antiulcer effect on
gastric ulcer models in mice. Phytomedicine. 12:345–350.
2005.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Liu J, Wang J, Shi Y, Su W, Chen J, Zhang
Z, Wang G and Wang F: Short chain fatty acid acetate protects
against ethanol-induced acute gastric mucosal lesion in mice. Biol
Pharm Bull. 40:1439–1446. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kangwan N, Park JM, Kim EH and Hahm KB:
Quality of healing of gastric ulcers: Natural products beyond acid
suppression. World J Gastrointest Pathophysiol. 5:40–47.
2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tarnawski A, Douglass TG, Stachura J and
Krause WJ: Quality of gastric ulcer healing: Histological and
ultrastructural assessment. Aliment Pharmacol Ther. 1 (5
Suppl):S79–S90. 1991.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Guilloteau P, Martin L, Eeckhaut V,
Ducatelle R, Zabielski R and Van Immerseel F: From the gut to the
peripheral tissues: The multiple effects of butyrate. Nutr Res Rev.
23:366–384. 2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Bloemen JG, Venema K, van de Poll MC, Olde
Damink SW, Buurman WA and Dejong CH: Short chain fatty acids
exchange across the gut and liver in humans measured at surgery.
Clin Nutr. 28:657–661. 2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Gaudier E, Jarry A, Blottiere HM, de
Coppet P, Buisine MP, Aubert JP, Laboisse C, Cherbut C and Hoebler
C: Butyrate specifically modulates MUC gene expression in
intestinal epithelial goblet cells deprived of glucose. Am J
Physiol Gastrointest Liver Physiol. 287:G1168–G1174.
2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liang JB, Wang P, Feng YH, Huang YL, Wang
FJ and Ren H: Effects of sodium butyrate on intestinal barrier of
severe scald mice and the related mechanism. Zhonghua shao shang za
zhi. 36:48–53. 2020.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
32
|
Barcenilla A, Pryde SE, Martin JC, Duncan
SH, Stewart CS, Henderson C and Flint HJ: Phylogenetic
relationships of butyrate-producing bacteria from the human gut.
Appl Environ Microbiol. 66:1654–1661. 2000.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Egorin MJ, Yuan ZM, Sentz DL, Plaisance K
and Eiseman JL: Plasma pharmacokinetics of butyrate after
intravenous administration of sodium butyrate or oral
administration of tributyrin or sodium butyrate to mice and rats.
Cancer Chemother Pharmacol. 43:445–453. 1999.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liang X, Wang RS, Wang F, Liu S, Guo F,
Sun L, Wang YJ, Sun YX and Chen XL: Sodium butyrate protects
against severe burn-induced remote acute lung injury in rats. PLoS
One. 8(e68786)2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wong JM, de Souza R, Kendall CW, Emam A
and Jenkins DJ: Colonic health: Fermentation and short chain fatty
acids. J Clin Gastroenterol. 40:235–243. 2006.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hijova E and Chmelarova A: Short chain
fatty acids and colonic health. Bratisl Lek Listy. 108:354–358.
2007.PubMed/NCBI
|
|
37
|
Wang FY, Liu JM, Luo HH, Liu AH and Jiang
Y: Potential protective effects of clostridium butyricum on
experimental gastric ulcers in mice. World J Gastroenterol.
21:8340–8351. 2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Lin Y, Fang ZF, Che LQ, Xu SY, Wu D, Wu CM
and Wu XQ: Use of sodium butyrate as an alternative to dietary
fiber: Effects on the embryonic development and anti-oxidative
capacity of rats. PLoS One. 9(e97838)2014.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Xing X, Jiang Z, Tang X, Wang P, Li Y, Sun
Y, Le G and Zou S: Sodium butyrate protects against oxidative
stress in HepG2 cells through modulating Nrf2 pathway and
mitochondrial function. J Physiol Biochem. 73:405–414.
2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Inan MS, Rasoulpour RJ, Yin L, Hubbard AK,
Rosenberg DW and Giardina C: The luminal short-chain fatty acid
butyrate modulates NF-kappaB activity in a human colonic epithelial
cell line. Gastroenterology. 118:724–734. 2000.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Yin L, Laevsky G and Giardina C: Butyrate
suppression of colonocyte NF-kappa B activation and cellular
proteasome activity. J Biol Chem. 276:44641–44646. 2001.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Luhrs H, Gerke T, Muller JG, Melcher R,
Schauber J, Boxberge F, Scheppach W and Menzel T: Butyrate inhibits
NF-kappaB activation in lamina propria macrophages of patients with
ulcerative colitis. Scand J Gastroenterol. 37:458–466.
2002.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Aziz RS, Siddiqua A, Shahzad M, Shabbir A
and Naseem N: Oxyresveratrol ameliorates ethanol-induced gastric
ulcer via downregulation of IL-6, TNF-α, NF-kB, and COX-2 levels,
and upregulation of TFF-2 levels. Biomed Pharmacother. 110:554–560.
2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
He H, Feng M, Xu H, Li X, He Y, Qin H,
Zhang Y, Tang H and Zou K: Total triterpenoids from the fruits of
chaenomeles speciosa exerted gastroprotective activities on
indomethacin-induced gastric damage via modulating
microRNA-423-5p-mediated TFF/NAG-1 and apoptotic pathways. Food
Funct. 11:662–679. 2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Longman RJ, Douthwaite J, Sylvester PA,
Poulsom R, Corfield AP, Thomas MG and Wright NA: Coordinated
localisation of mucins and trefoil peptides in the ulcer associated
cell lineage and the gastrointestinal mucosa. Gut. 47:792–800.
2000.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Katoh M: Trefoil factors and human gastric
cancer (review). Int J Mol Med. 12:3–9. 2003.PubMed/NCBI
|
|
47
|
Chen J, Zhao KN and Vitetta L: Effects of
intestinal microbial(-)elaborated butyrate on oncogenic signaling
pathways. Nutrients. 11(1026)2019.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Bultman SJ: Interplay between diet, gut
microbiota, epigenetic events, and colorectal cancer. Mol Nutr Food
Res. 61(10)2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Davie JR: Inhibition of histone
deacetylase activity by butyrate. J Nutr. 133 (7
Suppl):2485S–2493S. 2003.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Eslami M, Sadrifar S, Karbalaei M, Keikha
M, Kobyliak NM and Yousefi B: Importance of the microbiota
inhibitory mechanism on the warburg effect in colorectal cancer
cells. J Gastrointest Cancer. 51:738–747. 2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Hanson J, Gille A, Zwykiel S, Lukasova M,
Clausen BE, Ahmed K, Tunaru S, Wirth A and Offermanns S: Nicotinic
acid- and monomethyl fumarate-induced flushing involves GPR109A
expressed by keratinocytes and COX-2-dependent prostanoid formation
in mice. J Clin Invest. 120:2910–2919. 2010.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Benyo Z, Gille A, Kero J, Csiky M,
Suchánková MC, Nüsing RM, Moers A, Pfeffer K and Offermanns S:
GPR109A (PUMA-G/HM74A) mediates nicotinic acid-induced flushing. J
Clin Invest. 115:3634–3640. 2005.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Tarnawski AS: Cellular and molecular
mechanisms of gastrointestinal ulcer healing. Dig Dis Sci. 50
(Suppl 1):S24–S33. 2005.PubMed/NCBI View Article : Google Scholar
|