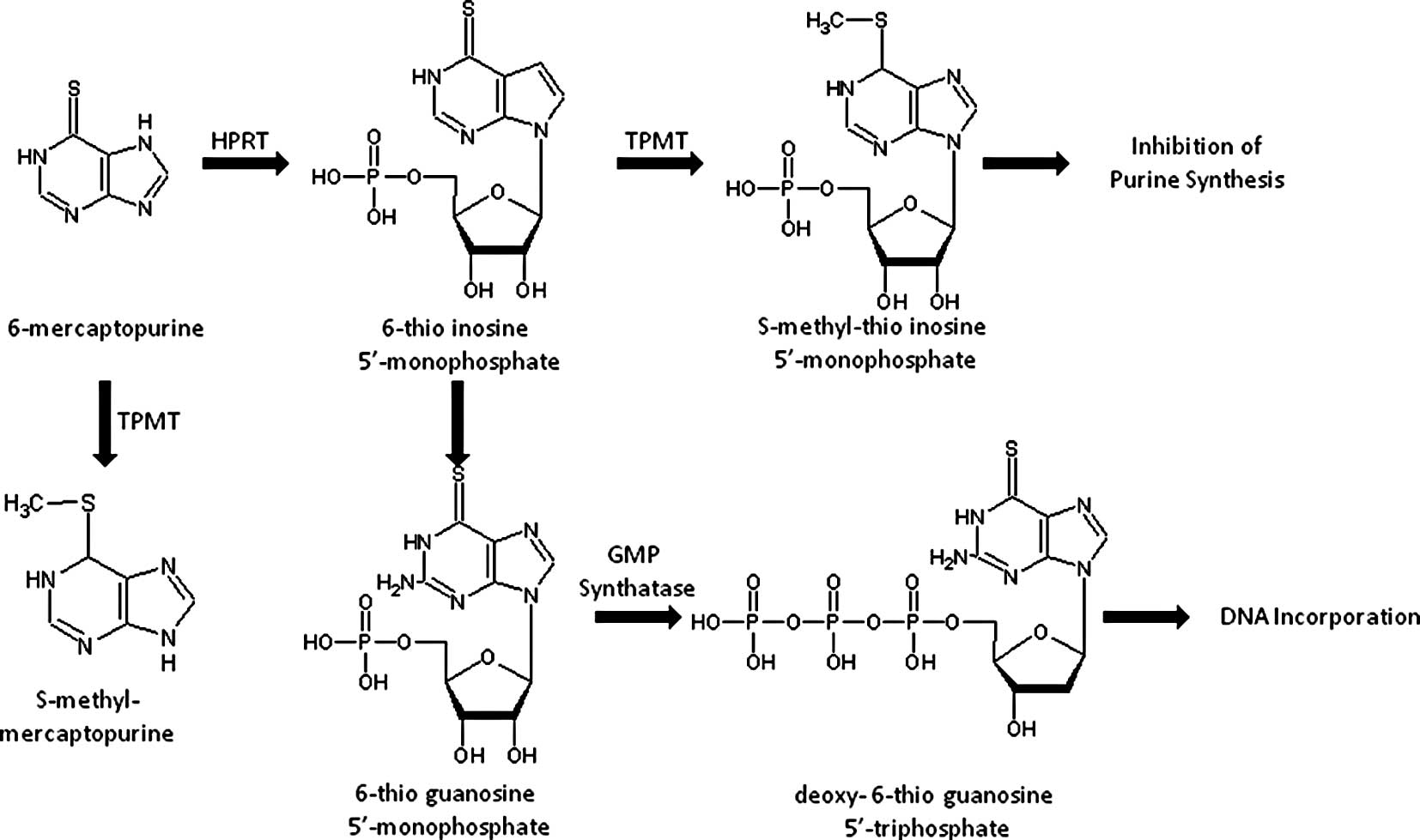
Introduction
Thiopurine S-methyltransferase [TPMT: EC 2.1.1.67
(MIM 187680)] is a cytoplasmic enzyme present in both prokaryotes
and eukaryotes. TPMT, first described by Remy (1), catalyzes the S-methylation of
aromatic compounds including thiopurine drugs such as
6-mercaptopurine, 6-thioguanine and azathiopurine (2,3).
Thiopurines are extensively used as chemotherapeutic agents in
patients with neoplasia and auto-immune disorders. The prodrugs,
thiopurines, are converted intracellularly to a thioguanine
nucleotide form which exerts a cytotoxic effect. The first step in
the intracellular activation of mercaptopurine involves being
catalyzed by hypoxanthine-guanine phosphoribosyltransferase (HPRT),
which yields thioinosine monophosphate (TIMP). Subsequently, TIMP
is converted to thioguanine triphosphate nucleotides, which are
incorporated into DNA. Excessive accumulation of thioguanine (TGN),
a cytotoxic compound, induces severe cytotoxicity. Fig. 1 illustrates the drug metabolic
pathway for thiopurine drugs by TPMT and the subsequent conversion
into an inactive substance thereby decreasing the amount of prodrug
available for active TGN (2).
The level of TPMT activity in human tissue is
controlled by genetic polymorphisms responsible for individual
differences in thiopurine toxicity and therapeutic efficacy
(4–6). Several studies have reported that
patients with low or undetectable TPMT activity are at high risk
for severe toxicity when treated with conventional dosages of
thiopurine drugs, while similar doses may be adequate to treat
patients with high levels of enzyme activity (5,6). Due
to these important clinical consequences, it is suggested that
patients be screened for TPMT genotype to determine the
enzyme activity before thiopurine therapy. The active gene for the
enzyme is 34 kb in length, consisting of 10 exons, and is localized
at 6q22.3 (4,7). The TPMT pseudogene has also
been reported and mapped to human chromosome band 18q21.1 (8).
TPMT enzyme activity shows trimodal distribution
with 89–94% of individuals possessing high activity, 6–11%
intermediate activity and 0.3% low activity. The wild-type allele
for high TPMT activity has been designated
TPMT*1. Several variant alleles for TPMT
(TPMT*2 to TPMT*20) have been
reported in many ethnic groups. A list of known alleles of the
TPMT gene is presented in Table
I, with the type of variations and the loci. Among these, four
mutant alleles namely TPMT*2,
TPMT*3A, TPMT*3B and
TPMT*3C have been identified as responsible for
enzyme deficiency in several populations. Another sixteen allelic
variants, TPMT*5 to TPMT*20
(Table I), have also been
suggested to be associated with deficient TPMT activity (9–11).
 | Table I.List of currently known mutant
alleles of the thiopurine S-methyltransferase (TPMT)
gene. |
Table I.
List of currently known mutant
alleles of the thiopurine S-methyltransferase (TPMT)
gene.
| Allele | Nucleotide
change | Amino acid
change | Exon position |
|---|
|
TPMT*1 | Wild | Wild | |
|
TPMT*1A | C178T | Silent | 1 |
|
TPMT*1S | T474C | Silent | 6 |
|
TPMT*2 | G238C | Ala80Pro | 5 |
|
TPMT*3A | G460A | Ala154Thr | 7 |
| A719G | Try240Thr | 10 |
|
TPMT*3B | G460A | Ala154Thr | 7 |
|
TPMT*3C | A719G | Try240Thr | 10 |
|
TPMT*3D | G292T | Glu98-Stop | 5 |
| G460A | Ala154Thr | 7 |
| A719G | Try240Thr | 10 |
|
TPMT*4 | G-A | Splice junction
intron 9/exon 10 | |
|
TPMT*5 | T146C | Leu49Ser | 4 |
|
TPMT*6 | A539T | Tyr180phe | 8 |
|
TPMT*7 | T681G | His227Glu | 10 |
|
TPMT*8 | G644A | Arg215His | 10 |
|
TPMT*9 | A356C | Lys119Thy | 5 |
|
TPMT*10 | G430C | Gly144Arg | 7 |
|
TPMT*11 | G395A | Cys132Thy | 6 |
|
TPMT*12 | C374T | Ser125Leu | 6 |
|
TPMT*13 | A83T | Glu28Val | 3 |
|
TPMT*14 | A1G | Met1Val | 3 |
|
TPMT*15 | G-A | Splice junction
intron 7/exon 8 | |
|
TPMT*16 | G488A | Arg163His | 7 |
|
TPMT*17 | C124G | Gln42Glu | 3 |
|
TPMT*18 | G211A | Gly71Arg | 4 |
|
TPMT*19 | A356C | Lys122Thr | 5 |
|
TPMT*20 | G106C | Gly36Ser | 3 |
The first identified variant allele,
TPMT*2, contains a transversion c.238G>C,
leading to substitution of p.Ala80Pro residue. It is a rare allele
reported to be found in European Caucasians and African-Americans
(12–14). The TPMT*3A mutant
allele contains two nucleotide transitions, c.460G>A and
c.719A>G, in the open reading frame, leading to the substitution
of amino acids p.Ala154Thr and p.Tyr240Cys, respectively, and are
found in African-American, European Caucasians and Southwest
Asians. It is the most common allele among the European Caucasian
population (13,15). TPMT*3B, that
contains the transition c.460G>A, is a common allele in
Caucasian populations. TPMT*3C contains
transversion c.719A>G and is the most prevalent allele among the
Chinese population. However, no similar data are available on
TPMT polymorphisms in Indian populations. Identifying the
most prevalent TPMT allele in Indian populations could
facilitate the deployment of rapid DNA-based assays for patients
before they are subjected to thiopurine drug therapy. Thus, the aim
of the study was to determine the frequency of TPMT variant
alleles in an Indian population in comparison to other populations.
The present study focused on the detection of signature alleles for
the TPMT gene, TPMT*2,
*3A, *3B and
*3C by using allele-specific (mutation-specific)
oligonucleotide polymerase chain reaction (ASO-PCR), polymerase
chain reaction, followed by restriction fragment length
polymorphism (PCR-RFLP) analysis and by confirmatory DNA sequencing
of the loci.
Materials and methods
Sample collection
Participants for the study were recruited randomly
from among the population of Southern India. A total of 326 (176
males; 150 females) unrelated healthy Indian individuals were
recruited, with a mean age of 31.4 years (range, 18–69 years).
Venous blood (4 ml) was obtained from each participant in an EDTA
vacutainer. The study was approved by the Institutional Ethics
Committee of Manipal University as per the guidelines of the Indian
Council of Medical Research, and written informed consent was
obtained from all participants.
Isolation of genomic DNA and polymerase
chain reaction
Genomic DNA was isolated from all the samples
collected by the standard phenol-chloroform extraction method
(16), and the three major
TPMT polymorphisms were genotyped in each sample. The
genotypes of the TPMT gene were analyzed for the three SNP
loci, NM_000367.2:c.238G>C, NM_000367.2:c.460G>A and
NM_000367.2:c.719A>G, to determine the frequencies of
TPMT*2, *3A,
*3B and *3C alleles and their
genotypes. Genotyping of the NM_000367.2:c.238G>C locus was
performed by ASO-PCR and those of NM_000367.2:c.460G>A and
NM_000367.2:c.719A>G loci were performed by polymerase chain
reaction, followed by PCR-RFLP. All PCR reactions were performed
with 30–50 ng of DNA in 25 μl of the total reaction mixture
containing 0.15 mM dNTPs, 20 pmol of each primer and 1 unit of Taq
DNA polymerase.
Allele-specific oligonucleotide
polymerase chain reaction for NM_000367.2:c.238G>C
The NM_000367.2:c.238G>C polymorphism was
analyzed by ASO-PCR to test for the presence of the
TPMT*2 allele. DNA was amplified using a common
reverse primer, 5′-TAAATAGGAACCATCGGAC AC-3′ and forward primer
with the single allele discriminating base at their 3′ end used in
a separate wild-type allele specific or mutant allele specific PCR
reaction was either a) 5′-GTA TGATTTTATGCAGGTTTG-3′ or b)
5′-GTATGATTTTATG CAGGTTTC-3′, respectively, as previously reported.
PCR thermal cycling conditions were: i) an initial 5-min
denaturation at 94°C; ii) 34 cycles at 94°C (30 sec), 56°C (30 sec)
and 72°C (60 sec); and iii) a final extension for 5 min at 72°C.
Amplified 254-bp PCR product was visualized by electrophoresis
using 2% of agarose gel containing ethidium bromide (0.5
μg/ml).
Genotyping of TPMT*3B and
TPMT*3C by polymerase chain reaction-restriction
fragment length polymorphism
The TPMT*3B
(NM_000367.2:c.460G>A) polymorphism was genotyped by employing
PCR-RFLP. The forward primer for PCR was
5′-AGGCAGCTAGGGAAAAAGAAAGGTG-3′ and reverse primer
5′-CAAGCCTTATAGCCTTACACCCAGG-3′. PCR thermal cycling conditions
were: i) an initial 5-min denaturation at 94°C; ii) 34 cycles at
94°C (30 sec), 62°C (30 sec) and 72°C (60 sec); and iii) a final
extension for 5 min at 72°C. The 694-bp PCR amplicon containing
c.460G>A was digested with the restriction enzyme MwoI,
and the DNA fragments were separated on a 2% agarose gel to
discriminate the alleles. The wild-type allele contains the
MwoI restriction site; however, the restriction site is
absent in the mutant allele.
The analysis of the TPMT*3C
polymorphism was performed by PCR-RFLP. DNA was amplified using the
forward primer 5′-GAGACAGAGTTTCACCATCTTGG-3′ and reverse primer
5′-CAGGCTTTAGCATAATTTTCAAT TCCTC-3′. PCR thermal cycling conditions
were: i) an initial 5-min denaturation at 94°C; ii) 34 cycles at
94°C (30 sec), 62°C (30 sec) and 72°C (60 sec); and iii) a final
extension for 5 min at 72°C. The 373-bp PCR amplicon containing
c.719A>G was digested with the restriction enzyme AccI,
and the DNA fragments were separated on a 2% agarose gel to
discriminate the alleles. The mutant type allele introduces an
AccI restriction site which is absent in the wild-type
allele.
DNA sequencing
The target TPMT PCR amplicons were sequenced
in both forward and reverse directions for all the available
genotypes for TPMT*2, *3B and
*3C. Amplified PCR products were purified after
eluting the fragments from the agarose gel using the standard
isopropanol precipitation method, followed by direct sequencing
using the Big Dye Terminator Cycle Sequencing Ready Reaction Mix
Kit v3.1 (ABI, Foster City, CA, USA) according to the
manufacturer's instructions. DNA sequencing was performed in an
ABI, 3130 genetic analyzer, and the electropherograms of the DNA
sequences in sense and antisense directions were analyzed for
detection of mutations using the Bioinformatics Tools and Human
Genome Nucleotide Reference Sequence NT_007592.14.
Statistical analysis
Subjects were genotyped, and allele frequencies were
calculated by counting the alleles. The 95% confidence interval
(CI) was calculated for all proportions of alleles and genotypes.
The Chi-square test was performed to test for deviation from
Hardy-Weinberg equilibrium for each locus.
Results
Our study determined the allele and genotype
frequencies of the three TPMT gene polymorphisms,
c.238G>C, c.460G>A and c.719A>G in a sample group of 326
individuals drawn from a Southern Indian population. Agarose gel
electrophoresis images representative of genotyping experiment
results by ASO-PCR and PCR-RFLP are depicted in Fig. 2. A summary of the results of allele
and genotype frequencies at the three loci (c.238G>C,
c.460G>A and c.719A>G) are presented with their Chi-square
test P-values in Table II. The
allele and genotype frequencies involving TPMT*1,
*2, *3A, *3B
and*3C alleles in the population sample were also
calculated with 95% CI and are presented in Tables III and IV, respectively.
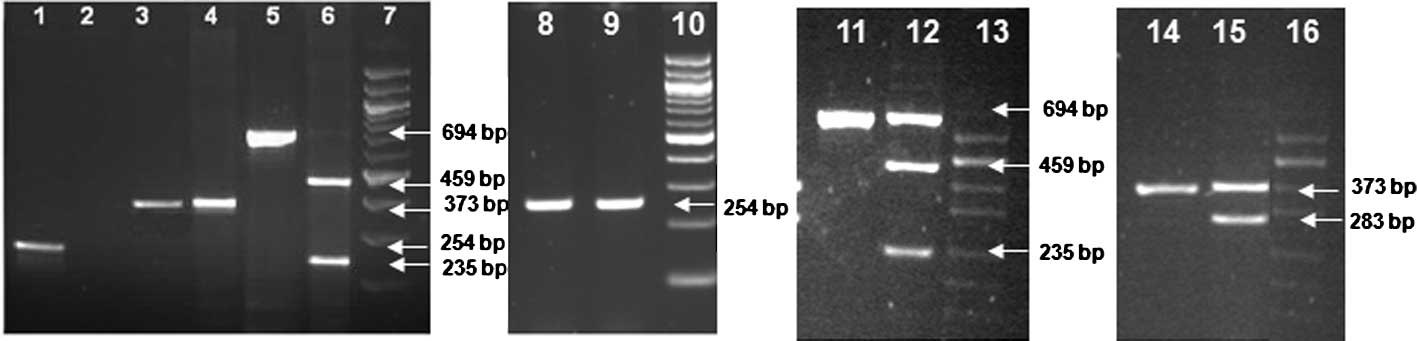 | Figure 2.Images of PCR amplicons containing
SNP loci of TPMT, NM_000367.2:c.238G>C,
NM_000367.2:c.460G>A and NM_000367.2:c.719A>G, and their
restriction fragments after agarose gel electrophoresis. All the
lanes in the consecutive gel images are numbered sequentially from
1 to 16. Sizes of the DNA fragments are indicated by arrows shown
in the image of 1.5% agarose gel electrophoresis. Lanes 7, 10, 13
and 16 show DNA size markers; 7 and 11 are 100-bp DNA ladder, and
lanes 13 and 16 are pUC/Hind III DNA ladder. Lanes 1–6 show
the wild-type alleles of the three TPMT polymorphisms
(NM_000367.2:c.238G>C, NM_000367.2:c.719A>G and
NM_000367.2:c.460G>A) and their restriction enzyme digestion
patterns; lane 1 shows the allele-specific amplicon representing
the wild-type c.238G allele; lane 2, which is blank, corresponds to
the mutant allele-specific reaction; lanes 3 and 4 represent PCR
amplicon with c.719A allele and the AccI restriction
enzyme-treated and undigested PCR amplicons, respectively; lanes 5
and 6 show the wild-type c.450G PCR amplicon and its MwoI
restriction endonuclease digested fragments of sizes 459 and 235
bp, respectively. Lanes 8–16 show the gel images of the PCR
amplicons with mutant alleles at the three TPMT SNP loci,
NM_000367.2:c.238G>C, NM_000367.2:c.460G>A and
NM_000367.2:c.719A>G, respectively. Lanes 8 and 9 show
allele-specific PCR amplicons of 254 bp from a heterozygote for the
c.238G>C polymorphism. Lane 11 shows PCR amplicon of the
heterozygous c.450G>A and the MwoI digested fragments of
the amplicon giving DNA fragments of 559 and 235 bp, respectively,
along with the wild-type undigested fragment of size 694 bp (lane
12). Lane 14 depicts the PCR amplicon of a heterozygote of
c.719A>G SNP and the AccI restriction enzyme-treated
undigested mutant allele, and the PCR fragments with wild-type
allele digested to fragments of 283 bp and a smaller one of 90 bp
that is not shown in the gel (lane 15). |
| Table II.Distribution of genotypes and alleles
of thiopurine S-methyltransferase, 238G>C, 460G>A and
719A>G single nucleotide polymorphismsa, in a Southern Indian
population, tested for Hardy-Weinberg equilibrium. |
Table II.
Distribution of genotypes and alleles
of thiopurine S-methyltransferase, 238G>C, 460G>A and
719A>G single nucleotide polymorphismsa, in a Southern Indian
population, tested for Hardy-Weinberg equilibrium.
| Locus | Genotype counts
(Frequency %) | Allele counts
(Frequency %) | P-valueb |
|---|
| c.238G>C
(rs1800462) | GG | GC | CC | G | C | |
|
| |
| 324 (99.39) | 2 (0.61) | 0 | 650 (99.69) | 2 (0.31) | 0.96 |
| c.460G>A
(rs1800460) | GG | GA | AA | G | A | |
|
| |
| 324 (99.39) | 2 (0.61) | 0 | 650 (99.69) | 2 (0.31) | 0.96 |
| c.719A>G
(rs1142345) | AA | AG | GG | A | G | |
|
| |
| 321 (98.47) | 5 (1.53) | 0 | 647 (99.23) | 5 (0.77) | 0.89 |
| Table III.Functional variants of thiopurine
S-methyltransferase (TPMT) gene alleles and their
distribution in a Southern Indian population with their 95%
confidence interval (CI). |
Table III.
Functional variants of thiopurine
S-methyltransferase (TPMT) gene alleles and their
distribution in a Southern Indian population with their 95%
confidence interval (CI).
| Type of allele | No. of alleles | Percentage of
allele | 95% CI |
|---|
|
TPMT*1 | 643 | 98.62 | 97.7–99.5 |
|
TPMT*2 | 2 | 0.31 | 0.0–0.7 |
|
TPMT*3A | 0 | 0.00 | - |
|
TPMT*3B | 2 | 0.31 | 0.0–0.7 |
|
TPMT*3C | 5 | 0.76 | 0.1–1.4 |
| Total | 652 | 100.00 | |
| Table IV.Distribution of genotype frequencies
of thiopurine S-methyltransferase (TPMT) gene functional
variants (involving TPMT*1, *2,
*3A, *3B and
*3C alleles) in a Southern Indian population and
their Chi-square test result for Hardy-Weinberg equilibrium. |
Table IV.
Distribution of genotype frequencies
of thiopurine S-methyltransferase (TPMT) gene functional
variants (involving TPMT*1, *2,
*3A, *3B and
*3C alleles) in a Southern Indian population and
their Chi-square test result for Hardy-Weinberg equilibrium.
| Genotype | No. of samples | Frequency (%) | 95% CI |
|---|
| Distribution of
genotypes for individual loci | | | |
|
TPMT*1/TPMT*1 | 317 | 97.24 | 95.5–99.0 |
|
TPMT*1/TPMT*2 | 2 | 0.61 | 0.0–1.5 |
|
TPMT*1/TPMT*3A | 0 | 0.00 | 0 |
|
TPMT*1/TPMT*3B | 2 | 0.61 | 0.0–1.5 |
|
TPMT*1/TPMT*3C | 5 | 1.53 | 0.2–2.9 |
| Total | 326 | 100.00 | |
| Distribution of
genotypes for all loci | | | |
| Homozygous wild
(Total) | 317 | 97.24 | 95.5–99.0 |
| Heterozygous mutant
(Total) | 9 | 2.76 | 0.98–4.5 |
| Homozygous
mutant | 0 | 0.00 | 0 |
| Total | 326 | 100.00 | |
Of the 326 healthy Indian participants investigated
in this study, 317 were found to be homozygous for
TPMT*1/*1 (97.24%; 95% CI, 95.5–99.0).
Two participants were heterozygous for the alleles
TPMT*1/*2 (0.61%; 95% CI, 0–1.5), 2
were heterozygous for alleles
TPMT*1/*3B (0.61%; 95% CI, 0–1.5) and
5 were heterozygous for alleles
TPMT*1/*3C (1.53%; 95% CI, 0.2–2.9)
among all the individuals of the group. The
TPMT*3A allele was not found in individuals from
the investigated population in this study, and no individual was
found homozygous for mutant alleles (Table IV). The allele and genotype
frequencies did not show any deviation from Hardy-Weinberg
equilibrium as shown by the Chi-square test results
(χ2=0.06; P=0.8).
Discussion
TPMT catalyzes the S-methylation of thiopurine drugs
such as 6-mercaptopurine and 6-thioguanine to form inactive
metabolites, a pathway that competes with xanthine oxidase-mediated
conversion of 6-MP to the inactive 6-thiouric acid and
HPRT-mediated conversion of 6-MP into the active 6-thioguanine
nucleotide. Patients with inherited low levels of TPMT enzyme
activity are at increased risk for thiopurine-induced toxicity when
treated with thiopurine drugs such as 6-mercaptopurine,
6-thioguanine and azathioprine; and those with inherited higher
levels of enzyme activity may be treated inadequately. Since these
drugs have a narrow therapeutic window, toxicities such as
life-threatening myelosuppression assume great importance. The
TPMT genetic polymorphism represents a good example of the
well established clinical importance of pharmacogenetic variation
of a drug-metabolizing enzyme. The prevalence of various
TPMT alleles in the Caucasian population is reported to be
0.3% which occur as homozygous mutants, 11% which occur as
heterozygous and 89% as homozygous wild-types (17,18).
Homozygous-mutated individuals are at increased risk for
potentially life-threatening toxicity when exposed to standard
doses of thiopurine drugs, while heterozygous carriers are advised
to begin therapy with 50–60% of the recommended dose (19). The molecular basis of TPMT
deficiency is well understood, where TPMT*2,
*3A, *3B and
*3C are the most common variants detected in more
than 80% of individuals with low or intermediate TPMT activity
(20,21). The frequencies of these alleles in
various populations of the world that are available in published
literature have been summarized in Table V.
| Table V.Frequencies (%) of thiopurine
S-methyltransferase (TPMT)*2,
*3A, *3B and
*3C alleles reported in different populations of
the world. |
Table V.
Frequencies (%) of thiopurine
S-methyltransferase (TPMT)*2,
*3A, *3B and
*3C alleles reported in different populations of
the world.
| Population | No. of participants
(n) |
TPMT*2 |
TPMT*3A |
TPMT*3B |
TPMT*3C | Reference |
|---|
| Indians | 326 | 0.61 | 0.0 | 0.61 | 1.53 | Present study |
| African
American | 248 | 0.40 | 0.8 | N/D | 2.40 | (9) |
| American
Caucasian | 282 | 0.20 | 3.2 | N/D | 0.20 | (9) |
| Brazilian | 306 | 0.80 | 2.0 | N/D | 2.50 | (26) |
| Kenya | 101 | 0.00 | 0.0 | 0.00 | 5.40 | (27) |
| British
Caucasian | 199 | 0.50 | 4.5 | 0.00 | 0.30 | (28) |
| South West
Asian | 99 | 0.00 | 1.0 | 0.00 | 0.00 | (28) |
| Egyptian | 200 | 0.00 | 0.3 | 0.00 | 1.30 | (30) |
| French
Caucasian | 469 | 3.00 | 0.7 | N/D | 0.40 | (31) |
| Italian | 103 | 0.50 | 3.9 | N/D | 1.00 | (32) |
| Norwegian
Caucasian | 66 | 0.00 | 3.4 | N/D | 0.30 | (35) |
| Saami | 194 | 0.00 | 0.0 | N/D | 3.30 | (35) |
| Polish | 358 | 0.40 | 2.7 | 0.00 | 0.10 | (37) |
| South East
Asian | 300 | 0.00 | 0.0 | 0.00 | 1.00 | (38) |
| Swedish | 800 | 0.10 | 3.8 | 0.10 | 0.40 | (39) |
| Thai | 200 | 0.00 | 0.0 | 0.00 | 5.00 | (40) |
To date, no studies have been carried out in the
Southern Indian population to determine the frequencies of the
TPMT alleles that are important in thiopurine drugs
pharmacogenomics. In this study, we did not include an analysis for
other alleles including rare variants such as
TPMT*3D, *4 and
*20. The promoter polymorphisms were also ignored
due to their minor effect on the modulation of TPMT activity
compared with variation in the open reading frame. In this study,
the TPMT*2 and *3B variants
were found at a low allele frequency of 0.31% for each. No
TPMT*3A allele was observed in the subjects
studied that appeared to be significant. However, the absence of
the TPMT*3A allele in this population needs to be
confirmed through study of an appropriately large sample of the
population. The most predominant mutant allele found in the
Southern Indian population in this study was the
TPMT*3C allele with a frequency of 0.76%,
representing 55.6% of the mutant alleles investigated. Therefore, a
test for this mutation alone in this population may prospectively
identify approximately half the proportion of individuals at risk
of hematopoietic toxicity in the event of administration of a
regular dose of thiopurine drugs. From an anthropological point of
view, the TPMT*3C allele was found in a Chinese
(22) and in a Ghanaian population
(23). It was the first
TPMT mutation to arise in humans, and this mutation is also
found in Caucasians, either alone in the TPMT*3C
allele or in combination with the TPMT*3B allele
in the form of TPMT*3A (24). It has been postulated that the
ancestral TPMT*3C allele founded prior to the
divergence of African and non-African populations was likely to
have evolved to contain a second mutation giving rise to the
TPMT*3A allele which predominated in American and
European Caucasians. The presence of the TPMT*3A
allele in African-Americans may be explained on the basis of genome
integration due to interethnic admixture (9,25,26).
Inhibitors of the activity of the enzyme may also have adverse
consequences for carriers of these mutant alleles, whether
homozygous or heterozygous. Furthermore, it should be noted that
the true significance of this human genetic variation may be better
explained in relation to the S-methylation of some endogenous
substrates yet to be identified, as opposed to the metabolism of
any group of drugs (41).
Our study was the first major study designed to
analyze the genetic polymorphisms of TPMT in 326 randomly
selected unrelated South Indian subjects from different locations
in this region of the country representing the heterogeneous ethnic
background of the population. Our study established the frequencies
of the TPMT alleles. Overall, 2.76% of the Indian subjects
were found to be heterozygous variants for TPMT alleles.
Conformity for Hardy-Weinberg equilibrium for the loci suggest that
no significant genotyping error was involved (Table II), that allele-based genetic
effects may not have been biased and that the mutations are not
relatively recent. Comparison of the mutation frequencies obtained
in this study with those reported in different ethnic groups shows
considerable difference between the Indian and other populations
(Table V). The present study
delineates more data on variations in the TPMT gene in
Indian populations. Identification of such variant allele
frequencies may help to predict both clinical efficacy and drug
toxicity of thiopurine drugs in Indian populations and to introduce
appropriate genetic screening protocols.
Acknowledgements
This study was supported by the
Technology Information Forecasting and Assessment Council-Centre of
Relevance and Excellence (TIFAC-CORE) in Pharmacogenomics, at the
Manipal Life Sciences Centre, Manipal University, India.
References
|
1.
|
Remy CN: Metabolism of thiopyrimidines and
thiopurines. S-Methylation with S-adenosylmethionine transmethylase
and catabolism in mammalian tissues. J Biol Chem. 238:1078–1084.
1963.
|
|
2.
|
Tidd DM and Paterson AR: A biochemical
mechanism for the delayed cytotoxic reaction of 6-mercaptopurine.
Cancer Res. 34:738–746. 1974.
|
|
3.
|
Lennard L, van Loon JA, Lilleyman JS and
Weinshilboum RM: Thiopurine pharmacogenetics in leukemia:
correlation of erythrocyte thiopurine methyltransferase activity
and 6-thioguanine nucleotide concentrations. Clin Pharmacol Ther.
41:18–25. 1987.
|
|
4.
|
Krynetski EY and Evans WE: Drug
methylation in cancer therapy: lessons from the TPMT polymorphism.
Oncogene. 22:7403–7413. 2003.
|
|
5.
|
Woodson LC, Dunnette JH and Weinshilboum
RM: Pharmacogenetics of human thiopurine methyltransferase:
kidney-erythrocyte correlation and immunotitration studies. J
Pharmacol Exp Ther. 222:174–181. 1982.
|
|
6.
|
Krynetski EY, Tai HL, Yates CR, Fessing
MY, Loennechen T, Schuetz JD, Relling MV and Evans WE: Genetic
polymorphism of thiopurine S-methyltransferase: clinical importance
and molecular mechanisms. Pharmacogenetics. 6:279–290. 1996.
|
|
7.
|
Snow JL and Gibson LE: The role of genetic
variation in thiopurine methyltransferase activity and the efficacy
and/or side effects of azathioprine therapy in dermatologic
patients. Arch Dermatol. 131:193–197. 1995.
|
|
8.
|
Lee D, Szumlanski C, Houtman J, Honchel R,
Rojas K, Overhauser J, Wieben ED and Weinshilboum RM: Thiopurine
methyltransferase pharmacogenetics. Cloning of human liver cDNA and
a processed pseudogene on human chromosome 18q21.1. Drug Metab
Dispos. 23:398–405. 1995.
|
|
9.
|
Hon YY, Fessing MY, Pui C-H, Relling MV,
Krynetski EY and Evans WE: Polymorphism of the thiopurine
S-methyltransferase gene in African-Americans. Hum Mol Genet.
8:371–376. 1999.
|
|
10.
|
Evans WE, Hon YY, Bomgaars L, Coutre S,
Holdsworth M, Janco R, Kalwinsky D, Keller F, Khatib Z, Margolin J,
Murray J, Quinn J, Ravindranath Y, Ritchey K, Roberts W, Rogers ZR,
Schiff D, Steuber C, Tucci F, Kornegay N, Krynetski EY and Relling
MV: Preponderance of thiopurine S-methyltransferase deficiency and
heterozygosity among patients intolerant to mercaptopurine or
azathioprine. J Clin Oncol. 19:2293–2301. 2001.
|
|
11.
|
Lindqvist M, Haglund S, Almer S, Peterson
C, Taipalensu J, Hertervig E, Lyrenäs E and Söderkvist P:
Identification of two novel sequence variants affecting thiopurine
methyltransferase enzyme activity. Pharmacogenetics. 14:261–265.
2004.
|
|
12.
|
Krynetski EY, Schuetz JD, Galpin AJ, Pui
C-H, Relling MV and Evans WE: A single point mutation leading to
loss of catalytic activity in human thiopurine S-methyltransferase.
Proc Natl Acad Sci USA. 92:949–953. 1995.
|
|
13.
|
Tai HL, Krynetski EY, Yates CR, Loennechen
T, Fessing MY, Krynetskaia NF and Evans WE: Thiopurine
S-methyltransferase deficiency: two nucleotide transitions define
the most prevalent mutant allele associated with loss of catalytic
activity in Caucasians. Am J Hum Genet. 58:694–702. 1996.
|
|
14.
|
Otterness D, Szumlanski C, Lennard L,
Klemetsdal B, Aarbakke J, Park-Hah JO, Iven H, Schmiegelow K,
Branum E, O'Brien J and Weinshilboum R: Human thiopurine
methyltransferase pharmacogenetics: gene sequence polymorphisms.
Clin Pharmacol Ther. 62:60–73. 1997.
|
|
15.
|
Otterness DM, Szumlanski CL, Wood TC and
Weinshilboum RM: Human thiopurine methyltransferase
pharmacogenetics. Kindred with a terminal exon splice junction
mutation that results in loss of activity. J Clin Invest.
101:1036–1044. 1998.
|
|
16.
|
Maniatis T, Fritsch EF and Sambrook J:
Analysis and cloning of eukaryotic genomic DNA. Molecular Cloning.
2nd edition. A Laboratory Manual. Cold Spring Harbor; New York: pp.
9.16–9.23. 1989
|
|
17.
|
Lennard L, Lilleyman JS, van Loon J and
Weinshilboum RM: Genetic variation in response to 6-mercaptopurine
for childhood acute lymphoblastic leukaemia. Lancet. 336:225–229.
1990.
|
|
18.
|
Weinshilboum R: Thiopurine
pharmacogenetics: clinical and molecular studies of thiopurine
methyltransferase. Drug Metab Dispos. 29:601–605. 2001.
|
|
19.
|
Coulthard SA, Hogarth LA, Little M,
Matheson EC, Redfern CPF, Minto L and Hall AG: The effect of
thiopurine methyltransferase expression on sensitivity to
thiopurine drugs. Mol Pharmacol. 62:102–109. 2002.
|
|
20.
|
Yates CR, Krynetski EY, Loennechen T,
Fessing MY, Tai HL, Pui CH, Relling MV and Evans WE: Molecular
diagnosis of thiopurine S-methyltransferase deficiency: genetic
basis for azathioprine and mercaptopurine intolerance. Ann Intern
Med. 126:608–614. 1997.
|
|
21.
|
Black AJ, McLeod HL, Capell HA, Powrie RH,
Matowe LK, Pritchard SC, Collie-Duguid ESR and Reid RM: Thiopurine
methyltransferase genotype predicts therapy-limiting severe
toxicity from azathioprine. Ann Intern Med. 129:716–718. 1998.
|
|
22.
|
Kham SK, Tan PL, Tay AH, Heng CK, Yeoh AE
and Quah TC: Thiopurine methyltransferase polymorphisms in a
multiracial Asian population and children with acute lymphoblastic
leukemia. J Pediatr Hematol Oncol. 24:353–359. 2002.
|
|
23.
|
Ameyaw MM, Collie-Duguid ES, Powrie RH,
Ofori-Adjei D and McLeod HL: Thiopurine methyltransferase alleles
in British and Ghanaian populations. Hum Mol Genet. 8:367–370.
1999.
|
|
24.
|
Leipold G, Schütz E, Haas JP and Oellerich
M: Azathioprine-induced severe pancytopenia due to a homozygous
two-point mutation of the thiopurine methyltransferase gene in a
patient with juvenile HLA-B27-associated spondylarthritis.
Arthritis Rheum. 40:1896–1898. 1997.
|
|
25.
|
Laróvere LE, de Kremer RD, Lambooy LH and
de Abreu RA: Genetic polymorphism of thiopurine S-methyltransferase
in Argentina. Ann Clin Biochem. 40:388–393. 2003.
|
|
26.
|
Reis M, Santoro A and Suarez-Kurtz G:
Thiopurine methyltransferase phenotypes and genotypes in
Brazilians. Pharmacogenetics. 13:371–373. 2003.
|
|
27.
|
McLeod HL, Pritchard SC, Githang'a J,
Indalo A, Ameyaw MM, Powrie RH, Booth L and Collie-Duguid ES:
Ethnic differences in thiopurine methyltransferase
pharmacogenetics: evidence for allele specificity in Caucasian and
Kenyan individuals. Pharmacogenetics. 9:773–776. 1999.
|
|
28.
|
Collie-Duguid ES, Pritchard SC, Powrie RH,
Sludden J, Collier DA, Li T and McLeod HL: The frequency and
distribution of thiopurine methyltransferase alleles in Caucasian
and Asian populations. Pharmacogenetics. 9:37–42. 1999.
|
|
29.
|
Isaza C, Henao J, López AM and Cacabelos
R: Allelic variants of the thiopurine methyltransferase
(TPMT) gene in the Colombian population. Methods Find Exp
Clin Pharmacol. 25:423–429. 2003.
|
|
30.
|
Hamdy SI, Hiratsuka M, Narahara K, Endo N,
El-Enany M, Moursi N, Ahmed MS-E and Mizugaki M: Genotype and
allele frequencies of TPMT, NAT2, GST, SULT1A1 and
MDR-1 in the Egyptian population. Br J Clin Pharmacol.
55:560–569. 2003.
|
|
31.
|
Ganiere-Monteil C, Medard Y, Lejus C,
Bruneau B, Pineau A, Fenneteau O, Bourin M and Jacqz-Aigrain E:
Phenotype and genotype for thiopurine methyltransferase activity in
the French Caucasian population: impact of age. Eur J Clin
Pharmacol. 60:89–96. 2004.
|
|
32.
|
Rossi AM, Bianchi M, Guarnieri C, Barale R
and Pacifici GM: Genotype-phenotype correlation for thiopurine
S-methyltransferase in healthy Italian subjects. Eur J Clin
Pharmacol. 57:51–54. 2001.
|
|
33.
|
Hiratsuka M, Inoue T, Omori F, Agatsuma Y,
Kishikawa Y and Mizugaki M: Detection assay of rare variants of the
thiopurine methyltransferase gene by PCR-RFLP using a mismatch
primer in a Japanese population. Biol Pharm Bull. 23:1090–1093.
2000.
|
|
34.
|
Wei H, Zhou S, Li C, Zhang J, Wu J and
Huang M: Phenotyping and genotyping studies of thiopurine
S-methyltransferase in Kazaks. Pharm Res. 22:1762–1766. 2005.
|
|
35.
|
Loennechen T, Utsi E, Hartz I, Lysaa R,
Kildalsen H and Aarbakke J: Detection of one single mutation
predicts thiopurine S-methyltransferase activity in a population of
Saami in northern Norway. Clin Pharmacol Ther. 70:183–188.
2001.
|
|
36.
|
Alves S, Prata MJ, Ferreira F and Amorim
A: Thiopurine methyltransferase pharmacogenetics: alternative
molecular diagnosis and preliminary data from Northern Portugal.
Pharmacogenetics. 9:257–261. 1999.
|
|
37.
|
Kurzawski M, Gawronska-Szklarz B and
Drozdzik M: Frequency distribution of thiopurine
S-methyltransferase alleles in a polish population. Ther Drug
Monit. 26:541–545. 2004.
|
|
38.
|
Chang JG, Lee LS, Chen CM, Shih MC, Wu MC,
Tsai FJ and Liang DC: Molecular analysis of thiopurine
S-methyltransferase alleles in South-east Asian populations.
Pharmacogenetics. 12:191–195. 2002.
|
|
39.
|
Haglund S, Lindqvist M, Almer S, Peterson
C and Taipalensuu J: Pyrosequencing of TPMT alleles in a general
Swedish population and in patients with inflammatory bowel disease.
Clin Chem. 50:288–295. 2004.
|
|
40.
|
Srimartpirom S and Tassaneeyakul W,
Kukongviriyapan V and Tassaneeyakul W: Thiopurine
S-methyltransferase genetic polymorphism in the Thai population. Br
J Clin Pharmacol. 58:66–70. 2004.
|
|
41.
|
Vesell ES: Therapeutic lessons from
pharmacogenetics. Ann Intern Med. 126:653–655. 1997.
|