Introduction
Chronic kidney disease (CKD) has a significant
impact on public health worldwide (1), and it is an important risk factor
for death and cardiovascular disease (2). CKD is characterized by the
progressive loss of renal function, chronic inflammation, oxidative
stress, vascular remodeling, as well as glomerular and
tubulointerstitial scarring (3).
Although the progression of CKD may depend upon the capacity of
residual nephrons to overcome stress, the mechanisms underlying the
progression of CKD remain poorly understood (2).
Although various animal CKD models of CKD have been
reported, rodent models tend to be more commonly employed in
experimental CKD studies (4). The
partially nephrectomized rat model has been used extensively to
investigate archetypal pathological changes in CKD. The remnant
kidney of nephrectomized rats exhibits adaptive, compensatory
growth in the days following injury, which is similar to the course
of human disease (4–6).
Shen-Kang (SK), a traditional Chinese medicine (TCM)
that confers protection against renal diseases (due to its
composition) (7–27), is comprised of 4 herbs used in
TCM: rhubarb (Rheum officinale), Salvia miltiorrhiza,
safflower (Carthamus tinctorius L.) and Astragalus
membranaceu). In our pre-experimental studies, in rats
subjected to 5/6 nephrectomy, treatment with SK attenuated chronic
clinical renal failure. However, the mechanisms underlying the
renal protective effects of SK remain to be elucidated.
Oxidative stress, which is characterized by
excessive levels of reactive oxygen species (ROS) due to an
imbalance between the oxidative and anti-oxidative systems, has
been implicated in various pathological conditions observed in CKD
(28–30). Renal proximal tubule epithelial
cells (HK-2 cells) are most frequently atrophic in CKD (1). During renal injury, elevated levels
of ROS in the proximal tubule promote lipid peroxidation and DNA
damage, eventually resulting in apoptotic cell death in the kidneys
(31–33).
In this study, we assessed the protective properties
of SK in rats subjected to 5/6 nephrectomy, as well as in renal
proximal tubule epithelial cells (HK-2 cells). An initial
evaluation of the beneficial effects of SK was performed using an
in vivo model of CKD (5/6 nephrectomized rats). These
observations were further evaluated using an in vitro model
with human renal proximal tubular epithelial HK-2 cells, in which
ROS production, apoptosis and cell viability were determined. In
addition, the molecular mechanisms underlying the potentially
protective effects of SK were also investigated.
Materials and methods
Materials
SK was obtained from Xi'an Shiji Shengkang
Pharmaceutical Industry Co., Ltd., (Xi'an, China). Enalapril (EN)
was purchased from Merck Millipore (Billerica, MA, USA).
Anti-extracellular signal-regulated kinase (ERK; #9102),
anti-phosphorylated ERK (p-ERK; #4370), anti-p38 (#9212), and
anti-phosphorylated p38 mitogen-activated protein kinase (p-p38
MAPK; #9216) antibodies were purchased from Cell Signaling
Technology (Danvers, MA, USA). Anti-c-Jun N-terminal kinase (JNK;
sc-571), anti-phosphorylated JNK (p-JNK; sc-6254),
anti-poly(ADP-ribose) polymerase (PARP; sc-8007) and anti-actin
(sc-47778) antibodies were obtained from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). Horseradish peroxidase (HRP)-conjugated
anti-mouse and anti-rabbit IgG antibodies were purchased from Merck
Millipore. Hydrogen peroxide (H2O2) was
purchased from Samchun Chemical Co. Ltd. (Seoul, Korea).
Animals
Six-week-old male Sprague-Dawley (SD) rats weighing
200±20 g were obtained from the Fourth Military Medical University
(Xi'an, China). The rats were maintained under a regular 12 h
light/dark cycle at stable room temperature for 1 week prior to the
commencement of the experiments. The rats were fed standard rodent
chow and had free access to tap water. All experimental procedures
were carried out according to the protocols approved by the Ethics
Committee for Animal Experimentation of the Fourth Military Medical
University and in accordance with the National Institutes of Health
Guide for the Care and Use of Laboratory Animals.
Surgical procedures and drug
administration
Each rat was anesthetized with chloral hydrate
solution (33 mg/100 g) via intraperitoneal injection. A total of 18
rats was subjected to 5/6 nephrectomy (5/6 Nx) in which, the upper
and lower poles of the left kidney and the entire right kidney were
removed, as previousy described (5,34,35). A sham operation was performed on 6
additional rats as a non-Nx control (sham-operated) group. The
nephrectomized rats were randomly divided into 3 groups as follows:
i) no treatment (5/6 Nx, n=6); ii) treatment with SK (5/6 Nx + SK,
n=6); and iii) treatment with EN (5/6 Nx + EN, n=6, positive
control). The rats in the treatment groups received either SK (450
mg/kg/day; via tail vein injection) or EN (5 mg/kg/day; via
intraperitoneal injection) daily for 1 week following surgery,
whereas the rats in the sham-operated group and 5/6 Nx (no
treatment) group received the vehicle (distilled water, 5
ml/kg/day) only. The animals were sacrificed by exsanguination at
day 30 post-surgery.
Biochemical analysis
Blood samples were collected from the orbital venous
plexus on days 0, 7 and 20 post-surgery. At the end of the
experiment, blood samples were obtained from the abdominal aorta,
immediately following sacrifice. The serum concentrations of blood
urea nitrogen (BUN) and serum creatinine (SCr) were determined
using standard laboratory procedures, as previously described
(36).
Histological analysis
After the rats were sacrificed, the kidney tissue
was removed from the abdominal cavity of the rats by surgical
methods (isolated kidney tissue from adipose tissue, renal
vasculature cut with scissors to obtain kidney tissue). Tissue was
fixed in formalin, dehydrated with ethanol, rendered transparent
with xylene, embedded with liquid paraffin and sliced with an
automatic slicing machine. Slices of renal tissue fixed in 10%
neutral-buffered formalin were embedded in paraffin, and
2-μm-thick sections were cut for morphological anlaysis.
These sections were stained with hematoxylin and eosin (H&E),
as previously descrbied (37).
All tissue samples were evaluated by an independent investigator
without prior knowledge of the group to which the rat belonged. All
glomeruli and the entire microscopic area of each specimen were
examined.
Cell culture
Renal proximal tubule epithelial cells (HK-2 cells)
were obtained from the American Type Culture Collection (ATCC,
Manassas, VA, USA). The HK-2 cells were passaged every 2–3 days in
100-mm dishes containing combined Dulbecco's modified Eagle's
medium/F-12 [DMEM/F12(1:1)] supplemented with 10% fetal bovine
serum, 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml
streptomycin (Life Technologies, Grand Island, NY, USA). The cells
were grown at 37°C in a humidified 5% CO2 atmosphere.
These cells were treated with H2O2 in the
presence or absence of SK at the indicated concentrations. For
experimental use, the cells were harvested at the end of the
treatment period for further analysis.
Cell viability assay
Cell viability was measured using the EZ-Cytox Cell
Viability Assay kit (MTT) assay (Itsbio, Seoul, Korea). MTT assay
is based on the cleavage of the tetrazolium salt, MTT, to the
water-insoluble formazan (38,39). The formazan dye produced by viable
cells can be quantified by measuring the absorbance of the dye
solution at 460 nm. The HK-2 cells were seeded in 96-well plates
(5×103 cells/well) at 37°C in a 5% CO2
incubator in DMEM/F12. Following an overnight incubation, the cells
were incubated for 24 h in the presence or absence of SK (300, 600
and 900 μg/ml) for 1 h prior to exposure to
H2O2 (200, 300, 400 and 500 μM). The
final incubation of the cells with 10 μl of kit reagent was
performed for 45 min at 37°C. The absorbance was measured at 460 nm
using a microplate reader (Bio-Rad, Hercules, CA, USA). Cell
viability was calculated and averaged. The cells from the control
group were treated in the same manner without
H2O2, and cell viability was expressed as a
percentage of the untreated controls.
Flow cytometric analysis
Early apoptosis and late apoptosis/necrosis induced
by H2O2 were detected quantitatively, using
an Annexin V-FITC apoptosis detection kit (Invitrogen, Grand
Island, NY, USA), as previously described (31). Briefly, the cells were treated
with H2O2 and/or SK, and then harvested by
centrifugation, washed with PBS, re-suspended in a
Ca2+-enriched binding buffer (10 mM HEPES, pH 7.4, 140
mM NaCl, and 2.5 mM CaCl2), incubated with 10 μM
fluorescein isothiocyanate (FITC)-conjugated Annexin V protein and
propidium iodide (PI) for 15 min in the dark and analyzed by
two-color flow cytometry. The cell samples were detected
immediately in the FL1-H and FL2-H channels of a FACSCalibur flow
cytometer (BD Biosciences, San Jose, CA, USA), which measured the
fluorescence at 488 nm excitation and 530 nm emission. The number
of apoptotic cells was quantified and the percentage of apoptotic
cells was calculated.
4′,6-Diamidino-2-phenylindole (DAPI)
staining
Changes in the nuclear morphology of apoptotic cells
were detected using the DNA-specific fluorescent dye, DAPI
(Vectashield; Vector Laboratories, Burlingame, CA, USA), as
previously described (31). The
HK-2 cells were grown on glass coverslips and treated with
H2O2 and/or SK for 24 h. The treated cells
were then fixed with 4% paraformaldehyde for 15 min at room
temperature, washed with PBS, permeablized with 0.2% Triton X-100
for 10 min at room temperature, washed with PBS again and 20
μl of mounting medium (DAPI) was then added to the fixed
cells for 5 min. The finalized slides were stored at 4°C, protected
from light and examined under a fluorescence microscope (Nikon,
Tokyo, Japan) in order to assess chromatin condensation and
fragmentation of the nuclei. Cells that exhibited a reduced nuclear
size, chromatin condensation, intense fluorescence and nuclear
fragmentation were considered apoptotic.
Determination of oxidative stress
The generation of intracellular ROS was measured
with dihydroethidium (DHE; Invitrogen) (a ROS fluorescent probe),
as previously described (1).
Briefly, the HK-2 cells were seeded onto 6 cm plates and incubated
with 500 μM H2O2 for 6 h in the
presence or absence of 300 μg/ml SK. At the end of the
experimental period, the cells were incubated with 5 μM DHE
for 30 min at 37°C, washed and then collected by centrifugation
(600 × g for 5 min at room temperature), and resuspended in PBS.
The fluorescence intensity was measured using a FACSCalibur™ flow
cytometer (BD Biosciences). An OxiSelect™ Total Antioxidant
Capacity (TAC) assay kit (Cell Biolabs, San Diego, CA, USA) was
employed to measure TAC, as previously described (40) and to estimate the reductive or
antioxidant capacity of biomolecules. Briefly, HK-2 cells were
incubated with 500 μM H2O2 for 6 h in
the presence or absence of 300 μg/ml SK, washed 3 times with
cold PBS, homogenized in cold PBS, and then centrifuged at 10,000 ×
g for 10 min at 4°C. The supernatant was stored at −80°C and
measured according to the manufacturer's instructions and with the
appropriate controls.
Western blot analysis
Following treatment with H2O2
and/or SK, the HK-2 cells were placed on ice, washed twice in
ice-cold PBS and lysed at 4°C for 30 min in cell lysis buffer
containing 50 mmol/l Tris-HCl, pH 7.5, 1% (v/v) Nonidet P-40, 250
mmol/l NaCl, 0.1 mmol/l phenylmethylsulfonyl fluoride, 0.1 mmol/l
sodium vanadate, 20 mmol/l β-glycerol phosphate, 2 mmol/l DTT, 1
mmol/l leupeptin and 10 mmol/l PNPP, as previously described
(31). The lysate was then
centrifuged at 16,000 × g for 20 min at 4°C. The supernatant was
collected for use in sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and the protein content was estimated
using a bovine serum albumin protein assay. Proteins were mixed
with sample buffer containing β-mercaptoethanol and heated at 100°C
for 2 min. A total of 40 μg of each cell lysate was
fractionated by SDS-PAGE on a 10% polyacrylamide gel and
transferred onto nitrocellulose membranes. After blocking with 5%
skim milk in Tris-buffered saline (TBS) containing 0.02% Tween-20
at room temperature for 1 h, the membranes were then incubated with
primary antibodies (diluted 1:1,000) overnight at 4°C. Actin
(diluted 1:5,000) was used as a loading control. Following
incubation with the primary antibodies, the blots were washed 4
times in TBS/Tween-20 prior to incubation for 1 h at room
temperature in goat anti-mouse or anti-rabbit HRP-conjugate
antibody at 1:2,000 dilution in TBS/Tween-20 containing 5% skim
milk. Following extensive washing in TBS/Tween-20, the blots were
processed for the detection of antigens using the enhanced
chemiluminescence system. Proteins were visualized with the
ECL-chemiluminescence kit (GE Healthcare Life Sciences, Logan, UT,
USA).
Statistical analysis
The quantification of the results of western blot
analysis was carried out using the ImageJ (1.47) software (version
1.47). Data are expressed as the means ± SEM of the 3 independent
experiments and analyzed by the Student's unpaired t-test (SPSS
version 17.0 software; SPSS Inc., Chicago, IL, USA). A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
Administration of SK attenuates the
development of glomerular lesions in rats induced by 5/6
nephrectomy
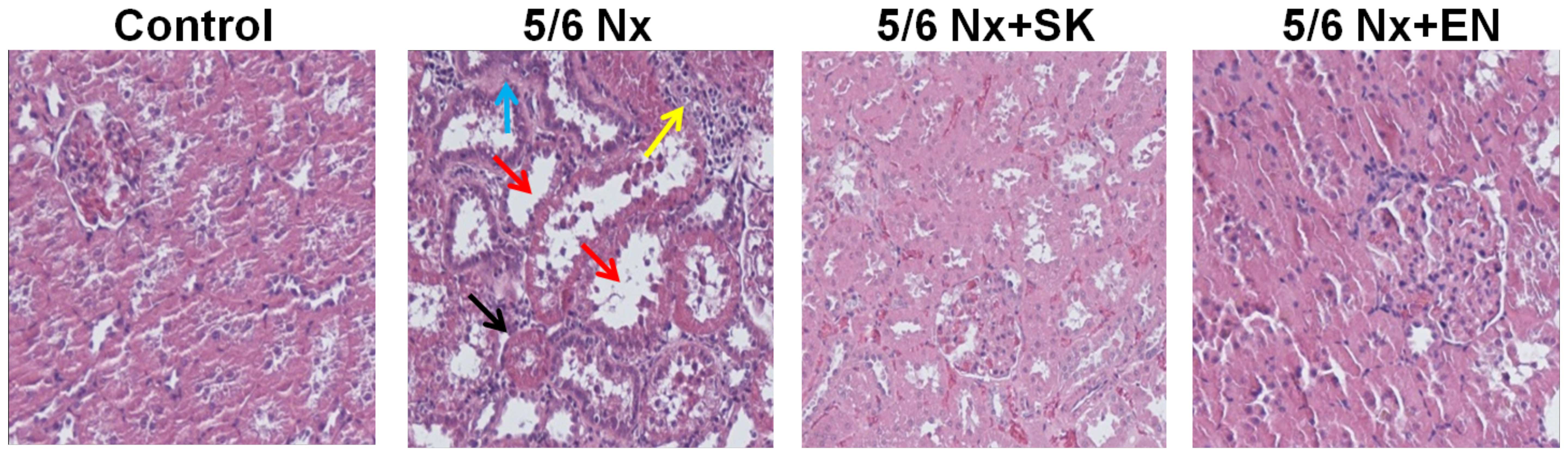
In order to evaluate the putative effects of SK on
renal function, histopathological changes in the renal sections of
5/6 nephrectomized rats were examined (Fig. 1). In the control (sham-operated)
group, proximal and distal tubules exhibited a normal structure (no
histological lesions; Fig. 1). A
significantly greater number of renal histological abnormalities
(glomerular sclerosis, tubular vacuoles, interstitial fibrosis and
inflammatory cell infiltration) was observed in the 5/6 Nx vs. the
sham-operated group. These abnormalities were markedly decreased in
the 5/6 Nx + SK group compared with the 5/6 Nx group, and similar
effects were also observed in the 5/6 Nx + EN-treated group
(Fig. 1). EN, an effective and
widely used drug in the treatment of CKD (41–47), was used as a positive control
treatment. These results indicate that SK may be beneficial in the
treatment of CKD.
Treatment with SK or EN improves renal
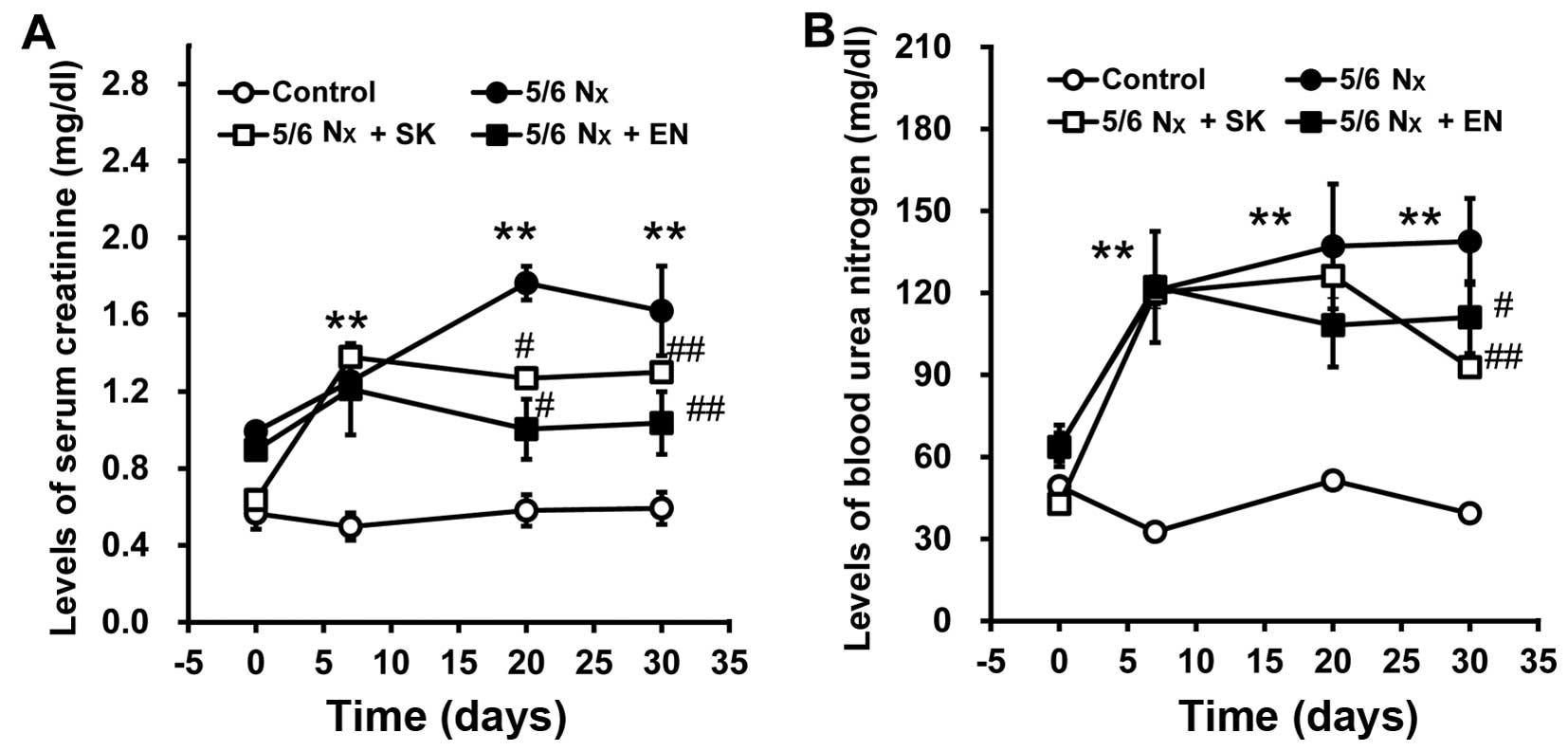
function in rats subjected to 5/6 nephrectomy
To evaluate renal function, SCr and BUN levels were
measured. The SCr and BUN levels significantly increased in the 5/6
Nx group vs. the sham-operated group (Fig. 2), indicating that the 5/6
nephrectomy model, reflecting an impairment in renal function, had
been successfully established. Notably, the SCr and BUN levels were
significantly decreased in the 5/6 Nx + SK and 5/6 Nx + EN groups
compared with the 5/6 Nx group (Fig.
2), which suggests that the administration of SK improves renal
function in a similar manner to EN.
SK protects against
H2O2-mediated cell death in HK-2 cells
To elucidate the molecular mechanisms underlying the
effects of SK, HK-2 cells were employed to examine the effects of
SK on the apoptosis of renal proximal tubule epithelial cells
induced by H2O2. The HK-2 cells were exposed
to H2O2 at various concentrations at 37°C for
24 h. Cell viability was then measured by MTT assay. As shown in
Fig. 3A, cell viability was
decreased in the H2O2-treated cells in a
dose-dependent manner (200–500 μM). Notably,
H2O2-induced cell death was inhibited by
treatment with SK (300, 600 and 900 μg/ml) (Fig. 3B) prior to exposure to
H2O2 (500 μM), suggesting that
treatment with SK effectively protects HK-2 cells against
H2O2-induced cell death.
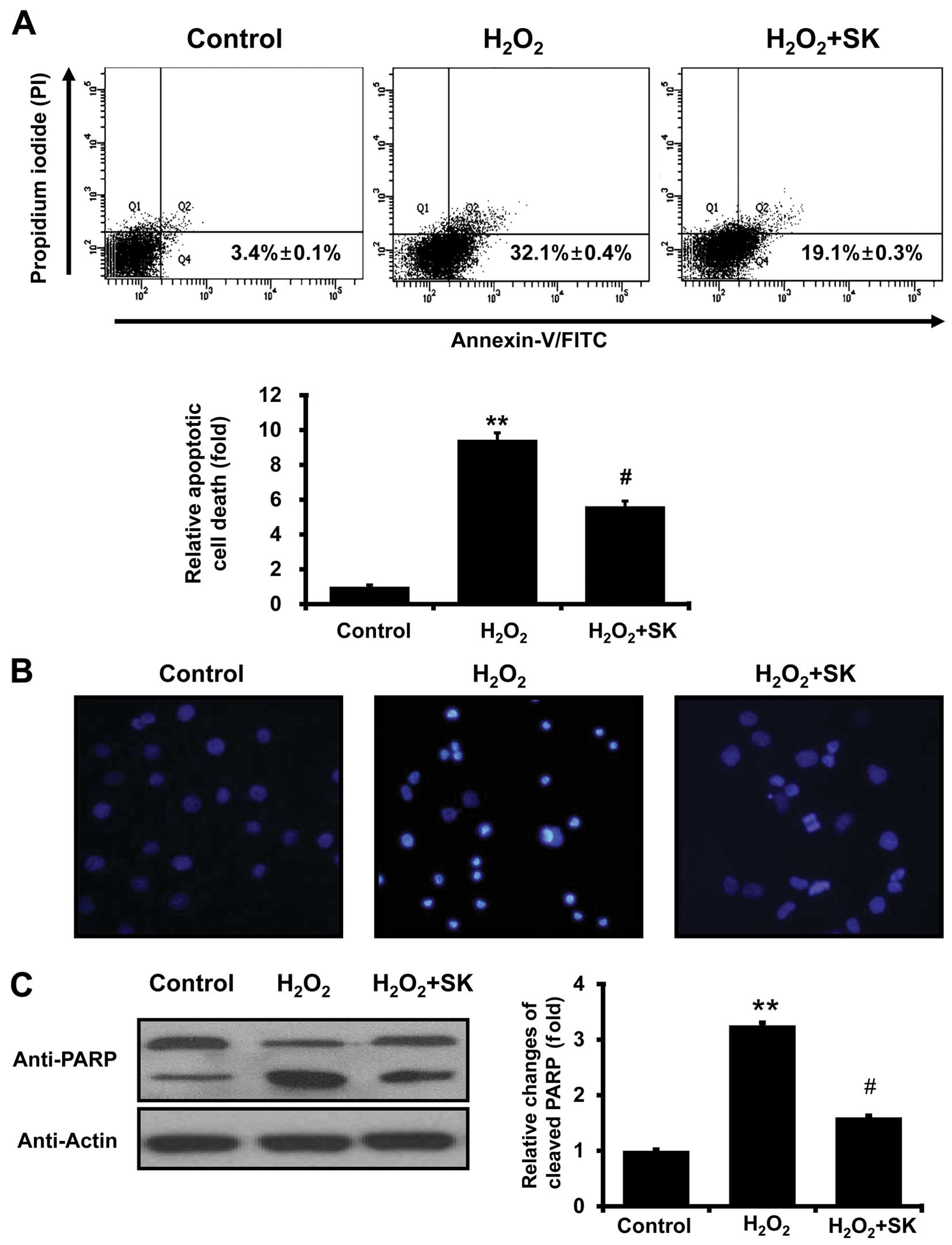
Morphological and biochemical effects of
H2O2 and SK on HK-2 cells
To further investigate the effects of SK on
H2O2-mediated cell death, Annexin V and PI
staining were employed to evaluate the events leading to cell
death. FACS analysis with Annexin V and PI staining revealed that
the apoptotic cell population, induced by exposure to
H2O2 for 24 h, was elevated from 3.4±0.1 to
32.1±0.4% (Fig. 4A); this
increase in the apoptotic cell population was prevented by
treatment with SK prior to exposure to H2O2
(apoptotic cell population decreased to 19.1±0.3%). Statistical
analysis further confirmed that treatment with SK protected the
HK-2 cells against apoptosis (induced by
H2O2; Fig.
4A, lower panel). To further evaluate the effects of SK,
morphological changes in the nuclei were observed using DAPI
staining with fluorescence microscopy. Normal nuclei were
characterized by homogeneous staining, and regular oval and rounded
shapes (Fig. 4B, left panel).
Apoptotic nuclei, indicated by condensed nuclei and nuclear
fragmentation, were apparent following exposure to 500 μM
H2O2 for 24 h (Fig. 4B, center panel). However, these
changes in nuclear characteristics were ameliorated by
pre-treatment with SK in the HK-2 cells (Fig. 4B, right panel). This was confirmed
by western blot analysis of PARP, which is cleaved under apoptotic
conditions (48,49). As illustrated in Fig. 4C, the cleaved form of PARP
significantly increased following exposure to
H2O2 for 24 h. The cleavage of PARP was
inhibited by pretreatment of the cells with SK (Fig. 4C), suggesting that SK prevents the
molecular events involved in apoptotic signaling.
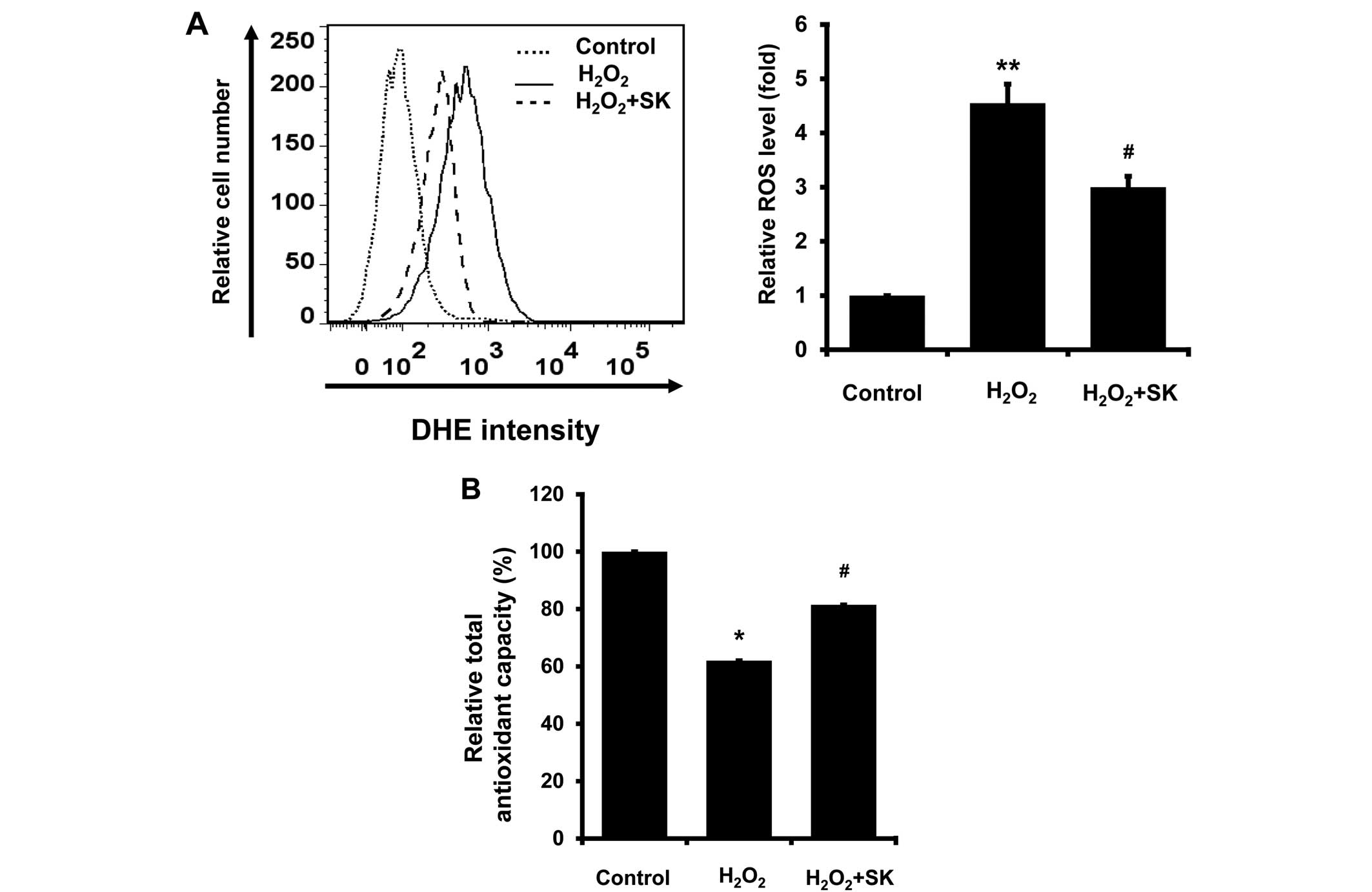
Inhibition of intracellular oxidative
stress following treatment with SK in HK-2 cells
It is well-established that oxidative stress
contributes to apoptosis (31–33). Therefore, the intracellular ROS
levels, and TAC, were measured in the HK-2 cells. ROS production,
measured by DHE staining, was elevated following exposure to
H2O2; treatment with SK significantly reduced
ROS production (Fig. 5A). In
addition, TAC was also enhanced by treatment with SK (Fig. 5B). Taken together, our findings
suggest that the SK-induced inhibition of ROS production is
mediated by enhanced TAC in HK-2 cells.
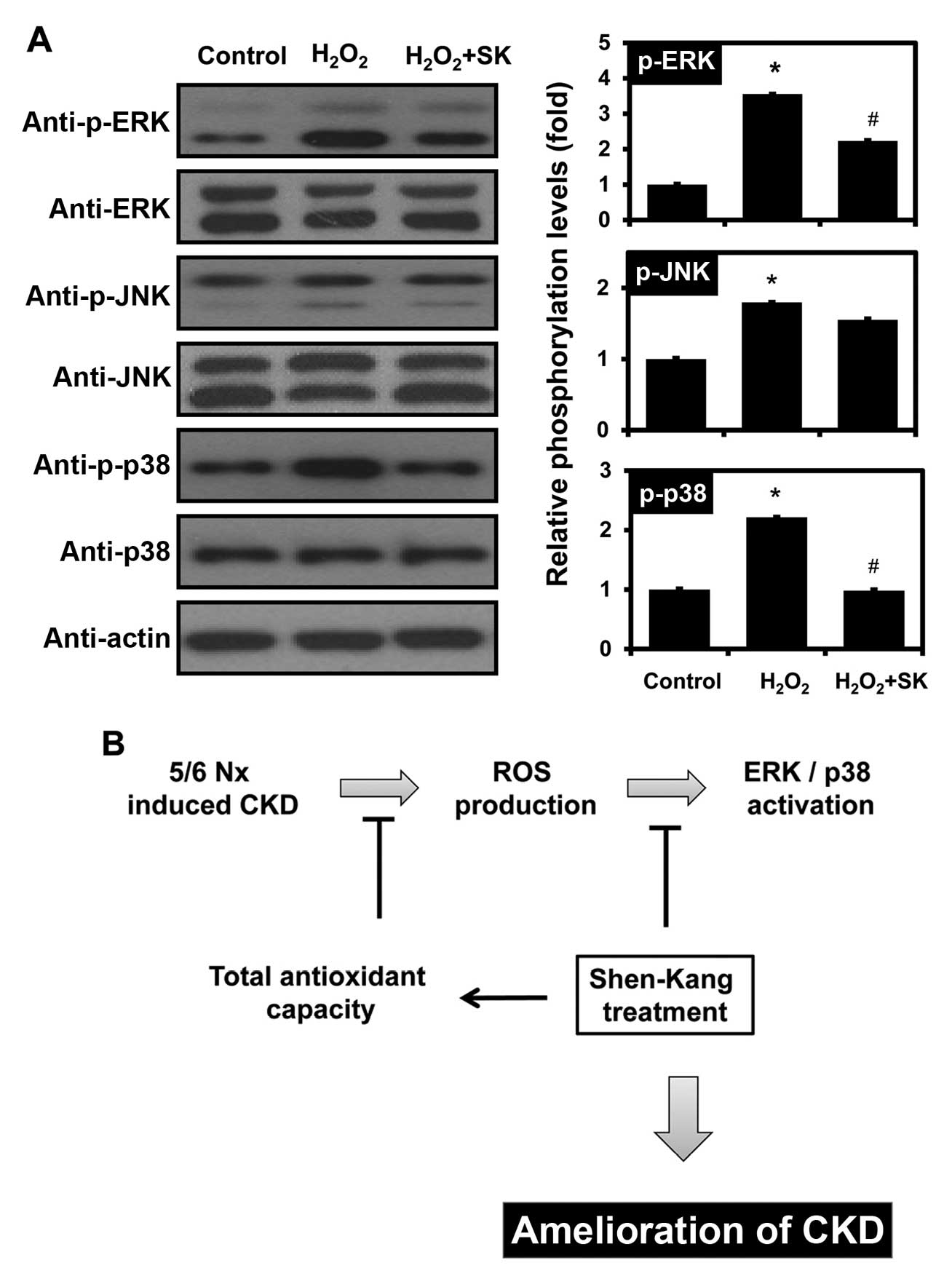
Modulation of MAPK signaling by SK in
HK-2 cells
To determine whether SK regulates the signaling
mechanisms responsible for H2O2-induced
apoptosis, the activation of different MAPK signaling pathways,
including ERK, JNK and p38 MAPK, was monitored by western blot
analysis. Notably, the H2O2-mediated
activation of ERK and p38 was observed, but JNK activation was not
affected by exposure to H2O2 (Fig. 6A). Furthermore, pre-treatment of
the HK-2 cells with SK inhibited the phosphorylation of ERK and
p38, but not that of JNK.
Discussion
CKD is a progressive, pathological condition
characterized by the progressive destruction of renal parenchyma
and the loss of functional nephrons over time (50). Various pathophysiological
conditions, including diabetes, hypertension, hyperlipidemia,
obesity, smoking and aging, are risk factors for the progression of
CKD (51–54). The clinical diagnosis of CKD is
dependent on the levels of BUN and SCr (50). In 2002, the National Kidney
Foundation Kidney Disease Outcomes Quality Initiative established a
novel CKD staging paradigm that characterized the progression of
CKD according to 5 stages (I–V) that result in unavoidable
end-stage renal disease (55–58). It is critical that novel
therapeutics or medications are developed to improve the
cost-effectiveness of renoprotective interventions, such as
TCM.
TCM has been used for over 2,500 years. Up to 80% of
the world's population uses herbs for medicinal purposes (59,60). TCM is frequently used in
conjunction with pharmacotherapy for the treatment of CKD in China
and many other Asian countries (61,62). TCM represents a major
complementary and alternative branch of CKD therapy (63). Clinical data indicate that SK and
its constituents, which include rhubarb, Salvia, safflower
and Astragalus, exert protective effects against renal
diseases (7–27). In China, SK is widely used in the
treatment of patients with renal diseases, such as CKD, diabetic
nephropathy, chronic renal failure, glomerulonephritis, chronic
nephritis and renal insufficiency. SK reduces pathological damage,
inhibits endothelial cell proliferation, attenuates proteinuria and
glomerular sclerosis and protects residual renal function and
attenuates disease progression (41–47).
The 5/6 nephrectomized rat model, involving
unilateral nephrectomy and either partial infarction or amputation
of the poles of the remaining kidney, represents the primary option
for the investigation of CKD (4,33,34). Lu et al reported kidney
dysfunction, with significantly elevated BUN and SCr levels and
histopathological changes (glomerular sclerosis and interstitial
fibrosis) in a 5/6 nephrectomized rat model (34). The 5/6 nephrectomized model of CKD
has demonstrable clinical relevance (5,35,64,65). SCr reflects the ability of the
renal system to remove creatinine from the blood and to concentrate
it in the urine. The diseased or damaged kidney is less able to
clear urea from the bloodstream, which results in elevated SCr and
BUN levels (66). In our 5/6
nephrectomized rat model, glomerular and interstitial pathogenesis
was significantly altered (Fig.
1); rats subjected to 5/6 nephrectomy exhibited markedly
increased SCr and BUN levels (Fig.
2), and this increase was reversed by 1 week of treatment with
SK, suggesting that SK restores kidney dysfunction in 5/6
nephrectomized rats.
Oxidative stress manifests as an increase in the
production of ROS, which arise from various cellular sources
(67,68). ROS represent a family consisting
of a large number of free-radical and non-free-radical molecules
derived from oxygen; the free radical reaction is a type of chain
reaction (29,69). Small et al (29), as well as others have clearly
demonstrated that oxidative stress is a unifying mechanism in CKD
pathogenesis (33,70). In addition, Lee et al
demonstrated that ROS are important mediators of apoptosis, as they
alter mitochondrial membrane potential (31). Tu et al provided evidence
that cell apoptosis, including in renal tubular epithelial HK-2
cells, is a critical determinant of renal fibrosis, which
eventually results in CKD (71).
Similarly, our results also indicated that treatment of HK-2 cells
with H2O2 promoted apoptosis (Figs. 3A and 4). This effect, attenuated by
concomitant SK treatment (Figs.
3B and 4), was further
evaluated by measuring ROS levels and TAC. The
H2O2-mediated ROS production in HK-2 cells
was reduced by pre-treatment of the cells with SK (Fig. 5). Of note, TAC was enhanced by SK
treatment (Fig. 5B), suggesting
that the upregulation of TAC by SK may underlie the SK-mediated
cell survival.
ROS also activate MAPKs, which include ERK, JNK and
p38 MAPK (72–75). The MAPK signaling pathways play a
vital role in the apoptosis of renal proximal tubule epithelial
cells, inflammation and renal injury (73,76–83). Shimizu et al reported that
the renal expression of monocyte chemotactic protein-1 is
upregulated by indoxyl sulfate through ROS production and the
activation of ERK and JNK in proximal tubular cells (77). Consistent with previous studies,
in this study, ERK and p38 MAPK phosphorylation was increased by
exposure of the cells to H2O2 (Fig. 6A). However, SK pretreatment
prevented the H2O2-mediated ERK and p38 MAPK
activation in HK-2 cells (Fig.
6A), indicating that SK protects HK-2 cells against
H2O2-induced apoptosis, by modulating TAC and
the MAPK signaling pathways (Fig.
6B).
Taken together, our data suggest that SK attenuates
glomerular sclerosis and interstitial fibrosis, reduces SCr and BUN
levels in rats subjected to 5/6 nephrectomy and abrogates the
H2O2-induced apoptosis of HK-2 cells. This
inhibitory effect may be partly mediated through the MAPK signaling
pathways and the upregulation of TAC. The association between TAC
and MAPK signaling requires further investigation.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81373947 and 81201985)
and by the National Research Foundation of Korea (NRF) grant funded
by the Korean government (MEST) (grant nos. 2007-0054932,
2012R1A1A2004714 and 2012M3A9B6055302).
Abbreviations:
|
CKD
|
chronic kidney disease
|
|
BUN
|
blood urea nitrogen
|
|
ROS
|
reactive oxygen species
|
|
PI
|
propidium iodide
|
References
|
1
|
Small DM and Gobe GC: Oxidative stress and
antioxidant therapy in chronic kidney and cardiovascular disease.
Oxidative Stress and Chronic Degenerative Diseases - A Role for
Antioxidants. Morales-Gonzales JA: InTech; pp. 233–264. 2013
|
|
2
|
Canaud G, Bienaimé F, Viau A, Treins C,
Baron W, Nguyen C, Burtin M, Berissi S, Giannakakis K, Muda AO, et
al: AKT2 is essential to maintain podocyte viability and function
during chronic kidney disease. Nat Med. 19:1288–1296. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Impellizzeri D, Esposito E, Attley J and
Cuzzocrea S: Targeting inflammation: new therapeutic approaches in
chronic kidney disease (CKD). Pharmacol Res. 81:91–102. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suzuki Y, Yamaguchi I, Onoda N, Saito T,
Myojo K, Imaizumi M, Takada C, Kimoto N, Takaba K and Yamate J:
Differential renal glomerular changes induced by 5/6
nephrectomization between common marmoset monkeys (Callithrix
jacchus) and rats. Exp Toxicol Pathol. 65:667–676. 2013. View Article : Google Scholar
|
|
5
|
Fedorova LV, Tamirisa A, Kennedy DJ,
Haller ST, Budnyy G, Shapiro JI and Malhotra D: Mitochondrial
impairment in the five-sixth nephrectomy model of chronic renal
failure: proteomic approach. BMC Nephrol. 14:2092013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen JF, Liu H, Ni HF, Lv LL, Zhang MH,
Zhang AH, Tang RN, Chen PS and Liu BC: Improved mitochondrial
function underlies the protective effect of pirfenidone against
tubulointerstitial fibrosis in 5/6 nephrectomized rats. PLoS One.
8:e835932013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Albersmeyer M, Hilge R, Schröttle A, Weiss
M, Sitter T and Vielhauer V: Acute kidney injury after ingestion of
rhubarb: secondary oxalate nephropathy in a patient with type 1
diabetes. BMC Nephrol. 13:1412012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hanzlicek AS, Roof CJ, Sanderson MW and
Grauer GF: The Effect of Chinese rhubarb, Rheum officinale, with
and without benazepril on the progression of naturally occurring
chronic kidney disease in cats. J Vet Intern Med. 28:1221–1228.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang J, Zhao Y, Xiao X, Li H, Zhao H,
Zhang P and Jin C: Assessment of the renal protection and
hepatotoxicity of rhubarb extract in rats. J Ethnopharmacol.
124:18–25. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan M, Zhang LY, Sun LX, Jiang ZZ and Xiao
XH: Nephrotoxicity study of total rhubarb anthraquinones on Sprague
Dawley rats using DNA microarrays. J Ethnopharmacol. 107:308–311.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ahn YM, Kim SK, Lee SH, Ahn SY, Kang SW,
Chung JH, Kim SD and Lee BC: Renoprotective effect of Tanshinone
IIA, an active component of Salvia miltiorrhiza, on rats with
chronic kidney disease. Phytother Res. 24:1886–1892. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Behradmanesh S, Derees F and
Rafieian-Kopaei M: Effect of Salvia officinalis on diabetic
patients. J Renal Inj Prev. 2:51–54. 2013.PubMed/NCBI
|
|
13
|
Chen G, Fu Y and Wu X: Protective effect
of Salvia miltiorrhiza extract against renal
ischemia-reperfusion-induced injury in rats. Molecules.
17:1191–1202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guan S, Ma J, Zhang Y, Gao Y, Zhang Y,
Zhang X, Wang N, Xie Y, Wang J, Zhang J, et al: Danshen (Salvia
miltiorrhiza) injection suppresses kidney injury induced by iron
overload in mice. PLoS One. 8:e743182013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee SH, Kim YS, Lee SJ and Lee BC: The
protective effect of Salvia miltiorrhiza in an animal model of
early experimentally induced diabetic nephropathy. J
Ethnopharmacol. 137:1409–1414. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li L, Zhang Y, Ma J, Dong W, Song Q, Zhang
J and Chu L: Salvia miltiorrhiza injection ameliorates renal damage
induced by lead exposure in mice. ScientificWorldJournal.
2014:5726972014.PubMed/NCBI
|
|
17
|
You Z, Xin Y, Liu Y, Han B, Zhang L, Chen
Y, Chen Y, Gu L, Gao H and Xuan Y: Protective effect of Salvia
miltiorrhiza e injection on N(G)-nitro-D-arginine induced nitric
oxide deficient and oxidative damage in rat kidney. Exp Toxicol
Pathol. 64:453–458. 2012. View Article : Google Scholar
|
|
18
|
Lo HC, Wang YH, Chiou HY, Lai SH and Yang
Y: Relative efficacy of casein or soya protein combined with palm
or safflower-seed oil on hyperuricaemia in rats. Br J Nutr.
104:67–75. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang YL, Chang SY, Teng HC, Liu YS, Lee
TC, Chuang LY, Guh JY, Chang FR, Liao TN, Huang JS, et al:
Safflower extract: a novel renal fibrosis antagonist that functions
by suppressing autocrine TGF-beta. J Cell Biochem. 104:908–919.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bai F, Makino T, Kono K, Nagatsu A, Ono T
and Mizukami H: Calycosin and formononetin from astragalus root
enhance dimethylarginine dimethylaminohydrolase 2 and nitric oxide
synthase expressions in Madin Darby Canine Kidney II cells. J Nat
Med. 67:782–789. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim J, Moon E and Kwon S: Effect of
Astragalus membranaceus extract on diabetic nephropathy. Endocrinol
Diabetes Metab Case Rep. 2014:1400632014.PubMed/NCBI
|
|
22
|
Li M, Wang W, Xue J, Gu Y and Lin S:
Meta-analysis of the clinical value of Astragalus membranaceus in
diabetic nephropathy. J Ethnopharmacol. 133:412–419. 2011.
View Article : Google Scholar
|
|
23
|
Lian Y, Xie L, Chen M and Chen L: Effects
of an astragalus polysaccharide and rhein combination on apoptosis
in rats with chronic renal failure. Evid Based Complement Alternat
Med. 2014:2718622014.PubMed/NCBI
|
|
24
|
Nie Y, Li S, Yi Y, Su W, Chai X, Jia D and
Wang Q: Effects of astragalus injection on the TGFβ/Smad pathway in
the kidney in type 2 diabetic mice. BMC Complement Altern Med.
14:1482014. View Article : Google Scholar
|
|
25
|
Song J, Meng L, Li S, Qu L and Li X: A
combination of Chinese herbs, Astragalus membranaceus var.
mongholicus and Angelica sinensis, improved renal microvascular
insufficiency in 5/6 nephrectomized rats. Vascul Pharmacol.
50:185–193. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang HW, Lin ZX, Xu C, Leung C and Chan
LS: Astragalus (a traditional Chinese medicine) for treating
chronic kidney disease. Cochrane Database Syst Rev.
10:CD0083692014.PubMed/NCBI
|
|
27
|
Zhang J, Xie X, Li C and Fu P: Systematic
review of the renal protective effect of Astragalus membranaceus
(root) on diabetic nephropathy in animal models. J Ethnopharmacol.
126:189–196. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miyamoto Y, Iwao Y, Mera K, Watanabe H,
Kadowaki D, Ishima Y, Chuang VT, Sato K, Otagiri M and Maruyama T:
A uremic toxin, 3-carboxy-4-methyl-5-propyl-2-furanpropionate
induces cell damage to proximal tubular cells via the generation of
a radical intermediate. Biochem Pharmacol. 84:1207–1214. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Small DM, Bennett NC, Roy S, Gabrielli BG,
Johnson DW and Gobe GC: Oxidative stress and cell senescence
combine to cause maximal renal tubular epithelial cell dysfunction
and loss in an in vitro model of kidney disease. Nephron Exp
Nephrol. 122:123–130. 2012. View Article : Google Scholar
|
|
30
|
Yu X, Liu H, Zou J, Zhu J, Xu X and Ding
X: Oxidative stress in 5/6 nephrectomized rat model: effect of
alpha-lipoic acid. Ren Fail. 34:907–914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee KE, Kim EY, Kim CS, Choi JS, Bae EH,
Ma SK, Park JS, Jung YD, Kim SH, Lee JU and Kim SW:
Macrophage-stimulating protein attenuates hydrogen peroxide-induced
apoptosis in human renal HK-2 cells. Eur J Pharmacol. 715:304–311.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zaza G, Granata S, Masola V, Rugiu C,
Fantin F, Gesualdo L, Schena FP and Lupo A: Downregulation of
nuclear-encoded genes of oxidative metabolism in dialyzed chronic
kidney disease patients. PLoS One. 8:e778472013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tapia E, Soto V, Ortiz-Vega KM,
Zarco-Márquez G, Molina-Jijón E, Cristóbal-García M, Santamaría J,
García-Niño WR, Correa F, Zazueta C and Pedraza-Chaverri J:
Curcumin induces Nrf2 nuclear translocation and prevents glomerular
hypertension, hyperfiltration, oxidant stress, and the decrease in
antioxidant enzymes in 5/6 nephrectomized rats. Oxid Med Cell
Longev. 2012:2690392012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lu JR, Han HY, Chen J, Xiong CX, Wang XH,
Hu J, Chen XF and Ma L: Protective effects of Bu-Shen-Huo-Xue
formula against 5/6 nephrectomy-induced chronic renal failure in
rats. Evid Based Complement Alternat Med.
2014:5898462014.PubMed/NCBI
|
|
35
|
Gava AL, Freitas FP, Balarini CM, Vasquez
EC and Meyrelles SS: Effects of 5/6 nephrectomy on renal function
and blood pressure in mice. Int J Physiol Pathophysiol Pharmacol.
4:167–173. 2012.PubMed/NCBI
|
|
36
|
Hamada K, Taniguchi Y, Shimamura Y, Inoue
K, Ogata K, Ishihara M, Horino T, Fujimoto S, Ohguro T, Yoshimoto
Y, et al: Serum level of soluble (pro)renin receptor is modulated
in chronic kidney disease. Clin Exp Nephrol. 17:848–856. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ding ZH, Xu LM, Wang SZ, Kou JQ, Xu YL,
Chen CX, Yu HP, Qin ZH and Xie Y: Ameliorating adriamycin-induced
chronic kidney disease in rats by orally administrated cardiotoxin
from Naja naja atra venom. Evid Based Complement Alternat Med.
2014:6217562014.PubMed/NCBI
|
|
38
|
Nagai J, Yamamoto A, Yumoto R and Takano
M: Albumin overload induces expression of hypoxia-inducible factor
1α and its target genes in HK-2 human renal proximal tubular cell
line. Biochem Biophys Res Commun. 434:670–675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kwon JT, Seo GB, Jo, Lee M, Kim HM, Shim
I, Lee BW, Yoon BI, Kim P and Choi K: Aluminum nanoparticles induce
ERK and p38MAPK activation in rat brain. Toxicol Res. 29:181–185.
2013. View Article : Google Scholar
|
|
40
|
Waly MI, Ali BH, Al-Lawati I and Nemmar A:
Protective effects of emodin against cisplatin-induced oxidative
stress in cultured human kidney (HEK 293) cells. J Appl Toxicol.
33:626–630. 2013. View Article : Google Scholar
|
|
41
|
Du J, Chen H and Wang XB: Effect of
shenkang injection on hypertrophy and expressions of p21 and p27 in
glomerular mesangial cells of rats cultured in high glucose.
Zhongguo Zhong Xi Yi Jie He Za Zhi. 26(Suppl): 68–71. 2006.In
Chinese.
|
|
42
|
Fu C, Song S and Shi Y: Observation on
effect of chongcao shenkang capsules in preventing and treating
acute renal failure in cases of hemorrhagic fever with renal
syndrome. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi.
13:188–190. 1999.In Chinese.
|
|
43
|
Guo L, Liu Y and Mao W: Contrast study on
effect of shenkang injection and benazepril on human glomerular
mesangial extracellular matrix. Zhongguo Zhong Xi Yi Jie He Za Zhi.
20:50–52. 2000.In Chinese.
|
|
44
|
Guo LZ, Mao W and Wang HM: Serological
study on inhibitory function of shenkang injection on glomerular
mesangial cell. Zhongguo Zhong Xi Yi Jie He Za Zhi. 21:531–533.
2001.In Chinese.
|
|
45
|
Xiao W, Wei LB, Ma Y, Long HB and Chen GB:
Renal protective effect of Shenkang pill on diabetic rats. Zhongguo
Zhong Yao Za Zhi. 31:1006–1009. 2006.In Chinese. PubMed/NCBI
|
|
46
|
Zhao Y, Wang JC, Li HY, Jia Q, Chen S, Xu
Z, Du X, Chen X, Lu L, Huang B and Long H: Shenkang pill
down-regulates AOPP-induced expression of inflammatory factor MCP-1
via a p38MAPK/NF-κB-dependent mechanism. Nan Fang Yi Ke Da Xue Xue
Bao. 34:1265–1271. 2014.In Chinese. PubMed/NCBI
|
|
47
|
Zhao Z, Li H and Zhang X: Effect of
shenkang injection on transforming growth factor-beta messenger
ribonucleic acid of LLC-PK1 renal tubular epithelial cells.
Zhongguo Zhong Xi Yi Jie He Za Zhi. 20:931–933. 2000.In
Chinese.
|
|
48
|
Chatterjee PK, Chatterjee BE, Pedersen H,
Sivarajah A, McDonald MC, Mota-Filipe H, Brown PA, Stewart KN,
Cuzzocrea S, Threadgill MD and Thiemermann C: 5-Aminoisoquinolinone
reduces renal injury and dysfunction caused by experimental
ischemia/reperfusion. Kidney Int. 65:499–509. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park SM, Jung HC, Koak IS, Na HY, Woo JS,
Jung JS and Kim YK: Oxidant-induced cell death in renal epithelial
cells: differential effects of inorganic and organic
hydroperoxides. Pharmacol Toxicol. 92:43–50. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Khan Z and Pandey M: Role of kidney
biomarkers of chronic kidney disease: An update. Saudi J Biol Sci.
21:294–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Padullés A, Rama I, Llaudó I and Lloberas
N: Developments in renal pharmacogenomics and applications in
chronic kidney disease. Pharmgenomics Pers Med. 7:251–266.
2014.PubMed/NCBI
|
|
52
|
Junaid Nazar CM, Kindratt TB, Ahmad SM,
Ahmed M and Anderson J: Barriers to the successful practice of
chronic kidney diseases at the primary health care level; a
systematic review. J Renal Inj Prev. 3:61–67. 2014.PubMed/NCBI
|
|
53
|
Lenz O, Mekala DP, Patel DV, Fornoni A,
Metz D and Roth D: Barriers to successful care for chronic kidney
disease. BMC Nephrol. 6:112005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Campbell RC, Sui X, Filippatos G, Love TE,
Wahle C, Sanders PW and Ahmed A: Association of chronic kidney
disease with outcomes in chronic heart failure: a
propensity-matched study. Nephrol Dial Transplant. 24:186–193.
2009. View Article : Google Scholar :
|
|
55
|
Onuigbo MA and Agbasi N: Chronic kidney
disease prediction is an inexact science: The concept of
'progressors' and 'nonprogressors'. World J Nephrol. 3:31–49. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Goolsby MJ: National kidney foundation
guidelines for chronic kidney disease: evaluation, classification,
and stratification. J Am Acad Nurse Pract. 14:238–242. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kopple JD: National kidney foundation
K/DOQI clinical practice guidelines for nutrition in chronic renal
failure. Am J Kidney Dis. 37(Suppl 2): S66–S70. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Levey AS, Coresh J, Balk E, Kausz AT,
Levin A, Steffes MW, Hogg RJ, Perrone RD, Lau J and Eknoyan G;
National Kidney Foundation: National Kidney Foundation practice
guidelines for chronic kidney disease: evaluation, classification,
and stratification. Ann Intern Med. 139:137–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li X and Wang H: Chinese herbal medicine
in the treatment of chronic kidney disease. Adv Chronic Kidney Dis.
12:276–281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Normile D: Asian medicine. The new face of
traditional Chinese medicine. Science. 299:188–190. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhong Y, Deng Y, Chen Y, Chuang PY and
Cijiang He J: Therapeutic use of traditional Chinese herbal
medications for chronic kidney diseases. Kidney Int. 84:1108–1118.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xu Q, Bauer R, Hendry BM, Fan TP, Zhao Z,
Duez P, Simmonds MS, Witt CM, Lu A, Robinson N, et al: The quest
for modernisation of traditional Chinese medicine. BMC Complement
Altern Med. 13:1322013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wojcikowski K, Johnson DW and Gobe G:
Herbs or natural substances as complementary therapies for chronic
kidney disease: ideas for future studies. J Lab Clin Med.
147:160–166. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Santos LS, Chin EW, Ioshii SO and Tambara
Filho R: Surgical reduction of the renal mass in rats: morphologic
and functional analysis on the remnant kidney. Acta Cir Bras.
21:252–257. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kren S and Hostetter TH: The course of the
remnant kidney model in mice. Kidney Int. 56:333–337. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
He L, Shen P, Fu Q, Li J, Dan M, Wang X
and Jia W: Nephro-protective effect of Kangqianling decoction on
chronic renal failure rats. J Ethnopharmacol. 122:367–373. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wen X, Wu J, Wang F, Liu B, Huang C and
Wei Y: Deconvoluting the role of reactive oxygen species and
autophagy in human diseases. Free Radic Biol Med. 65:402–410. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Dettmering T, Zahnreich S, Colindres-Rojas
M, Durante M, Taucher-Scholz G and Fournier C: Increased
effectiveness of carbon ions in the production of reactive oxygen
species in normal human fibroblasts. J Radiat Res. 56:67–76. 2015.
View Article : Google Scholar
|
|
69
|
Qin XJ, He W, Hai CX, Liang X and Liu R:
Protection of multiple antioxidants Chinese herbal medicine on the
oxidative stress induced by adriamycin chemotherapy. J Appl
Toxicol. 28:271–282. 2008. View Article : Google Scholar
|
|
70
|
Bongartz LG, Cramer MJ, Doevendans PA,
Joles JA and Braam B: The severe cardiorenal syndrome: 'Guyton
revisited'. Eur Heart J. 26:11–17. 2005. View Article : Google Scholar
|
|
71
|
Tu Y, Sun W, Wan YG, Gao K, Liu H, Yu BY,
Hu H and Huang YR: Dahuang Fuzi Decoction ameliorates tubular
epithelial apoptosis and renal damage via inhibiting TGF-β1-JNK
signaling pathway activation in vivo. J Ethnopharmacol.
156:115–124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wankun X, Wenzhen Y, Min Z, Weiyan Z, Huan
C, Wei D, Lvzhen H, Xu Y and Xiaoxin L: Protective effect of
paeoniflorin against oxidative stress in human retinal pigment
epithelium in vitro. Mol Vis. 17:3512–3522. 2011.
|
|
73
|
Cao Y, Zhang Y, Wang N and He L:
Antioxidant effect of imperatorin from Angelica dahurica in
hypertension via inhibiting NADPH oxidase activation and MAPK
pathway. J Am Soc Hypertens. 8:527–536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chakraborti S and Chakraborti T:
Oxidant-mediated activation of mitogen-activated protein kinases
and nuclear transcription factors in the cardiovascular system: a
brief overview. Cell Signal. 10:675–683. 1998. View Article : Google Scholar
|
|
75
|
Li ZJ, Li XM, Piao YJ, Choi DK, Kim SJ,
Kim JW, Sohn KC, Kim CD and Lee JH: Genkwadaphnin induces reactive
oxygen species (ROS)-mediated apoptosis of squamous cell carcinoma
(SCC) cells. Biochem Biophys Res Commun. 450:1115–1119. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhang W and Liu HT: MAPK signal pathways
in the regulation of cell proliferation in mammalian cells. Cell
Res. 12:9–18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Shimizu H, Bolati D, Higashiyama Y,
Nishijima F, Shimizu K and Niwa T: Indoxyl sulfate upregulates
renal expression of MCP-1 via production of ROS and activation of
NF-κB, p53, ERK, and JNK in proximal tubular cells. Life Sci.
90:525–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Sung CC, Hsu YC, Chen CC, Lin YF and Wu
CC: Oxidative stress and nucleic acid oxidation in patients with
chronic kidney disease. Oxid Med Cell Longev. 2013:3019822013.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Khan SR: Stress oxidative: nephrolithiasis
and chronic kidney diseases. Minerva Med. 104:23–30.
2013.PubMed/NCBI
|
|
80
|
Popolo A, Autore G, Pinto A and Marzocco
S: Oxidative stress in patients with cardiovascular disease and
chronic renal failure. Free Radic Res. 47:346–356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Huang C, Day ML, Poronnik P, Pollock CA
and Chen XM: Inhibition of KCa3.1 suppresses TGF-β1 induced MCP-1
expression in human proximal tubular cells through Smad3, p38 and
ERK1/2 signaling pathways. Int J Biochem Cell Biol. 47:1–10. 2014.
View Article : Google Scholar
|
|
82
|
An P, Dang HM, Shi XM, Ye BY and Wu XL:
'Qufeng Tongluo' acupuncture prevents the progression of
glomerulonephritis by decreasing renal sympathetic nerve activity.
J Ethnopharmacol. 155:277–284. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Alcorn JF, van der Velden J, Brown AL,
McElhinney B, Irvin CG and Janssen-Heininger YM: c-Jun N-terminal
kinase 1 is required for the development of pulmonary fibrosis. Am
J Respir Cell Mol Biol. 40:422–432. 2009. View Article : Google Scholar :
|