Introduction
Telomeres are DNA regions positioned at the tips of
chromosomes. They are composed of repetitive nucleotide sequences
(5'-TTAGGG-3'). These functional non-coding repeats protect the
ends of eukaryotic chromosomes from attrition and damage. Telomeres
shorten during mitotic cell division due to the inability of DNA
polymerase to fully replicate the end part of the lagging strand of
DNA. This is commonly referred to as the 'end-replication
problem'.
Telomere length (TL) gradually decreases, by
approximately 20-40 base pairs per year (1). The shortening of telomeres can be
reversed by the enzyme telomerase, which is active in
high-proliferating cells, such as male germ cells, activated
lymphocytes, stem cells and cancer cells (2,3).
In most adult human somatic cells though, the expression of
telomerase is deficient, and TL decreases with age (1). Telomerase activity has been
associated with pregnancy complications (4,5).
Furthermore, attention has been paid to telomere shortening in
various diseases and age-related diseases, such as cancer,
cardiovascular diseases (CAD), diabetes, rheumatoid arthritis and
psychiatric disorders (6-10). In addition, TL is a biomarker of
the replicative past and the replicative potential of somatic
cells. Previous studies have focused on the estimation of mean TL
(MTL) in age-related diseases (11,12); however, current concepts highlight
the pivotal role of critically short telomeres in cell homeostasis
(13). Vakonaki et al
previously demonstrated an association between TL and drug abuse,
which leads to premature biological aging (14). Recently, the creation of a
database named 'BIOTEL version 2.4', Telomere Length Database
Project (TLDP), was reported (15). This database is a functional
semi-automated worksheet that calculates a wide range of TL
statistics and is thus a useful tool with applications in research
on telomere biology, as well as in biological age estimation
(15). It is important to mention
that when the length of the telomeres shortens below a threshold
limit, cell growth is restricted, and cells undergo cellular
senescence or apoptosis (16).
Continuous telomere shortening in cells that fail to undergo
senescence, leads to critically short telomeres (<3 kb), prone
to chromosome-end fusion. Fused chromosomes break unequally in
mitosis, leading to genomic instability and chromosomal
rearrangements during cell divisions (17). Indeed, a short telomere load has
emerged as a more potent biomarker than MTL or the expression of
telomerase (18,19). Actually, recent studies have
indicated that a short TL is associated with several age-associated
diseases, including CAD, stroke, cancer, arthritis, osteoporosis,
cataracts, diabetes type2, hypertension, mental diseases, chronic
obstructive pulmonary disease and dementia (9,20-22). Goglin et al suggested that
individuals with shorter telomeres have a higher mortality rate, as
compared to those with longer telomeres (23). Human studies have shown that TL
can be inherited through the direct inheritance of the telomeres
and through the specific genetic factors found in the parental
gametes (24). Furthermore,
telomere shortening can be affected by environmental factors,
including physical activity, body mass index (BMI), hormone
replacement therapy, smoking, chronic inflammation, oxidative
stress, dietary antioxidants and importantly vitamin intake
(25-28).
Several studies have examined the correlation of
individual nutrients with TL. Indeed, B12 vitamin deficiency has
been associated with TL, and an altered DNA methylation (26). In addition, a lack of vitamin B12
and folate can lead to inadequate DNA methylation, genomic
instability and telomere shortening (29-31). In a previous study, the beneficial
effects of omega-3 (n-3) fatty acids on TL observed in patients
with chronic kidney disease, was attributed to reduced oxidative
stress (32). Furthermore, it has
been shown that omega-3 polyunsaturated fatty acids (PUFAs) can
reduce inflammation and decrease oxidative stress (33-35) and thus, protect telomeres from
attrition (36). Another study
suggested that a shorter TL is associated with a significantly
increased risk for breast cancer in pre-menopausal women and women
with a lower dietary and supplemental β-carotene vitamin C or E
intake (37). In vitro
experiments have demonstrated that telomere shortening is delayed
by ascorbic acid or its derivatives (38-40) and the lifespan of certain somatic
cells has been shown to be increased by α-tocophenol (41). To date, TL has been associated
with single vitamin or micronutrient intake, while little is known
about the effects of their combination on telomere status. A recent
study by Tsoukalas et al indicated that the synergistic
effects of nutrients and natural compounds can activate telomerase
enzyme in vitro, in human peripheral blood mononuclear
cells; human clinical studies with these formulations are required
in order to reveal the health benefits of telomerase activation
through natural molecules (42).
Epidemiological studies have demonstrated that the deficiency of
multiple concurrent micronutrients is very common globally and is
associated with chronic diseases; these studies stress the need to
restore these deficiencies through diet and nutraceutical
supplements (NS) (43,44).
The aim of the present study was to analyze the TL
profile of a population using a combination of NS and to evaluate
the putative effects of supplements on TL, in comparison to a
healthy population not receiving any supplements.
Materials and methods
Materials and reagents
One capsule of My Health (Lumis Research SA)
contains the following: Mix Vitamin (ascorbic acid, vitamin E
acetate 50% natural, niacin, vitamin B1, vitamin K2 Mena Q7 0.2%,
vitamin B6, β-carotene, vitamin B12 1%), anti-caking agents
(microcrystalline cellulose, mono- and diglycerides of fatty acids,
magnesium stearate and silica dioxide. One drop of vitamin D (Pure
D3, Natural Doctor) contains 2,000 IU vitamin D. One capsule of
Complete D3&K2 Cofactors (Natural Doctor) contains 2,000 IU
vitamin D3, 100 µg K2 and 56 mg magnesium. Two capsules of
Omega 3-6-9 (Lumis Research SA) contain the following: 740 mg fish
oil, 740 mg linseed oil, 740 mg borage oil (Borrgo
officinalis L.), 200 mg Krill oil, 370 mg alpha linolenic acid,
312,6 mg eicosapentaenoic acid (EPA), 154,2 mg docosa-hexaenoic
acid (DHA), 162,8 mg gamma linolenic acid (GLA), 400 mg linoleic
acid and 273,8 mg oleic acid. Four capsules of My Antioxidant
(Lumis Research SA) contain the following: Mix Vitamin (ascorbic
acid, calcium ascorbate, ascorbyl palmitate, natural vitamin E
acetate 50%, vitamin B3, vitamin B12 1%, β-carotene, vitamin B5,
vitamin B6, vitamin B2, vitamin B1, folic acid, vitamin D3 biotin),
One capsule of My Probiotics (Lumis Research SA) contains the
following: Lyophilized kefir grains, Lactobacillus casei
(LMG-S27763), Lactobscillus reuteri (LMG S-27759),
Bifidobacterium bifidum (B. bifidum; LMG S-27761),
Lactobacillus acidophilus (LMG S-27762), inulin, acacia
fiber and hydroxypropyl cellulose. One scoop of My Gastro (Lumis
Research SA) contains the following: 2,500 mg L-glutamine
(Kyowa®) and 500 mg oligo fructose (FOS). One capsule of
My Ubiquinol (Q10) (Lumis Research SA) contains 50 mg of
ubiquinol.
Study design
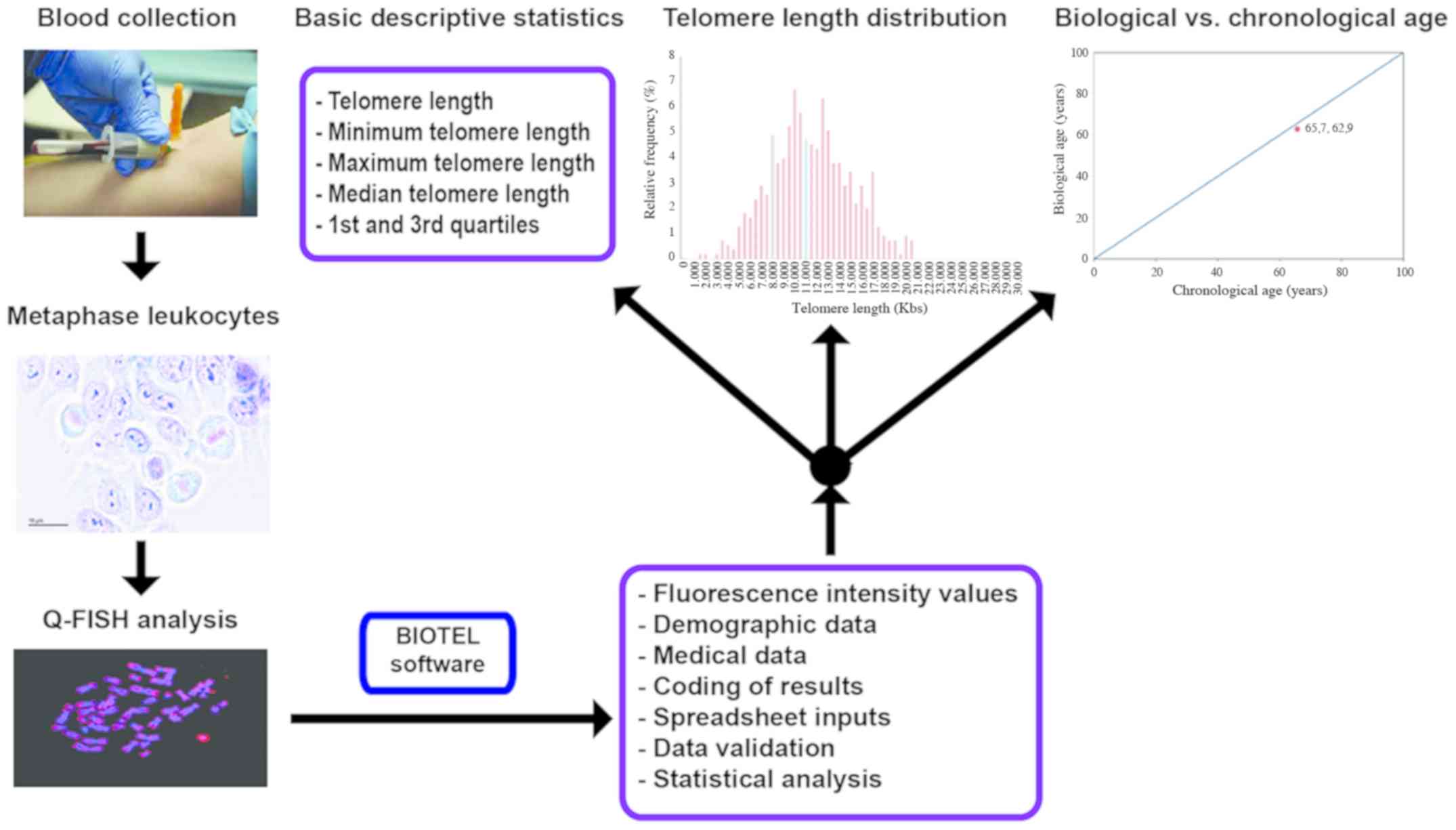
The process of TL estimation is depicted in Fig. 1. The participants were selected
from individuals visiting a private clinic in Athens for routine
checkup, as well as from personnel of the University Hospital of
Heraklion. Basic demographic data and a medical history of chronic
diseases were obtained. Demographic data included the name,
surname, date of birth and sex. Medical history included all the
self-reported statements for chronic diseases, such as diabetes,
cancer, rheumatoid arthritis, etc., possible exposure to pesticides
(occupational) and use of antidepressant medication. Individuals
that reported having a medical history of any chronic disease were
excluded. Based on these criteria, 47 individuals were selected for
the study. They had never taken any supplements prior to the study
and they were in the 40-55 age group. The participants were healthy
and were not on a restricted diet. They were divided into 2 groups
as follows: An experimental group that consisted of 16 individuals
supplemented with NS and the control group with 31 individuals that
did not receive any supplements. The daily dose of all supplements
was assigned according to the suggestions of the manufacturers.
Blood samples were collected from all individuals and citrate was
used as an anticoagulant. The supplementation period for all
participants lasted for 6 to 12 months. The protocol of this study
was approved by the Ethics Committee for Patients and Biological
Material of the University of Crete with reference no.
63/22.03.2019. Biological Material and information of patients were
obtained with written informed consent according to the EU General
Data Protection Regulation (GDPR). All procedures performed in
studies involving human participants were under the ethical
standards with the 1964 Helsinki declaration and its later
amendments, or comparable ethical standards.
Quantitative-fluorescent in situ
hybridization (Q-FISH) analysis
Peripheral blood samples (2.5 ml) were collected
from all participants. Heparinized blood was cultured in RPMI-1640
culture medium supplemented with 10% fetal bovine serum, 1%
L-glutamine, 1% penicillin and streptomycin (all from
Sigma-Aldrich) and stimulated for 72 h in a CO2
incubator with phytohemagglutinin. For metaphase chromosome
preparations, 10 µg/ml colcemid was added to each culture 2
h before harvesting, followed by KCl hypotonic shock and
methanol/acetic (3:1) fixation. Q-FISH analysis on metaphases was
performed as previously described (14,15).
Images were captured for each slide, using a 63X
objective and a charge-coupled device camera at 1,024×1,024 pixels
resolution and 8-bit depth. For each slide, >10 different
scanned images were obtained from 3 independent biological
replicates. Maximum projections and deconvolution of the images
were performed using Leica Q-FISH software [Leica Application
Suite-Advanced Fluorescence (LAS-AF) version 3.1.3 for Leica TCS
SP8] and telomere fluorescence intensity was analysed using ImageJ
(https://imagej.nih.gov/ij/) by two
independent investigators. Telomere fluorescence values were
converted into kb by using L5178Y-S cells (cat. no. 93050408;
Culture Collections, Public Health England) with a stable and known
TL, which was estimated to be approximately 7 kb (45).
Statistical analysis
Percentiles of TL (of the whole telomere genome) and
the TL of short telomeres (TLS) (TL <20th percentile) were
estimated. The examination of TL and TLS normality was assessed
using Kolmogorov-Smirnov's test with Liliefors correction.
Differences in proportions were examined using Pearson's Chi-square
test or Fisher's exact test when assumptions were not reached. Age
comparison was tested using an independent samples t-test. Multiple
linear regressions were applied for examining the effect of NS on
TL and TLS length following adjustment for age and sex. Box and
Whisker plot and bar-charts were used for data graphical
representation. IBM SPSS Statistics 23.0 software was used for data
analysis and a level of acceptance of null hypothesis was set at
P=0.05.
Results
The demographic characteristics of the participants
in the control group (CG) and NS group (NSG) are presented in
Table I. The sex distribution
between the 2 groups did not differ significantly (P=0.609), with a
similar proportion of males in both the NSG (n=9, 37.5%) and the CG
(n=15, 62.5%). Age distribution, after dividing the population to
5-year interval groups, revealed that the 2 groups were age-matched
(P=0.505). To elucidate whether demographics had any effect on TL,
we performed statistical analysis in both groups with respect to
sex. A separate analysis was performed in the respective age
groups. As shown in Table II,
females had a longer TL compared to males with a difference of 248
to 474 bases in TL and a difference of 195 to 200 bases in TLS.
These differences however, did not reach statistical significance,
as none of the quartiles in whole or in short telomeres exhibited a
statistically significant difference between the 2 sexes
(P>0.300).
 | Table IDemographic characteristics of the
participants in the nutraceutical supplements group (NSG) and
control group (CG). |
Table I
Demographic characteristics of the
participants in the nutraceutical supplements group (NSG) and
control group (CG).
| NSG (n=16) | CG (n=31) | Total | P-value |
|---|
| Sex (n, %)a | | | | | | |
| Male | 9 | 37.5% | 15 | 62.5% | 24 | 0.609 |
| Female | 7 | 30.4% | 16 | 69.6% | 23 | |
| Age, years (means ±
SD)b | 46.4 | 3.5 | 47.6 | 4.0 | 47.1 (3.8) | 0.323 |
| Age groups, (n,
%)c | | | | | | |
| 40-45 | 6 | 42.9% | 8 | 57.1% | 14 | 0.505 |
| 45-50 | 8 | 34.8% | 15 | 65.2% | 23 | |
| 50-55 | 2 | 20.0% | 8 | 80.0% | 10 | |
| Table IIComparison of whole telomere genome
(TL) and short telomere length (TLS) (<20th percentile) (in
bases) between the sexes. |
Table II
Comparison of whole telomere genome
(TL) and short telomere length (TLS) (<20th percentile) (in
bases) between the sexes.
| Sex
| P-value |
|---|
Male (n=24)
| Female (n=23)
|
|---|
| Mean | SD | Mean | SD |
|---|
| TLa | | | | | |
| 1st Quartile | 8,444 | 1,296 | 8,692 | 1,374 | 0,528 |
| Median | 10,142 | 1,315 | 10,490 | 1,537 | 0,407 |
| 3rd Quartile | 12,107 | 1,505 | 12,582 | 1,910 | 0,348 |
| TLSa | | | | | |
| 1st Quartile | 6,321 | 1,288 | 6,516 | 1,079 | 0,577 |
| Median | 7,052 | 1,270 | 7,247 | 1,173 | 0,587 |
| 3rd Quartile | 7,592 | 1,283 | 7,792 | 1,293 | 0,598 |
TL (of the whole telomere genome) and TLS (<20th
percentile) was estimated in the participants of both groups.
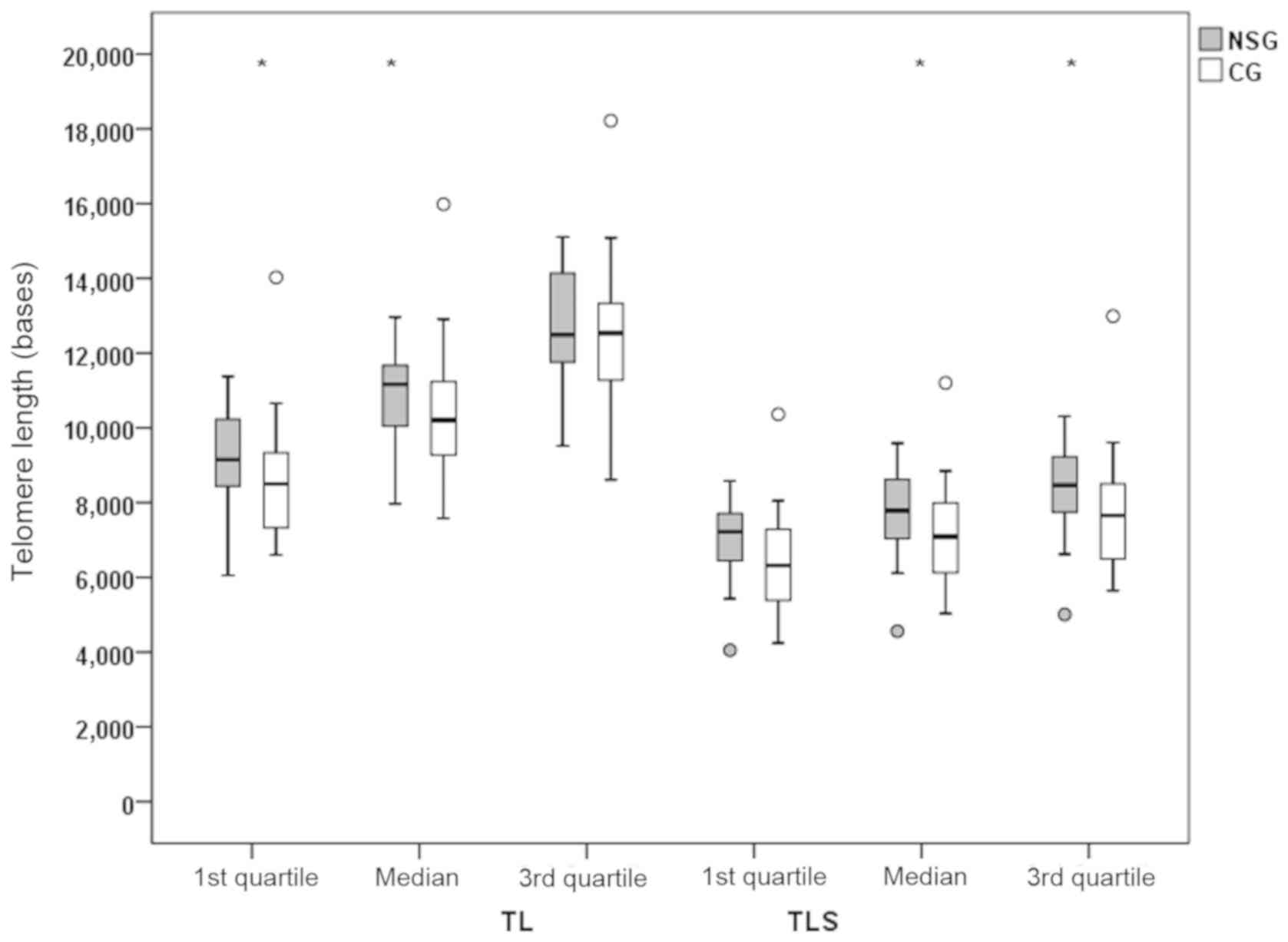
Differences in TL and TLS between the NSG and CG are shown in
Figs. 2 and 3. Quartiles of TL and TLS were estimated
from raw data for each patient separately. There was a
statistically significant difference (P<0.05) in the TL values
of the NSG group, compared to the CG, with increased mean quartiles
values for the 1st and 2nd quartile (median). TL measurements were
844 and 953 bases higher in the NSG group compared to the CG.
Accordingly, there were statistically significant increased values
of TLS in the 2nd (median) and 3rd quartile with values of P=0.044
and P=0.033, respectively. A tendency for difference was found for
the 1st quartile comparisons (P=0.051).
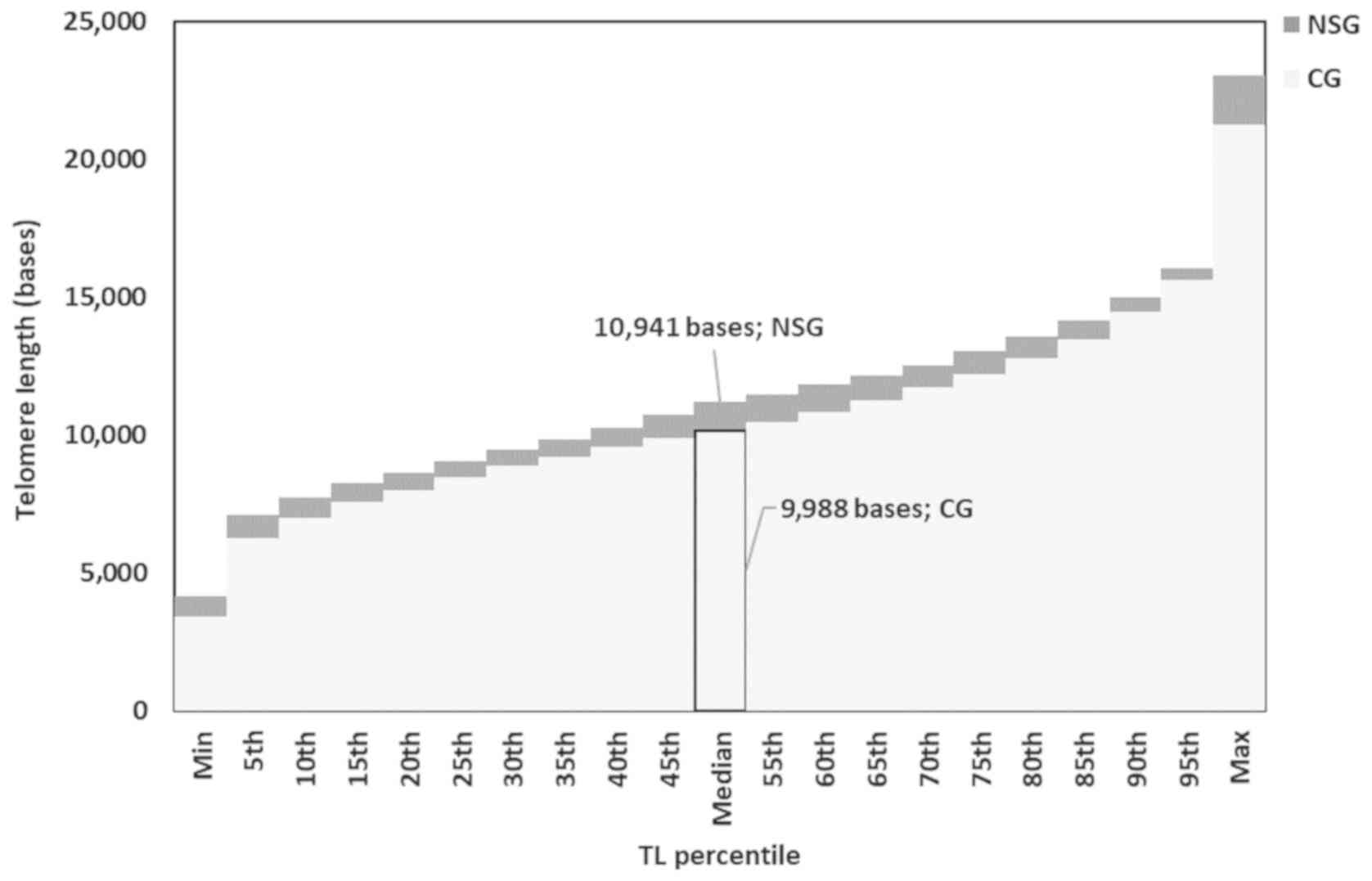
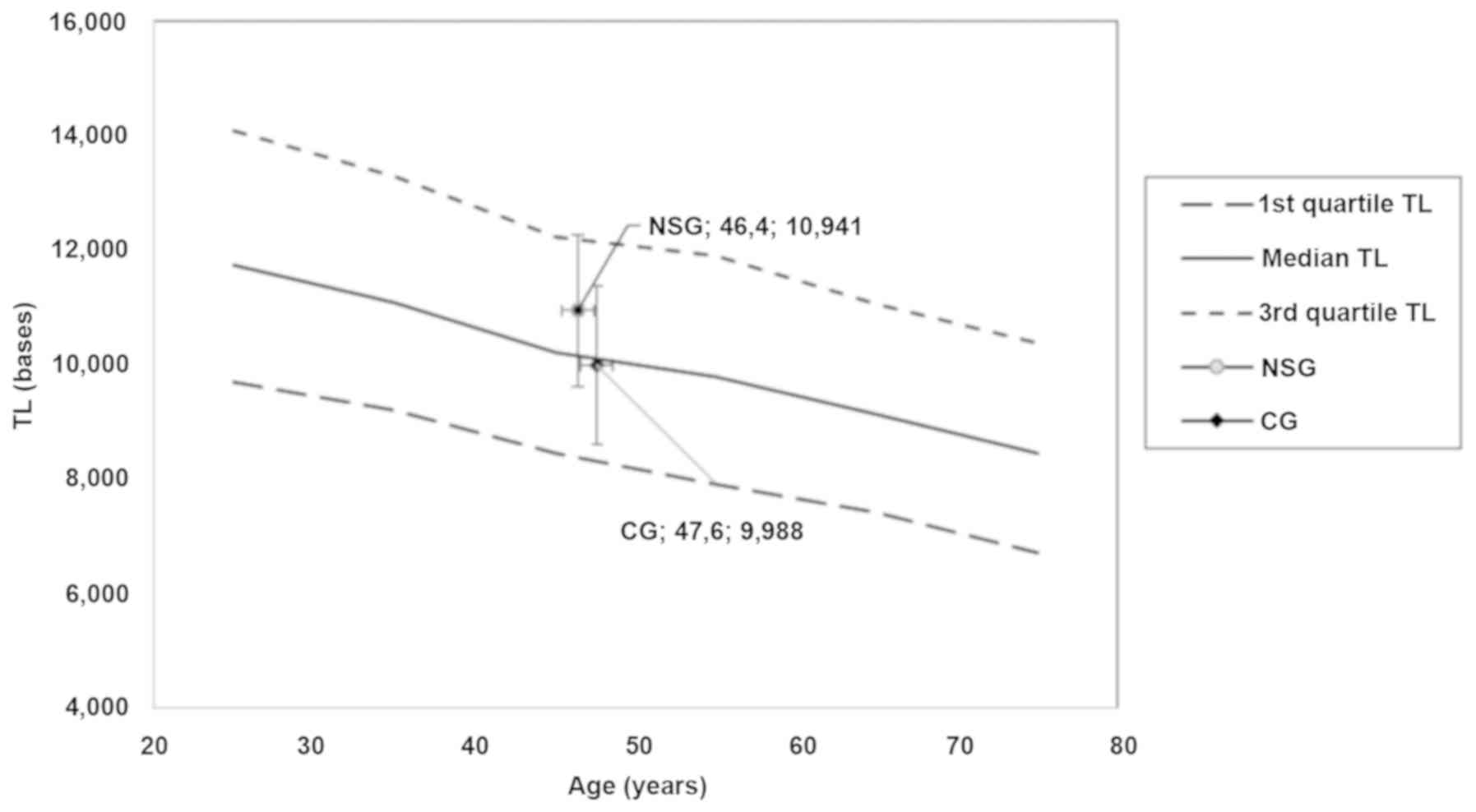
The position of NSG and CG in the nomograms produced
from TLDP data (13) is shown in
Fig. 4. Lines of 1st, 2nd
(median) and 3rd quartile of TL vs. age were presented, showing the
age decrement. Mean age with 2SD of age and TL with 2SD of both
groups were placed onto the diagram. It is clear that the mean TL
levels of the NSG are over the line of the median of a healthy
population, while the mean TL levels of the CG are under the line
of the median of a healthy population. A difference of
approximately 1,000 bases was observed.
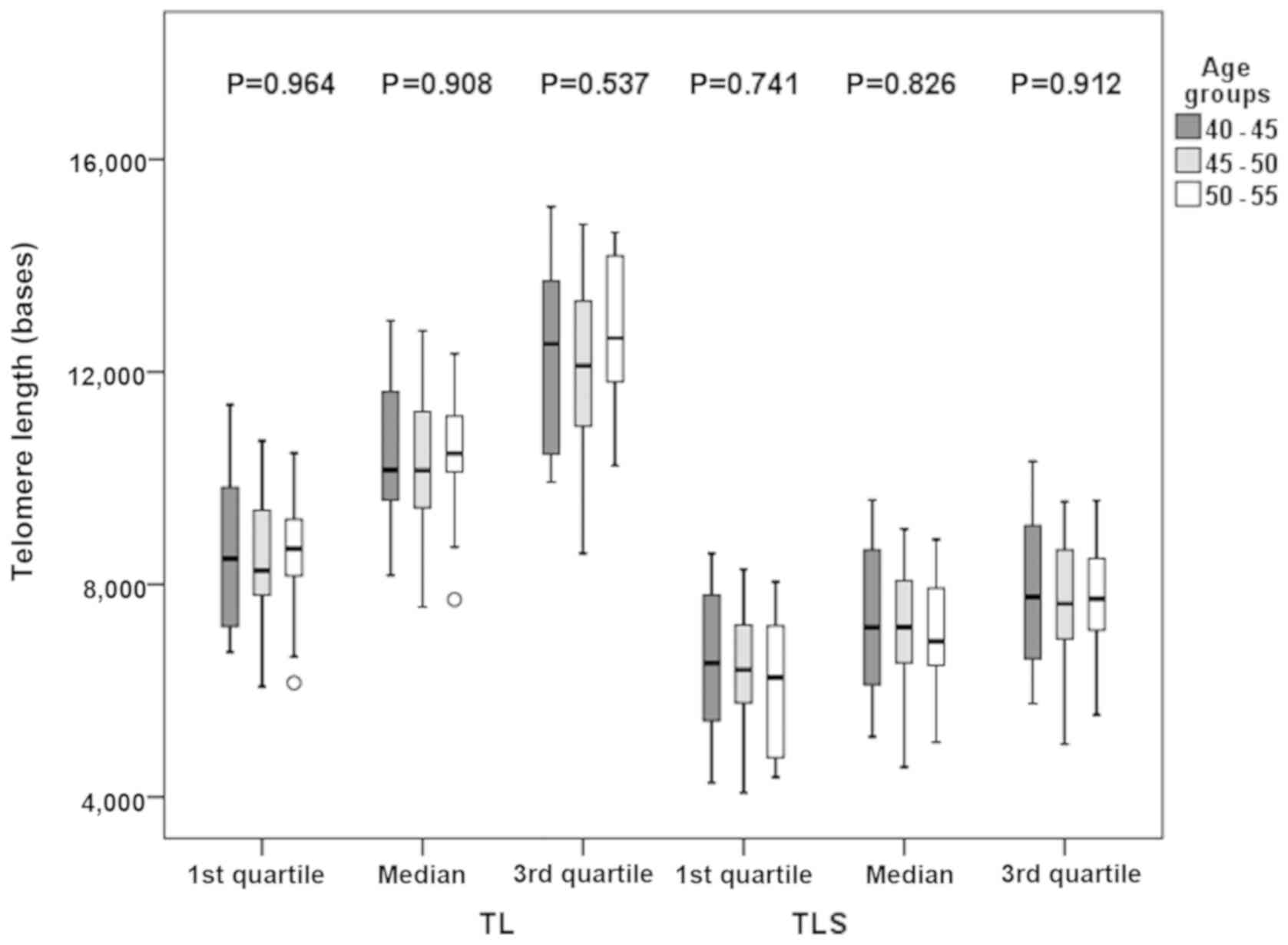
Furthermore, as demonstrated in Fig. 5, and in more details in Table SI, there were no significant
differences in TL among the age groups examined (P>0.500).
P-values of TL vs. age were 0.964 for the 1st quartile, 0.908 for
the median and 0.537 for the 3rd quartile, while P-values for TLS
vs. age were 0.741 for the 1st quartile, 0.826 for the median and
0.912 for the 3rd quartile.
Multiple linear regression models were applied for
mean TL (or TLS) and their quartiles were set as dependent
variables. Explanatory variables were NSG, age and sex of
participants. The results of each regression model are presented in
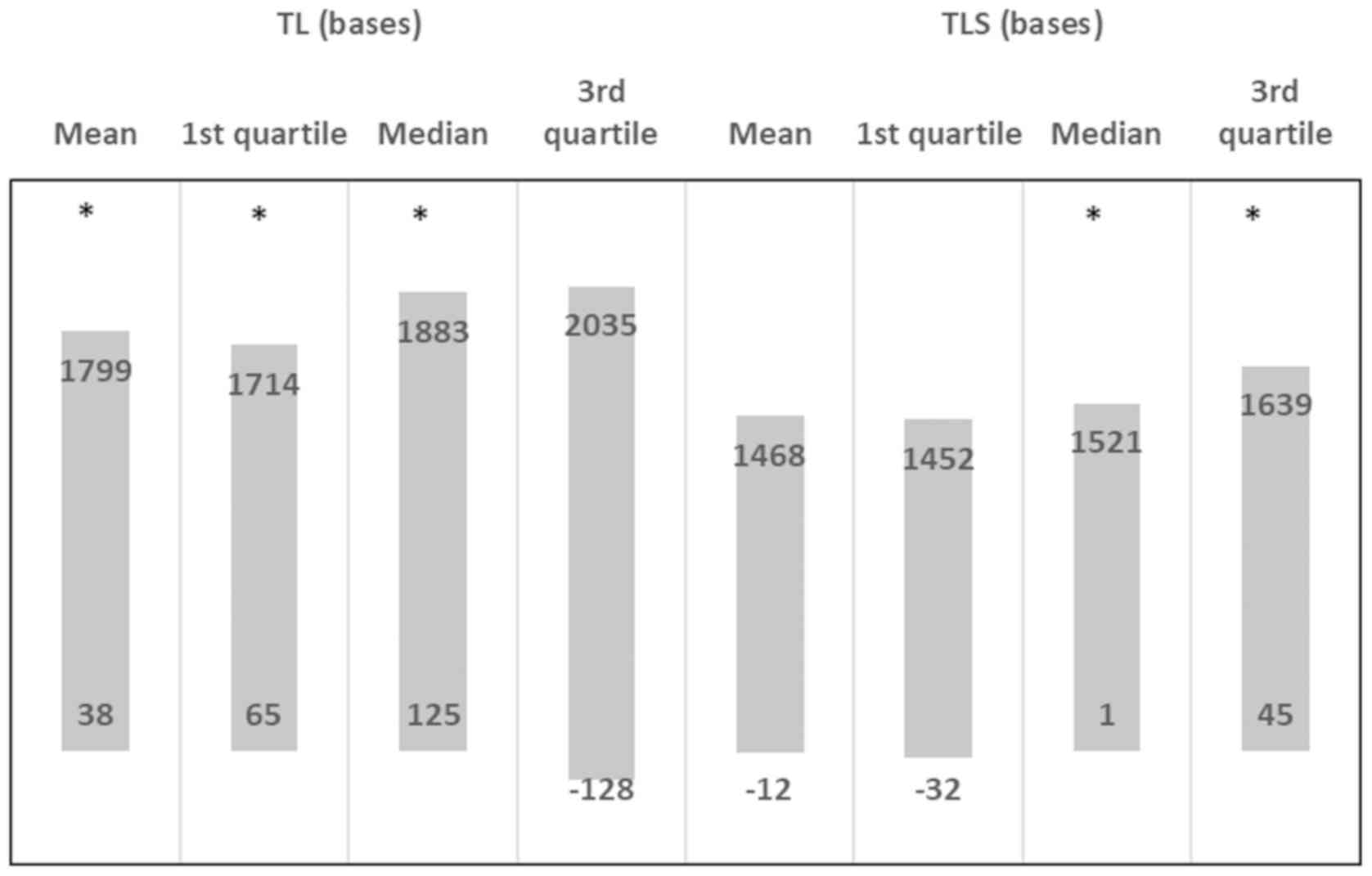
Fig. 6. There was a positive
effect for mean TL (beta: 918; 95% CI: 38-1799), for the 1st
quartile TL (beta: 889; 95% CI: 65-1714), for the median TL (beta:
1004; 95% CI: 125-1883), but not for the 3rd quartile TL (beta:
953; 95% CI: -128-2035) in the NSG. There was also a positive
effect on the median TLS (beta: 0.728; 95% CI: 1-1521) and 3rd
quartile TLS (beta: 842; 95% CI: 45-1639), and tendencies for an
effect were observed in the mean TLS (P=0.051) and in 1st quartile
TLS (P=0.060). Furthermore, Fig.
6 shows the effect (beta coefficients) as derived from multiple
linear regression, following adjustment for age and sex (beta and
95% CI).
Discussion
The results of the present study suggest that the
daily administration of NS favorably affects the length of whole
and short telomeres, independent of sex and age. Although the
underlying mechanisms of the effects of NS remain unclear, their
effects on several key mechanisms, including oxidative stress and
chronic inflammation, could be linked to TL maintenance (46). Indeed, studies have demonstrated
that increased oxidative stress and inflammation are associated
with a reduced leukocyte TL in patients with periodontitis
(47), as well as in patients
with Parkinson's disease (48).
Furthermore, Ma et al reported that older individuals with
central obesity, hyperglycemia, insulin resistance and lower
antioxidant levels had a shorter leukocyte TL; specifically, the
authors demonstrated that a decreased TL correlated with increased
oxidative stress in patients with type 1 and type 2 diabetes
(49). Furthermore, oxidative
stress has been linked to TL shortening during aging, in a
comparative study that concluded that elderly males living in
Greece have lower indices of oxidative stress and higher
antioxidant levels, compared to a corresponding population of
elderly Dutch males (50). The
authors suggested that the length of telomeres in leukocytes was
significantly greater in the Greek compared to the Dutch
population, as the latter live in a more stressful urban setting
and follow a diet poor in antioxidant nutrients. Indeed,
antioxidants and anti-inflammatory agents have been shown to reduce
the pace of TL shortening (51).
In a previous study, Paul associated the availability of vitamin B
and D with leukocyte (LTL) (52).
Furthermore, in that study, serum folate and its metabolites were
positively associated with LTL. It has been suggested that the
mechanisms through which these nutritional factors attenuate
telomere attrition are antioxidant activity, DNA methylation and
the prevention of DNA damage (53). In addition, it has been indicated
that carotenoids, omega-3-fatty acid, coenzyme Q10, selenium and
vitamin D can reduce the rate of telomeric shortening (51). In 2013, Kiecolt-Glaser et
al demonstrated that men and women aged 40-85 years who had
been taking supplementation, including omega-3 fatty acids, had
reduced oxidative stress and an increased TL (54).
In this study, we present findings from the
evaluation of TL in participants receiving a combination of vitamin
supplements. An advantage of the present study is the methodology
of TL measurement. We measured the TL of peripheral blood
leukocytes using a method of high sensitivity (Q-FISH) to produce
data on TL and TLS (<20th percentile) percentages using BIOTEL
(15). We determined a
statistically significant increase in TLS and TL by NS. This is a
very interesting finding, as there is evidence indicating that a
short telomere load is more critical to cell homeostasis than MTL,
or the expression of telomerase (18,17). The increase of short telomeres,
measured as a percentage, predicts longevity in mammals and is
reversely associated with the majority of age-related diseases.
Although the majority of measurements of TL in leukocyte
subpopulations have been performed by flow-FISH or real-time PCR,
which gives the average length of short telomeres, Q-FISH provides
results on median length of single telomeres, the percentage of
short telomeres and provides an overview of the rate of telomere
shortening and longevity (13).
In this study, we performed an analysis of the TL
and TLS between the 2 sexes and did not find a statistically
significant difference between males and females. This finding may
be attributed to the relatively small size of the group. A previous
study by Herrmann et al suggested that telomeres are longer
in women due to higher estrogen levels, which increase telomerase
activity and present antioxidant effects (53).
The limitations of this study include the small
number of participants, as well as the fact that not all lifestyle
parameteres that can affect TL and telomerase activity were
determined. Taking this limitation into account, the effects of NS
on TL presented herein highlight the need for the further
evaluation of the underlying mechanisms.
In conclusion, in this study, we demonstrate that NS
may affect the length of short telomeres. Short telomeres are
critical to cell homeostasis and the only reliable method for the
evaluation of short telomere distribution, to date, is Q-FISH.
Therefore, this study provides evidence that the antioxidant
effects of NS is implicated in TL maintainance. Future research is
required to focus on the mechanisms through which NS affect
telomere shortening.
Supplementary Materials
Funding
This study was funded by Metabolomic Medicine S.A.
and Toxplus S.A. and supported by the Special Research Account of
University of Crete (ELKE nos. 4602, 4920 and 3963).
Availability of data and materials
The datasets presented in this study are available
from the corresponding author upon reasonable request.
Authors' contributions
DT, PF, AT, DAS and DC conceived and designed the
study and wrote the manuscript. PF, EV and AOD performed the data
processing and quality control assessment. EvS, AKA and DN
performed the statistical analysis and data interpretation. LK
interpreted the data and critically revised the article. EK and ErS
performed the imaging analysis. All authors reviewed and approved
the manuscript prior to submission and have approved the final
manuscript.
Ethics approval and consent to
participate
The protocol of this study was approved by the
Ethics Committee for Patients and Biological Material of the
University of Crete with reference no. 63/22.03.2019. Biological
Material and information of patients were obtained with written
informed consent according to the EU General Data Protection
Regulation (GDPR). All procedures performed in studies involving
human participants were under the ethical standards with the 1964
Helsinki declaration and its later amendments, or comparable
ethical standards.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
DT is a scientific advisor for Lumis Research SA and Natural Doctor
SA. The other authors declare no competing interests. To avoid any
bias in the collection of the experimental data, the experiments
were conducted by the Laboratory of Toxicology of the Medical
School of the University of Crete. Lumis Research SA and Natural
Doctor SA had no involvement in the preparation of the manuscript,
the results and the supervision of the study.
Abbreviations:
|
MTL
|
mean telomere length
|
|
CAD
|
cardiovascular disease
|
|
BMI
|
body mass index
|
|
TL
|
telomere length (of the whole
genome)
|
|
TLS
|
telomere length of short telomeres
(20th percentile)
|
|
NS
|
nutraceutical supplements
|
|
TLDP
|
Telomere Length Database Project
|
Acknowledgments
The study is part of the special part of PhD thesis
from University of Medicine and Pharmacy and Craiova. The authors
would like to thank all the administrative, technical and medical
staff of Toxplus, the Metabolomic Medicine health clinic, and the
Laboratory of Toxicology for their dedicated involvement in this
study.
References
|
1
|
Starkweather AR, Alhaeeri AA, Montpetit A,
Brumelle J, Filler K, Montpetit M, Mohanraj L, Lyon DE and
Jackson-Cook CK: An integrative review of factors associated with
telomere length and implications for biobehavioral research. Nurs
Res. 63. pp. 36–50. 2014, View Article : Google Scholar
|
|
2
|
Blackburn EH, Chan S, Chang J, Fulton TB,
Krauskopf A, McEachern M, Prescott J, Roy J, Smith C and Wang H:
Molecular manifestations and molecular determinants of telomere
capping. Cold Spring Harb Symp Quant Biol. 65:253–263. 2000.
View Article : Google Scholar
|
|
3
|
Beyne-Rauzy O, Prade-Houdellier N, Demur
C, Recher C, Ayel J, Laurent G and Mansat-De Mas V: Tumor necrosis
factor- alpha inhibits hTERT gene expression in human myeloid
normal and leukemic cells. Blood. 106:3200–3205. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Westin ER, Aykin-Burns N, Buckingham EM,
Spitz DR, Goldman FD and Klingelhutz AJ: The p53/p21(WAF/CIP)
pathway mediates oxidative stress and senescence in dyskeratosis
congenita cells with telomerase insufficiency. Antioxid Redox
Signal. 14:985–997. 2011. View Article : Google Scholar :
|
|
5
|
Fragkiadaki P, Tsoukalas D, Fragkiadoulaki
I, Psycharakis C, Nikitovic D, Spandidos DA and Tsatsakis AM:
Telomerase activity in pregnancy complications (Review). Mol Med
Rep. 14:16–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fujii H, Shao L, Colmegna I, Goronzy JJ
and Weyand CM: Telomerase insufficiency in rheumatoid arthritis.
Proc Natl Acad Sci USA. 106:4360–4365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vakonaki E, Tsiminikaki K, Plaitis S,
Fragkiadaki P, Tsoukalas D, Katsikantami I, Vaki G, Tzatzarakis MN,
Spandidos DA and Tsatsakis AM: Common mental disorders and
association with telomere length. Biomed Rep. 8:111–116.
2018.PubMed/NCBI
|
|
8
|
Wu X, Amos CI, Zhu Y, Zhao H, Grossman BH,
Shay JW, Luo S, Hong WK and Spitz MR: Telomere dysfunction: A
potential cancer predisposition factor. J Natl Cancer Inst. 95. pp.
1211–1218. 2003, View Article : Google Scholar
|
|
9
|
Willeit P, Raschenberger J, Heydon EE,
Tsimikas S, Haun M, Mayr A, Weger S, Witztum JL, Butterworth AS,
Willeit J, et al: Leucocyte telomere length and risk of type 2
diabetes mellitus: New prospective cohort study and
literature-based meta-analysis. PLoS One. 9:e112483. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Calado RT and Young NS: Telomere diseases.
N Engl J Med. 361:2353–2365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Turner KJ, Vasu V and Griffin DK: Telomere
biology and human phenotype. Cells. 8:E732019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sanders JL and Newman AB: Telomere length
in epidemiology: A biomarker of aging, age-related disease, both,
or neither? Epidemiol Rev. 35:112–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vera E, Bernardes de Jesus B, Foronda M,
Flores JM and Blasco MA: The rate of increase of short telomeres
predicts longevity in mammals. Cell Rep. 2:732–737. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vakonaki E, Tzatzarakis M, Tsiminikaki K,
Nathena D, Fragkiadaki P, Kalliantasi K, Kanaki K, Vaki G, Plaitis
S, Tsoukalas D, et al: Effect of chronic and heavy drug abuse on
biological aging. World Acad Sci. 1:67–73. 2019.
|
|
15
|
Tsatsakis A, Tsoukalas D, Fragkiadaki P,
Vakonaki E, Tzatzarakis M, Sarandi E, Nikitovic D, Tsilimidos G and
Alegakis AK: Developing BIOTEL: A semi-automated spreadsheet for
estimating telomere length and biological age. Front Genet.
10:842019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shay JW and Wright WE: Hallmarks of
telomeres in ageing research. J Pathol. 211:114–123. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Muraki K, Nyhan K, Han L and Murnane JP:
Mechanisms of telomere loss and their consequences for chromosome
instability. Front Oncol. 2:1352012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hemann MT, Strong MA, Hao LY and Greider
CW: The shortest telomere, not average telomere length, is critical
for cell viability and chromosome stability. Cell. 107:67–77. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hao LY, Armanios M, Strong MA, Karim B,
Feldser DM, Huso D and Greider CW: Short telomeres, even in the
presence of telomerase, limit tissue renewal capacity. Cell.
123:1121–1131. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Baltzis D, Meimeti E, Grammatikopoulou MG,
Roustit M, Mavrogonatou E, Kletsas D, Efraimidou S, Manes C,
Nikolouzakis TK, Tsiaoussis J, et al: Assessment of telomerase
activity in leukocytes of type 2 diabetes mellitus patients having
or not foot ulcer: Possible correlation with other clinical
parameters. Exp Ther Med. 15:3420–3424. 2018.PubMed/NCBI
|
|
21
|
Nikolouzakis TK, Vassilopoulou L,
Fragkiadaki P, Mariolis Sapsakos T, Papadakis GZ, Spandidos DA,
Tsatsakis AM and Tsiaoussis J: Improving diagnosis, prognosis and
prediction by using biomarkers in CRC patients (Review). Oncol Rep.
39:2455–2472. 2018.PubMed/NCBI
|
|
22
|
Haycock PC, Heydon EE, Kaptoge S,
Butterworth AS, Thompson A and Willeit P: Leucocyte telomere length
and risk of cardiovascular disease: Systematic review and
meta-analysis. BMJ. 349. pp. g42272014, View Article : Google Scholar
|
|
23
|
Goglin SE, Farzaneh-Far R, Epel ES, Lin J,
Blackburn EH and Whooley MA: Change in leukocyte telomere length
predicts mortality in patients with stable coronary heart; disease
from the heart and soul study. PLoS One. 11:e01607482016.
View Article : Google Scholar
|
|
24
|
De Meyer T, Vandepitte K, Denil S, De
Buyzere ML, Rietzschel ER and Bekaert S: A non-genetic,
epigenetic-like mechanism of telomere length inheritance? Eur J Hum
Genet. 22:10–11. 2014. View Article : Google Scholar
|
|
25
|
Armanios M: Telomeres and age-related
disease: How telomere biology informs clinical paradigms. J Clin
Invest. 123:996–1002. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pusceddu I, Herrmann M, Kirsch SH, Werner
C, Hübner U, Bodis M, Laufs U, Wagenpfeil S, Geisel J and Herrmann
W: Prospective study of telomere length and LINE-1 methylation in
peripheral blood cells: The role of B vitamins supplementation. Eur
J Nutr. 55:1863–1873. 2016. View Article : Google Scholar
|
|
27
|
Pusceddu I, Herrmann M, Kirsch SH, Werner
C, Hübner U, Bodis M, Laufs U, Widmann T, Wagenpfeil S, Geisel J,
et al: One-carbon metabolites and telomere length in a prospective
and randomized study of B- and/or D-vitamin supplementation. Eur J
Nutr. 56:1887–1898. 2017. View Article : Google Scholar
|
|
28
|
Richards JB, Valdes AM, Gardner JP,
Paximadas D, Kimura M, Nessa A, Lu X, Surdulescu GL, Swaminathan R,
Spector TD, et al: Higher serum vitamin D concentrations are
associated with longer leukocyte telomere length in women. Am J
Clin Nutr. 86:1420–1425. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moores CJ, Fenech M and O'Callaghan NJ:
Telomere dynamics: The influence of folate and DNA methylation. Ann
NY Acad Sci. 1229:76–88. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Holick MF: Vitamin D deficiency. N Engl J
Med. 357:266–281. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nazki FH, Sameer AS and Ganaie BA: Folate:
Metabolism, genes, polymorphisms and the associated diseases. Gene.
533:11–20. 2014. View Article : Google Scholar
|
|
32
|
Barden A, O'Callaghan N, Burke V, Mas E,
Beilin LJ, Fenech M, Irish AB, Watts GF, Puddey IB, Huang RC, et
al: n-3 fatty acid supplementation and leukocyte telomere length in
patients with chronic kidney disease. Nutrients. 8:1752016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Calder PC: Polyunsaturated fatty acids and
inflammation. Biochem Soc Trans. 33:423–427. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kiecolt-Glaser JK, Belury MA, Andridge R,
Malarkey WB and Glaser R: Omega- 3 supplementation lowers
inflammation and anxiety in medical students: A randomized
controlled trial. Brain Behav Immun. 25:1725–1734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nälsén C, Vessby B, Berglund L, Uusitupa
M, Hermansen K, Riccardi G, Rivellese A, Storlien L, Erkkilä A,
Ylä-Herttuala S, et al: Dietary (n-3) fatty acids reduce plasma
F2-isoprostanes but not prostaglandin F2alpha in healthy humans. J
Nutr. 136:1222–1228. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang J, Rane G, Dai X, Shanmugam MK,
Arfuso F, Samy RP, Lai MK, Kappei D, Kumar AP and Sethi G: Ageing
and the telomere connection: An intimate relationship with
inflammation. Ageing Res Rev. 25:55–69. 2016. View Article : Google Scholar
|
|
37
|
Shen J, Gammon MD, Terry MB, Wang Q,
Bradshaw P, Teitelbaum SL, Neugut AI and Santella RM: Telomere
length, oxidative damage, antioxidants and breast cancer risk. Int
J Cancer. 124:1637–1643. 2009. View Article : Google Scholar :
|
|
38
|
Furumoto K, Inoue E, Nagao N, Hiyama E and
Miwa N: Age-dependent telomere shortening is slowed down by
enrichment of intracellular vitamin C via suppression of oxidative
stress. Life Sci. 63:935–948. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yokoo S, Furumoto K, Hiyama E and Miwa N:
Slow- down of age-dependent telomere shortening is executed in
human skin keratinocytes by hormesis-like-effects of trace hydrogen
peroxide or by anti-oxidative effects of pro-vitamin C in common
concurrently with reduction of intracellular oxidative stress. J
Cell Biochem. 93:588–597. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kashino G, Kodama S, Nakayama Y, Suzuki K,
Fukase K, Goto M and Watanabe M: Relief of oxidative stress by
ascorbic acid delays cellular senescence of normal human and Werner
syndrome fibroblast cells. Free Radic Biol Med. 35:438–443. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tanaka Y, Moritoh Y and Miwa N: Age-
dependent telomere- shortening is repressed by phosphorylated
alpha-tocopherol together with cellular longevity and intracellular
oxidative-stress reduction in human brain microvascular
endotheliocytes. J Cell Biochem. 102:689–703. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tsoukalas D, Fragkiadaki P, Docea AO,
Alegakis AK, Sarandi E, Spandidos DA, Tsatsakis A, Razgonova MP and
Calina D: Discovering novel telomerase activators. Mol Med Rep. In
press.
|
|
43
|
Bailey RL, West KP Jr and Black RE: The
epidemiology of global micronutrient deficiencies. Ann Nutr Metab.
66(Suppl 2): 22–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fairfield KM and Fletcher RH: Vitamins for
chronic disease prevention in adults: Scientific review. JAMA.
287:3116–3126. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
McIlrath J, Bouffler SD, Samper E,
Cuthbert A, Wojcik A, Szumiel I, Bryant PE, Riches AC, Thompson A,
Blasco MA, et al: Telomere length abnormalities in mammalian
radiosensitive cells. Cancer Res. 61:912–915. 2001.PubMed/NCBI
|
|
46
|
Lindqvist D, Epel ES, Mellon SH, Penninx
BW, Révész D, Verhoeven JE, Reus VI, Lin J, Mahan L, Hough CM, et
al: Psychiatric disorders and leukocyte telomere length: Underlying
mechanisms linking mental illness with cellular aging. Neurosci
Biobehav Rev. 55:333–364. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Masi S, Salpea KD, Li K, Parkar M, Nibali
L, Donos N, Patel K, Taddei S, Deanfield JE, D'Aiuto F, et al:
Oxidative stress, chronic inflammation, and telomere length in
patients with periodontitis. Free Radic Biol Med. 50:730–735. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Watfa G, Dragonas C, Brosche T, Dittrich
R, Sieber CC, Alecu C, Benetos A and Nzietchueng R: Study of
telomere length and different markers of oxidative stress in
patients with Parkinson's disease. J Nutr Health Aging. 15:277–281.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ma D, Zhu W, Hu S, Yu X and Yang Y:
Association between oxidative stress and telomere length in Type 1
and Type 2 diabetic patients. J Endocrinol Invest. 36:1032–1037.
2013.PubMed/NCBI
|
|
50
|
de Vos-Houben JM, Ottenheim NR, Kafatos A,
Buijsse B, Hageman GJ, Kromhout D and Giltay EJ: Telomere length,
oxidative stress, and antioxidant status in elderly men in Zutphen
and Crete. Mech Ageing Dev. 133:373–377. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Prasad KN, Wu M and Bondy SC: Telomere
shortening during aging: Attenuation by antioxidants and anti-
inflammatory agents. Mech Ageing Dev. 164:61–66. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Paul L: Diet, nutrition and telomere
length. J Nutr Biochem. 22:895–901. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Herrmann M, Pusceddu I, Marz W and
Herrmann W: Telomere biology and age- related diseases. Clin Chem
Lab Med. 56:1210–1222. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kiecolt-Glaser JK, Epel ES, Belury MA,
Andridge R, Lin J, Glaser R, Malarkey WB, Hwang BS and Blackburn E:
Omega- 3 fatty acids, oxidative stress, and leukocyte telomere
length: A randomized controlled trial. Brain Behav Immun. 28:16–24.
2013. View Article : Google Scholar
|