The liver serves a vital role in various
physiological processes, including glucose metabolism, fatty acid
metabolism, lipid metabolism, immune response and the secretion of
various cytokines (such as TGF-β, IL-6 and IL-10) (11-13). Liver diseases encompass
non-alcoholic fatty liver (NAFL) disease (NAFLD), liver fibrosis,
liver cirrhosis, acute liver failure (ALF) and hepatocellular
carcinoma (HCC) (11). Liver
diseases pose a notable global health burden, with an estimated 150
million cases of liver diseases resulting in ~2 million mortalities
annually (14,15). In the normal liver, there is a net
lactate uptake due to gluconeogenesis, and the liver has the
highest net lactate clearance rate compared with the other organs
of the body, which is expected to be ≤70% of the systemic clearance
(9,16). Thus, lactate and lactate
metabolism serve an important role in liver diseases. However,
there is not currently a systematic review that summarizes the role
of lactate and lactate metabolism in liver diseases. The aim of the
present review was to summarize the roles of lactate and lactate
metabolism in the liver and in the development of liver diseases.
Additionally, the present review may provide new directions and
guidance for future research and the treatment of liver
diseases.
The studies cited in the present review were
published between 1994 and 2024, with the majority published
between 2011 and 2024. All of the studies cited in the present
review were found on the PubMed database using the following
keywords: Lactate, lactate metabolism, lactate metabolism-related
genes (LMRGs), glucose metabolism, glycolysis, lactate shuttle,
monocarboxylate transporter (MCT), G protein-coupled receptor 81
(GPR81), histone lactylation, non-histone lactylation, NAFLD, liver
fibrosis, ALF and HCC.
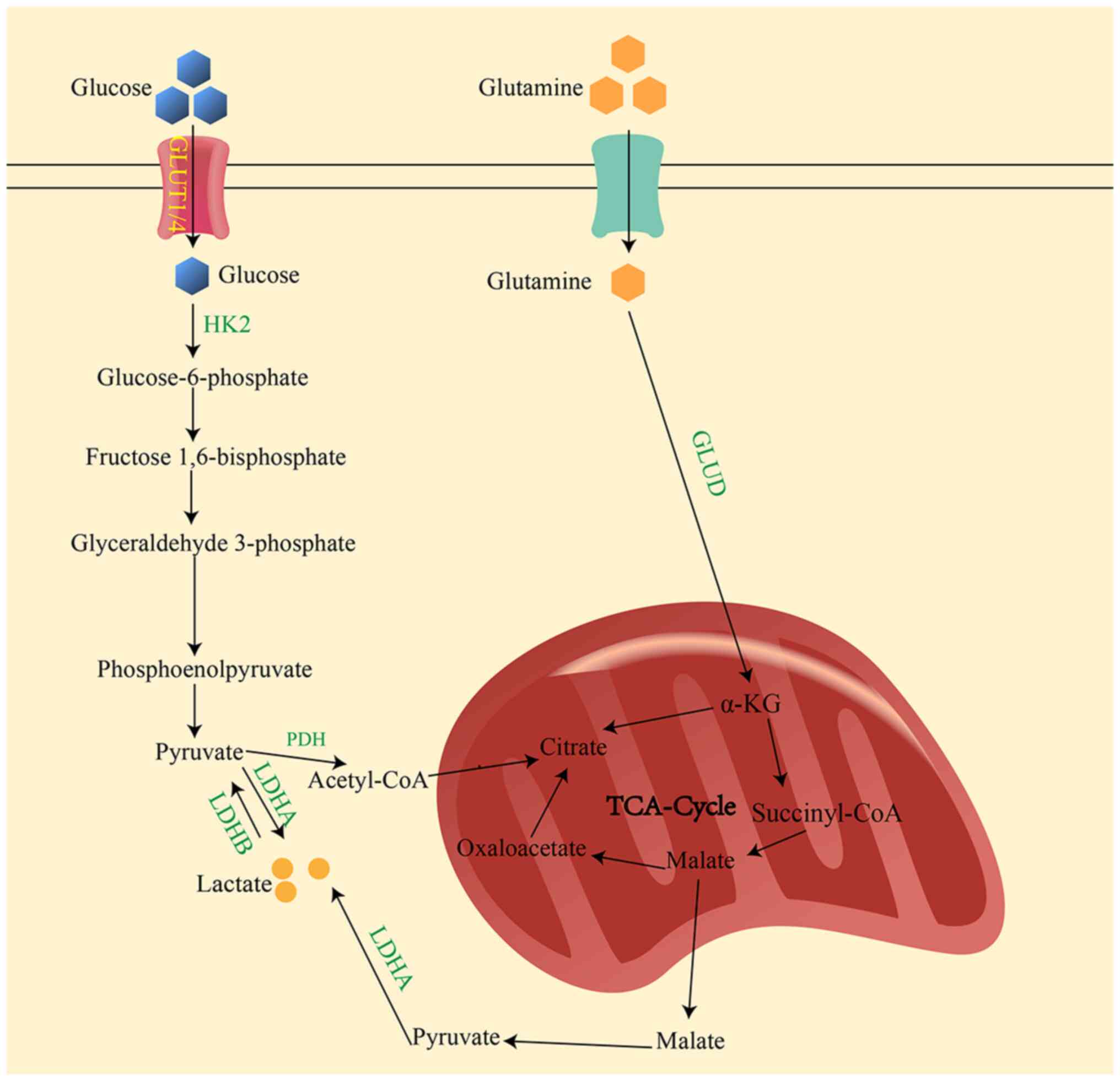
As the final product of glycolysis, lactate is
primarily produced under hypoxia conditions (7). In the cytoplasm, glucose undergoes a
series of catalytic reactions to form pyruvate, which is
subsequently converted to lactate in the presence of lactate
dehydrogenase (LDH)A (Fig. 1).
Another metabolic pathway that produces lactate includes the
conversion of alanine to glutamate via alanine aminotransferase,
which is also a minor source of lactate production in tumor cells
(17). Lactate exists in the body
as D-lactate, L-lactate and racemic DL-lactate. Among these,
L-lactate is the primary form in the human body, participating in
various biological processes (such as energy regulation and the
regulation of fatty acid metabolism) (9). D-lactate serves as the main
metabolite in bacteria (such as Lactobacillus and
colibacillus) found in the gut, and may be involved in the
transport of metabolic substrates (such as H+, pyruvic
acid and malate) within the body (9,18).
Accumulation of lactate in the body can be hazardous
and causes lactic acidosis (19).
Consequently, the body should efficiently and rapidly remove
lactate from tissues and circulation through metabolism. The
primary method of lactate clearance is through the oxidative
formation of pyruvate. This is followed by the formation of
acetyl-CoA, catalyzed by pyruvate dehydrogenase, which is then used
in the tricarboxylic acid cycle for the formation of
CO2, water and for providing energy (20,21). Another clearance pathway involves
activating gluconeogenesis in the liver and skeletal muscle cells
in response to hormones such as glucagon and cortisol, in which
lactate is converted into glucose, which is then released into the
bloodstream and further metabolized to provide energy to the body
(9,22). Under normal circumstances, the
liver exhibits the highest lactate clearance rate in the body
(9,16). When liver function is impaired, it
leads to dysregulation of lactate metabolism, and in patients with
chronic liver disease, lactate clearance is markedly reduced
leading to lactate accumulation (23).
Lactate, and its function as a signaling molecule,
has been investigated. It primarily exerts its effects by being
transported into cells via MCTs or by signaling through its
specific receptor, GPR81 (24,25).
GPR81 is widely distributed in tissues and organs
such as fat, kidney and liver (44). Lactate may inhibit IL1β expression
in macrophages by acting on GPR81 and arrestin β 2 to inhibit
Toll-like receptor (TLR) 4-triggered NLR family pyrin domain
containing 3 activation. Furthermore, low concentrations of lactate
can attenuate acute liver injury (45). Additionally, lactate can inhibit
lipolysis by activating GPR81 on the surface of adipocytes, which
downregulates cAMP levels to balance the energy metabolism between
glucose and lipids (46).
Metformin can increase GPR81 expression, improving mouse NAFLD
symptoms in a GPR81-dependent manner (47). However, knowledge of the role of
lactate/GRP81 in liver disease is limited and additional studies
are required.
As a metabolic substrate, the main function of
lactate is to generate pyruvate, which is catalyzed by LDHB.
Lactate also serves as a precursor of gluconeogenesis in the
synthesis of glucose, which is then used an energy source (Table II) (7,48-61). Additionally, lactate regulates
fatty acid metabolism and acts as a signaling molecule to modulate
cellular functions including the modulation of inflammatory
responses and cell proliferation (9). Besides these functions, Zhang et
al (62) showed that lactate
can regulate transcription through an epigenetic modification known
as lactylation. Lactylation is a post-translational modification
that occurs after the translation of proteins, directly promoting
gene transcription (62,63). When lactate levels are increased,
lactate is converted into lactyl CoA due to the action of a
currently unknown enzyme, and histone lysine residues are
lactylated by an effector protein (P300). Lactylation can be
modulated by effector proteins such as P300/cyclic AMP response
element-binding protein (CBP) (9,62).
Due to lactylation being considered a common post-translational
modification, investigating the role of lactylated proteins in the
occurrence and development of liver diseases in future research
will broaden the understanding of the mechanisms underlying
disrupted lactate metabolism in liver diseases.
Lactate serves an important role in inflammation,
immune energy metabolism and signaling pathway activation,
affecting inflammation processes and tumor immune tolerance
(9). The liver is an important
metabolic organ that coordinates various metabolic activities (such
as lipid fatty acid metabolism and immune responses) and serves an
essential role in several glucose metabolic pathways, including
gluconeogenesis, glycogenolysis and glycolysis (11,64,65). Normal lactate levels are 0.5-1.7
mmol/l (66), but lactate
clearance rates do not have a defined normal range. Elevated
lactate levels are observed in liver diseases, especially in
patients with chronic liver disease (67). Arterial serum lactate levels >2
mmol/l are associated with higher organ failure scores and higher
mortality (68). Comparison of
the 28-day survival in patients with liver cirrhosis admitted to
the intensive care unit (ICU) reveals that admission lactate
(1.2-3.4 mmol/l) are notably lower in surviving patients compared
with in those who died (2-9.7 mmol/l), whereas lactate clearance
(-9 to 50%) was markedly higher in surviving patients compared with
in those who died (-33 to 43%). Therefore, lactate is associated
with short-term mortality in critically ill patients with cirrhosis
and can be used as a prognostic indicator (68). Lactate and lactate metabolism are
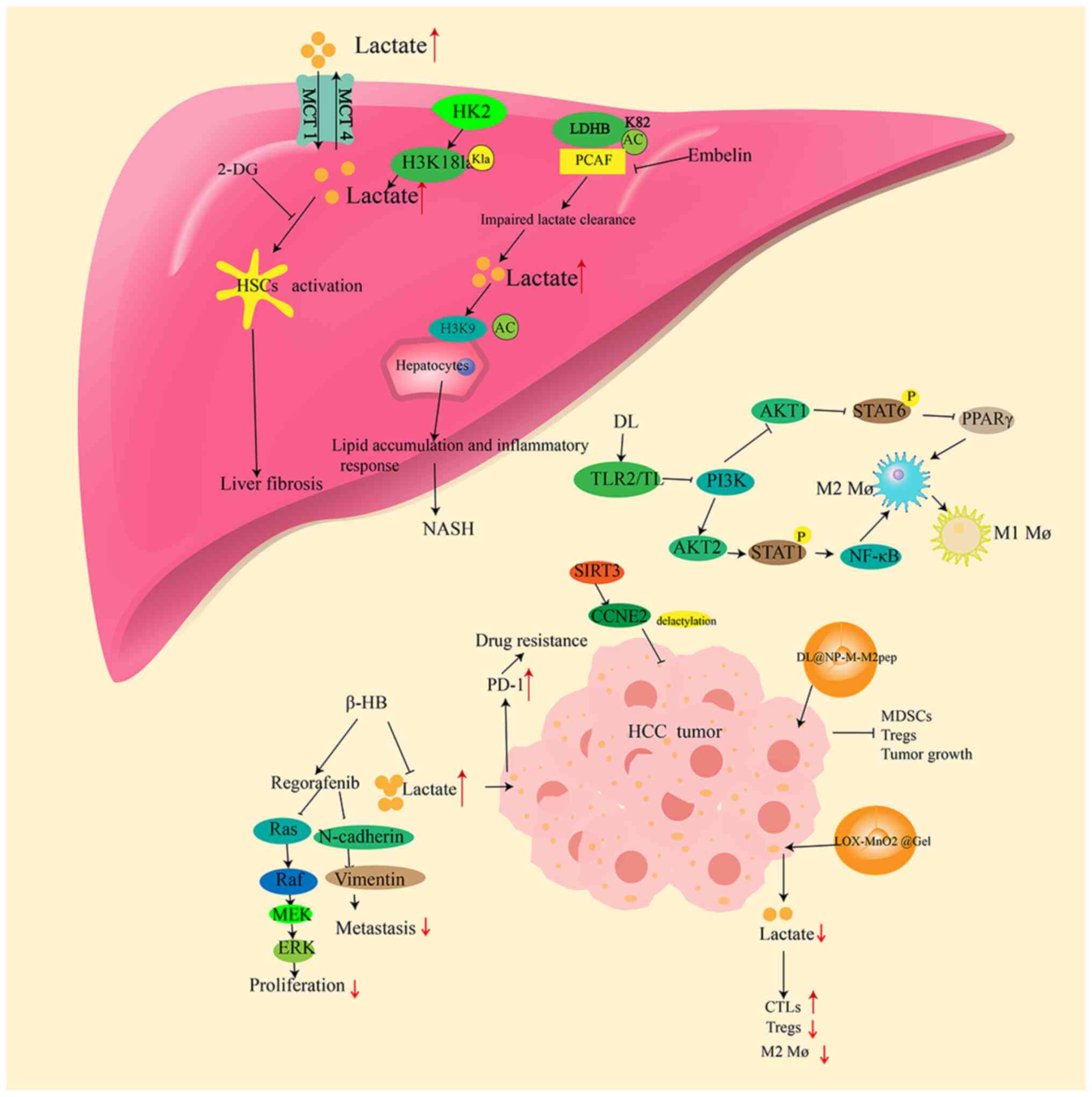
involved in liver fibrosis, NAFLD and HCC development (Fig. 2) (55-57). Lactate, LDH and LMRGs may be used
as predictors of liver failure and HCC prognosis (Table III) (59,68,69). Lactate can not only be used as a
clinical prognostic marker for liver disease, but can also be a
target for studying the pathogenesis and potential therapeutic
approaches for liver disease.
Liver fibrosis is a wound healing response to
various injuries to the liver and has a high morbidity rate
affecting >100 million individuals worldwide (11). Liver fibrosis is caused by a
variety of factors, including viral hepatitis, alcoholic liver
disease and NAFLD (11).
Additionally, as liver fibrosis progresses, liver function becomes
impaired, and further progression to liver cirrhosis will cause
ascites and esophagogastric fundus venous hypertension, which
decreases the quality of life of the patient, and it may progress
to HCC, affecting the prognosis of the patient (70,71). At present, besides liver
transplantation, there are no effective methods to cure liver
fibrosis (72). Liver fibrosis
mainly occurs due to the activation and transformation of quiescent
hepatic stellate cells (HSCs) into myofibroblasts, leading to
excessive extracellular matrix deposition (11,73). Research indicates that metabolic
reprogramming, including aerobic glycolysis, can activate HSCs
(74,75), and inhibition of aerobic
glycolysis can suppress HSC activation. After HSC activation,
lactate is involved in subsequent processes, including gene
expression (74,75).
In highly glycolytic proliferating cells, such as
cancer cells, hexokinase (HK)2 expression accelerates glucose
metabolism (76). In HSCs, HK2
deletion or pharmacological inhibition of lactate production can
reduce histone lactylation at H3K181a in HSCs, thereby inhibiting
HSC activation. This inhibition is reversed by supplementing
exogenous lactate (55).
Furthermore, HSC activation is inhibited, and hepatic fibrosis is
attenuated in mice with an HSC-specific knockout of HK2 (55). Thus, intervention of the
HK2/H3K18la axis is a potential therapeutic strategy for liver
fibrosis. Increased expression of MCT1 also promotes liver fibrosis
formation in non-alcoholic steatohepatitis (NASH) mouse models
(34). Increases in lactate
levels promotes HSC activation as well as lactate transport and
lactylation, which are associated with the development of liver
fibrosis. Therefore, the role of lactate metabolism in the
development of liver fibrosis should be investigated further in the
future.
The prevalence of NAFLD has made it a major global
health issue and the overall prevalence of NAFLD worldwide is
estimated to be 32.4% (77).
NAFLD comprises a spectrum of liver conditions, in which hepatic
steatosis (fatty liver) alone is referred to as NAFL and NASH is
defined as a more serious condition with inflammation and
hepatocyte damage (steatohepatitis) (77,78). As the disease progresses, a number
of patients may develop liver cirrhosis or HCC (79). Studies show that with an
increasing severity of liver disease, especially when the disease
progresses from steatosis to NASH, lactate levels gradually
increase in both the blood and liver (80,81). Protein acetylation is a major
mechanism in the development of chronic liver diseases (82). LDHB activity is markedly reduced
in the liver of patients with NAFLD or NASH (23). Similarly, in high fat diet
(HFD)-induced NAFLD mouse models, the LDHB activity is decreased
(23). Mass spectrometry reveals
that during the construction of the mouse model, the HFD
exacerbates lactate accumulation, decreases the liver lactate
clearance rate and alters the expression of the acetyltransferases
P300/CBP-associated factor (PCAF) (23). PCAF is the major regulatory factor
for LDHB acetylation at the K82 site (23). PCAF-dependent K82 acetylation
reduces LDHB activity and inhibits lactate clearance. Another study
suggests that H3K9 acetylation may aggravate lipid accumulation,
and overexpression of LDHB-K82Q (which inhibits the activation of
LDHB) in mice increases H3K9 histone acetylation, promotes lipid
accumulation and inflammatory reactions, and results in an
exacerbation of NAFLD (23).
Inhibition of PCAF reduces LDHB acetylation and alleviates hepatic
steatosis in NASH mice. This provides a potential therapeutic
target for NAFLD. MCT1 can promote liver steatosis, and the
knockdown of MCT1 in a mouse model attenuates the symptoms of NAFLD
(35-37). However, further studies are
required to investigate the role of lactate metabolic processes in
NAFLD.
ALF is a syndrome characterized by brain
dysfunction, coagulation disorders and multi-organ dysfunction
resulting from acute liver injury (83). A number of studies have
investigated the role of lactate in ALF. Bernal et al
(84) demonstrate that arterial
blood lactate levels can predict a poor prognosis of
acetaminophen-induced ALF. There is a notable association between
lactate levels and survival in response to ALF, including
acetaminophen-induced, non-acetaminophen-induced and edible
mushroom-induced AFL, and despite the low specificity of lactate as
a predictor of prognosis in patients with ALF, patients with high
lactate levels have lower survival rates (68,85-93). Furthermore, a study suggests that
lactate levels lack specificity as a criterion for urgent liver
transplantation in patients with ALF (94). Bernal (95) argues that the decision-making
process for liver transplantation is a dynamic one, and the level
of lactate remains an important component of the overall assessment
of patients with ALF, aiding transplant teams in making decisions
that contribute to the prognosis of the patients. Early
postoperative lactate levels are effective markers for clinically
relevant post-hepatectomy liver failure (PHLF) (96). Furthermore, elevated perioperative
lactate levels and decreased lactate clearance are associated with
the incidence of PHLF (97).
Therefore, lactate levels may assist in clinical decision-making
for patients with a liver transplant, such as timely administration
of preventive treatment and enhanced observation. The lactate
clearance rate serves as an independent predictor of mortality in
critically ill patients with liver cirrhosis and acute-on-chronic
liver failure (ACLF) (98).
Additionally, in patients with ACLF requiring ICU admission, a
lactate and organ failures predictive model, constructed from
lactate levels and a number of organ failures, demonstrates that
the lactate level and the number of organ failures at the time of
admission to the ICU predicts patient prognosis. Therefore, this
may allow for an improved risk stratification in order to optimize
strategies for organ support (99).
In patients with ALF, serum LDH levels are markedly
higher compared with those in patients with acute or chronic
hepatitis, or liver cirrhosis. Immunohistochemistry also shows a
relative increase in LDH expression levels (100). While LDH was previously
considered to have low diagnostic value in liver diseases due to
its production by various cells throughout the body (101), a recent study reveals that a
high lactate/albumin ratio is associated with an increased
mortality during hospitalization in patients with liver cirrhosis
(102). This ratio serves as an
independent predictive indicator of in-hospital mortality in
patients with cirrhosis (102).
Therefore, the present study suggests that lactate can be used as a
prognostic predictor for cirrhosis (103) as well as ALF. During the
development of ALF, the proteins involved in lactate metabolism
must be altered, therefore, it may be possible to identify more
specific LMRGs to predict the prognosis of ALF in the future.
Liver cancer is the 6th most common cancer and the
4th leading cause of cancer-related mortality in the world
(104). The most common type of
primary liver cancer is HCC, which accounts for 80-90% of cases
(104). Current effective
treatments for HCC include surgery, liver transplantation,
chemotherapy and targeted therapy, but overall survival (OS)
remains unsatisfactory (105).
Further research is required for the treatment of HCC.
The tumor microenvironment (TME) serves an important
role in cancer progression, consisting of tumor cells, immune
cells, stromal cells, blood vessels and the extracellular matrix
(106). Lactate accumulation
exacerbates hypoxia thereby activating hypoxia-inducible factor-1α
to further promote lactate production (107). Lactate accumulation in the TME
can lead to extracellular acidification, inhibiting the function of
T cells and natural killer (NK) cells, while enhancing the
immunosuppressive functions of tumor-associated macrophages (TAMs),
myeloid-derived suppressor cells (MDSCs) and regulatory T cells
(Tregs), thereby promoting tumor progression (9). Lactate also promotes hypoxia and
angiogenesis, further contributing to the immunosuppressive
functions of the TME (9).
Lactate induces the expression of programmed death-1
(PD-1) in monocytes and neutrophils (108). In addition, lactate can induce
PD-1 expression by activating signaling pathways mediated by TGF-β,
IFN-γ and TNF-α (105-108). Increased lactate activates the
TGF-β/Smad signaling pathway and subsequently activates
epithelial-mesenchymal transition (EMT)-related genes to promote
tumor progression (109,110). Lactate can lead to an increase
in the levels of hepatocyte growth factor (HGF) in
cancer-associated fibroblasts and subsequently activate the
mesenchymal epithelial transition-dependent signaling pathway in
cancer cells, maintaining resistance to tyrosine kinase inhibitors
(TKIs) (111). In HCC, lactate
promoted HCC progression by modulating HGF (112). Lactate upregulates IFN-γ
expression in M2 TAMs and promotes T-cell apoptosis through the
PD-1/PD-ligand (L)1 pathway (113). In HCC, lactate promotes PD-L1
upregulation by increasing TNF-α expression; blocking TNF-α
inhibits PD-L1 expression in TAMs (114). Both lactate and lactylation
promotes IL-6 secretion to promote tumor progression (115,116). Hence, increases in lactate
levels increases HCC resistance and decreases HCC treatment
efficacy (117).
Elevated lactate levels are observed in the tumor
and surrounding tissues, and in the serum of patients with HCC
(108,118). Lenvatinib administered in
combination with celecoxib reduces lactate-induced PD-L1 neutrophil
survival and thus, reduces the levels of PD-L1 neutrophils
increasing the antitumor effect of lenvatinib in subcutaneous and
orthotopic HCC mouse models (108). Overexpression of LDHA in HCC
cells increases the invasive capacity of HCC cells, and knockdown
of LDHA inhibits the metastatic potential in xenograft mice
(119). Ketogenesis primarily
occurs in the liver, and β-hydroxybutyrate (β-HB) is a ketone
produced during this process (120). Exogenous administration of β-HB
decreases LDHA expression and lactate production in
sorafenib-resistant HCC cells, enhancing the drug sensitivity of
HCC sorafenib-resistant cells by inhibiting the B-Raf/MAPK pathway
and EMT (117). However, in
xenograft models, β-HB inhibits the expression of proliferating
cell nuclear antigen and LDHA without markedly improving tumor size
and weight (117). While β-HB
treatment may reverse sorafenib resistance by downregulating
lactate production, further research is needed to confirm its
effectiveness in vivo (117). microRNA (miR)-34a serves an
anticancer role in HCC cells. By establishing a radioresistant HCC
cell line, miR-34a levels are markedly downregulated in HepG2
radioresistant cells, and overexpression of miR-34a re-sensitizes
cells to radiation treatment by inhibiting LDHA (121). The potassium inwardly-rectifying
channel, subfamily J, member 11 potassium channel can interact with
LDHA and enhance its enzymatic activity to promote HCC progression
(122). Knockdown of LDHA in
mice notably inhibits the growth of HCC, while the selective loss
of CD8+ and increases of CD4+ T lymphocytes
in the TME are observed (123).
However, further studies are needed to investigate the mechanism of
HCC development inhibition by knockdown of LDHA. A number of drugs
that target LDHA are effective in in vitro experiments but
are found to be ineffective when experimenting in vivo
(123,124). Additionally, a number of
effective in vivo LDHA inhibitors often exhibit off-target
effects, suggesting that their antitumor activity may not be solely
due to LDHA inhibition (125,126). Thus, further investigation is
required to determine whether LDHA is a suitable target for HCC
treatment.
Research shows that D-lactate can reach the liver
through the portal vein and enhance the ability of Kupffer cells to
clear pathogens from the bloodstream (127). In addition, D-lactate also
interacts with TLR2 and/or TLR9 on macrophages to inhibit the
PI3K/Akt pathway while activating the NF-κB pathway to promote the
transition of M2 TAMs to M1 TAMs (128). Targeted delivery of D-lactate to
macrophages through the nanoformulation DL@NP-M-M2
macrophage-binding peptide (DL@NP-M-M2pep) markedly inhibits tumor
growth in mouse models, improving survival rates (128). It reverses the immunosuppression
in TME by inhibiting MDSCs and Tregs while activating NK cells and
DCs (128). Another study used
nanotechnology to construct a nanoparticle-hydrogel composite
system, LOX-MnO2@Gel; this system depletes lactate from the TME
through a cascade catalytic reaction, then restores intratumoral
cytotoxic T lymphocyte function, reduces the ratio of Tregs/M2-like
macrophages, enhances antitumor immune responses and transforms the
immunosuppressive TME into an immunocompetent one (129). This approach markedly inhibits
residual tumor growth, suppresses lung metastasis and prolongs
mouse survival in subcutaneous and in situ HCC mouse models
(129). Combining metabolic
therapy with immunotherapy provides new insights for the treatment
of HCC recurrence post-ablation, but long-term research is
required. Targeting lactate has also been shown to be a possible
treatment for HCC.
LDH can serve a predictive role in the treatment of
sorafenib in patients with renal cell, rectal and lung cancer
(130-132). Earlier studies have also shown
the same predictive value in HCC (133). However, another study suggests
that baseline LDH levels are not associated with the prognosis of
patients with HCC undergoing sorafenib treatment (134). In a previous study, baseline LDH
levels in patients with HCC were found to be influenced by the
degree of liver fibrosis, independent of HCC staging (135). Lower baseline LDH levels were
identified as an independent prognostic factor for an improved
response to sorafenib. The study also found that a marked increase
in serum LDH levels during sorafenib administration may indicate
the potential development of ALF (135). In patients with HCC undergoing
liver resection, low LDH levels are associated with improved OS and
recurrence-free survival (136).
Preoperative serum LDH levels can assess the long-term prognosis of
patients with HCC undergoing transarterial chemoembolization (TACE)
(137). In addition, an increase
in LDH after undergoing TACE also implies poorer OS (138). Another study involving 2,327
patients with HCC indicated that high LDH levels and a high ratio
of alkaline phosphatase/LDH are associated with poor OS (139). The positive rates of LDHC mRNA
expression in serum and in serum exosomes of patients with HCC were
68 and 60%, respectively. The LDHC expression levels were
negatively associated with HCC prognosis, serving as a predictor
for HCC prognosis (140).
Therefore, current research supports LDH as a prognostic indicator
for HCC treatment.
Lactate transport also serves a role in the
development of HCC. MCT1 is highly expressed in HCC tissues
compared with adjacent tissues (38). Additionally, MCT4 is highly
expressed in HCC cells and tissues compared with normal hepatocytes
and adjacent tissues. MCT4 can promote tumor cell proliferation,
invasion and metastasis, and is strongly associated with the poor
prognosis of patients with HCC (39,40). It is also involved in HCC
progression by promoting the expression of trafficking protein
particle complex subunit 5 (41).
When MCT4 is inhibited, it leads to the disruption of pH
homeostasis in HCC cells, which induces apoptosis and inhibits
migration and invasion (42).
Lactylation primarily affects enzymes involved in
metabolism, and is associated with cellular energy metabolism.
Adenylate kinase 2 (AK2) is a key enzyme in the transfer of
phosphate groups between adenosine monophosphate and ATP to produce
adenosine diphosphate. AK2 lactylation markedly reduces AK2 enzyme
activity, leading to energetic disturbances in HCC cells, which
promotes HCC progression (56).
The NAD-dependent deacetylase sirtuin 3 can inhibit the development
of HCC by promoting cell cycle protein E2 delactylation (141). Pan et al (57) also reveal that histone lactate
levels in tumor tissues of patients with HCC are notably higher
compared with those in adjacent tissues, and that lactylation of H3
histones can promote the progression of HCC. Silencing HK2 in HCC
cells inhibits tumorigenesis and promotes cell death, and knockdown
of HK2 in mice suppresses the incidence of HCC by inhibiting
lactate production (55). By
analyzing the relevant lactonization genes of patients with HCC in
The Cancer Genome Atlas and the International Cancer Genome
Consortium databases, Cheng et al (142) found that 16 lactylation-related
genes were associated with the prognosis of HCC, and eight
differential genes, which were further filtered to be included in
the lactylation score, were found to be negatively associated with
prognosis; therefore, lactylation-related genes have the potential
to serve as a prognostic biomarker for HCC in the future. Lactate
production, lactate shuttling and lactylation are involved in the
development of HCC, therefore, targeting lactate metabolism is a
potential approach to treating HCC; however, additional research is
required.
Efforts have been made to identify effective
treatment methods and diagnostic markers for liver diseases. When
liver diseases progress to liver cirrhosis and HCC, the prognosis
for patients is poor. Lactate levels can be useful for assessing
the prognosis of ALF and liver cirrhosis (84-103), and LDH can also be used as a
predictor of therapeutic sensitivity in HCC (133-140). LDHA, HK2, MCTs and HK2 are also
potential targets for the treatment of liver diseases (23,35-37,42,43,55,123). Lactate-targeted amelioration of
liver disease mainly occurs through the regulation of lactate
production, lactate transport and lactylation (Table IV).
Glycolysis is an important source of lactate
production, and glucose transport is also a regulator of lactate
production. Lactate production can be regulated by targeting
glycolytic pathway-related proteins including LDH, glucose
transporter, MCT, HK2 and pyruvate kinase M2 (PKM2) (143,144). Reducing lactate can inhibit the
activation of HSCs, thereby suppressing the occurrence and
development of liver fibrosis (55). Kruppel like factor 5 (KLF5)
promotes glycolysis, leading to an increased LDHA expression.
Curcumol inhibits liver fibrosis by blocking the KLF5/LDHA feedback
loop (145). The Wnt/β-catenin
signaling pathway enhances LDHA stability, promoting glycolysis and
liver fibrosis. In mice, the specific deletion of LDHA in HSCs
alleviates liver fibrosis (146). Therefore, the inhibition of LDHA
may be an effective treatment for liver fibrosis. By inhibiting HK,
2-deoxy-D-glucose (2-DG) inhibits glycolysis, which improves liver
fibrosis (147). The combination
of 2-DG with sorafenib inhibits HCC cell proliferation and improves
sorafenib resistance (148,149). However, a number of studies
suggest that 2-DG has no marked impact on tumor growth at doses
that do not cause severe adverse reactions (150,151). A recent study, in which 2-DG is
delivered to the liver via nanoparticles, demonstrates that it
increases the antitumor effects of sorafenib while producing
antitumor effects in anti-PD-1-resistant tumors (152). Additionally, 3-bromopyruvate is
a HK2 inhibitor that suppresses HCC cell proliferation and
movement, enhances sorafenib efficacy, and is considered a
potential sensitizer for clinical chemotherapy (153-155). Quercetin is a bioactive
flavonoid that can inhibit HK2-dependent glycolysis and thus,
inhibit HCC progression (156).
Oviductus ranae protein hydrolysate (ORPH) has immunomodulatory and
anti-glioma activities. ORPH can inhibit HCC progression by
targeting the miR-491-5p/PKM2 axis to inhibit glycolysis (157). Galloflavin, an LDHA inhibitor,
alleviates liver damage in ALF mouse models (158). Quinoline-3-sulfonamides are also
inhibitors of LDHA, and quinoline-3-sulfonamides and galloflavin
also inhibit the proliferation of HCC cells (159). As an inhibitor of LDHA, oxamate
enhances the antitumor activity of sorafenib, imatinib and
sunitinib in HCC (160). Liver
fibrosis, liver injury and HCC can be ameliorated by targeting
lactate production; however, long-term studies are required to
investigate the application of this in clinical treatment.
MCT1 and 4 serve important roles in the occurrence
and development of liver diseases. Inhibiting the transport of
lactate is a potential target for cancer therapy (161). Therefore, inhibiting lactate
transport may be beneficial for improving liver diseases. In mice,
knocking out MCT1 alleviates symptoms of NAFLD (162). Upregulation of MCT1 in Tregs
promotes resistance to PD-1 therapy in patients with HCC (43). A previous study demonstrates that
ARC155858, an inhibitor of MCT1, can inhibit proliferation and
lipid synthesis in HCC cells, but further in vivo studies
are required to confirm this finding (163). Thus, in-depth studies are
required to determine whether an MCT1 inhibitor can improve liver
diseases, and further research is warranted in the future.
Inhibition of MCT4 disrupts the intracellular pH homeostasis and
initiates apoptosis in HCC cells (42). The MCT4 inhibitor VB124 enhances T
cell infiltration and the efficacy of anti-PD-1 immunotherapy in a
HCC mouse model (164).
Inhibiting lactylation may also be a therapeutic
strategy for HCC. Demethylzeylasteral, a triterpenoid anti-tumor
compound, can inhibit the development of HCC by suppressing the
lactylation of H3 histones, thereby inhibiting the tumorigenicity
induced by liver cancer stem cells (57). Royal jelly acid (RJA), a major
unsaturated fatty acid in natural compound royal jelly, inhibits
the proliferation and migration of HCC cells and promotes
apoptosis. In a subcutaneous HCC model, RJA inhibits tumor growth
by inhibiting the lactylation of H3K9la and H3K14la sites on H3
histone (165). Glypican-3
(GPC3), a member of the glypican family, is expressed at high
levels in HCC and has diagnostic value (166). Recent research indicates that
GPC3 promotes lactate production, contributing to HCC development
by enhancing the overall lactate levels and c-myc lactylation
(167). In future research, a
new direction may be to target lactylation to study the treatment
of liver diseases. In addition, LMRGs are also potential markers
and therapeutic targets for predicting the prognosis of liver
diseases.
Lactate and lactate metabolism serve an essential
role in the development and progression of liver diseases.
Abnormalities in lactate production and transport, and lactylation
contribute to the development of liver disease, while lactate
levels can predict the prognosis of ALF and liver cirrhosis. LDH
can be used as a predictor of the therapeutic sensitivity of HCC.
Targeting lactate production and transport, regulating circulating
lactate levels and inhibiting lactylation may serve as potential
future strategies for the treatment of liver disease. Several
studies have already been conducted (57,145,147-160,164,165), but the role of lactate transport
and lactylation in liver disease should be further investigated in
the future. Lactate metabolism involves a number of genes; LMRGs
may exist as biomarkers and therapeutic targets for liver diseases
such as liver fibrosis, ALF and HCC. In-depth basic and clinical
studies are required to confirm the role of lactate metabolism in
liver diseases. Summarizing the currently available studies may
help guide future research.
Not applicable.
SY drafted the manuscript. HC, TT, LZ, XY and ZY
participated in the literature search and analysis of the data to
be included in the review. XL, YW, JA and GW were involved in the
design of the study and assisted in the preparation of the figures
and tables. HJ and BT edited and revised the manuscript. All
authors read and approved the final version of the manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81960507, 82073087
and 82160112), the collaborative Innovation Center of Chinese
Ministry of Education (grant no. 2020-39), the Science and
Technology Bureau fund of Zunyi city [grant no. ZUN SHI KE HE HZ ZI
(2019)93-Hao], and the Science and Technology Plan Project of
Guizhou Province [grant nos. QIAN KE HE JI cHU-ZK (2021) YI BAN451
and QIAN KE HE LH ZI (2017)7095 HAO].
|
1
|
Ferguson BS, Rogatzki MJ, Goodwin ML, Kane
DA, Rightmire Z and Gladden LB: Lactate metabolism: Historical
context, prior misinterpretations, and current understanding. Eur J
Appl Physiol. 118:691–728. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liberti MV and Locasale JW: The warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Certo M, Tsai CH, Pucino V, Ho PC and
Mauro C: Lactate modulation of immune responses in inflammatory
versus tumour microenvironments. Nat Rev Immunol. 21:151–161. 2021.
View Article : Google Scholar
|
|
4
|
Syed M, Kammala AK, Callahan B,
Oskeritzian CA and Subramanian H: Lactic acid suppresses MRGPRX2
mediated mast cell responses. Cell Immunol. 368:1044222021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Souto-Carneiro MM, Klika KD, Abreu MT,
Meyer AP, Saffrich R, Sandhoff R, Jennemann R, Kraus FV, Tykocinski
L, Eckstein V, et al: Effect of increased lactate dehydrogenase a
activity and aerobic glycolysis on the proinflammatory profile of
autoimmune CD8+ T cells in rheumatoid arthritis. Arthritis
Rheumatol. 72:2050–2064. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brooks GA: Lactate shuttles in nature.
Biochem Soc Trans. 30:258–264. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brooks GA: Cell-cell and intracellular
lactate shuttles. J Physiol. 587(Pt 23): 5591–5600. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang C, Pan RY, Guan F and Yuan Z: Lactate
metabolism in neurodegenerative diseases. Neural Regen Res.
19:69–74. 2024. View Article : Google Scholar
|
|
9
|
Li X, Yang Y, Zhang B, Lin X, Fu X, An Y,
Zou Y, Wang JX, Wang Z and Yu T: Lactate metabolism in human health
and disease. Signal Transduct Target Ther. 7:3052022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gaffney DO, Jennings EQ, Anderson CC,
Marentette JO, Shi T, Schou Oxvig AM, Streeter MD, Johannsen M,
Spiegel DA, Chapman E, et al: Non-enzymatic lysine lactoylation of
glycolytic enzymes. Cell Chem Biol. 27:206–213.e6. 2020. View Article : Google Scholar :
|
|
11
|
Yao S, Yang X, An J, Jin H, Wen G, Wang H
and Tuo B: Role of the S100 protein family in liver disease
(Review). Int J Mol Med. 48:1662021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kang JH, Toita R and Murata M: Liver
cell-targeted delivery of therapeutic molecules. Crit Rev
Biotechnol. 36:132–143. 2016. View Article : Google Scholar
|
|
13
|
Gao B: Hepatoprotective and
anti-inflammatory cytokines in alcoholic liver disease. J
Gastroenterol Hepatol. 27(Suppl 2): S89–S93. 2012. View Article : Google Scholar
|
|
14
|
Asrani SK, Devarbhavi H, Eaton J and
Kamath PS: Burden of liver diseases in the world. J Hepatol.
70:151–171. 2019. View Article : Google Scholar
|
|
15
|
Paik JM, Golabi P, Younossi Y, Mishra A
and Younossi ZM: Changes in the global burden of chronic liver
diseases from 2012 to 2017: The growing impact of NAFLD.
Hepatology. 72:1605–1616. 2020. View Article : Google Scholar
|
|
16
|
van Hall G: Lactate kinetics in human
tissues at rest and during exercise. Acta Physiol (Oxf).
199:499–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feron O: Pyruvate into lactate and back:
From the Warburg effect to symbiotic energy fuel exchange in cancer
cells. Radiother Oncol. 92:329–333. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
de Bari L, Atlante A, Guaragnella N,
Principato G and Passarella S: D-Lactate transport and metabolism
in rat liver mitochondria. Biochem J. 365(Pt 2): 391–403. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bennis Y, Bodeau S, Batteux B,
Gras-Champel V, Masmoudi K, Maizel J, De Broe ME, Lalau JD and
Lemaire-Hurtel AS: A study of associations between plasma metformin
concentration, lactic acidosis, and mortality in an emergency
hospitalization context. Crit Care Med. 48:e1194–e1202. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jha MK, Lee IK and Suk K: Metabolic
reprogramming by the pyruvate dehydrogenase kinase-lactic acid
axis: Linking metabolism and diverse neuropathophysiologies.
Neurosci Biobehav Rev. 68:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Soreze Y, Boutron A, Habarou F, Barnerias
C, Nonnenmacher L, Delpech H, Mamoune A, Chrétien D, Hubert L,
Bole-Feysot C, et al: Mutations in human lipoyltransferase gene
LIPT1 cause a Leigh disease with secondary deficiency for pyruvate
and alpha-ketoglutarate dehydrogenase. Orphanet J Rare Dis.
8:1922013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Emhoff CA, Messonnier LA, Horning MA,
Fattor JA, Carlson TJ and Brooks GA: Gluconeogenesis and hepatic
glycogenolysis during exercise at the lactate threshold. J Appl
Physiol (1985). 114:297–306. 2013. View Article : Google Scholar
|
|
23
|
Wang T, Chen K, Yao W, Zheng R, He Q, Xia
J, Li J, Shao Y, Zhang L, Huang L, et al: Acetylation of lactate
dehydrogenase B drives NAFLD progression by impairing lactate
clearance. J Hepatol. 74:1038–1052. 2021. View Article : Google Scholar
|
|
24
|
Brown TP and Ganapathy V: Lactate/GPR81
signaling and proton motive force in cancer: Role in angiogenesis,
immune escape, nutrition, and Warburg phenomenon. Pharmacol Ther.
206:1074512020. View Article : Google Scholar
|
|
25
|
Felmlee MA, Jones RS, Rodriguez-Cruz V,
Follman KE and Morris ME: Monocarboxylate transporters (SLC16):
Function, regulation, and role in health and disease. Pharmacol
Rev. 72:466–485. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Halestrap AP: The SLC16 gene
family-structure, role and regulation in health and disease. Mol
Aspects Med. 34:337–349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun S, Li H, Chen J and Qian Q: Lactic
Acid: No longer an inert and end-product of glycolysis. Physiology
(Bethesda). 32:453–463. 2017.PubMed/NCBI
|
|
28
|
Contreras-Baeza Y, Sandoval PY, Alarcón R,
Galaz A, Cortés-Molina F, Alegría K, Baeza-Lehnert F, Arce-Molina
R, Guequén A, Flores CA, et al: Monocarboxylate transporter 4
(MCT4) is a high affinity transporter capable of exporting lactate
in high-lactate microenvironments. J Biol Chem. 294:20135–20147.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Halestrap AP: Monocarboxylic acid
transport. Compr Physiol. 3:1611–1643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Valença I, Ferreira AR, Correia M, Kühl S,
van Roermund C, Waterham HR, Máximo V, Islinger M and Ribeiro D:
Prostate cancer proliferation is affected by the subcellular
localization of MCT2 and accompanied by significant peroxisomal
alterations. Cancers (Basel). 12:31522020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang G, Zhang Y, Dong D, Wang F, Ma X,
Guan F and Sun L: MCT1 regulates aggressive and metabolic
phenotypes in bladder cancer. J Cancer. 9:2492–2501. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Halestrap AP: The monocarboxylate
transporter family-Structure and functional characterization. IUBMB
Life. 64:1–9. 2012. View Article : Google Scholar
|
|
33
|
Droździk M, Szeląg-Pieniek S,
Grzegółkowska J, Łapczuk-Romańska J, Post M, Domagała P,
Miętkiewski J, Oswald S and Kurzawski M: Monocarboxylate
transporter 1 (MCT1) in liver pathology. Int J Mol Sci.
21:16062020. View Article : Google Scholar
|
|
34
|
Min K, Yenilmez B, Kelly M, Echeverria D,
Elleby M, Lifshitz LM, Raymond N, Tsagkaraki E, Harney SM, DiMarzio
C, et al: Lactate transporter MCT1 in hepatic stellate cells
promotes fibrotic collagen expression in nonalcoholic
steatohepatitis. bioRxiv [Preprint] 2023.05.03.539244. 2023.
|
|
35
|
Martini T, Ripperger JA, Chavan R, Stumpe
M, Netzahualcoyotzi C, Pellerin L and Albrecht U: The hepatic
monocarboxylate transporter 1 (MCT1) contributes to the regulation
of food anticipation in mice. Front Physiol. 12:6654762021.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carneiro L, Asrih M, Repond C, Sempoux C,
Stehle JC, Leloup C, Jornayvaz FR and Pellerin L: AMPK activation
caused by reduced liver lactate metabolism protects against hepatic
steatosis in MCT1 haploinsufficient mice. Mol Metab. 6:1625–1633.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lengacher S, Nehiri-Sitayeb T, Steiner N,
Carneiro L, Favrod C, Preitner F, Thorens B, Stehle JC, Dix L,
Pralong F, et al: Resistance to diet-induced obesity and associated
metabolic perturbations in haploinsufficient monocarboxylate
transporter 1 mice. PLoS One. 8:e825052013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fan Q, Yang L, Zhang X, Ma Y, Li Y, Dong
L, Zong Z, Hua X, Su D, Li H and Liu J: Autophagy promotes
metastasis and glycolysis by upregulating MCT1 expression and
Wnt/β-catenin signaling pathway activation in hepatocellular
carcinoma cells. J Exp Clin Cancer Res. 37:92018. View Article : Google Scholar
|
|
39
|
Gao HJ, Zhao MC, Zhang YJ, Zhou DS, Xu L,
Li GB, Chen MS and Liu J: Monocarboxylate transporter 4 predicts
poor prognosis in hepatocellular carcinoma and is associated with
cell proliferation and migration. J Cancer Res Clin Oncol.
141:1151–1162. 2015. View Article : Google Scholar
|
|
40
|
Chen HL, OuYang HY, Le Y, Jiang P, Tang H,
Yu ZS, He MK, Tang YQ and Shi M: Aberrant MCT4 and GLUT1 expression
is correlated with early recurrence and poor prognosis of
hepatocellular carcinoma after hepatectomy. Cancer Med.
7:5339–5350. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Niu Z, Yang F, Li H, Wang J, Ni Q, Ma C,
Zhu H, Chang H, Zhou X, Lu J and Gao H: MCT4 promotes
hepatocellular carcinoma progression by upregulating TRAPPC5 gene.
J Hepatocell Carcinoma. 9:289–300. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao Y, Li W, Li M, Hu Y, Zhang H, Song G,
Yang L, Cai K and Luo Z: Targeted inhibition of MCT4 disrupts
intracellular pH homeostasis and confers self-regulated apoptosis
on hepatocellular carcinoma. Exp Cell Res. 384:1115912019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhou J, Shao Q, Lu Y, Li Y, Xu Z, Zhou B,
Chen Q, Li X, Xu X, Pan Y, et al: Monocarboxylate transporter
upregulation in induced regulatory T cells promotes resistance to
anti-PD-1 therapy in hepatocellular carcinoma patients. Front
Oncol. 12:9600662022. View Article : Google Scholar
|
|
44
|
Hu J, Cai M, Liu Y, Liu B, Xue X, Ji R,
Bian X and Lou S: The roles of GRP81 as a metabolic sensor and
inflammatory mediator. J Cell Physiol. 235:8938–8950. 2020.
View Article : Google Scholar
|
|
45
|
Hoque R, Farooq A, Ghani A, Gorelick F and
Mehal WZ: Lactate reduces liver and pancreatic injury in Toll-like
receptorand inflammasome-mediated inflammation via GPR81-mediated
suppression of innate immunity. Gastroenterology. 146:1763–1774.
2014. View Article : Google Scholar
|
|
46
|
Ahmed K, Tunaru S, Tang C, Müller M, Gille
A, Sassmann A, Hanson J and Offermanns S: An autocrine lactate loop
mediates insulin-dependent inhibition of lipolysis through GPR81.
Cell Metab. 11:311–319. 2010. View Article : Google Scholar
|
|
47
|
Wu G, Dai Y, Yan Y, Zheng X, Zhang H, Li H
and Chen W: The lactate receptor GPR81 mediates hepatic lipid
metabolism and the therapeutic effect of metformin on experimental
NAFLDs. Eur J Pharmacol. 924:1749592022. View Article : Google Scholar
|
|
48
|
Hui S, Ghergurovich JM, Morscher RJ, Jang
C, Teng X, Lu W, Esparza LA, Reya T, Le Zhan, Yanxiang Guo J, et
al: Glucose feeds the TCA cycle via circulating lactate. Nature.
551:115–118. 2017. View Article : Google Scholar
|
|
49
|
Dienel GA: Brain glucose metabolism:
Integration of energetics with function. Physiol Rev. 99:949–1045.
2019. View Article : Google Scholar
|
|
50
|
Lhomme T, Clasadonte J, Imbernon M,
Fernandois D, Sauve F, Caron E, da Silva Lima N, Heras V,
Martinez-Corral I, Mueller-Fielitz H, et al: Tanycytic networks
mediate energy balance by feeding lactate to glucose-insensitive
POMC neurons. J Clin Invest. 131:e1405212021. View Article : Google Scholar
|
|
51
|
Gómez-Valadés AG, Pozo M, Varela L,
Boudjadja MB, Ramírez S, Chivite I, Eyre E, Haddad-Tóvolli R, Obri
A, Milà-Guasch M, et al: Mitochondrial cristae-remodeling protein
OPA1 in POMC neurons couples Ca(2+) homeostasis with adipose tissue
lipolysis. Cell Metab. 33:1820–1835.e9. 2021. View Article : Google Scholar
|
|
52
|
Faubert B, Li KY, Cai L, Hensley CT, Kim
J, Zacharias LG, Yang C, Do QN, Doucette S, Burguete D, et al:
Lactate metabolism in human lung tumors. Cell. 171:358–371.e9.
2017. View Article : Google Scholar
|
|
53
|
Pucino V, Certo M, Bulusu V, Cucchi D,
Goldmann K, Pontarini E, Haas R, Smith J, Headland SE, Blighe K, et
al: Lactate buildup at the site of chronic inflammation promotes
disease by inducing CD4(+) T cell metabolic rewiring. Cell Metab.
30:1055–1074.e8. 2019. View Article : Google Scholar
|
|
54
|
Irizarry-Caro RA, McDaniel MM, Overcast
GR, Jain VG, Troutman TD and Pasare C: TLR signaling adapter BCAP
regulates inflammatory to reparatory macrophage transition by
promoting histone lactylation. Proc Natl Acad Sci USA.
117:30628–30638. 2020. View Article : Google Scholar
|
|
55
|
Rho H, Terry AR, Chronis C and Hay N:
Hexokinase 2-mediated gene expression via histone lactylation is
required for hepatic stellate cell activation and liver fibrosis.
Cell Metab. 35:1406–1423.e8. 2023. View Article : Google Scholar
|
|
56
|
Yang Z, Yan C, Ma J, Peng P, Ren X, Cai S,
Shen X, Wu Y, Zhang S, Wang X, et al: Lactylome analysis suggests
lactylation-dependent mechanisms of metabolic adaptation in
hepatocellular carcinoma. Nat Metab. 5:61–79. 2023. View Article : Google Scholar
|
|
57
|
Pan L, Feng F, Wu J, Fan S, Han J, Wang S,
Yang L, Liu W, Wang C and Xu K: Demethylzeylasteral targets lactate
by inhibiting histone lactylation to suppress the tumorigenicity of
liver cancer stem cells. Pharmacol Res. 181:1062702022. View Article : Google Scholar
|
|
58
|
Hayes C, Donohoe CL, Davern M and Donlon
NE: The oncogenic and clinical implications of lactate induced
immunosuppression in the tumour microenvironment. Cancer Lett.
500:75–86. 2021. View Article : Google Scholar
|
|
59
|
Li Y, Mo H, Wu S, Liu X and Tu K: A novel
lactate metabolism-related gene signature for predicting clinical
outcome and tumor microenvironment in hepatocellular carcinoma.
Front Cell Dev Biol. 9:8019592022. View Article : Google Scholar
|
|
60
|
Yang L, Tan P, Sun H, Zeng Z and Pan Y:
Integrative dissection of novel lactate metabolism-related
signature in the tumor immune microenvironment and prognostic
prediction in breast cancer. Front Oncol. 12:8747312022. View Article : Google Scholar
|
|
61
|
Wang Z, Zhang S, Li J, Yuan Y, Chen S, Zuo
M, Li W, Feng W, Chen M and Liu Y: Prognostic value of lactate
metabolism-related gene expression signature in adult primary
gliomas and its impact on the tumor immune microenvironment. Front
Oncol. 12:10082192022. View Article : Google Scholar
|
|
62
|
Zhang D, Tang Z, Huang H, Zhou G, Cui C,
Weng Y, Liu W, Kim S, Lee S, Perez-Neut M, et al: Metabolic
regulation of gene expression by histone lactylation. Nature.
574:575–580. 2019. View Article : Google Scholar
|
|
63
|
Izzo LT and Wellen KE: Histone lactylation
links metabolism and gene regulation. Nature. 574:492–493. 2019.
View Article : Google Scholar
|
|
64
|
Oosterveer MH and Schoonjans K: Hepatic
glucose sensing and integrative pathways in the liver. Cell Mol
Life Sci. 71:1453–1467. 2014. View Article : Google Scholar
|
|
65
|
Lei Y, Han P, Chen Y, Wang H, Wang S, Wang
M, Liu J, Yan W, Tian D and Liu M: Protein arginine
methyltransferase 3 promotes glycolysis and hepatocellular
carcinoma growth by enhancing arginine methylation of lactate
dehydrogenase A. Clin Transl Med. 12:e6862022. View Article : Google Scholar
|
|
66
|
Lazzeri C, Gensini GF, Sori A, Bernardo P,
Chiostri M, Tommasi E, Grossi F and Valente S: Dynamic behaviour of
lactate values during mild hypothermia in patients with cardiac
arrest. Eur Heart J Acute Cardiovasc Care. 3:176–182. 2014.
View Article : Google Scholar
|
|
67
|
Scheiner B, Lindner G, Reiberger T,
Schneeweiss B, Trauner M, Zauner C and Funk GC: Acid-base disorders
in liver disease. J Hepatol. 67:1062–1073. 2017. View Article : Google Scholar
|
|
68
|
Drolz A, Horvatits T, Rutter K, Landahl F,
Roedl K, Meersseman P, Wilmer A, Kluwe J, Lohse AW, Kluge S, et al:
Lactate improves prediction of short-term mortality in Critically
Ill patients with cirrhosis: A multinational study. Hepatology.
69:258–269. 2019. View Article : Google Scholar
|
|
69
|
Gao Y, Zhang H, Zhong H, Yang S and Wang
Q: Lactate and blood ammonia on admission as biomarkers to predict
the prognosis of patients with acute mushroom poisoning and liver
failure: A retrospective study. Toxicol Res (Camb). 10:850–855.
2021. View Article : Google Scholar
|
|
70
|
Roehlen N, Crouchet E and Baumert TF:
Liver fibrosis: Mechanistic concepts and therapeutic perspectives.
Cells. 9:8752020. View Article : Google Scholar
|
|
71
|
Dulai PS, Singh S, Patel J, Soni M, Prokop
LJ, Younossi Z, Sebastiani G, Ekstedt M, Hagstrom H, Nasr P, et al:
Increased risk of mortality by fibrosis stage in nonalcoholic fatty
liver disease: Systematic review and meta-analysis. Hepatology.
65:1557–1565. 2017. View Article : Google Scholar
|
|
72
|
Tan Z, Sun H, Xue T, Gan C, Liu H, Xie Y,
Yao Y and Ye T: Liver fibrosis: Therapeutic targets and advances in
drug therapy. Front Cell Dev Biol. 9:7301762021. View Article : Google Scholar
|
|
73
|
Sherman MH: Stellate cells in tissue
repair, inflammation, and cancer. Annu Rev Cell Dev Biol.
34:333–355. 2018. View Article : Google Scholar
|
|
74
|
Mejias M, Gallego J, Naranjo-Suarez S,
Ramirez M, Pell N, Manzano A, Suñer C, Bartrons R, Mendez R and
Fernandez M: CPEB4 increases expression of PFKFB3 to induce
glycolysis and activate mouse and human hepatic stellate cells,
promoting liver fibrosis. Gastroenterology. 159:273–288. 2020.
View Article : Google Scholar
|
|
75
|
Trivedi P, Wang S and Friedman SL: The
power of plasticity-metabolic regulation of hepatic stellate cells.
Cell Metab. 33:242–257. 2021. View Article : Google Scholar
|
|
76
|
Shangguan X, He J, Ma Z, Zhang W, Ji Y,
Shen K, Yue Z, Li W, Xin Z, Zheng Q, et al: SUMOylation controls
the binding of hexokinase 2 to mitochondria and protects against
prostate cancer tumorigenesis. Nat Commun. 12:18122021. View Article : Google Scholar
|
|
77
|
Riazi K, Azhari H, Charette JH, Underwood
FE, King JA, Afshar EE, Swain MG, Congly SE, Kaplan GG and Shaheen
AA: The prevalence and incidence of NAFLD worldwide: A systematic
review and meta-analysis. Lancet Gastroenterol Hepatol. 7:851–861.
2022. View Article : Google Scholar
|
|
78
|
Estes C, Razavi H, Loomba R, Younossi Z
and Sanyal AJ: Modeling the epidemic of nonalcoholic fatty liver
disease demonstrates an exponential increase in burden of disease.
Hepatology. 67:123–133. 2018. View Article : Google Scholar
|
|
79
|
Afonso MB, Rodrigues PM, Simão AL and
Castro RE: Circulating microRNAs as potential biomarkers in
non-alcoholic fatty liver disease and hepatocellular carcinoma. J
Clin Med. 5:302016. View Article : Google Scholar
|
|
80
|
Jeppesen JB, Mortensen C, Bendtsen F and
Møller S: Lactate metabolism in chronic liver disease. Scand J Clin
Lab Invest. 73:293–299. 2013. View Article : Google Scholar
|
|
81
|
Ha TS, Shin TG, Jo IJ, Hwang SY, Chung CR,
Suh GY and Jeon K: Lactate clearance and mortality in septic
patients with hepatic dysfunction. Am J Emerg Med. 34:1011–1015.
2016. View Article : Google Scholar
|
|
82
|
Li J, Wang T, Xia J, Yao W and Huang F:
Enzymatic and nonenzymatic protein acetylations control glycolysis
process in liver diseases. FASEB J. 33:11640–11654. 2019.
View Article : Google Scholar
|
|
83
|
Vazquez JH, Kennon-McGill S, Byrum SD,
Mackintosh SG, Jaeschke H, Williams DK, Lee WM, Dranoff JA and
McGill MR; Acute Liver Failure Study Group: Proteomics indicates
lactate dehydrogenase is prognostic in acetaminophen-induced acute
liver failure patients and reveals altered signaling pathways.
Toxicol Sci. 187:25–34. 2022. View Article : Google Scholar
|
|
84
|
Bernal W, Donaldson N, Wyncoll D and
Wendon J: Blood lactate as an early predictor of outcome in
paracetamol-induced acute liver failure: A cohort study. Lancet.
359:558–563. 2002. View Article : Google Scholar
|
|
85
|
Macquillan GC, Seyam MS, Nightingale P,
Neuberger JM and Murphy N: Blood lactate but not serum phosphate
levels can predict patient outcome in fulminant hepatic failure.
Liver Transpl. 11:1073–1079. 2005. View Article : Google Scholar
|
|
86
|
Dabos KJ, Newsome PN, Parkinson JA,
Davidson JS, Sadler IH, Plevris JN and Hayes PC: A biochemical
prognostic model of outcome in paracetamol-induced acute liver
injury. Transplantation. 80:1712–1717. 2005. View Article : Google Scholar
|
|
87
|
Schmidt LE and Larsen FS: Prognostic
implications of hyperlactatemia, multiple organ failure, and
systemic inflammatory response syndrome in patients with
acetaminophen-induced acute liver failure. Crit Care Med.
34:337–343. 2006. View Article : Google Scholar
|
|
88
|
Cholongitas EB, Betrossian A, Leandro G,
Shaw S, Patch D and Burroughs AK: King's criteria, APACHE II, and
SOFA scores in acute liver failure. Hepatology. 43:881author reply
882. 2006. View Article : Google Scholar
|
|
89
|
Gow PJ, Warrilow S, Lontos S, Lubel J,
Wongseelashote S, MacQuillan GC, Jones RM, Bellomo R and Angus PW:
Time to review the selection criteria for transplantation in
paracetamol-induced fulminant hepatic failure? Liver Transpl.
13:1762–1763. 2007. View Article : Google Scholar
|
|
90
|
Agrawal T, Maiwall R, Rajan V, Bajpai M,
Jagdish RK, Sarin SK and Trehanpati N: Higher circulating natural
killer cells and lower lactate levels at admission predict
spontaneous survival in non-acetaminophen induced acute liver
failure. Clin Immunol. 231:1088292021. View Article : Google Scholar
|
|
91
|
Karvellas CJ, Tillman H, Leung AA, Lee WM,
Schilsky ML, Hameed B, Stravitz RT, McGuire BM and Fix OK; United
States Acute Liver Failure Study Group: Acute liver injury and
acute liver failure from mushroom poisoning in North America. Liver
Int. 36:1043–1050. 2016. View Article : Google Scholar
|
|
92
|
Feldman AG, Sokol RJ, Hardison RM, Alonso
EM, Squires RH and Narkewicz MR; Pediatric Acute Liver Failure
Study Group: Lactate and Lactate: Pyruvate ratio in the diagnosis
and outcomes of pediatric acute liver failure. J Pediatr.
182:217–222.e3. 2017. View Article : Google Scholar
|
|
93
|
Haidar MK, Morton N, Roederer T, Mayronne
S, Bawo L, Kerkula J, Porten K and Baud FJ: Evaluating lactate
prognostic value in children suspected of acetaminophen-induced
liver failure in Liberia. Pediatr Res. 88:605–611. 2020. View Article : Google Scholar
|
|
94
|
Schmidt LE and Larsen FS: Is lactate
concentration of major value in determining the prognosis in
patients with acute liver failure? Hardly. J Hepatol. 53:211–212.
2010. View Article : Google Scholar
|
|
95
|
Bernal W: Lactate is important in
determining prognosis in acute liver failure. J Hepatol.
53:209–210. 2010. View Article : Google Scholar
|
|
96
|
Niederwieser T, Braunwarth E, Dasari BVM,
Pufal K, Szatmary P, Hackl H, Haselmann C, Connolly CE, Cardini B,
Öfner D, et al: Early postoperative arterial lactate concentrations
to stratify risk of post-hepatectomy liver failure. Br J Surg.
108:1360–1370. 2021. View Article : Google Scholar
|
|
97
|
Popescu M, Dima S, Brasoveanu V, Tudor A,
Simionescu M and Tomescu D: High perioperative lactate levels and
decreased lactate clearance are associated with increased incidence
of posthepatectomy liver failure. Hepatobiliary Pancreat Dis Int.
20:592–594. 2021. View Article : Google Scholar
|
|
98
|
Gao F, Huang XL, Cai MX, Lin MT, Wang BF,
Wu W and Huang ZM: Prognostic value of serum lactate kinetics in
critically ill patients with cirrhosis and acute-on-chronic liver
failure: A multicenter study. Aging (Albany NY). 11:4446–4462.
2019. View Article : Google Scholar
|
|
99
|
Cardoso FS, Abraldes JG, Sy E, Ronco JJ,
Bagulho L, Mcphail MJ and Karvellas CJ: Lactate and number of organ
failures predict intensive care unit mortality in patients with
acute-on-chronic liver failure. Liver Int. 39:1271–1280. 2019.
View Article : Google Scholar
|
|
100
|
Kotoh K, Kato M, Kohjima M, Tanaka M,
Miyazaki M, Nakamura K, Enjoji M, Nakamuta M and Takayanagi R:
Lactate dehydrogenase production in hepatocytes is increased at an
early stage of acute liver failure. Exp Ther Med. 2:195–199. 2011.
View Article : Google Scholar
|
|
101
|
Cassidy WM and Reynolds TB: Serum lactic
dehydrogenase in the differential diagnosis of acute hepatocellular
injury. J Clin Gastroenterol. 19:118–121. 1994. View Article : Google Scholar
|
|
102
|
Krispin I, Mahamid M, Goldin E and Fteiha
B: Elevated lactate/albumin ratio as a novel predictor of
in-hospital mortality in hospitalized cirrhotics. Ann Hepatol.
28:1008972023. View Article : Google Scholar
|
|
103
|
Nie Y, Liu LX, Chen T, Zhang Y and Zhu X:
Serum lactate level predicts 6-months mortality in patients with
hepatitis B virus-related decompensated cirrhosis: A retrospective
study. Epidemiol Infect. 149:e262021. View Article : Google Scholar
|
|
104
|
Forner A, Reig M and Bruix J:
Hepatocellular carcinoma. Lancet. 391:1301–1314. 2018. View Article : Google Scholar
|
|
105
|
Chen Z, Xie H, Hu M, Huang T, Hu Y, Sang N
and Zhao Y: Recent progress in treatment of hepatocellular
carcinoma. Am J Cancer Res. 10:2993–3036. 2020.
|
|
106
|
Hanahan D and Coussens LM: Accessories to
the crime: Functions of cells recruited to the tumor
microenvironment. Cancer Cell. 21:309–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Lee DC, Sohn HA, Park ZY, Oh S, Kang YK,
Lee KM, Kang M, Jang YJ, Yang SJ, Hong YK, et al: A lactate-induced
response to hypoxia. Cell. 161:595–609. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Deng H, Kan A, Lyu N, He M, Huang X, Qiao
S, Li S, Lu W, Xie Q, Chen H, et al: Tumor-derived lactate inhibit
the efficacy of lenvatinib through regulating PD-L1 expression on
neutrophil in hepatocellular carcinoma. J Immunother Cancer.
9:e0023052021. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Xu Y, Hao X, Ren Y, Xu Q, Liu X, Song S
and Wang Y: Research progress of abnormal lactate metabolism and
lactate modification in immunotherapy of hepatocellular carcinoma.
Front Oncol. 12:10634232022. View Article : Google Scholar
|
|
110
|
Tu CE, Hu Y, Zhou P, Guo X, Gu C, Zhang Y,
Li A and Liu S: Lactate and TGF-β antagonistically regulate
inflammasome activation in the tumor microenvironment. J Cell
Physiol. 236:4528–4537. 2021. View Article : Google Scholar
|
|
111
|
Apicella M, Giannoni E, Fiore S, Ferrari
KJ, Fernández-Pérez D, Isella C, Granchi C, Minutolo F, Sottile A,
Comoglio PM, et al: Increased lactate secretion by cancer cells
sustains non-cell-autonomous adaptive resistance to MET and EGFR
targeted therapies. Cell Metab. 28:848–65 e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Huang X, Gan G, Wang X, Xu T and Xie W:
The HGF-MET axis coordinates liver cancer metabolism and autophagy
for chemotherapeutic resistance. Autophagy. 15:1258–1279. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Shan T, Chen S, Chen X, Wu T, Yang Y, Li
S, Ma J, Zhao J, Lin W, Li W, et al: M2-TAM subsets altered by
lactic acid promote T-cell apoptosis through the PD-L1/PD-1
pathway. Oncol Rep. 44:1885–1894. 2020.PubMed/NCBI
|
|
114
|
Lu LG, Zhou ZL, Wang XY, Liu BY, Lu JY,
Liu S, Zhang GB, Zhan MX and Chen Y: PD-L1 blockade liberates
intrinsic antitumourigenic properties of glycolytic macrophages in
hepatocellular carcinoma. Gut. 71:2551–2560. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Stone SC, Rossetti RAM, Alvarez KLF,
Carvalho JP, Margarido PFR, Baracat EC, Tacla M, Boccardo E,
Yokochi K, Lorenzi NP and Lepique AP: Lactate secreted by cervical
cancer cells modulates macrophage phenotype. J Leukoc Biol.
105:1041–1054. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Chu X, Di C, Chang P, Li L, Feng Z, Xiao
S, Yan X, Xu X, Li H, Qi R, et al: Lactylated histone H3K18 as a
potential biomarker for the diagnosis and predicting the severity
of septic shock. Front Immunol. 12:7866662022. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Suk FM, Wu CY, Fang CC, Chen TL and Liao
YJ: β-HB treatment reverses sorafenib resistance by shifting
glycolysis-lactate metabolism in HCC. Biomed Pharmacother.
166:1152932023. View Article : Google Scholar
|
|
118
|
Baltazar F, Afonso J, Costa M and Granja
S: Lactate beyond a waste metabolite: Metabolic affairs and
signaling in malignancy. Front Oncol. 10:2312020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Sheng SL, Liu JJ, Dai YH, Sun XG, Xiong XP
and Huang G: Knockdown of lactate dehydrogenase A suppresses tumor
growth and metastasis of human hepatocellular carcinoma. FEBS J.
279:3898–3910. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Fukao T, Lopaschuk GD and Mitchell GA:
Pathways and control of ketone body metabolism: On the fringe of
lipid biochemistry. Prostaglandins Leukot Essent Fatty Acids.
70:243–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Li X, Lu P, Li B, Yang R, Chu Y, Zhang Z,
Wan H, Niu C, Wang C and Luo K: Sensitization of hepatocellular
carcinoma cells to irradiation by miR-34a through targeting lactate
dehydrogenase-A. Mol Med Rep. 13:3661–3667. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhang K, Mu L, Ding MC, Xu R, Ding ZJ and
Liang J: NFκB mediated elevation of KCNJ11 promotes tumor
progression of hepatocellular carcinoma through interaction of
lactate dehydrogenase A. Biochem Biophys Res Commun. 495:246–253.
2018. View Article : Google Scholar
|
|
123
|
Serra M, Di Matteo M, Serneels J, Pal R,
Cafarello ST, Lanza M, Sanchez-Martin C, Evert M, Castegna A,
Calvisi DF, et al: Deletion of lactate dehydrogenase-a impairs
oncogene-induced mouse hepatocellular carcinoma development. Cell
Mol Gastroenterol Hepatol. 14:609–624. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Cui W, Lv W, Qu Y, Ma R, Wang YW, Xu YJ,
Wu D and Chen X: Discovery of
2-((3-cyanopyridin-2-yl)thio)acetamides as human lactate
dehydrogenase A inhibitors to reduce the growth of MG-63
osteosarcoma cells: Virtual screening and biological validation.
Bioorg Med Chem Lett. 26:3984–3987. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Le A, Cooper CR, Gouw AM, Dinavahi R,
Maitra A, Deck LM, Royer RE, Vander Jagt DL, Semenza GL and Dang
CV: Inhibition of lactate dehydrogenase A induces oxidative stress
and inhibits tumor progression. Proc Natl Acad Sci USA.
107:2037–2042. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Chen CY, Feng Y, Chen JY and Deng H:
Identification of a potent inhibitor targeting human lactate
dehydrogenase A and its metabolic modulation for cancer cell line.
Bioorg Med Chem Lett. 26:72–75. 2016. View Article : Google Scholar
|
|
127
|
McDonald B, Zucoloto AZ, Yu IL, Burkhard
R, Brown K, Geuking MB and McCoy KD: Programing of an intravascular
immune firewall by the gut microbiota protects against pathogen
dissemination during infection. Cell Host Microbe. 28:660–668.e4.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Han S, Bao X, Zou Y, Wang L, Li Y, Yang L,
Liao A, Zhang X, Jiang X, Liang D, et al: d-lactate modulates M2
tumor-associated macrophages and remodels immunosuppressive tumor
microenvironment for hepatocellular carcinoma. Sci Adv.
9:eadg26972023. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Chen Y, Bei J, Chen M, Cai W, Zhou Z, Cai
M, Huang W, Lin L, Guo Y, Liu M, et al: Intratumoral lactate
depletion based on injectable nanoparticles-hydrogel composite
system synergizes with immunotherapy against postablative
hepatocellular carcinoma recurrence. Adv Healthc Mater.
13:e23030312024. View Article : Google Scholar
|
|
130
|
Kubackova K, Bortlicek Z, Pavlik T,
Melichar B, Linke Z, Pokorna P, Vyzula R, Prausova J and Buchler T;
Czech Renal Cancer Cooperative Group: Prognostic factors in renal
cell carcinoma patients treated with sorafenib: Results from the
Czech registry. Target Oncol. 10:385–392. 2015. View Article : Google Scholar
|
|
131
|
Scartozzi M, Giampieri R, Maccaroni E, Del
Prete M, Faloppi L, Bianconi M, Galizia E, Loretelli C, Belvederesi
L, Bittoni A and Cascinu S: Pre-treatment lactate dehydrogenase
levels as predictor of efficacy of first-line bevacizumab-based
therapy in metastatic colorectal cancer patients. Br J Cancer.
106:799–804. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Hermes A, Gatzemeier U, Waschki B and Reck
M: Lactate dehydrogenase as prognostic factor in limited and
extensive disease stage small cell lung cancer-a retrospective
single institution analysis. Respir Med. 104:1937–1942. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Faloppi L, Scartozzi M, Bianconi M,
Svegliati Baroni G, Toniutto P, Giampieri R, Del Prete M, De
Minicis S, Bitetto D, Loretelli C, et al: The role of LDH serum
levels in predicting global outcome in HCC patients treated with
sorafenib: implications for clinical management. BMC Cancer.
14:1102014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Sacco R, Mismas V, Granito A, Musettini G,
Masi G, Caparello C, Vivaldi C, Felder M, Bresci G and Fornaro L;
Italian Liver Cancer (IT.LI.CA) group: Correlation between LDH
levels and response to sorafenib in HCC patients: An analysis of
the ITA. LI.CA database. Int J Biol Markers. 30:e65–e72. 2015.
View Article : Google Scholar
|
|
135
|
Yada M, Miyazaki M, Motomura K, Masumoto
A, Nakamuta M, Kohjima M, Sugimoto R, Aratake Y, Higashi N,
Morizono S, et al: The prognostic role of lactate dehydrogenase
serum levels in patients with hepatocellular carcinoma who are
treated with sorafenib: the influence of liver fibrosis. J
Gastrointest Oncol. 7:615–623. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Wu SJ, Lin YX, Ye H, Xiong XZ, Li FY and
Cheng NS: Prognostic value of alkaline phosphatase, gamma-glutamyl
transpeptidase and lactate dehydrogenase in hepatocellular
carcinoma patients treated with liver resection. Int J Surg. 36(Pt
A): 143–151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Gan Y, Gao F, Du B, Liu Y, Xue Q and Fu J:
Effects of preoperative serum lactate dehydrogenase levels on
long-term prognosis in elderly patients with hepatocellular
carcinoma undergoing transcatheter arterial chemoembolization.
Front Surg. 9:9821142022. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhuang G, Xie Y, Hong J, Lin S, Chen T and
Fang W: Arterial chemoembolization for patients with hepatocellular
carcinoma and elevated lactate dehydrogenase is associated with low
survival: A cohort study. Infect Agent Cancer. 17:312022.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Su K, Huang W, Li X, Xu K, Gu T, Liu Y,
Song J, Qian K, Xu Y, Zeng H, et al: Evaluation of lactate
dehydrogenase and alkaline phosphatase as predictive biomarkers in
the prognosis of hepatocellular carcinoma and development of a new
nomogram. J Hepatocell Carcinoma. 10:69–79. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Cui Z, Li Y, Gao Y, Kong L, Lin Y and Chen
Y: Cancer-testis antigen lactate dehydrogenase C4 in hepatocellular
carcinoma: A promising biomarker for early diagnosis, efficacy
evaluation and prognosis prediction. Aging (Albany NY).
12:19455–19467. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Jin J, Bai L, Wang D, Ding W, Cao Z, Yan
P, Li Y, Xi L, Wang Y, Zheng X, et al: SIRT3-dependent
delactylation of cyclin E2 prevents hepatocellular carcinoma
growth. EMBO Rep. 24:e560522023. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Cheng Z, Huang H, Li M, Liang X, Tan Y and
Chen Y: Lactylation-Related gene signature effectively predicts
prognosis and treatment responsiveness in hepatocellular carcinoma.
Pharmaceuticals (Basel). 16:6442023. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Doherty JR and Cleveland JL: Targeting
lactate metabolism for cancer therapeutics. J Clin Invest.
123:3685–3692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Kooshki L, Mahdavi P, Fakhri S, Akkol EK
and Khan H: Targeting lactate metabolism and glycolytic pathways in
the tumor microenvironment by natural products: A promising
strategy in combating cancer. BioFactors. 48:359–383. 2022.
View Article : Google Scholar
|
|
145
|
Li Y, Zhou Y, Xia S, Chen L, Yang T, Zhao
D, Zhang Z, Shao J, Xu X, Zhang F and Zheng S: Blockade of
KLF5/LDH-A feedback loop contributes to Curcumol inhibition of
sinusoidal endothelial cell glycolysis and mitigation of liver
fibrosis. Phytomedicine. 114:1547592023. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Wang F, Chen L, Kong D, Zhang X, Xia S,
Liang B, Li Y, Zhou Y, Zhang Z, Shao J, et al: Canonical Wnt
signaling promotes HSC glycolysis and liver fibrosis through an
LDH-A/HIF-1α transcriptional complex. Hepatolog. 79:606–623. 2024.
View Article : Google Scholar
|
|
147
|
Ban D, Hua S, Zhang W, Shen C, Miao X and
Liu W: Costunolide reduces glycolysis-associated activation of
hepatic stellate cells via inhibition of hexokinase-2. Cell Mol
Biol Lett. 24:522019. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Reyes R, Wani NA, Ghoshal K, Jacob ST and
Motiwala T: Sorafenib and 2-deoxyglucose synergistically inhibit
proliferation of both sorafenib-sensitive and -resistant HCC cells
by inhibiting ATP production. Gene Expr. 17:129–140. 2017.
View Article : Google Scholar :
|
|
149
|
Tomizawa M, Shinozaki F, Motoyoshi Y,
Sugiyama T, Yamamoto S and Ishige N: 2-Deoxyglucose and sorafenib
synergistically suppress the proliferation and motility of
hepatocellular carcinoma cells. Oncol Lett. 13:800–804. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Raez LE, Papadopoulos K, Ricart AD,
Chiorean EG, Dipaola RS, Stein MN, Rocha Lima CM, Schlesselman JJ,
Tolba K, Langmuir VK, et al: A phase I dose-escalation trial of
2-deoxy-D-glucose alone or combined with docetaxel in patients with
advanced solid tumors. Cancer Chemother Pharmacol. 71:523–530.
2013. View Article : Google Scholar
|
|
151
|
Stein M, Lin H, Jeyamohan C, Dvorzhinski
D, Gounder M, Bray K, Eddy S, Goodin S, White E and Dipaola RS:
Targeting tumor metabolism with 2-deoxyglucose in patients with
castrate-resistant prostate cancer and advanced malignancies.
Prostate. 70:1388–1394. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Sasaki K, Nishina S, Yamauchi A, Fukuda K,
Hara Y, Yamamura M, Egashira K and Hino K: Nanoparticle-Mediated
Delivery of 2-Deoxy-D-Glucose induces antitumor immunity and
cytotoxicity in liver tumors in mice. Cell Mol Gastroenterol
Hepatol. 11:739–762. 2021. View Article : Google Scholar :
|
|
153
|
Tomizawa M, Shinozaki F, Motoyoshi Y,
Sugiyama T, Yamamoto S and Ishige N: Suppressive effects of
3-bromopyruvate on the proliferation and the motility of
hepatocellular carcinoma cells. Oncol Rep. 35:59–63. 2016.
View Article : Google Scholar
|
|
154
|
Yoo JJ, Yu SJ, Na J, Kim K, Cho YY, Lee
YB, Cho EJ, Lee JH, Kim YJ, Youn H and Yoon JH: Hexokinase-II
inhibition synergistically augments the anti-tumor efficacy of
sorafenib in hepatocellular carcinoma. Int J Mol Sci. 20:12922019.
View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Sun X, Sun G, Huang Y, Hao Y, Tang X,
Zhang N, Zhao L, Zhong R and Peng Y: 3-Bromopyruvate regulates the
status of glycolysis and BCNU sensitivity in human hepatocellular
carcinoma cells. Biochem Pharmacol. 177:1139882020. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Wu H, Pan L, Gao C, Xu H, Li Y, Zhang L,
Ma L, Meng L, Sun X and Qin H: Quercetin inhibits the proliferation
of glycolysis-addicted HCC cells by reducing hexokinase 2 and
Akt-mTOR pathway. Molecules. 24:19932019. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Xu Q, Dou C, Liu X, Yang L, Ni C, Wang J,
Guo Y, Yang W, Tong X and Huang D: Oviductus ranae protein
hydrolysate (ORPH) inhibits the growth, metastasis and glycolysis
of HCC by targeting miR-491-5p/PKM2 axis. Biomed Pharmacother.
107:1692–1704. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Ferriero R, Nusco E, De Cegli R, Carissimo
A, Manco G and Brunetti-Pierri N: Pyruvate dehydrogenase complex
and lactate dehydrogenase are targets for therapy of acute liver
failure. J Hepatol. 69:325–335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Billiard J, Dennison JB, Briand J, Annan
RS, Chai D, Colón M, Dodson CS, Gilbert SA, Greshock J, Jing J, et
al: Quinoline 3-sulfonamides inhibit lactate dehydrogenase A and
reverse aerobic glycolysis in cancer cells. Cancer Metab. 1:192013.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Manerba M, Di Ianni L, Govoni M, Roberti
M, Recanatini M and Di Stefano G: LDH inhibition impacts on heat
shock response and induces senescence of hepatocellular carcinoma
cells. Eur J Pharm Sci. 105:91–98. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Payen VL, Mina E, Van Hée VF, Porporato PE
and Sonveaux P: Monocarboxylate transporters in cancer. Mol Metab.
33:48–66. 2020. View Article : Google Scholar :
|
|
162
|
Hadjihambi A, Konstantinou C, Klohs J,
Monsorno K, Le Guennec A, Donnelly C, Cox IJ, Kusumbe A, Hosford
PS, Soffientini U, et al: Partial MCT1 invalidation protects
against diet-induced non-alcoholic fatty liver disease and the
associated brain dysfunction. J Hepatol. 78:180–190. 2023.
View Article : Google Scholar
|
|
163
|
Jeon JY, Lee M, Whang SH, Kim JW, Cho A
and Yun M: Regulation of acetate utilization by monocarboxylate
transporter 1 (MCT1) in hepatocellular carcinoma (HCC). Oncol Res.
26:71–81. 2018. View Article : Google Scholar
|
|
164
|
Fang Y, Liu W, Tang Z, Ji X, Zhou Y, Song
S, Tian M, Tao C, Huang R, Zhu G, et al: Monocarboxylate
transporter 4 inhibition potentiates hepatocellular carcinoma
immunotherapy through enhancing T cell infiltration and immune
attack. Hepatology. 77:109–123. 2023. View Article : Google Scholar
|
|
165
|
Xu H, Li L, Wang S, Wang Z, Qu L, Wang C
and Xu K: Royal jelly acid suppresses hepatocellular carcinoma
tumorigenicity by inhibiting H3 histone lactylation at H3K9la and
H3K14la sites. Phytomedicine. 118:1549402023. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Zhou F, Shang W, Yu X and Tian J:
Glypican-3: A promising biomarker for hepatocellular carcinoma
diagnosis and treatment. Med Res Rev. 38:741–767. 2018. View Article : Google Scholar
|
|
167
|
Yao G and Yang Z: Glypican-3 knockdown
inhibits the cell growth, stemness, and glycolysis development of
hepatocellular carcinoma cells under hypoxic microenvironment
through lactylation. Arch Physiol Biochem. May 2–2023.Online ahead
of print. View Article : Google Scholar : PubMed/NCBI
|