Globally, stress is a prevalent issue. Mental health
obstacles, such as social isolation, fear, loneliness,
unemployment, financial instability and loss of family or friends,
all contribute to psychological stress (1). Mental stress causes physical health
problems, including weakened immune function, hypertension,
gastrointestinal issues, cardiovascular disease, sleep disturbance
and ovarian dysfunction (2,3).
Stress has been widely acknowledged as a key factor that impacts
the advancement and onset of ovarian damage (2,4).
To the best of our knowledge, however, no consistent conclusions
have been drawn on the impact of stress on ovarian function based
on epidemiological data, clinical trials and experimental studies
(2,4,5).
There is a growing body of evidence indicating that
psychological stress leads to ovarian dysfunction (2,6,7).
Notably, for patients undergoing in vitro fertilization
(IVF) treatment, higher anxiety leads to lower clinical pregnancy
(8) and fertilization rates
(9), decreased embryo quality
(10) and fewer oocytes
retrieved (11). A prospective
study of 274 patients showed that stress reduced the likelihood of
conception during the fertile window each day (12) and participation in stress
reduction interventions improves IVF outcomes (11). Mental stress can also affect
ovarian reserve, leading to decreased levels of antral follicle
count (AFC) (13) and
anti-Müllerian hormone (AMH) (14). Moreover, mental stress is also
involved in an increased risk of premature ovarian insufficiency
(POI) (15), functional
hypothalamic amenorrhea (FHA) (16) and polycystic ovary syndrome
(PCOS) (17). Furthermore,
ovarian insufficiency, such as alteration in female sex hormones,
also increases the incidence of depression (18). Similarly, ovarian dysfunction is
observed in mental stress models; both chronic unpredictable stress
(CUS) (19) and chronic
restraint stress (CRS) (20)
models lead to depression-like behavior and decreased fertility.
Stress can also cause general neuroendocrine system changes, which
decrease oocyte quality (21).
In recent years, there has been a growing recognition that stress
is an emerging risk factor for metabolic disorder (22,23). Metabolic disturbances triggered
by stress could lead to conditions such as obesity, type 2 diabetes
mellitus (T2DM) and ovary damage (2).
The present study aimed to investigate the
association between psychological stress and ovarian function based
on existing clinical evidence. Research on psychological-associated
stress and female reproductive endocrine function from a
neuroendocrine-metabolic (NEM) perspective and the underlying
molecular mechanisms are summarized. Additionally, strategies to
safeguard ovarian function from psychological stress are explored.
The present review may provide new perspectives and directions for
research and treatment of ovarian dysfunction and infertility.
Research and review articles focusing on the role of
stress on ovarian function were included. By contrast, research and
review articles focusing on the effect of disorders associated with
ovarian dysfunction on mental stress, including communications,
case reports, guidelines, comments and protocols, were excluded
from the present review. Articles written not in English or without
available full text were also excluded.
The title and abstract of every article were checked
by two authors (YH and WW). Disagreements were resolved by a third
author (FF). A total of 7,623 articles were found by the initial
search; 7,316 papers were removed after checking the title and
abstract. Next, 307 papers were filtered for full text, of which
183 met the inclusion criteria; 36 were population studies on the
effect of stress on ovarian function, 71 were animal models and 26
were psychological and 50 were metabolic intervention studies.
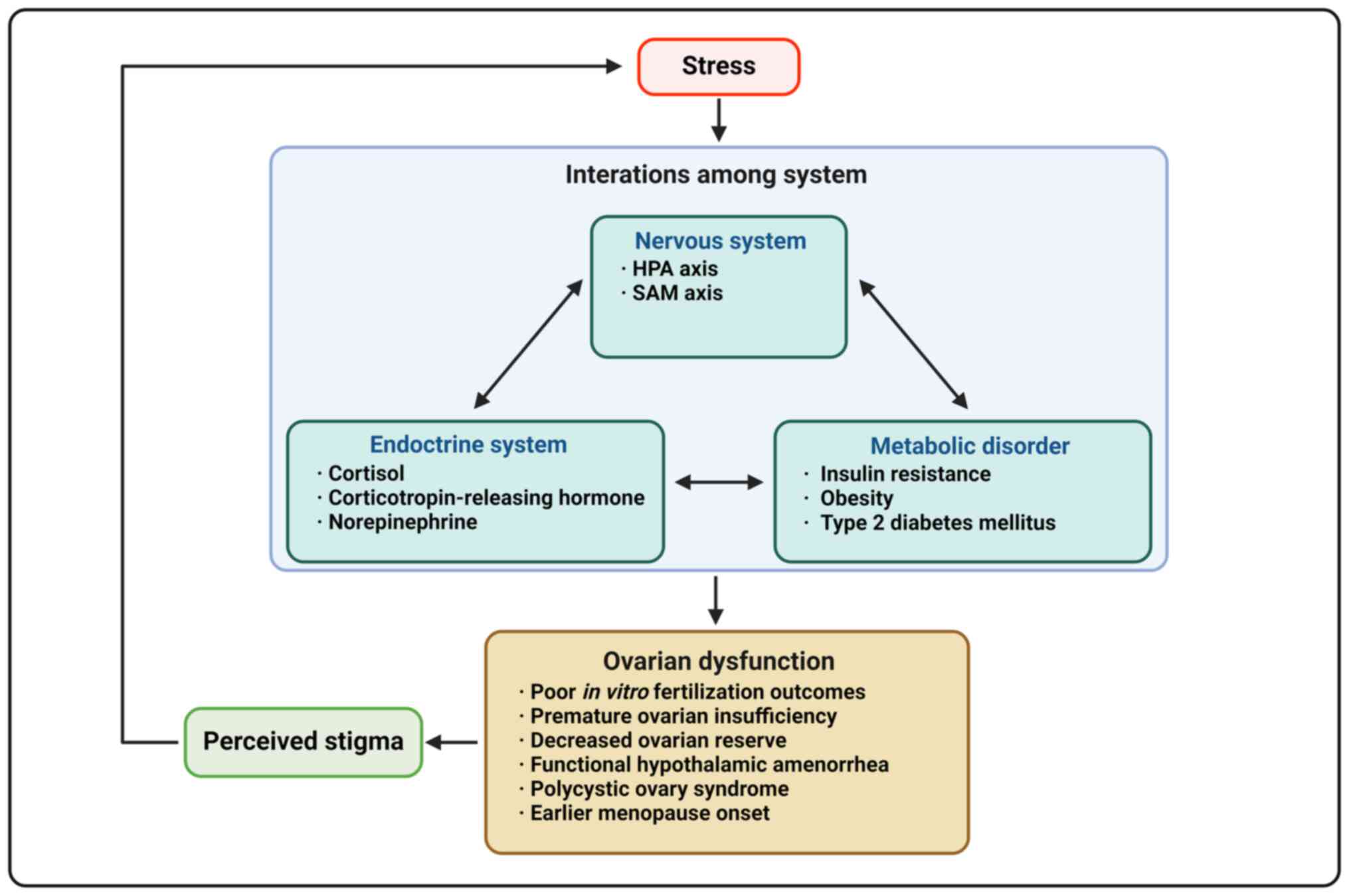
It has been hypothesized that stress may affect
ovarian function, however it is unclear whether ovarian dysfunction
precedes anxiety or stress precedes ovarian damage (5). Stress is commonly overlooked as a
cause of reproductive vulnerability (2). Activation of the stress response
system suppresses the hypothalamic-pituitary-ovarian (HPO) axis
(24,25). Daily perceived stress can lead to
a decrease in estrogen, progesterone and luteinizing hormone (LH),
as well as an increase in follicle-stimulating hormone (FSH) in
reproductive-aged patients (24). Patients with a history of
depression also had higher FSH and LH levels and lower estradiol
levels and those having more significant depressive symptoms were
twice as likely to experience earlier perimenopause transition than
those without such history (25). Additionally, female patients may
be more sensitive to lower levels of cortisol than male patients
due to increased limbic system activation following repeated
stressors (26). The
psychological distress caused by decreased ovarian function may
further exacerbate the association between psychological stress and
impaired ovarian function (Fig.
1; Table I).
In assisted reproductive medicine, one of the most
controversial areas is the underlying influence of psychological
factors on pregnancy outcomes. Infertility is a serious health
problem around the world and female infertility factors account for
at least 35% of all infertility cases (27). The risk of anxiety, depression
and distress is generally high in infertile patients (28,29). A 2004 study that interviewed 122
patients before their first infertility clinic visit utilizing a
structured psychiatric interview found that 40.2% had a psychiatric
disorder, including major depressive, generalized anxiety and
dysthymic disorder (30).
Volgsten et al (31)
reported that, of 413 infertile patients undergoing IVF treatment,
127 (30.8%) were diagnosed with psychiatric disorders, with major
depression being the most common. In a study involving 352 female
and 274 male patients attending infertility clinics in northern
California, 56.5% of patients exhibited notable symptoms of
depression and 76% showed notable symptoms of anxiety (32). Infertile patients have higher
Beck Depression Inventory (BDI) (14.94±12.90 vs. 8.95±10.49,
P<0.0001) and Spielberger Trait Anxiety Inventory (STAI-T) score
(48.76±10.96 vs. 41.18±11.26, P<0.0001), indicating a greater
experience of anxiety and depression compared with fertile
individuals (33).
Numerous studies have explored the link between
psychological symptoms before and during ART cycles and subsequent
pregnancy rates, yielding conflicting results: Certain studies
suggest that higher distress/anxiety/depression levels before and
during treatment are associated with lower pregnancy rates
(8,9,11,34-39), while others do not (10,40-50). These inconsistencies may be
attributed to challenges in accurately assessing stress levels in
female patients with infertility and heterogeneity of studies
(5,28). Future research should therefore
focus on developing more precise psychological assessment methods
and establishing consistent time points for data collection to
enhance quality of research findings.
POI is characterized by premature loss of ovarian
function and permanent cessation of menstruation before the age of
40 years (51). Demographic
studies confirm that psychological stress, such as emotional
trauma, major life events or chronic stress, may be as a potential
trigger for POI in certain cases (52,53). Individuals with POI report
continuous exposure to family and work stress and sleep problems,
suggesting that adverse life events may contribute to development
of POI (52).
Among mental disorders associated with POI,
depression is most closely associated to POI. A population study
involving 290 patients found that 43% of those with POI had a
history of depression, with 26% receiving a depression diagnosis
before being diagnosed with POI (15). Depression typically emerges
following signs of altered ovarian function but before POI
diagnosis (54), possibly due to
a more fragile stress system. Female patients are more likely than
male patients to develop depression and changes in estrogen levels
can increase susceptibility to this condition (18,55). Given the lack of prospective
studies on pre-diagnosis stress in patients with POI, more
conclusive evidence is needed to determine whether there is an
overlapping process or causal association between POI and
psychological stress.
Various indicators are used to assess ovarian
reserve (OR), with AFC and AMH being particularly promising
(51). A study on 979
premenopausal patients aged 25-45 years used AFC measured via
transvaginal ultrasound to evaluate OR; the finding indicated a
positive association between higher stress levels and AFC in
younger patients (β=0.545, P=0.005), and stress accelerates
age-related AFC decline (β=−0.036, P=0.031), suggesting women with
higher stress experienced faster AFC loss (56). Environmental stress might enhance
fertility temporarily by increasing the volume of growing
follicles, potentially depleting OR in the long-term (56). Additionally, a cross-sectional
study suggested that psychological stress accelerates AFC decline
(β=−0.070, P=0.021), implying that low positive affectivity may
aggravate the adverse impact of stress on AFC decline (57). Consistent findings across
populations reveal that higher levels of perceived stress are
linked to decreased AFC and AMH levels (13). A cross-sectional study on 576
infertile patients demonstrated a negative association between
salivary α amylase (SAA), a stress marker, and AMH levels
(r=−0.315, P<0.001; adjusted for age, r=−0.336, P<0.001)
(58). Another study noted
modest inverse associations between depression and low AMH among
patients aged 36-40 years (adjusted P=0.75, 95%CI, 0.52, 1.09) and
nulliparous patients (adjusted P=0.77, 95%CI, 0.59, 1.00) (14). Taken together, data from the
aforementioned population studies suggested that elevated levels of
perceived stress, as well as biomarkers of stress, are associated
with reduced OR.
FHA or anovulation is characterized by the absence
of menstruation and ovulation without an identifiable organic
cause. The main determinant of the disease is a combination of
psychosocial and metabolic stress (59). Predisposing factors include low
energy use, nutritional deficiencies, abnormal sleep, emotional and
chronic severe stress and overexertion or physical activity
(59,60). More severe depressive and anxious
symptoms contribute to onset of FHA (16,61-63). Patients with FHA often exhibit
dysfunctional attitudes, such as perfectionism, higher need for
approval, and struggle to cope with daily stressors (59). Cognitive behavioral therapy
accelerates the recovery of ovarian function in patients with FHA,
underscoring the role of mental stress in development of this
condition (59). As with POI,
more prospective studies are needed to explore the effect of
psychological stress on FHA.
PCOS is a prevalent endocrine disorder in patients
of reproductive age, characterized by systemic metabolic issue and
disruptions in neuroendocrine-immune function (64). Patients with PCOS are at a higher
risk of developing cardiovascular risk factors and associated
insulin resistance, dyslipidemia and diabetes (65). Patients with PCOS also face an
elevated likelihood of mood and anxiety disorders (17,66,67). There is a notable prevalence of
psychiatric issues among patients with PCOS, including depression
(26-40%), anxiety (11.6%) and binge-eating (23.3%) (17), indicating a potential role of
psychological factors in pathophysiology of PCOS. Psychological
aspects have been identified as a notable risk factor for PCOS
(68) and emotional distress
potentially stems from psychosocial and/or pathophysiological
origins (66). Moreover,
patients with PCOS may perceive symptom such as hirsutism and acne
or potential outcomes such as infertility and obesity as
stigmatizing, leading to distress and potentially exacerbating
progression of the disease (67). However, as meta-analysis has
indicated, few studies have focused on the effect of psychological
stress on disease and PCOS (17).
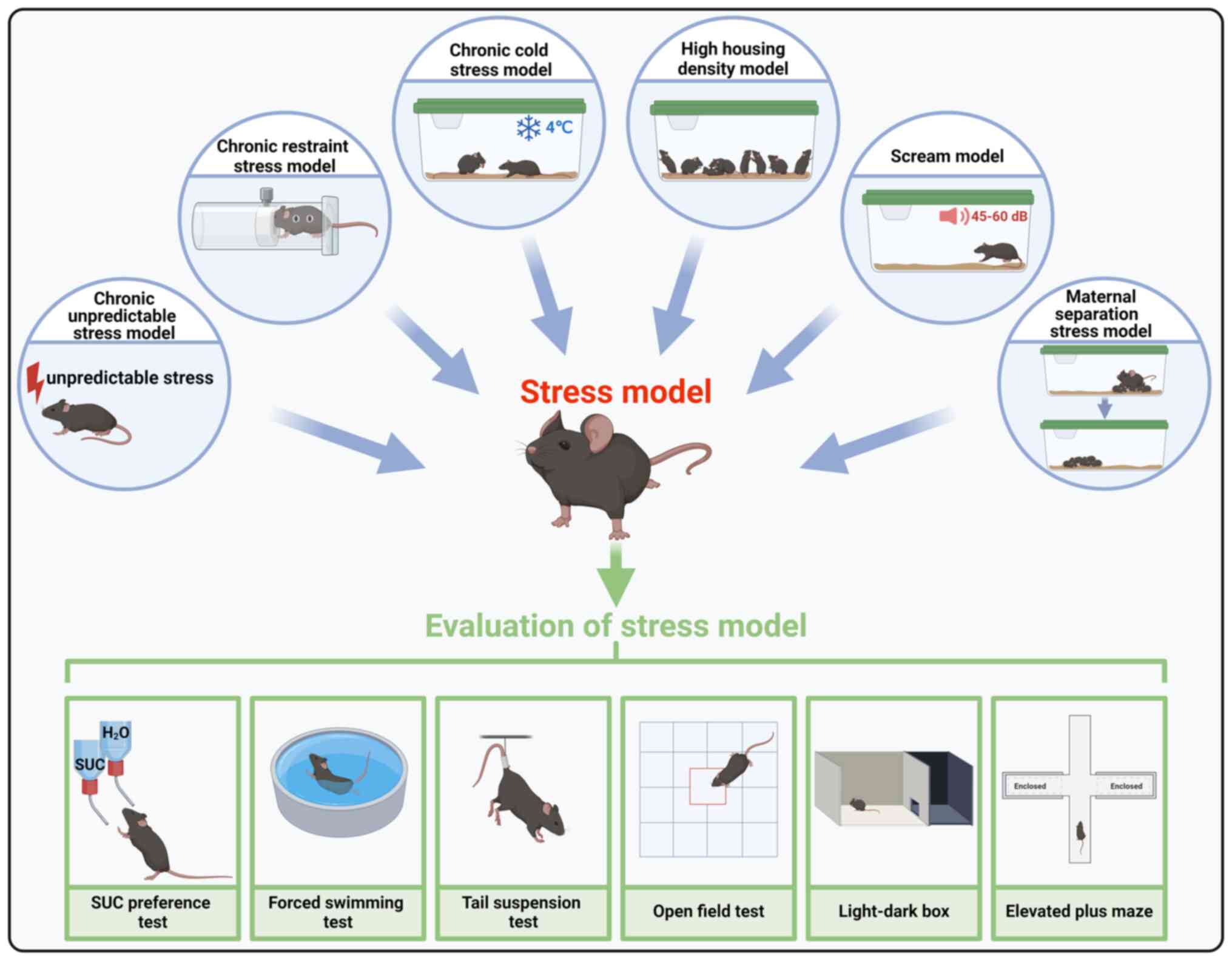
CUS is a well-established method where animals are
subjected to unpredictable stressors over a prolonged period to
induce emotional changes, which lead to lasting alterations in
animal behavior, gut microbiota and neurological function (72-74). The CUS stressors include
restraint stress, tail suspension and swimming, with the intensity
of stressors gradually increasing to prevent tolerance (19). Depression-like behaviors induced
by CUS are evaluated using tests such as the forced swimming test
(FST) and sucrose preference test (SPT) (75). CUS exposure has been shown to
disrupt the estrous cycle, decrease the number of primordial and
preantral follicles and corpus luteum, elevate serum FSH and
cortisol levels and reduce serum estradiol, AMH and
gonadotropin-releasing hormone (GnRH) levels, making it an ideal
animal model for studying impaired ovarian function (19,76-80).
CUS has been found to negatively impact oocyte
developmental potential, with transcriptome analysis revealing
disruption in expression of apoptotic genes and cell cycle
regulators (81). CUS decreases
brain-derived neurotrophic factor (BDNF) expression in antral
follicles, resulting in decreased number of retrieved oocytes and
rate of blastocyst formation, which could be rescued by exogenous
BDNF treatment (82). Another
study indicated that CUS significantly decreased the number of
retrieved oocytes, fertilization rate and number of embryos
(including high-quality embryos and blastocysts), which may be due
to different expression of heat shock protein 70 in the embryo
after CUS (83). Additionally,
supplementation with exogenous growth differentiation factor 9
(GDF9) rescues development of both secondary and antral follicles
in stressed mice (84).
Granulosa cells in the ovary of depression-like mice induced by CUS
exhibit increased apoptosis, possibly due to accumulation of
reactive oxygen species (ROS) and activation of the
mitogen-activated protein kinase (MAPK) pathway resulting from
downregulation of isocitrate dehydrogenase-1 expression (85,86). This suggests CUS can lead to
elevated ovarian oxidative stress and mitochondrial dyshomeostasis,
supported by increased expression of superoxide dismutase (SOD),
mitochondrial fusion protein optic atrophy 1, mitofusin 1 and
nucleus-encoded protein succinate dehydrogenase complex A and
decreased expression of mitochondrial fission protein 1 in ovary of
stressed mice (87,88).
Mice subjected to CRS display elevated serum
cortisol and corticotropin-releasing hormone (CRH) levels,
indicating activation of the hypothalamic-pituitary-adrenal (HPA)
axis (21,93-98). Additionally, CRS results in
decreased ovarian weight, disruption in estrous cycles, increased
ovarian fibrosis and overactivation of primordial follicles, which
may be attributed to the upregulation of Kit expression and
activation of the phosphatidylinositol 3-kinase (PI3K)/PTEN/Akt
pathway (21). CRS increases
oocyte aneuploidy, decreases the percentage of blastocysts and
number of cells/blastocyst and impaired corpora lutea function
during gestation in mice and rats, leading to decreased pregnancy
rates and litter size (99-101). The decline in fertility
associated with CRS may be linked to diminished oocyte quality,
with germinal vesicle breakdown (GVBD) being a key indicator of
oocyte quality (102).
Following CRS, female mice exhibit compromised oocyte competence,
which was characterized by an increase in spindles defects and
chromatin misalignment, decrease in GVBD percentage and prolonged
GVBD time, which may be due to abnormalities of the Fas/FasL system
or TNF-α signaling in oocytes and granulosa cells (93,97,98,103,104). Moreover, CRS decreases the
expression of cyclin B1 (CCNB1), a key regulator of M-phase/mature
promoting factor (93).
Treatment with MG132, an inhibitor of anaphase-promoting
complex/cyclosome, rescues prolonged GVBD time and elevates CCNB1
expression in oocytes of CRS-exposed mice (93). Furthermore, CRS elevates meiotic
arrest failure in oocytes by suppressing the cyclic AMP (cAMP)
signaling pathway, particularly via the downregulation of
G-protein-coupled receptor 3, natriuretic peptide C and natriuretic
peptide receptor 2, leading to accelerated oocyte loss and
premature ovarian aging (20).
Short-time restraint stress accelerates postovulatory oocyte aging
due to elevated cytoplasmic calcium levels and increased oxidative
stress (105).
Animals exposed to cold stress experience heightened
sympathetic activity and elevated levels of norepinephrine in the
ovary (106-108), while corticosterone and
gonadotropin (Gn) levels are unaffected (109,110). As a result, chronic cold stress
is commonly used in the examination of overactivation effects of
the sympathetic-adrenal-medullary (SAM) axis. The cold stress
regimen typically spans 21-28 days, involving activities such as
daily 15-min swims in 15°C water or 3-h exposures to 4°C
temperature (111,112).
The majority of studies show that prolonged exposure
to cold stress may result in extended estrous cycle, reduced level
of estrogen and progesterone, decreased corpus luteum count and
increased follicular atresia in rats (111,113-115). The observed hindered follicular
development may be attributed to increased nerve growth factor
levels in the ovary stemming from sympathetic activation, breakdown
of the gap junctions between oocyte and cumulus cells and
dysregulation of the NRF2/Connexin-43 (CX43)/steroidogenic acute
regulatory (StAR)/progesterone pathway in granulosa cells following
cold exposure (106). Cold
stress triggers the upregulation of endothelin-1 (ET-1) and
ET-receptor type A, alongside downregulation of ET-receptor type B
protein, thereby disrupting the ovarian vascular endothelin system,
leading to vasoconstriction and microcirculation issues in rats
(113). This compromised blood
flow may impede ovarian function by decreasing oxygen and nutrient
delivery to the ovary (116).
However, a previous study indicated that prepubertal rats that
experience chronic cold stress do not exhibit altered ovarian
architecture that affects the ovulation in adulthood (117). When animals were exposed to
both chronic restraint and cold water stress, the results were
different (118,119): A study showed that the number
of atretic follicles and the percentage of apoptotic granulosa
cells are notably increased following exposure to restraint stress,
while another study indicated that there were no effects on the
ovaries after exposure to cold stress (118,119).
A model of high housing density is conducted by
housing eight C57BL/6 mice in a 484 cm2 ventilated cage
which results in increased stocking density and elevated stress
hormones among the mice (120).
Mice housed in more densely packed conditions exhibit higher
corticosterone and lower AMH levels, alongside a great number of
atretic ovarian follicles (120). The scream model subjects mice
to noise levels between 45 and 60 dB for 6 h daily over 3 weeks,
leading to decreased levels of AMH and estradiol, loss of
primordial and preantral follicles, increased apoptosis in
granulosa cell and ultimately decreased litter sizes (121). An additional study indicated
that maternal separation stress could simulate early life stress
(122), with population studies
suggesting that early life stress may be associated with an earlier
onset of menarche and menopause (123-125). In a previous study, female
litters were separated from their mothers between postnatal days 2
and 16 for 6 h/day, with the home cages of the litters placed on a
warming pad set at 33-35°C to prevent undue stress (122). After 14 days, there was a
significant decrease in the activity of SOD, glutathione peroxidase
and catalase, accompanied by oocyte development disorder (122). A predatory stress model
suggests that the rate of blastocyst of formation, and the number
of cells/blastocyst significantly decreases in stressed mice
(126). Chronic immobilization
stress could disrupt the estrous cycle, decrease serum levels of
AMH, increase the serum levels of FSH and cortisol and reduce the
number of primordial follicles (127). In addition, cortisol injection
simulates the impact of stress-induced cortisol elevation on
ovarian function (128).
While animal models enhance the current
understanding of psychiatric disorders, they have certain
limitations; for example, animals are incapable of conveying
sadness, guilt or suicidal thoughts, which are symptoms mostly
confined to humans (129).
Following successful modeling, the experimental
animals display indications such as anhedonia and melancholy, in
addition to other signs of depression such as disturbed sleep,
alterations in weight and appetite, anxiousness and isolation from
social interaction, mirroring the symptoms observed in humans with
major depressive disorder (130).
To evaluate behaviors associated with hopelessness,
FST is employed. The animal is positioned in a cylinder filled with
water; if the subject recognizes that escape is unattainable, it
may exhibit depressive-like behavior characterized by limited
movement except that necessary to keep the snout above the water
level (134).
TST is a common method for assessing depressive-like
behavior, with the time spent motionless by the subject over a
6-min period recorded (135).
LDB box test exploits instinctual aversion to
well-lit areas and spontaneous exploration in response to mild
stressors, such as novel environments and light exposure (139). The delay in a mouse entry into
the dark section indicates an avoidance of the bright area due to
anxiety, while movements between compartments reflect exploratory
behavior.
EPM is extensively utilized for evaluating
anxiety-related behavior in laboratory animals, particularly
rodents (140-142). Similar to OFT and LDB, EPM
exploits the emotional adaptive responses that animals exhibit when
confronted with unavoidable aversive situations (143).
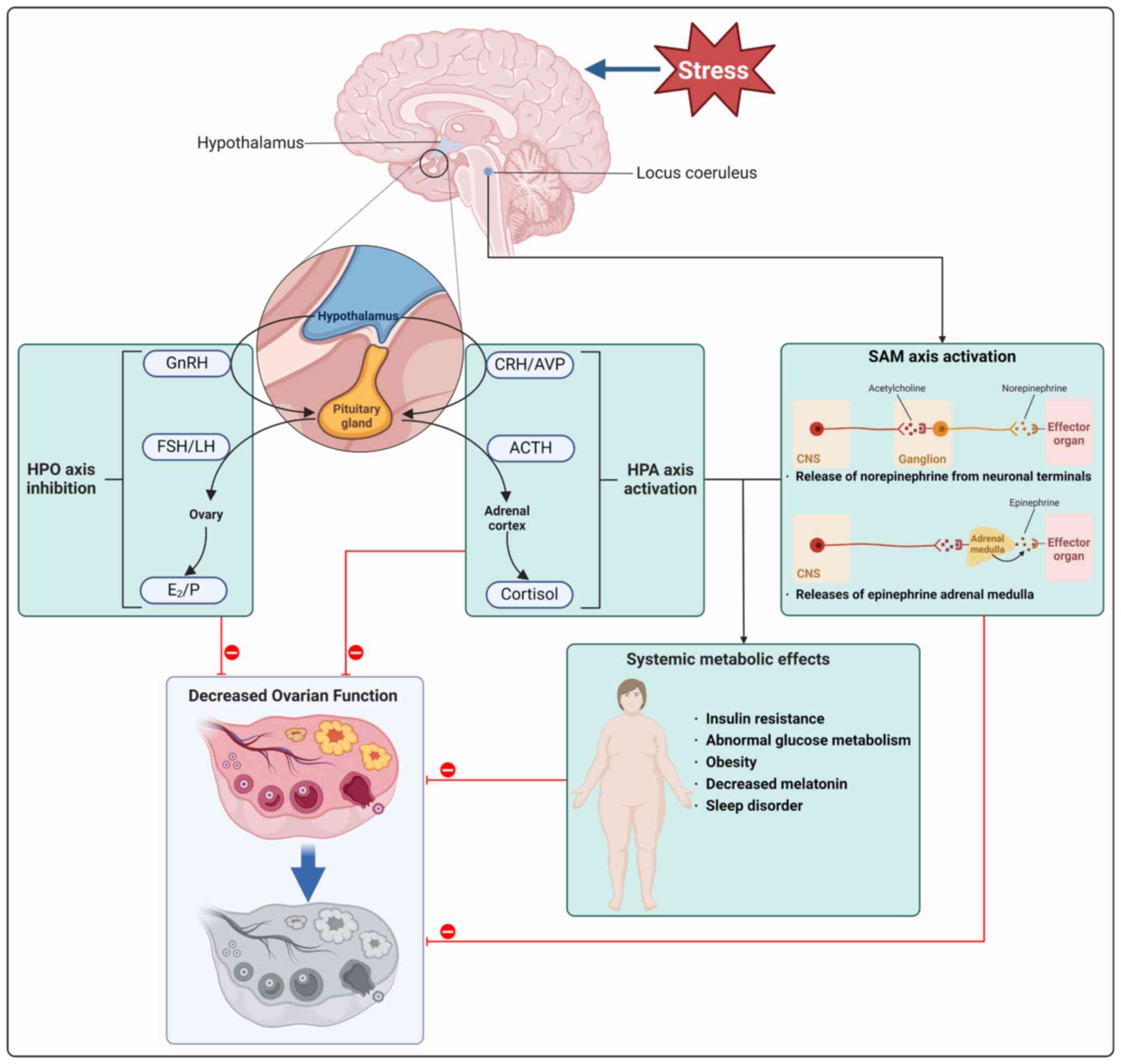
During the stress response, the autonomic nervous
system reacts instantly by triggering the SAM axis and activating
the HPA axis (144).
Stimulation of autonomic nerves prompts release of norepinephrine
(NE) from neuronal terminals, leading to the activation of the
adrenal medulla and subsequent discharge of epinephrine, resulting
in activation of the SAM axis (145). The activation of the
hypothalamic paraventricular nucleus (PVN) in the brain induces
release of CRH and arginine vasopressin (AVP) into the pituitary
portal system. CRH and AVP synergistically stimulate secretion of
pituitary adrenocorticotropic hormone, which enters the systemic
circulation, reaches the adrenal cortex and promotes the production
and release of cortisol (146).
Additionally, CRH that is released from the PVN area of the
inferior colliculus can activate locus coeruleus neurons,
increasing NE production (147). The neuroendocrine system serves
critical roles in regulating basal homeostasis and responding to
threats and it is also involved in the pathogenesis of diseases
marked by imbalance or dyshomeostasis (148-150). Prolonged exposure to life
stressors in individuals with a susceptible genetic makeup can lead
to visceral fat build-up, attributed to persistent
hypercortisolism, reactive insulin hypersecretion and reduced
growth hormone secretion (68).
Furthermore, in adults, stress can induce hypogonadism, and lead to
symptoms such as loss of libido and subfertility (68).
Elevated levels of cortisol have the potential to
disturb the equilibrium of hormones key for reproductive function,
such as GnRH, LH and FSH, leading to a disruption in ovarian
function (151,152). Additionally, stress hormones
released by the SAM axis impact the HPO axis by influencing the
secretion of GnRH and other regulatory hormones responsible for
ovarian function (145).
Furthermore, premature cessation of ovulation may be related to
elevated serum triglycerides and high-density lipoprotein
cholesterol levels (153).
Patients with POI exhibit irregularities in lipid and glucose
metabolism (154). Fig. 3 demonstrates associations between
psychological stress, HPA and HPO axes, sympathetic adrenal system
and metabolic disorders influencing ovarian function, highlighting
the role of HPA and SAM axis activation along with metabolic
disorder (Fig. 4; Tables III and IV).
The HPA axis serves a key role in the functioning of
the neuroendocrine system, overseeing responses to stress and
managing bodily functions such as digestion, immunity, mood, sexual
behavior and energy regulation (68). Activation of the HPA axis impacts
ovarian function by boosting the levels of cortisol and CRH.
Additionally, it can suppress release of gonadotropins, such as FSH
and LH, thereby affecting synthesis of steroid hormones, ovulation
and menstrual cycle (155).
Previous research has indicated that the surge in cortisol and CRH
resulting from HPA axis activation, along with the inhibition of
the HPO axis, are key factors in impairing ovarian function
(156).
In a CRS model, activation of the HPA axis results
in impaired oocyte developmental potential and decreased fertility
(21,93-95). Previous studies indicated that
patients undergoing IVF experienced higher levels of state anxiety
and serum cortisol (39,157). Additionally, patients with high
anticipatory state anxiety and cortisol concentrations have lower
pregnancy rates (158),
supporting the link between HPA axis activation and decreased
fertility. However, a cross-sectional study of 89 infertile
patients revealed that increased perceived mental stress is an
independent predictor of increasing FSH levels (β=0.22, P=0.045)
and likelihood for DOR (OR=9.97, 95%CI, 1.66, 55.99, P=0.012), but
there was no significant difference in serum cortisol levels
between patients with DOR and normal OR (r=−0.176, P=0.287), which
suggests that serum cortisol levels, representing current stress,
may not significantly be associated with ovarian reserve or
infertility (159). In direct
exposure studies, mouse oocytes exposed to physiological or
stress-induced levels of cortisol during in vitro maturation
did not show any impact on nuclear maturation or embryo development
(94,160). Conversely, injection of
cortisol into female mice resulted in impaired oocyte development,
decreased mitochondrial membrane potential and increased oxidative
stress and granulosa cell apoptosis through activating the TNF-α
system (128). These effects
may be linked to decreased expression of insulin-like growth factor
1 (IGF-1) and BDNF in granulosa cells and decreased FasL/Fas
secretion in ovaries and oocytes (161). The differing impacts of
cortisol in vivo and in vitro may be attributed to
the indirect influence of cortisol on oocytes via the NEM network.
In rats, exposure to CRS leads to increased expression of
hypothalamic RFamide-related peptide-3 (RFRP3) and levels of
circulating corticosterone, resulting in decreased HPG axis
function (95). However,
following a complete estrous cycle, female rats exhibit decreased
inclination towards mating; the stress-induced reproductive
dysfunction in female rats is alleviated by suppressing RFRP3
expression using a cytomegalovirus promoter-driven lentivirus
vector. This indicates that elevated levels of RFRP3 under stress
may have enduring detrimental impacts on fertility (95).
During stress, the hypothalamus produces a
neuropeptide known as CRH, which serves a vital role in the
regulation of ovarian functions, as its receptors have been
identified in reproductive organs, including ovary, uterus and
placenta (162). Previous
research has shown that CRH is responsible for activation of
primordial follicles in cultured newborn mouse ovaries in
vitro (21). Furthermore,
CRS could result in a decrease in oocyte competence and an increase
in both CRH and CRH receptor (CRH-R) expression in the ovary
(97). Co-culturing mouse
granulosa cells with CRH activates the Fas/FasL and TNF-α systems,
leading to heightened apoptosis of granulosa cells and oocytes, as
well as impaired early embryonic development (97,103). CRH-R antagonists such as
antalarmin are effective in inhibiting these damaging effects
(97). In another study,
injection of CRH in female rats for 12 days post-mating resulted in
a 40% disruption in pregnancy (163). These findings suggest that CRH
negatively impacts reproductive function.
Stress-induced activation of the HPA axis results
in increased levels of glucocorticoids in the bloodstream, along
with disrupted functioning of the HPO axis and ovarian cycle. In
female mice, chronic stress reduces the synthesis and release of LH
mediated by GnRH (164). One
explanation for decreased LH secretion may be the elevated levels
of cortisol due to stress. A previous study on mice exposed to
long-term corticosterone demonstrated a notable decrease in LH
levels, accompanied by alteration in neuronal activity in the
pituitary gonadotroph cells (165). CRS in rats inhibits the
preovulatory LH surge (166),
while CUS has been shown to raise cortisol levels and SOD activity
(87), both of which impact
ovulation. Moreover, patients diagnosed with FHA exhibit high
activation of the HPA axis, indicated by increased levels of
cortisol in the blood (167).
Furthermore, patients with FHA who experience spontaneous recovery
of ovarian function show lower serum cortisol levels post-recovery
compared with those who do not recover from FHA (168).
Ovarian function is regulated by the autonomic
nervous system, which cooperates with the HPO axis (169). Sympathetic fibers from the
celiac ganglia extend to ovarian follicles, mainly targeting the
theca layer, where NE, the principal neurotransmitter, interacts
with β-adrenergic receptors situated in the theca and granulosa
cells (170-172). Neural and hormonal signals
integrate to control steroid production and development of ovarian
follicles (173). Notably,
during stressful conditions, excessive activation of the SAM axis
disrupts this equilibrium, leading to compromised ovarian function
(174).
Previous research on rats has shown that normal
aging process is associated with elevated NE levels and increased
sympathetic activity within the ovary (175), implying that SAM axis
activation promotes ovarian aging. Chronic exposure to cold stress
activates the sympathetic nervous system, causing prolonged estrous
cycle, decreased levels of estrogen and progesterone and corpus
luteum count and heightened follicular atresia in rats (114,115,117,176).
Surgical denervation of the ovary and blocking the
β-adrenergic receptor using propranolol decrease ovarian
sympathetic activity, leading to a decrease in ovarian kisspeptin
(KISS1) and an increase in FSH receptor (FSHR) expression. This may
enhance follicle growth, suggesting activation of the SAM axis
could hinder ovarian function by elevating ovarian KISS1 levels
(177). NE is detected in human
follicular fluid (178).
However, physiological levels of NE are unable to trigger NE
receptor activation but can induce the generation of ROS in
granulosa cells (178). In
vitro, rat granulosa cells display increased apoptosis upon
co-culture with NE (179).
Furthermore, following recovery from cold stress model, rats
exhibited sustained elevated levels of ovarian NE, disrupted
estrous cycle and reduced fertility, indicating that SAM axis
activation may have lasting impacts on ovarian function in rats
(111).
Data from population studies also corroborate the
harm caused by SAM axis activation on ovarian function (12,58,180,181). A previous population-based
study including 274 patients showed significant decreases in the
likelihood of conceiving during the fertile window in the first
cycle of attempting pregnancy for patients with high SAA levels
compared with those with low levels (fecundability odds ratio,
FOR=0.85, 95%CI, 0.67, 1.09) (12). Although SAA is a marker for
stress and SAM activity, these findings support the idea that
stress impacts female fertility through the SAM pathway (12). Similarly, a study on 401 patients
found that those in the top tertile for SAA levels had a 29%
decrease in fecundity (longer time to pregnancy) compared with
those in the bottom tertile (FOR=0.71, 95%CI, 1.04, 4.11), and this
decrease in fecundity leads to an increased risk of infertility
(Relative Risk, RR=2.07, 95%CI=1.04, 4.11) (181). Aside from its potential impact
on fertility rates, SAM axis activation may also influence OR. A
study on 576 infertile patients found that higher levels of SAA
were linked to lower serum AMH (r=−0.315; adjusted r=−0.336, both
P<0.0001) (58). Another
population study involving 107 patients revealed that follicular NE
levels had a negative association with proportion of good quality
embryos (r=−0.62, P<0.05) but not with clinical pregnancy rates
(180).
The activation of stress-associated neuroendocrine
systems plays a key role in maintaining body equilibrium, yet
excessive stress can disrupt homeostasis (148). Previous studies indicate that
life stress poses a risk and has prognostic implications for
metabolic conditions such as obesity, T2DM and metabolic syndrome
(3,182). Moreover, involvement of the HPA
and SAM axes due to mental stress is linked to increased metabolic
disorder (183,184). Notably, patients with metabolic
syndrome or T2DM exhibit lower OR compared with healthy
counterparts (185,186), highlighting the connection
between stress, metabolic disorder and ovarian function.
Previous research suggests that stressors such as
stressful life events, emotional distress, anger, poor sleep and
work stress disrupt glucose balance and promote insulin resistance
(187). As a result, these
stressors are considered independent risk factors for T2DM
(188,189) and individuals experiencing
chronic stress are at a higher risk of developing T2DM compared
with non-stressed individuals (190). T2DM is a chronic metabolic
disorder characterized by hyperglycemia and insulin resistance,
both of which may contribute to ovarian dysfunction (191,192).
Previous population studies have indicated an
association between abnormal glucose metabolism, insulin resistance
and ovarian function (173-180). The incidence of oligomenorrhea
is notably higher in patients with T2DM compared with those with
normal blood glucose levels (203,204). Additionally, Study of Women's
Health Across the Nation Bone, a longitudinal and ethnically
diverse research project (n=2,171), observed that patients with
diabetes at the start of the study reached menopause ~3 years
sooner than non-diabetic counterparts (P=0.002) (205). Similarly, Sekhar et al
(206) reported that the mean
age of menopause is lower in patients with T2DM (44.65 years)
compared with those without T2DM (48.2 years; P<0.01). A
retrospective cohort study by Qin et al (207) found that patients with T2DM had
significantly decreased AMH levels (2.43±2.31 vs. 3.58±2.58
μg/l, P<0.001), and T2DM was an independent risk factor
for clinical pregnancy rate (adjusted OR=0.458, adjusted 95% CI,
0.235, 0.891, P=0.022) and live birth rate (adjusted OR=0.227,
adjusted 95%CI, 0.101, 0.513, P<0.001). Furthermore, early
menopause (onset at <45 years of age) was found to be more
prevalent among patients with T2DM compared with those without T2DM
(206). Decreased estrogen
secretion post-menopause can increase visceral fat deposition and
insulin resistance, significantly elevating risk of T2DM (208), and hormone replacement therapy
has been shown to mitigate this risk (209,210). Similarly, decreased insulin
sensitivity and impaired glucose metabolism are observed in animal
models that underwent ovariectomy, but these effects are reversed
by chronic estrogen administration (211,212). These findings highlight the key
role of estrogens in maintaining glucose homeostasis. Consequently,
stress-induced impaired glucose metabolism may lead to ovarian
dysfunction, which can cause further abnormal glucose
metabolism.
In summary, stress-induced abnormal glucose
metabolism has detrimental effects on ovarian function throughout
the reproductive lifecycle. However, more animal studies and cohort
studies are required to elucidate the interplay between these
factors.
Obesity has emerged as a growing health crisis with
notable repercussions for public health. In recent years,
accumulating evidence indicated that stress, notably increased
levels of cortisol, plays a crucial role in the onset of obesity
(213-216). Chronic stress activates the HPA
axis, which results in elevated circulating cortisol levels. This
causes the redistribution of white adipose tissue towards the
abdominal area, and also increases appetite, particularly for
calorie-dense foods (217).
Notably, the obesity epidemic coincides with factors such as
elevated cortisol production, prolonged stress, consumption of high
glycemic index foods and poor sleep quality (3). The Prospective Urban and Rural
Epidemiological study involving 120,000 individuals found that
participants with higher life stress scores (encompassing work,
family and financial stress and major life events) exhibited a
higher prevalence of abdominal obesity compared with those with
lower life stress (218).
Moreover, in elderly female patients, central obesity becomes
common due to redistribution of adipose tissue associated with
aging (219). Similar patterns
observed in fat redistribution due to chronic stress and aging in
female patients suggest a potential connection between these
factors (220). Recent studies
increasingly highlight that obesity is associated not only with
stress but also with diminished ovarian function (221-223).
Population-based studies have demonstrated an
inverse association between obesity and OR (186-192). A cross-sectional study on 36
healthy patients aged 40-52 years showed that AMH and inhibin
levels of obese patients were reduced by 77 (0.06, 95%CI, 0.12,
0.67 vs. 0.02 ng/ml, 95%CI, 0.12, 0.67, P=0.02) and 24% (12.5,
95%CI, 10.1, 15.4 ng/ml vs. 9.5 ng/ml, 95% CI, 8.0, 11.3, P=0.08),
respectively, compared with normal-weight patients after adjusting
for age, body mass index (BMI), race, smoking status and alcohol
use (224). In a previous study
involving 1,654 African American patients aged 23-35 years, it was
observed that obese participants exhibited 23.7% lower AMH
concentration (2.9 vs. 3.8 ng/ml) compared with those with a BMI
≤25 kg/m2 (225).
This finding has been corroborated by several other research
studies (226-228). Additionally, obesity is linked
to anovulatory cycles, irregular menstrual period and, diminished
implantation and pregnancy rate and it may also be associated with
PCOS (229,230). Research on the effects of a
high-fat diet (HFD) on ovarian follicular development has indicated
a decrease in number of primordial follicles (231). Exposure to HFD results in
follicular atresia, and accelerated follicle loss in rats via the
activation of mTOR and the inhibition of sirtuin 1 (SIRT1)
signaling, leading to premature ovarian failure (POF) (232). Previous studies on diet-induced
obesity in mice reveal increased oocyte apoptosis (233), the presence of multiple
spindles (double spindles) (234) and oocyte mitochondrial
dysfunction such as decreased mitochondrial membrane potential,
mitochondrial distribution and ATP levels (235). These findings are supported by
population studies: For example, Machtinger et al (236) found that severe obesity is
associated with a higher incidence of spindle anomalies in failed
fertilized oocytes, with observations of double spindles (OR=2.68,
95%CI, 1.39-5.15, P=0.003) and disorganized single spindles
(OR=4.58, 95%CI, 1.05-19.86, P=0.04). A recent study reported that
an increase in BMI was negatively associated with OR measured by
AMH levels in patients with PCOS (β=−0.03, 95%CI, −0.06-0.00,
P=0.034) (237). Although
direct evidence linking stress-induced obesity with impaired
ovarian function is lacking, the aforementioned indirect evidence
supports this connection.
Sleep disorder caused by mental stress are widely
hypothesized to be associated with infertility and ovarian
dysfunction (238-240). Previous studies on animals have
indicated that sleep deprivation (SD) disrupts the regularity of
the estrous cycle and results in anovulation, potentially leading
to reproductive dysfunction (241,242). Negative effects of SD may
impede follicular development during ovarian hyperstimulation due
to an increase in corticosterone levels in rats (243). SD has been shown to
significantly elevate serotonin levels, which decreases estradiol
production induced by FSH and FSH-driven expression of StAR protein
in follicles, resulting in decreased estrogen levels in the
bloodstream (244). This is
also supported by population-based research: A large-scale
prospective cohort study that monitored 80,840 patients from the
Nurses' Health Study between 1991 and 2013 discovered that working
rotating night shifts for 2 or >10 years is associated with
earlier onset of menopause (245).
Melatonin secretion decreases with age, which
serves a crucial role in regulating sleep and metabolic balance,
with its lipophilic nature giving it potent antioxidant properties
(246). Melatonin is a key
player in combating aging by protecting cells from oxidative
damage, a process exacerbated by the natural decline in melatonin
levels over time (247,248). Previous findings linked CUS to
increased ovarian ROS levels, with ROS-induced oxidative stress
being a notable contributor to ovarian damage and aging (85). Previous studies have demonstrated
that melatonin safeguards mouse ovarian granulosa cells by
mitigating oxidative stress-induced DNA damage, mitochondrial
dysfunction, lipid peroxidation and apoptosis (249), which are associated with the
SIRT1/FOXO1 and FOS pathways (249-252). Moreover, melatonin improves
oocyte quality by neutralizing ROS generated during follicle
maturation and ovulation and fertilization via NADPH, glutathione
(GSH), bone Morphogenetic Protein 15 (BMP15) and
SIRT1/SOD2-associated mechanisms (253-256). On the other hand, melatonin
supplements also significantly alleviated spindle/chromosome
disorganization, which may involve the SIRT2-dependent H4K16
deacetylation pathway (257,258). Clinical trials have indicated
that melatonin treatment in infertile patients can elevate
follicular melatonin levels, reduce oxidative damage and enhance
fertilization and pregnancy rates (253,257,259). Importantly, decreased melatonin
secretion has been linked to a higher risk of diabetes (260), underscoring the association
between melatonin levels, metabolic health and ovarian
function.
The likelihood of depression, anxiety and distress
is notably high among patients experiencing infertility and
psychological symptoms adversely affect fertility (261). Additionally, onset of
depression often occurs before the diagnosis of POI, implying that
depression and/or its treatment may contribute to development of
POI (15).
The effectiveness of psychological interventions in
decreasing psychological distress and association with significant
increases in pregnancy rates has been shown (5). After receiving antidepressant
medication and psychotherapy, infertile patients with depression
have higher pregnancy rates (262). Patients with POF/POI may
benefit from psychological interventions, as emotional and other
menopausal symptoms may be prolonged and not always alleviated by
hormone therapy (15). Stress
management and psychological therapy have been linked to
spontaneous pregnancy in certain infertile couples. Multiple
studies have documented cases of infertile couples conceiving
naturally after adopting a child (263,264). The aforementioned data suggests
that psychological interventions may be a promising approach for
these individuals to enhance quality of life.
CBT can help patients alleviate stress and achieve
relaxation by identifying and analyzing problems, recognizing
unproductive thoughts and behaviors, creating and implementing
replacement thoughts and behaviors, regular review and adjustment
of strategies, and applying learned skills to daily life (265). CBT has been demonstrated to be
an effective approach in treating depressive disorder, with
meta-analyses of randomized controlled trials corroborating its
efficacy for both anxiety (266-268) and depression (269,270).
A randomized controlled trial involving 56 pregnant
patients with a history of primary infertility indicated that CBT
counseling alleviates perceived stress, and anxiety, as well as
enhancing the quality of life score (271). Domar et al (272) showed that infertile patients
assigned to either structured CBT or standard support groups had a
higher pregnancy rate (52 vs. 20%, P=0.05), compared with those who
did not participate in these programs during their second IVF
cycle; however, P-value decreased slightly to 0.038 after adjusting
for intracytoplasmic sperm injection (ICSI) through logistic
regression. Mean stress scores after CBT treatment in infertile
patients were significantly decreased compared with before
(2.7±0.62 vs. 3.5±0.62; P<0.05) (273), which was confirmed by a
previous web-based CBT study (274). Researchers have identified CBT
as an effective treatment for enhancing the quality of life
(vasomotor symptoms, physical, psychosocial, sexual and total
domain) of perimenopausal patients (275-280). A randomized controlled trial
conducted with 50 patients with POI revealed mean scores of stress
[adjusted mean difference (AMD), −10.97; 95%CI, −11.64, −10.29;
P<0.001], state anxiety (AMD, −14.76; 95%CI, −15.77, −13.74;
P<0.001), trait anxiety (AMD, −14.41; 95%CI, −15.47, −13.74;
P<0.001) and depression (AMD, −7.44; 95%CI, −8.41, −6.46;
P<0.001) were significantly lower in the CBT group compared with
control group receiving routine care (281), suggesting healthcare providers
should employ this method to enhance mental health in these
patients.
A study allocated 16 patients with FHA equally to
CBT group or an observation group for 20 weeks; 6/8 patients in the
CBT group experienced recovery of ovarian activity compared with
2/88 in the observation group (87.5 vs. 25.0%, χ2=7.14)
(282). Besides promoting
ovulation, CBT has also been shown to lower cortisol levels in
patients with FHA (283). CBT
not only reinstates ovarian function but also impacts metabolic
processes (284). A pilot study
on 12 adolescents with PCOS demonstrated that CBT notably decreases
weight (104±26 vs. 93±18 kg, P<0.05) and improves depression
score (17±3 vs. 9.6±2, P<0.01) (285). Another study involving 15
overweight/obese adolescents with PCOS revealed that weekly CBT +
lifestyle modification sessions (LS) for 8 weeks significantly led
to weight loss, enhanced quality of life and reduced depressive
symptoms compared with those only receiving LS intervention
(286).
MBSR, a strategy of lessening stress and anxiety
through meditation, breathwork and body awareness, is a methodical
mindfulness meditation plan that is progressively accessible in
medical and healthcare environments to boost mental health and
general wellbeing (287). MBSR
has been demonstrated to decrease mental suffering and enhance life
quality in patients with cancer, heart disease and depressions
(288,289).
Individuals diagnosed with POF or POI often
experience prolonged mood disturbance and other menopausal symptoms
that may not be effectively alleviated by hormone therapy (15), indicating that MBSR may offer
potential benefits. A randomized controlled trial including 197
symptomatic peri- and postmenopausal patients found that both MBSR
and menopause education control (MEC) decreases total Greene
Climacteric Scale score and symptom score reduction of anxiety and
depression is significant in the MBSR compared with the MEC group
(anxiety, 5.97±2.49 vs. 6.97±2.57, P=0.07; depression, 4.78±2.60
vs. 5.39±2.41, P=0.031) (290).
In patients with POI, mindfulness practices have been shown to
decrease scores of menopause-specific quality of life (compared
with baseline, 48.32±4.96 vs. 95.6±9.77, P<0.0001; compared with
control, 48.32±4.96 vs. 102.6±14.9, P<0.0001) and the frequency
of hot flushes (6.74±6.34 vs. 23.4±13.9, P<0.0001) (291). MBSR improves emotional
wellbeing and biological outcomes in infertile patients (292-294).
Previous research has shown that music therapy has
the potential to decrease stress, improve quality of life and
alleviate depression and anxiety (295). In a randomized-controlled study
of 48 postmenopausal patients, music therapy significantly
decreased BDI score (11.81±8.62 vs. 16.44±6.09, P=0.031) compared
with control and the Menopause Rating Scale (MRS) total and
sub-scale scores were significantly decreased when comparing before
and after intervention (296).
In a study of 40 patients with perimenopause
syndrome, music therapy was more likely to improve psychological
and emotional symptoms compared with CBT (MRS total, 9.2 vs. 3.5,
P=0.008; MRS-psychological, 6.5 vs. 0.9, P=0.004; Patient Health
Questionnaire 9 score, 4.3 vs. 1.6, P=0.009) (297). A comprehensive meta-analysis
indicated that music therapy decreases the anxiety (MD=−3.09,
95%CI, −5.57, −0.61, P=0.01) and pain score (MD=−2.93, 95%CI,
−3.86, −2.00, P<0.0001) and increased the satisfaction score
(MD=1.51, 95%CI, 0.40, 2.61, P=0.008) in infertile patients
undergoing ART (298). While
music therapy increases the clinical pregnancy rates, the
difference is not statistically significant (RR=1.08, 95%CI, 0.94,
1.26, P=0.28) (298,299).
Alongside seeking advice from professional
psychologists, individuals may opt for self-care. An evaluation of
166 patients undergoing first-time IVF through a randomized
controlled prospective study, examining the effectiveness of a
cognitive coping and relaxation intervention (CCRI) that could be
self-administered indicated that patients who engaged in the CCRI
demonstrated enhanced positive reappraisal coping, better Fertility
Quality of Life score and reduced levels of anxiety (300). Another randomized controlled
prospective pilot study explored the impact of an online version of
the mind/body program (301),
where patients assigned to the intervention group showed
significant decreases in anxiety and depression, along with an
increased likelihood of pregnancy.
Yoga has been shown to be a beneficial intervention
for anxiety and depression, stress reduction and enhancement of the
overall wellbeing among the general public (302). Furthermore, yoga can alleviate
anxiety and depression in individuals undergoing IVF treatment and
increase the ART success rate (303). Yoga can enhance the quality of
life in relation to infertility, with a specific focus on
diminishing adverse emotions and cognition linked to infertility
(304). A separate study
investigating yoga as a complementary approach for patients
receiving IVF treatment yielded comparable results (305). Yoga alleviates the menopausal
symptoms, anxiety and depression of menopausal patients and
decreases the levels of FSH and LH (306,307).
As aforementioned, the activation of
stress-associated neuroendocrine systems causes metabolic imbalance
in the body, and metabolic disorders lead to ovary dysfunction.
CR refers to a reduction in calorie consumption
without eliminating essential nutrients (310). CR is hypothesized to extend the
lifespan of rodents, yeast, worms, flies and primates by decreasing
body weight and slowing aging process (311). CR has been shown to mitigate
age-associated diseases such as neurodegeneration, hepatic
steatosis and T2DM (312).
Notably, recent studies have shown that CR can
protect ovarian function. CR decreases aneuploidy, chromosomal
misalignment and meiotic spindle abnormality in oocytes in aged
mice (313-315). CR can attenuate
aging-associated decrease in chromosomal cohesion and protect the
normal progress of meiosis (315). CR enhances ovarian estrogen
sensitivity and inhibits ovulation, thereby extending reproductive
lifespan (316,317). CR can also reduce primordial
follicle activation and lead to increased ovarian Forkhead box O3
(Foxo3a) expression, activation of the SIRT1 signaling pathway and
suppression of the mTOR signaling pathway, which may increase
preservation of the ovarian primordial follicular reserve and delay
menopause onset (318-322).
Obesity impairs ovarian function and obese patients
are often encouraged to lose weight before conception to increase
the chances of a successful pregnancy (323). However, the protective effect
of CR on ovarian function lacks large-scale multicenter clinical
trials and CR is a long-term dietary adjustment process that is
difficult to maintain (324).
Recently, there has been considerable interest in identifying drugs
that can mimic the effects of CR. CR mimetics (CRMs) promote
lifespan and sustain the health benefits of CR without requiring
dietary restriction (325-329). CRMs have good clinical
prospects because they do not require long-term dietary restriction
and have better patient compliance.
Metformin has been demonstrated to increase OR and
protect ovarian function through regulatory mechanisms such as
inhibiting the mTOR pathway, enhancing SIRT1 expression and
decreasing oxidative stress damage (335,336). Furthermore, in aged mice,
metformin decreases mitochondrial ROS via SIRT3-mediated
acetylation of SOD2K68 in oocytes (337). For chemotherapy-induced ovarian
damage, metformin may attenuate carboplatin-induced ovarian damage,
potentially through its antioxidative effects (338). In addition, metformin decreases
aging-related ovarian fibrosis. Metformin-responsive macrophage
subpopulations appear in aged mice treated with metformin, which
may decrease ovarian fibrosis by clearing fibroblasts that produce
the senescence-associated secretory phenotype in aged ovaries
(339). Therefore, metformin
may serve as a promising drug to protect reproductive function.
In humans, resveratrol can improve the follicular
microenvironment to improve IVF outcomes (344). Intraperitoneal injection of
resveratrol during superovulation improves oocyte quality in both
young and old mice, and increased the number of oocytes retrieved
from old mice (345).
Resveratrol improves fertility of reproductive age female mice,
which may be achieved by activating SIRT1 and increasing the
expression of Foxo3a (346-349). In vitro, resveratrol
increased first polar body emission rates and improves oocyte
mitochondrial function (350-352). In vitro, resveratrol
protects granulosa cells by activating autophagy to resist
oxidative stress (353,354). In conclusion, resveratrol is
beneficial to ovarian function and protects oocytes.
Psychological stress is associated with poor health
and impaired ovarian function (2,4).
The present review summarizes the adverse effects of mental stress
on ovarian function, which are associated with decreased IVF
success rates and OR and the occurrence of POI, FHA and PCOS.
Mental stress is involved in the pathogenesis of POI, FHA and POCS
(15,17,61). However, more prospective studies
are needed to explore the effect of psychological stress in
initiation and progression of these diseases. Elevated levels of
perceived stress, as well as SAA, could reduce OR (13,56,58). However, salivary cortisol,
another biomarker used to measure psychological stress, has no
association with fecundability (181). Cortisol exhibits a distinct
circadian rhythm, with peak levels occurring in the morning and
decreasing throughout the day; thus, it may be important to obtain
multiple samples/day from a patient in order to investigate its
effects on human health (355).
Mental stress has an adverse effect on the outcome of IVF or ICSI,
although some studies suggest that there is no association between
them (48). To the best of our
knowledge, no studies have reported that psychological stress has a
positive effect on pregnancy outcomes. Anderheim et al
(48) found no evidence that
mental stress impairs IVF outcomes. However, Aimagambetova et
al (36) found that higher
STAI-S and STAI-T are negatively associated with clinical pregnancy
outcomes. A potential reason for this discrepancy is that, when
filling out psychological questionnaires, patients may not provide
accurate assessments of distress. Different inclusion criteria,
assessment tools for psychological status and study design may be
additional reasons for the inconsistent results. The influence of
stress on ART outcomes remains controversial, but, for infertile
patients, psychological intervention can alleviate anxiety and
depression, potentially resulting in notably improved pregnancy
rates. Multi-center clinical trials with large sample sizes are
necessary in future research. Under ideal conditions, prospective
studies through puberty to perimenopause should regularly collect
psychological questionnaires/scales, metabolic and ovarian function
indexes to demonstrate causal mechanisms linking stress and ovarian
dysfunction.
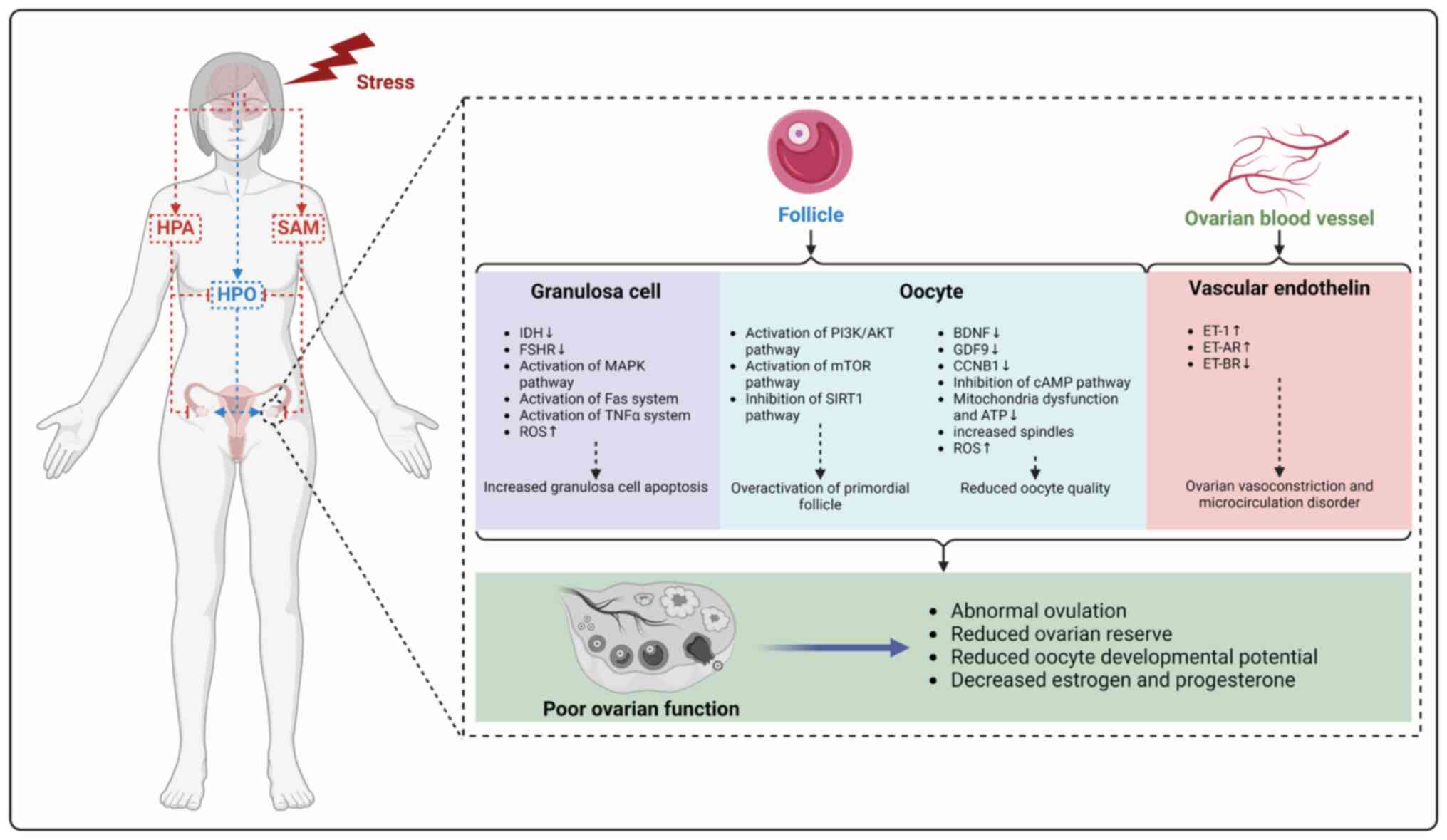
The present study summarizes animal models used to
study the impact of mental stress on ovarian function, such as CUS,
CRS and chronic cold stress, as well as evaluation methods. Mental
stress affects the follicle and ovarian vascular function via the
HPA, SAM and HPO axis and neurometabolic network, in which stress
induces granulosa cell apoptosis by activating the MAPK, FAS and
TNF-α pathway and increasing ROS, leading to overactivation of
primordial follicles and decreasing the number of follicles by
activating the PI3K/AKT pathway, as well as oocyte quality by
downregulating BDNF, GDF9 and CCNB1, inhibiting the cAMP pathway,
mitochondrial energy metabolism imbalance and inducing ROS
accumulation. In addition, stress can lead to disorder of ovarian
vascular structure and microenvironment by upregulating expression
of ET-1 and ET-AR and downregulating expression of ET-BR, leading
to a decline in ovarian function (Fig. 4).
Multiple studies have examined the impact of
chronic stress on the neuroendocrine system over extended periods
(156,356). In the existing studies,
traditional biological stress markers (cortisol and α-amylase) have
been widely used to assess stress levels of patients and
experimental animals (10,58,85,181). However, due to the differences
in susceptibility between individuals and evaluation methods,
previous results are inconsistent. Obayashi (357) summarized advantages and
weaknesses of different salivary stress markers in the assessment
of stress, such as cortisol, α-amylase, chromogranin A and IgA and
proposed optimization of the method of saliva collection in order
to improve the consistency of stress levels. As Shah et al
(358) noted, when exploring
the association between psychological stress and ovarian function,
a comprehensive assessment of stress levels should include
physiological markers (such as heart rate variability, blood
pressure and cytokine levels) and biochemical indicators (cortisol,
catecholamines, copeptin, SAA, IL-6 and C-reactive protein levels),
as well as individual subjective assessments (self-reported stress
levels, perceived stress scale or psychometric evaluations).
Neuroendocrine changes caused by mental stress lead to systemic
inflammation, leading to an increase in systemic inflammatory
markers and health issues such as cardiovascular disease, cognitive
dysfunction and accelerated aging (359). Therefore, inflammatory
biomarkers such as TNF-α, HIF-1α and asymmetric dimethylarginine
(ADMA) may serve as biological indicators to measure mental stress
and its impact on physical health. Combinations of different
markers and approaches such as wearable device-based stress
detection (360) may establish
effective stress markers. However, its effects need more clinical
research for verification.
Existing studies focus on the direct role of
psychological stress on the onset of POI, FHA and PCOS (15,17,61). For example, most studies have
explored the impact of PCOS on the mental stress or
psychological-associated disease (361-363). Large-scale prospective studies
with lager samples are needed to support whether mental stress
causes PCOS. Furthermore, the existing evidence that mental stress
impairs ovarian function through metabolic disturbances is indirect
(207,236). cohort studies and animal
experiments are required to determine whether mental stress
directly affects ovarian function through metabolism-related
diseases.
It is key to elucidate the specific molecular
mechanism underlying the effect of on the ovary and provide
psychological intervention strategies for patients with ovarian
dysfunction, especially for those in need of ART intervention.
Hence, psychological interventions (CBT, MBSR and music therapy)
may be beneficial to reduce adverse influence of stress and improve
ovarian function and reproductive outcomes. The present review may
provide an alternative intervention strategy for improvement of
ovarian dysfunction and infertility.
Not applicable.
YL and FF conceived and designed the study. YH and
FF wrote the manuscript and constructed figures and tables. WuW,
WM, WeW, WR and SW revised the manuscript. All the authors have
read and approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Key Research and
Development Program of China (grant nos. 2022YFC2704100 and
2021YFC2700402-02) and National Natural Science Foundation of China
(grant no. 81902669).
|
1
|
Wang S, Quan L, Chavarro JE, Slopen N,
Kubzansky LD, Koenen KC, Kang JH, Weisskopf MG, Branch-Elliman W
and Roberts AL: Associations of depression, anxiety, worry,
perceived stress, and loneliness prior to infection with risk of
post-COVID-19 conditions. JAMA Psychiatry. 79:1081–1091. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valsamakis G, Chrousos G and Mastorakos G:
Stress, female reproduction and pregnancy.
Psychoneuroendocrinology. 100:48–57. 2019. View Article : Google Scholar
|
|
3
|
Kivimäki M, Bartolomucci A and Kawachi I:
The multiple roles of life stress in metabolic disorders. Nat Rev
Endocrinol. 19:10–27. 2023. View Article : Google Scholar
|
|
4
|
Bala R, Singh V, Rajender S and Singh K:
Environment, lifestyle, and female infertility. Reprod Sci.
28:617–638. 2021. View Article : Google Scholar
|
|
5
|
Rooney KL and Domar AD: The relationship
between stress and infertility. Dialogues Clin Neurosci. 20:41–47.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maeda E, Nomura K, Hiraike O, Sugimori H,
Kinoshita A and Osuga Y: Domestic work stress and self-rated
psychological health among women: A cross-sectional study in Japan.
Environ Health Prev Med. 24:752019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Y and Aungsuroch Y: Work stress,
perceived social support, self-efficacy and burnout among Chinese
registered nurses. J Nurs Manag. 27:1445–1453. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bapayeva G, Aimagambetova G, Issanov A,
Terzic S, Ukybassova T, Aldiyarova A, Utepova G, Daribay Z,
Bekbossinova G, Balykov A, et al: The effect of stress, anxiety and
depression on in vitro fertilization outcome in kazakhstani public
clinical setting: A cross-sectional study. J Clin Med. 10:9372021.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu H, Ouyang N, Li R, Tuo P, Mai M and
Wang W: The effects of anxiety and depression on in vitro
fertilisation outcomes of infertile Chinese women. Psychol Health
Med. 22:37–43. 2017. View Article : Google Scholar
|
|
10
|
Sallem A, Essoussi H, Mustapha HB, Zaouali
M and Ajina M: Impact of psychological stress on the outcomes of
assisted reproduction in Tunisian infertile women. Pan Afr Med J.
40:2502021. View Article : Google Scholar
|
|
11
|
Klonoff-Cohen H, Chu E, Natarajan L and
Sieber W: A prospective study of stress among women undergoing in
vitro fertilization or gamete intrafallopian transfer. Fertil
Steril. 76:675–687. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Buck Louis GM, Lum KJ, Sundaram R, Chen Z,
Kim S, Lynch CD, Schisterman EF and Pyper C: Stress reduces
conception probabilities across the fertile window: Evidence in
support of relaxation. Fertil Steril. 95:2184–2189. 2011.
View Article : Google Scholar
|
|
13
|
Mínguez-Alarcón L, Williams PL, Souter I,
Ford JB, Hauser R and Chavarro JE; Earth Study Team: Perceived
stress and markers of ovarian reserve among subfertile women.
Reprod Biomed Online. 46:956–964. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Golenbock SW, Wise LA, Lambert-Messerlian
GM, Eklund EE and Harlow BL: Association between a history of
depression and anti-müllerian hormone among late-reproductive aged
women: The Harvard study of moods and cycles. Womens Midlife
Health. 6:92020. View Article : Google Scholar
|
|
15
|
Allshouse AA, Semple AL and Santoro NF:
Evidence for prolonged and unique amenorrhea-related symptoms in
women with premature ovarian failure/primary ovarian insufficiency.
Menopause. 22:166–174. 2015. View Article : Google Scholar
|
|
16
|
Lawson EA, Donoho D, Miller KK, Misra M,
Meenaghan E, Lydecker J, Wexler T, Herzog DB and Klibanski A:
Hypercortisolemia is associated with severity of bone loss and
depression in hypothalamic amenorrhea and anorexia nervosa. J Clin
Endocrinol Metab. 94:4710–4716. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goodarzi MO, Dumesic DA, Chazenbalk G and
Azziz R: Polycystic ovary syndrome: Etiology, pathogenesis and
diagnosis. Nat Rev Endocrinol. 7:219–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Freeman EW, Sammel MD, Lin H and Nelson
DB: Associations of hormones and menopausal status with depressed
mood in women with no history of depression. Arch Gen Psychiatry.
63:375–382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao L, Zhao F, Zhang Y, Wang W and Cao Q:
Diminished ovarian reserve induced by chronic unpredictable stress
in C57BL/6 mice. Gynecol Endocrinol. 36:49–54. 2020. View Article : Google Scholar
|
|
20
|
Jiang Y, Xu J, Tao C, Lin Y, Lin X, Li K,
Liu Q, Saiyin H, Hu S, Yao G, et al: Chronic stress induces meiotic
arrest failure and ovarian reserve decline via the cAMP signaling
pathway. Front Endocrinol (Lausanne). 14:11770612023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu M, Sun J, Wang Q, Zhang Q, Wei C and
Lai D: Chronic restraint stress induces excessive activation of
primordial follicles in mice ovaries. PLoS One. 13:e01948942018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Poole L and Hackett RA: Diabetes distress:
The psychological burden of living with diabetes. Lancet Diabetes
Endocrinol. 12:439–441. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bartoskova Polcrova A, Dalecka A, Szabo D,
Gonzalez Rivas JP, Bobak M and Pikhart H: Social and environmental
stressors of cardiometabolic health. Sci Rep. 14:141792024.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schliep KC, Mumford SL, Vladutiu CJ,
Ahrens KA, Perkins NJ, Sjaarda LA, Kissell KA, Prasad A,
Wactawski-Wende J and Schisterman EF: Perceived stress,
reproductive hormones, and ovulatory function: A prospective cohort
study. Epidemiology. 26:177–184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harlow BL, Wise LA, Otto MW, Soares CN and
Cohen LS: Depression and its influence on reproductive endocrine
and menstrual cycle markers associated with perimenopause: The
Harvard study of moods and cycles. Arch Gen Psychiatry. 60:29–36.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Albert KM and Newhouse PA: Estrogen,
stress, and depression: cognitive and biological interactions. Annu
Rev Clin Psychol. 15:399–423. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yatsenko SA and Rajkovic A: Genetics of
human female infertility. Biol Reprod. 101:549–566. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matthiesen SMS, Frederiksen Y, Ingerslev
HJ and Zachariae R: Stress, distress and outcome of assisted
reproductive technology (ART): A meta-analysis. Hum Reprod.
26:2763–2776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Massey AJ, Campbell B, Raine-Fenning N,
Aujla N and Vedhara K: The association of physiological cortisol
and IVF treatment outcomes: A systematic review. Reprod Med Biol.
13:161–176. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen TH, Chang SP, Tsai CF and Juang KD:
Prevalence of depressive and anxiety disorders in an assisted
reproductive technique clinic. Hum Reprod. 19:2313–2318. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Volgsten H, Skoog Svanberg A, Ekselius L,
Lundkvist O and Sundström Poromaa I: Prevalence of psychiatric
disorders in infertile women and men undergoing in vitro
fertilization treatment. Hum Reprod. 23:2056–2063. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pasch LA, Holley SR, Bleil ME, Shehab D,
Katz PP and Adler NE: Addressing the needs of fertility treatment
patients and their partners: Are they informed of and do they
receive mental health services? Fertil Steril. 106:209–215.e2.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lakatos E, Szigeti JF, Ujma PP, Sexty R
and Balog P: Anxiety and depression among infertile women: A
cross-sectional survey from Hungary. BMC Womens Health. 17:482017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
An Y, Sun Z, Li L, Zhang Y and Ji H:
Relationship between psychological stress and reproductive outcome
in women undergoing in vitro fertilization treatment: Psychological
and neurohormonal assessment. J Assist Reprod Genet. 30:35–41.
2013. View Article : Google Scholar :
|
|
35
|
Terzioglu F, Turk R, Yucel C, Dilbaz S,
Cinar O and Karahalil B: The effect of anxiety and depression
scores of couples who underwent assisted reproductive techniques on
the pregnancy outcomes. Afr Health Sci. 16:441–450. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aimagambetova G, Issanov A, Terzic S,
Bapayeva G, Ukybassova T, Baikoshkarova S, Aldiyarova A, Shauyen F
and Terzic M: The effect of psychological distress on IVF outcomes:
Reality or speculations? PLoS One. 15:e02420242020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Turner K, Reynolds-May MF, Zitek EM,
Tisdale RL, Carlisle AB and Westphal LM: Stress and anxiety scores
in first and repeat IVF cycles: A pilot study. PLoS One.
8:e637432013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Quant HS, Zapantis A, Nihsen M, Bevilacqua
K, Jindal S and Pal L: Reproductive implications of psychological
distress for couples undergoing IVF. J Assist Reprod Genet.
30:1451–1458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Csemiczky G, Landgren BM and Collins A:
The influence of stress and state anxiety on the outcome of
IVF-treatment: Psychological and endocrinological assessment of
Swedish women entering IVF-treatment: Stress and state anxiety on
outcome of IVF-treatment. Acta Obstet Gynecol Scand. 79:113–118.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Boivin J, Griffiths E and Venetis CA:
Emotional distress in infertile women and failure of assisted
reproductive technologies: Meta-analysis of prospective
psychosocial studies. BMJ. 342:d2232011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pasch LA, Gregorich SE, Katz PK, Millstein
SG, Nachtigall RD, Bleil ME and Adler NE: Psychological distress
and in vitro fertilization outcome. Fertil Steril. 98:459–464.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chai Y, Li Q, Wang Y, Niu B, Chen H, Fan
T, Ke X and Zou H: Cortisol dysregulation in anxiety infertile
women and the influence on IVF treatment outcome. Front Endocrinol
(Lausanne). 14:11077652023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cesta CE, Johansson ALV, Hreinsson J,
Rodriguez-Wallberg KA, Olofsson JI, Holte J, Wramsby H, Wramsby M,
Cnattingius S, Skalkidou A and Nyman Iliadou A: A prospective
investigation of perceived stress infertility-related stress and
cortisol levels in women undergoing in vitro fertilization:
Influence on embryo quality and clinical pregnancy rate. Acta
Obstet Gynecol Scand. 97:258–268. 2018. View Article : Google Scholar
|
|
44
|
Mirzaasgari H, Momeni F, Pourshahbaz A,
Keshavarzi F and Hatami M: The relationship between coping
strategies and infertility self-efficacy with pregnancy outcomes of
women undergoing in vitro fertilization: A prospective cohort
study. Int J Reprod Biomed. 20:539–548. 2022.PubMed/NCBI
|
|
45
|
Peng M, Wen M, Jiang T, Jiang Y, Lv H,
Chen T, Ling X, Li H, Meng Q, Huang B, et al: Stress, anxiety, and
depression in infertile couples are not associated with a first IVF
or ICSI treatment outcome. BMC Pregnancy Childbirth. 21:7252021.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Park J, Stanford JB, Porucznik CA,
Christensen K and Schliep KC: Daily perceived stress and time to
pregnancy: A prospective cohort study of women trying to conceive.
Psychoneuroendocrinology. 110:1044462019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lynch CD, Sundaram R, Buck Louis GM, Lum
KJ and Pyper C: Are increased levels of self-reported psychosocial
stress, anxiety, and depression associated with fecundity? Fertil
Steril. 98:453–458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Anderheim L, Holter H, Bergh C and Möller
A: Does psychological stress affect the outcome of in vitro
fertilization? Hum Reprod. 20:2969–2975. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Milad MP, Klock SC, Moses S and Chatterton
R: Stress and anxiety do not result in pregnancy wastage. Hum
Reprod. 13:2296–2300. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Butts CD, Bloom MS, Frye CA, Walf AA,
Parsons PJ, Steuerwald AJ, Ilonze C and Fujimoto VY: Urine cortisol
concentration as a biomarker of stress is unrelated to IVF outcomes
in women and men. J Assist Reprod Genet. 31:1647–1653. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stuenkel CA and Gompel A: Primary ovarian
insufficiency. N Engl J Med. 388:154–163. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sun J, Fan Y, Guo Y, Pan H, Zhang C, Mao
G, Huang Y, Li B, Gu T, Wang L, et al: Chronic and cumulative
adverse life events in women with primary ovarian insufficiency: An
exploratory qualitative study. Front Endocrinol (Lausanne).
13:8560442022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xu YP, Fu JC, Hong ZL, Zeng DF, Guo CQ, Li
P and Wu JX: Psychological stressors involved in the pathogenesis
of premature ovarian insufficiency and potential intervention
measures. Gynecol Endocrinol. 40:23600852024. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Schmidt PJ, Luff JA, Haq NA, Vanderhoof
VH, Koziol DE, Calis KA, Rubinow DR and Nelson LM: Depression in
women with spontaneous 46, XX primary ovarian insufficiency. J Clin
Endocrinol Metab. 96:E278–E287. 2011. View Article : Google Scholar :
|
|
55
|
Freeman EW, Sammel MD, Liu L, Gracia CR,
Nelson DB and Hollander L: Hormones and menopausal status as
predictors of depression in womenin transition to menopause. Arch
Gen Psychiatry. 61:62–70. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bleil ME, Adler NE, Pasch LA, Sternfeld B,
Gregorich SE, Rosen MP and Cedars MI: Psychological stress and
reproductive aging among pre-menopausal women. Hum Reprod.
27:2720–2728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bleil ME, Adler NE, Pasch LA, Sternfeld B,
Gregorich SE, Rosen MP and Cedars MI: Depressive symptomatology,
psychological stress, and ovarian reserve: A role for psychological
factors in ovarian aging? Menopause. 19:1176–1185. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dong YZ, Zhou FJ and Sun YP: Psychological
stress is related to a decrease of serum anti-müllerian hormone
level in infertile women. Reprod Biol Endocrinol. 15:512017.
View Article : Google Scholar
|
|
59
|
Lania A, Gianotti L, Gagliardi I,
Bondanelli M, Vena W and Ambrosio MR: Functional hypothalamic and
drug-induced amenorrhea: An overview. J Endocrinol Invest.
42:1001–1010. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gordon CM, Ackerman KE, Berga SL, Kaplan
JR, Mastorakos G, Misra M, Murad MH, Santoro NF and Warren MP:
Functional hypothalamic amenorrhea: An endocrine society clinical
practice guideline. J Clin Endocrinol Metab. 102:1413–1439. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Bomba M, Gambera A, Bonini L, Peroni M,
Neri F, Scagliola P and Nacinovich R: Endocrine profiles and
neuropsychologic correlates of functional hypothalamic amenorrhea
in adolescents. Fertil Steril. 87:876–885. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Marcus MD, Loucks TL and Berga SL:
Psychological correlates of functional hypothalamic amenorrhea.
Fertil Steril. 76:310–316. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Dundon CM, Rellini AH, Tonani S,
Santamaria V and Nappi R: Mood disorders and sexual functioning in
women with functional hypothalamic amenorrhea. Fertil Steril.
94:2239–2243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cai J, Zhang Y, Wang Y, Li S, Wang L,
Zheng J, Jiang Y, Dong Y, Zhou H, Hu Y, et al: High thyroid
stimulating hormone level is associated with hyperandrogenism in
euthyroid polycystic ovary syndrome (PCOS) women, independent of
age, BMI, and thyroid autoimmunity: A cross-sectional analysis.
Front Endocrinol (Lausanne). 10:2222019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chang AY, Lalia AZ, Jenkins GD, Dutta T,
Carter RE, Singh RJ and Nair KS: Combining a nontargeted and
targeted metabolomics approach to identify metabolic pathways
significantly altered in polycystic ovary syndrome. Metabolism.
71:52–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Farrell K and Antoni MH: Insulin
resistance, obesity, inflammation, and depression in polycystic
ovary syndrome: Biobehavioral mechanisms and interventions. Fertil
Steril. 94:1565–1574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hillman SC, Bryce C, Caleyachetty R and
Dale J: Women's experiences of diagnosis and management of
polycystic ovary syndrome: A mixed-methods study in general
practice. Br J Gen Pract. 70:e322–e329. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chrousos GP: Stress and disorders of the
stress system. Nat Rev Endocrinol. 5:374–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kleinert M, Clemmensen C, Hofmann SM,
Moore MC, Renner S, Woods SC, Huypens P, Beckers J, de Angelis MH,
Schürmann A, et al: Animal models of obesity and diabetes mellitus.
Nat Rev Endocrinol. 14:140–162. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Seemann S, Zohles F and Lupp A:
Comprehensive comparison of three different animal models for
systemic inflammation. J Biomed Sci. 24:602017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cheon DJ and Orsulic S: Mouse models of
cancer. Annu Rev Pathol. 6:95–119. 2011. View Article : Google Scholar
|
|
72
|
Antoniuk S, Bijata M, Ponimaskin E and
Wlodarczyk J: Chronic unpredictable mild stress for modeling
depression in rodents: Meta-analysis of model reliability. Neurosci
Biobehav Rev. 99:101–116. 2019. View Article : Google Scholar
|
|
73
|
Du Preez A, Onorato D, Eiben I, Musaelyan
K, Egeland M, Zunszain PA, Fernandes C, Thuret S and Pariante CM:
Chronic stress followed by social isolation promotes
depressive-like behaviour, alters microglial and astrocyte biology
and reduces hippocampal neurogenesis in male mice. Brain Behav
Immun. 91:24–47. 2021. View Article : Google Scholar
|
|
74
|
Khan AR, Geiger L, Wiborg O and Czéh B:
Stress-induced morphological, cellular and molecular changes in the
brain-lessons learned from the chronic mild stress model of
depression. Cells. 9:10262020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Xiang Y, Jiang L, Gou J, Sun Y, Zhang D,
Xin X, Song Z and Huang J: Chronic unpredictable mild
stress-induced mouse ovarian insufficiency by interrupting lipid
homeostasis in the ovary. Front Cell Dev Biol. 10:9336742022.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Fu XY, Chen HH, Zhang N, Ding MX, Qiu YE,
Pan XM, Fang YS, Lin YP, Zheng Q and Wang WQ: Effects of chronic
unpredictable mild stress on ovarian reserve in female rats:
Feasibility analysis of a rat model of premature ovarian failure.
Mol Med Rep. 18:532–540. 2018.PubMed/NCBI
|
|
77
|
Ma J, Wang L, Yang D, Luo J, Gao J, Wang
J, Guo H, Li J, Wang F, Wu J and Hu R: Chronic stress causes
ovarian fibrosis to impair female fertility in mice. Cell Signal.
122:1113342024. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ding SM, Shi LG, Xing F, Cui SS, Cheng HR,
Liu Y, Ji DM, Liang D, Cao YX and Liu YJ: Melatonin protects
against mitochondrial dyshomeostasis and ovarian damage caused by
chronic unpredictable mild stress through the eIF2α-AFT4 signaling
pathway in mice. Reprod Sci. 31:3191–3201. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Fu X, Zheng Q, Zhang N, Ding M, Pan X,
Wang W and Chen H: CUMS promotes the development of premature
ovarian insufficiency mediated by nerve growth factor and its
receptor in rats. BioMed Res Int. 2020:19468532020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Xu HX, Lin SX, Gong Y, Huo ZX, Zhao CY,
Zhu HM and Xi SY: Chaiyu-dixian formula exerts protective effects
on ovarian follicular abnormal development in chronic unpredictable
mild stress (CUMS) rat model. Front Pharmacol. 11:2452020.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhao X, Ma R, Zhang X, Cheng R, Jiang N,
Guo M, Rong B, Liu Y, Chen M, Feng W and Xia T: Reduced growth
capacity of preimplantation mouse embryos in chronic unpredictable
stress model. Mol Reprod Dev. 88:80–95. 2021. View Article : Google Scholar
|
|
82
|
Wu LM, Hu MH, Tong XH, Han H, Shen N, Jin
RT, Wang W, Zhou GX, He GP and Liu YS: Chronic unpredictable stress
decreases expression of brain-derived neurotrophic factor (BDNF) in
mouse ovaries: relationship to oocytes developmental potential.
PLoS One. 7:e523312012. View Article : Google Scholar
|
|
83
|
Li XH, Pang HQ, Qin L, Jin S, Zeng X, Bai
Y and Li SW: HSP70 overexpression may play a protective role in the
mouse embryos stimulated by CUMS. Reprod Biol Endocrinol.
13:1252015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wu LM, Liu YS, Tong XH, Shen N, Jin RT,
Han H, Hu MH, Wang W and Zhou GX: Inhibition of follicular
development induced by chronic unpredictable stress is associated
with growth and differentiation factor 9 and gonadotropin in mice.
Biol Reprod. 86:1212012. View Article : Google Scholar
|
|
85
|
Sun J, Guo Y, Fan Y, Wang Q, Zhang Q and
Lai D: Decreased expression of IDH1 by chronic unpredictable stress
suppresses proliferation and accelerates senescence of granulosa
cells through ROS activated MAPK signaling pathways. Free Radic
Biol Med. 169:122–136. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Du Y, Ma J, Wu T, Li F, Pan J, Du L, Zhang
M, Diao X and Wu R: Downgrading breast imaging reporting and data
system categories in ultrasound using strain elastography and
computer-aided diagnosis system: A multicenter, prospective study.
Br J Radiol. 97:1653–1660. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kala M and Nivsarkar M: Role of cortisol
and superoxide dismutase in psychological stress induced
anovulation. Gen Comp Endocrinol. 225:117–124. 2016. View Article : Google Scholar
|
|
88
|
Gao Y, Chen F, Kong QQ, Ning SF, Yuan HJ,
Lian HY, Luo MJ and Tan JH: Stresses on female mice impair oocyte
developmental potential: Effects of stress severity and duration on
oocytes at the growing follicle stage. Reprod Sci. 23:1148–1157.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Scott K, Phan TT, Boukelmoune N, Heijnen
CJ and Dantzer R: Chronic restraint stress impairs voluntary wheel
running but has no effect on food-motivated behavior in mice. Brain
Behav Immun. 107:319–329. 2023. View Article : Google Scholar
|
|
90
|
Chen H, Fu S, Li X, Shi M, Qian J, Zhao S,
Yuan P, Ding L, Xia X and Zheng JC: Microglial glutaminase 1
mediates chronic restraint stress-induced depression-like behaviors
and synaptic damages. Signal Transduct Target Ther. 8:4522023.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Chiba S, Numakawa T, Ninomiya M, Richards
MC, Wakabayashi C and Kunugi H: Chronic restraint stress causes
anxiety- and depression-like behaviors, downregulates
glucocorticoid receptor expression, and attenuates glutamate
release induced by brain-derived neurotrophic factor in the
prefrontal cortex. Prog Neuropsychopharmacol Biol Psychiatry.
39:112–119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Buynitsky T and Mostofsky DI: Restraint
stress in biobehavioral research: Recent developments. Neurosci
Biobehav Rev. 33:1089–1098. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Sun J, Guo Y, Zhang Q, Bu S, Li B, Wang Q
and Lai D: Chronic restraint stress disturbs meiotic resumption
through APC/C-mediated cyclin B1 excessive degradation in mouse
oocytes. Cell Cycle. 17:1591–1601. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang SY, Wang JZ, Li JJ, Wei DL, Sui HS,
Zhang ZH, Zhou P and Tan JH: Maternal restraint stress diminishes
the developmental potential of oocytes. Biol Reprod. 84:672–681.
2011. View Article : Google Scholar
|
|
95
|
Geraghty AC, Muroy SE, Zhao S, Bentley GE,
Kriegsfeld LJ and Kaufer D: Knockdown of hypothalamic RFRP3
prevents chronic stress-induced infertility and embryo resorption.
Life. 4:e043162015.
|
|
96
|
Liang B, Wei DL, Cheng YN, Yuan HJ, Lin J,
Cui XZ, Luo MJ and Tan JH: Restraint stress impairs oocyte
developmental potential in mice: Role of CRH-induced apoptosis of
ovarian cells. Biol Reprod. 89:642013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li CY, Li ZB, Kong QQ, Han X, Xiao B, Li
X, Chang ZL and Tan JH: Restraint-induced corticotrophin-releasing
hormone elevation triggers apoptosis of ovarian cells and impairs
oocyte competence via activation of the Fas/FasL system. Biol
Reprod. 99:828–837. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Huang Y, Liu Q, Huang G, Wen J and Chen G:
Hypothalamic kisspeptin neurons regulates energy metabolism and
reproduction under chronic stress. Front Endocrinol (Lausanne).
13:8443972022. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Sugino N, Nakamura Y, Okuno N, Shimamura
K, Teyama T, Ishimatsu M and Kato H: Effects of restraint stress on
luteal function in rats during mid-pregnancy. J Reprod Fertil.
101:23–26. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhou P, Lian HY, Cui W, Wei DL, Li Q, Liu
YX, Liu XY and Tan JH: Maternal-restraint stress increases oocyte
aneuploidy by impairing metaphase I spindle assembly and reducing
spindle assembly checkpoint proteins in mice. Biol Reprod.
86:832012. View Article : Google Scholar
|
|
101
|
Lian HY, Gao Y, Jiao GZ, Sun MJ, Wu XF,
Wang TY, Li H and Tan JH: Antioxidant supplementation overcomes the
deleterious effects of maternal restraint stress-induced oxidative
stress on mouse oocytes. Reproduction. 146:559–568. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Higaki S, Kishi M, Koyama K, Nagano M,
Katagiri S, Takada T and Takahashi Y: Early germinal vesicle
breakdown is a predictor of high preimplantation developmental
competent oocytes in mice. Zygote. 25:41–48. 2017. View Article : Google Scholar
|
|
103
|
Zhao XY, Li ZB, Yuan HJ, Han X, Wu JS,
Feng XY, Zhang M and Tan JH: Restraint stress and elevation of
corticotrophin-releasing hormone in female mice impair oocyte
competence through activation of the tumour necrosis factor α
(TNF-α) system. Reprod Fertil Dev. 32:862–872. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Guo Y, Sun J, Bu S, Li B, Zhang Q, Wang Q
and Lai D: Melatonin protects against chronic stress-induced
oxidative meiotic defects in mice MII oocytes by regulating SIRT1.
Cell Cycle. 19:1677–1695. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Chen RR, Wang J, Zhang M, Kong QQ, Sun GY,
Jin CH and Tan JH: Restraint stress of female mice during oocyte
development facilitates oocyte postovulatory aging. Aging (Albany
NY). 14:9186–9199. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Dorfman M, Arancibia S, Fiedler JL and
Lara HE: Chronic intermittent cold stress activates ovarian
sympathetic nerves and modifies ovarian follicular development in
the rat. Biol Reprod. 68:2038–2043. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Bernuci MP, Szawka RE, Helena CVV, Leite
CM, Lara HE and Anselmo-Franci JA: Locus coeruleus mediates cold
stress-induced polycystic ovary in rats. Endocrinology.
149:2907–2916. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Barra R, Cruz G, Mayerhofer A, Paredes A
and Lara HE: Maternal sympathetic stress impairs follicular
development and puberty of the offspring. Reproduction.
148:137–145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Bernuci MP, Leite CM, Barros P, Kalil B,
Leoni GB, Del Bianco-Borges B, Franci CR, Szawka RE, Lara HE and
Anselmo-Franci JA: Transitory activation of the central and ovarian
norepinephrine systems during cold stress-induced polycystic ovary
in rats. J Neuroendocrinol. 25:23–33. 2013. View Article : Google Scholar
|
|
110
|
Pacak K, Palkovits M, Yadid G, Kvetnansky
R, Kopin IJ and Goldstein DS: Heterogeneous neurochemical responses
to different stressors: A test of Selye's doctrine of
nonspecificity. Am J Physiol. 275:R1247–R1255. 1998.PubMed/NCBI
|
|
111
|
Riquelme R, Ruz F, Mayerhofer A and Lara
HE: Role of ovarian sympathetic nerves and cholinergic local system
during cold stress. J Endocrinol. 242:115–124. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Casillas F, Betancourt M, Juárez-Rojas L,
Ducolomb Y, López A, Ávila-Quintero A, Zamora J, Ommati MM and
Retana-Márquez S: Chronic stress detrimentally affects in vivo
maturation in rat oocytes and oocyte viability at all phases of the
estrous cycle. Animals (Basel). 11:24782021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wang D, Cheng X, Fang H, Ren Y, Li X, Ren
W, Xue B and Yang C: Effect of cold stress on ovarian & uterine
microcirculation in rats and the role of endothelin system. Reprod
Biol Endocrinol. 18:292020. View Article : Google Scholar
|
|
114
|
Retana-Márquez S, Juárez-Rojas L,
Ávila-Quintero A, Rojas-Maya S, Perera G, Casillas F, Betancourt M
and Gómez-Quiroz L: Neuroendocrine disruption is associated to
infertility in chronically stressed female rats. Reprod Biol.
20:474–483. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Keremu A, Yaoliwasi A, Tuerhong M, Kadeer
N, Heyi, Yiming A and Yilike X: Research on the establishment of
chronic stress-induced premature ovarian failure the rat model and
effects of Chinese medicine Muniziqi treatment. Mol Reprod Dev.
86:175–186. 2019. View Article : Google Scholar
|
|
116
|
Ernsberger U, Deller T and Rohrer H: The
sympathies of the body: Functional organization and neuronal
differentiation in the peripheral sympathetic nervous system. Cell
Tissue Res. 386:455–475. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Espinoza JA, Navarrete MI, Linares R,
Chaparro-Ortega A, Ramírez DA, Rosas G, Vieyra E, Domínguez R and
Morales-Ledesma L: Effects of chronic exposure to cold stress on
ovarian functions in prepubertal rats. Reprod Biol. 23:1007562023.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Divyashree S and Yajurvedi HN: Long-term
chronic stress exposure induces PCO phenotype in rat. Reproduction.
152:765–774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Bunsueb S, Lapyuneyong N, Tongpan S, Arun
S and Iamsaard S: Chronic stress increases the tyrosine
phosphorylation in female reproductive organs: An experimental
study. Int J Reprod Biomed. 19:87–96. 2021.PubMed/NCBI
|
|
120
|
Kim J and You S: High housing
density-induced chronic stress diminishes ovarian reserve via
granulosa cell apoptosis by angiotensin II overexpression in mice.
Int J Mol Sci. 23:86142022. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Xi W, Mao H, Cui Z, Yao H, Shi R and Gao
Y: Scream sound-induced chronic psychological stress results in
diminished ovarian reserve in adult female rat. Endocrinology.
163:bqac0422022. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Ghatebi M, Zavareh S, Lashkarbolouki T and
Elahdadi Salmani M: Implications from early life stress on the
development of mouse ovarian follicles: Focus on oxidative stress.
J Obstet Gynaecol Res. 45:1506–1514. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Yuan J, Yu Y, Liu D and Sun Y:
Associations between distinct dimensions of early life adversity
and accelerated reproductive strategy among middle-aged women in
China. Am J Obstet Gynecol. 226:104.e1–104.e14. 2022. View Article : Google Scholar
|
|
124
|
Mishra GD, Cooper R, Tom SE and Kuh D:
Early life circumstances and their impact on menarche and
menopause. Womens Health (Lond). 5:175–190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wang N, Huang Y, Wen J, Su Q, Huang Y, Cai
L, Lin W, Zong L, Huang H, Qian X, et al: Early life exposure to
famine and reproductive aging among Chinese women. Menopause.
26:463–468. 2019. View Article : Google Scholar
|
|
126
|
Liu YX, Cheng YN, Miao YL, Wei DL, Zhao
LH, Luo MJ and Tan JH: Psychological stress on female mice
diminishes the developmental potential of oocytes: A study using
the predatory stress model. PLoS One. 7:e480832012. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Onalan E, Erbay B, Buran İK, Erol D,
Tektemur A, Kuloglu T and Ozercan IH: Effects and mechanism of AP39
on ovarian functions in rats exposed to cisplatin and chronic
immobilization stress. J Menopausal Med. 30:104–119. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Yuan HJ, Li ZB, Zhao XY, Sun GY, Wang GL,
Zhao YQ, Zhang M and Tan JH: Glucocorticoids impair oocyte
competence and trigger apoptosis of ovarian cells via activating
the TNF-α system. Reproduction. 160:129–140. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Becker M, Pinhasov A and Ornoy A: Animal
models of depression: What can they teach Us about the human
disease? Diagnostics (Basel). 11:1232021. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Kendler KS: The genealogy of major
depression: Symptoms and signs of melancholia from 1880 to 1900.
Mol Psychiatry. 22:1539–1553. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Liu MY, Yin CY, Zhu LJ, Zhu XH, Xu C, Luo
CX, Chen H, Zhu DY and Zhou QG: Sucrose preference test for
measurement of stress-induced anhedonia in mice. Nat Protoc.
13:1686–1698. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Primo MJ, Fonseca-Rodrigues D, Almeida A,
Teixeira PM and Pinto-Ribeiro F: Sucrose preference test: A
systematic review of protocols for the assessment of anhedonia in
rodents. Eur Neuropsychopharmacol. 77:80–92. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Verharen JPH, de Jong JW, Zhu Y and Lammel
S: A computational analysis of mouse behavior in the sucrose
preference test. Nat Commun. 14:24192023. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Porsolt RD, Le Pichon M and Jalfre M:
Depression: A new animal model sensitive to antidepressant
treatments. Nature. 266:730–732. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Cryan JF, Mombereau C and Vassout A: The
tail suspension test as a model for assessing antidepressant
activity: Review of pharmacological and genetic studies in mice.
Neurosci Biobehav Rev. 29:571–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Prut L and Belzung C: The open field as a
paradigm to measure the effects of drugs on anxiety-like behaviors:
A review. Eur J Pharmacol. 463:3–33. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Kraeuter AK, Guest PC and Sarnyai Z: The
open field test for measuring locomotor activity and anxiety-like
behavior. Methods Mol Biol. 1916:99–103. 2019. View Article : Google Scholar
|
|
138
|
Seibenhener ML and Wooten MC: Use of the
open field maze to measure locomotor and anxiety-like behavior in
mice. J Vis Exp. e524342015.PubMed/NCBI
|
|
139
|
Bourin M and Hascoët M: The mouse
light/dark box test. Eur J Pharmacol. 463:55–65. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Walf AA and Frye CA: The use of the
elevated plus maze as an assay of anxiety-related behavior in
rodents. Nat Protoc. 2:322–328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Kraeuter AK, Guest PC and Sarnyai Z: The
elevated plus maze test for measuring anxiety-like behavior in
rodents. Methods Mol Biol. 1916:69–74. 2019. View Article : Google Scholar
|
|
142
|
Schrader AJ, Taylor RM, Lowery-Gionta EG
and Moore NLT: Repeated elevated plus maze trials as a measure for
tracking within-subjects behavioral performance in rats (Rattus
norvegicus). PLoS One. 13:e02078042018. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Pietropaolo S: Mood and anxiety-related
phenotypes in mice: Characterization using behavioral tests-Edited
by T. D. Gould. Genes Brain Behav. 9:544. 2010. View Article : Google Scholar
|
|
144
|
Bae J, Lynch CD, Kim S, Sundaram R, Sapra
KJ and Buck Louis GM: Preconception stress and the secondary sex
ratio in a population-based preconception cohort. Fertil Steril.
107:714–722. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Ulrich-Lai YM and Herman JP: Neural
regulation of endocrine and autonomic stress responses. Nat Rev
Neurosci. 10:397–409. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Jacobson L:
Hypothalamic-pituitary-adrenocortical axis regulation. Endocrinol
Metab Clin North Am. 34:271–292. vii2005. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
McCall JG, Al-Hasani R, Siuda ER, Hong DY,
Norris AJ, Ford CP and Bruchas MR: CRH engagement of the locus
coeruleus noradrenergic system mediates stress-induced anxiety.
Neuron. 87:605–620. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Agorastos A and Chrousos GP: The
neuroendocrinology of stress: The stress-related continuum of
chronic disease development. Mol Psychiatry. 27:502–513. 2022.
View Article : Google Scholar
|
|
149
|
Yoo ES, Yu J and Sohn JW: Neuroendocrine
control of appetite and metabolism. Exp Mol Med. 53:505–516. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Di Comite G, Grazia Sabbadini M, Corti A,
Rovere-Querini P and Manfredi AA: Conversation galante: How the
immune and the neuroendocrine systems talk to each other. Autoimmun
Rev. 7:23–29. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Smeenk JMJ, Verhaak CM, Vingerhoets AJJM,
Sweep CGJ, Merkus JMWM, Willemsen SJ, van Minnen A, Straatman H and
Braat DD: Stress and outcome success in IVF: The role of
self-reports and endocrine variables. Hum Reprod. 20:991–996. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Hjollund NHI, Jensen TK, Bonde JP,
Henriksen TB, Andersson AM, Kolstad HA, Ernst E, Giwercman A,
Skakkebaek NE and Olsen J: Distress and reduced fertility: A
follow-up study of first-pregnancy planners. Fertil Steril.
72:47–53. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Ates S, Yesil G, Sevket O, Molla T and
Yildiz S: Comparison of metabolic profile and abdominal fat
distribution between karyotypically normal women with premature
ovarian insufficiency and age matched controls. Maturitas.
79:306–310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Huang Y, Lv Y, Qi T, Luo Z, Meng X, Ying
Q, Li D, Li C, Lan Y, Chu K, et al: Metabolic profile of women with
premature ovarian insufficiency compared with that of age-matched
healthy controls. Maturitas. 148:33–39. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Ferin M: Clinical review 105: Stress and
the reproductive cycle. J Clin Endocrinol Metab. 84:1768–1774.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Chrousos GP, Torpy DJ and Gold PW:
Interactions between the hypothalamic-pituitary-adrenal axis and
the female reproductive system: Clinical implications. Ann Intern
Med. 129:229–240. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Harlow CR, Fahy UM, Talbot WM, Wardle PG
and Hull MG: Stress and stress-related hormones during in-vitro
fertilization treatment. Hum Reprod. 11:274–279. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Demyttenaere K, Nijs P, Evers-Kiebooms G
and Koninckx PR: Coping and the ineffectiveness of coping influence
the outcome of in vitro fertilization through stress responses.
Psychoneuroendocrinology. 17:655–665. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Pal L, Bevilacqua K and Santoro NF:
Chronic psychosocial stressors are detrimental to ovarian reserve:
A study of infertile women. J Psychosom Obstet Gynecol. 31:130–139.
2010. View Article : Google Scholar
|
|
160
|
Andersen CY: Effect of glucocorticoids on
spontaneous and follicle-stimulating hormone induced oocyte
maturation in mouse oocytes during culture. J Steroid Biochem Mol
Biol. 85:423–427. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Yuan HJ, Han X, He N, Wang GL, Gong S, Lin
J, Gao M and Tan JH: Glucocorticoids impair oocyte developmental
potential by triggering apoptosis of ovarian cells via activating
the Fas system. Sci Rep. 6:240362016. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Kalantaridou SN, Zoumakis E,
Makrigiannakis A, Lavasidis LG, Vrekoussis T and Chrousos GP:
Corticotropin-releasing hormone, stress and human reproduction: An
update. J Reprod Immunol. 85:33–39. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Rivier C and Vale W: Influence of
corticotropin-releasing factor on reproductive functions in the
rat. Endocrinology. 114:914–921. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Breen KM, Thackray VG, Hsu T, Mak-McCully
RA, Coss D and Mellon PL: Stress levels of glucocorticoids inhibit
LHβ-subunit gene expression in gonadotrope cells. Mol Endocrinol.
26:1716–1731. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Luo E, Stephens SBZ, Chaing S, Munaganuru
N, Kauffman AS and Breen KM: Corticosterone blocks ovarian
cyclicity and the LH surge via decreased kisspeptin neuron
activation in female mice. Endocrinology. 157:1187–1199. 2016.
View Article : Google Scholar :
|
|
166
|
Roozendaal MM, Swarts JJM, Van Maanen JC,
Wiegant VM and Mattheij JA: Inhibition of the LH surge in cyclic
rats by stress is not mediated by opioids. Life Sci. 60:735–742.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Horowitz MA and Zunszain PA: Neuroimmune
and neuroendocrine abnormalities in depression: Two sides of the
same coin. Ann N Y Acad Sci. 1351:68–79. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Berga SL, Daniels TL and Giles DE: Women
with functional hypothalamic amenorrhea but not other forms of
anovulation display amplified cortisol concentrations. Fertil
Steril. 67:1024–1030. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Deady LD and Sun J: A follicle rupture
assay reveals an essential role for follicular adrenergic signaling
in drosophila ovulation. PLoS Genet. 11:e10056042015. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Aguado LI: Role of the central and
peripheral nervous system in the ovarian function. Microsc Res
Tech. 59:462–473. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Lara HE, Dorfman M, Venegas M, Luza SM,
Luna SL, Mayerhofer A, Guimaraes MA, Rosa E Silva AA and Ramírez
VD: Changes in sympathetic nerve activity of the mammalian ovary
during a normal estrous cycle and in polycystic ovary syndrome:
Studies on norepinephrine release. Microsc Res Tech. 59:495–502.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Morales-Ledesma L, Linares R, Rosas G,
Morán C, Chavira R, Cárdenas M and Domínguez R: Unilateral
sectioning of the superior ovarian nerve of rats with polycystic
ovarian syndrome restores ovulation in the innervated ovary. Reprod
Biol Endocrinol. 8:992010. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Dissen GA, Romero C, Paredes A and Ojeda
SR: Neurotrophic control of ovarian development. Microsc Res Tech.
59:509–515. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Uchida S and Kagitani F: Autonomic nervous
regulation of ovarian function by noxious somatic afferent
stimulation. J Physiol Sci. 65:1–9. 2015. View Article : Google Scholar
|
|
175
|
Acuña E, Fornes R, Fernandois D, Garrido
MP, Greiner M, Lara HE and Paredes AH: Increases in norepinephrine
release and ovarian cyst formation during ageing in the rat. Reprod
Biol Endocrinol. 7:642009. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Casillas F, Flores-González A,
Juárez-Rojas L, López A, Betancourt M, Casas E, Bahena I, Bonilla E
and Retana-Márquez S: Chronic stress decreases fertility parameters
in female rats. Syst Biol Reprod Med. 69:234–244. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Fernandois D, Cruz G, Na EK, Lara HE and
Paredes AH: Kisspeptin level in the aging ovary is regulated by the
sympathetic nervous system. J Endocrinol. 232:97–105. 2017.
View Article : Google Scholar
|
|
178
|
Saller S, Merz-Lange J, Raffael S, Hecht
S, Pavlik R, Thaler C, Berg D, Berg U, Kunz L and Mayerhofer A:
Norepinephrine, active norepinephrine transporter, and
norepinephrine-metabolism are involved in the generation of
reactive oxygen species in human ovarian granulosa cells.
Endocrinology. 153:1472–1483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Zhang L, Gao J and Cui S: miR-21 is
involved in norepinephrine-mediated rat granulosa cell apoptosis by
targeting SMAD7. J Mol Endocrinol. 58:199–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Li XH, Ma YG, Geng LH, Qin L, Hu H and Li
SW: Baseline psychological stress and ovarian norepinephrine levels
negatively affect the outcome of in vitro fertilisation. Gynecol
Endocrinol. 27:139–143. 2011. View Article : Google Scholar
|
|
181
|
Lynch CD, Sundaram R, Maisog JM, Sweeney
AM and Louis GMB: Preconception stress increases the risk of
infertility: Results from a couple-based prospective cohort
study-the LIFE study. Hum Reprod. 29:1067–1075. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Hackett RA and Steptoe A: Type 2 diabetes
mellitus and psychological stress-a modifiable risk factor. Nat Rev
Endocrinol. 13:547–560. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Afrisham R, Paknejad M, Soliemanifar O,
Sadegh-Nejadi S, Meshkani R and Ashtary-Larky D: The influence of
psychological stress on the initiation and progression of diabetes
and cancer. Int J Endocrinol Metab. 17:e674002019.
|
|
184
|
Joseph JJ and Golden SH: Cortisol
dysregulation: The bidirectional link between stress, depression,
and type 2 diabetes mellitus. Ann N Y Acad Sci. 1391:20–34. 2017.
View Article : Google Scholar :
|
|
185
|
Balkan F, Cetin N, Usluogullari CA, Unal
OK and Usluogullari B: Evaluation of the ovarian reserve function
in patients with metabolic syndrome in relation to healthy controls
and different age groups. J Ovarian Res. 7:632014. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Isik S, Ozcan HN, Ozuguz U, Tutuncu YA,
Berker D, Alimli AG, Akbaba G, Karademir MA and Guler S: Evaluation
of ovarian reserve based on hormonal parameters, ovarian volume,
and antral follicle count in women with type 2 diabetes mellitus. J
Clin Endocrinol Metab. 97:261–269. 2012. View Article : Google Scholar
|
|
187
|
Lustman PJ, Anderson RJ, Freedland KE, De
Groot M, Carney RM and Clouse RE: Depression and poor glycemic
control: A meta-analytic review of the literature. Diabetes Care.
23:934–942. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Yaribeygi H, Sathyapalan T, Atkin SL and
Sahebkar A: Molecular mechanisms linking oxidative stress and
diabetes mellitus. Oxid Med Cell Longev. 2020:86092132020.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Grigsby AB, Anderson RJ, Freedland KE,
Clouse RE and Lustman PJ: Prevalence of anxiety in adults with
diabetes: A systematic review. J Psychosom Res. 53:1053–1060. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Razzoli M, Pearson C, Crow S and
Bartolomucci A: Stress, overeating, and obesity: Insights from
human studies and preclinical models. Neurosci Biobehav Rev.
76:154–162. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Codner E, Merino PM and Tena-Sempere M:
Female reproduction and type 1 diabetes: From mechanisms to
clinical findings. Hum Reprod Update. 18:568–585. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Nandi A and Poretsky L: Diabetes and the
female reproductive system. Endocrinol Metab Clin North Am.
42:915–946. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Dri M, Klinger FG and De Felici M: The
ovarian reserve as target of insulin/IGF and ROS in metabolic
disorder-dependent ovarian dysfunctions. Reprod Fertil.
2:R103–R112. 2021. View Article : Google Scholar
|
|
194
|
Barbieri RL, Makris A and Ryan KJ: Effects
of insulin on steroidogenesis in cultured porcine ovarian theca.
Fertil Steril. 40:237–241. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Barbieri RL, Makris A, Randall RW, Daniels
G, Kistner RW and Ryan KJ: Insulin stimulates androgen accumulation
in incubations of ovarian stroma obtained from women with
hyperandrogenism. J Clin Endocrinol Metab. 62:904–910. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Cara JF and Rosenfield RL: Insulin-like
growth factor I and insulin potentiate luteinizing hormone-induced
androgen synthesis by rat ovarian thecal-interstitial cells.
Endocrinology. 123:733–739. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Hudish LI, Reusch JE and Sussel L: β Cell
dysfunction during progression of metabolic syndrome to type 2
diabetes. J Clin Invest. 129:4001–4008. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Escobar-Morreale HF, Roldán B, Barrio R,
Alonso M, Sancho J, De La Calle H and García-Robles R: High
prevalence of the polycystic ovary syndrome and hirsutism in women
with type 1 diabetes mellitus. J Clin Endocrinol Metab.
85:4182–4187. 2000.PubMed/NCBI
|
|
199
|
Diamanti-Kandarakis E, Piperi C, Patsouris
E, Korkolopoulou P, Panidis D, Pawelczyk L, Papavassiliou AG and
Duleba AJ: Immunohistochemical localization of advanced glycation
end-products (AGEs) and their receptor (RAGE) in polycystic and
normal ovaries. Histochem Cell Biol. 127:581–589. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Secomandi L, Borghesan M, Velarde M and
Demaria M: The role of cellular senescence in female reproductive
aging and the potential for senotherapeutic interventions. Hum
Reprod Update. 28:172–189. 2022. View Article : Google Scholar :
|
|
201
|
Willis DS, Watson H, Mason HD, Galea R,
Brincat M and Franks S: Premature response to luteinizing hormone
of granulosa cells from anovulatory women with polycystic ovary
syndrome: Relevance to mechanism of anovulation. J Clin Endocrinol
Metab. 83:3984–3991. 1998.PubMed/NCBI
|
|
202
|
Lee J, Lee HC, Kim SY, Cho GJ and Woodruff
TK: Poorly-controlled type 1 diabetes mellitus impairs LH-LHCGR
signaling in the ovaries and decreases female fertility in mice.
Yonsei Med J. 60:667–678. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Zargar AH, Gupta VK, Wani AI, Masoodi SR,
Bashir MI, Laway BA, Ganie MA and Salahuddin M: Prevalence of
ultrasonography proved polycystic ovaries in North Indian women
with type 2 diabetes mellitus. Reprod Biol Endocrinol. 3:352005.
View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Peppard HR, Marfori J, Iuorno MJ and
Nestler JE: Prevalence of polycystic ovary syndrome among
premenopausal women with type 2 diabetes. Diabetes Care.
24:1050–1052. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Khalil N, Sutton-Tyrrell K, Strotmeyer ES,
Greendale GA, Vuga M, Selzer F, Crandall CJ and Cauley JA:
Menopausal bone changes and incident fractures in diabetic women: A
cohort study. Osteoporos Int. 22:1367–1376. 2011. View Article : Google Scholar
|
|
206
|
Sekhar TVDS, Medarametla S, Rahman A and
Adapa SS: Early menopause in type 2 diabetes-A study from a South
Indian tertiary care centre. J Clin Diagn Res. 9:OC08–OC10.
2015.PubMed/NCBI
|
|
207
|
Qin X, Du J, He R, Li Y, Zhu Q, Li Y, Li H
and Liang X: Adverse effects of type 2 diabetes mellitus on ovarian
reserve and pregnancy outcomes during the assisted reproductive
technology process. Front Endocrinol (Lausanne). 14:12743272023.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Carr MC: The emergence of the metabolic
syndrome with menopause. J Clin Endocrinol Metab. 88:2404–2411.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Kanaya AM, Herrington D, Vittinghoff E,
Lin F, Grady D, Bittner V, Cauley JA and Barrett-Connor E; Heart
and Estrogen/progestin Replacement Study: Glycemic effects of
postmenopausal hormone therapy: The heart and estrogen/progestin
replacement study. A randomized, double-blind, placebo-controlled
trial. Ann Intern Med. 138:1–9. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Margolis KL, Bonds DE, Rodabough RJ,
Tinker L, Phillips LS, Allen C, Bassford T, Burke G, Torrens J and
Howard BV; Women's Health Initiative Investigators: Effect of
oestrogen plus progestin on the incidence of diabetes in
postmenopausal women: Results from the women's health initiative
hormone trial. Diabetologia. 47:1175–1187. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Wagner JD, Thomas MJ, Williams JK, Zhang
L, Greaves KA and Cefalu WT: Insulin sensitivity and cardiovascular
risk factors in ovariectomized monkeys with estradiol alone or
combined with nomegestrol acetate. J Clin Endocrinol Metab.
83:896–901. 1998.PubMed/NCBI
|
|
212
|
Kumagai S, Holmäng A and Björntorp P: The
effects of oestrogen and progesterone on insulin sensitivity in
female rats. Acta Physiol Scand. 149:91–97. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Cantave CY, Ruttle PL, Coté SM, Lupien SJ,
Geoffroy MC, Vitaro F, Brendgen M, Tremblay R, Boivin M and
Ouellet-Morin I: Body mass index across development and adolescent
hair cortisol: The role of persistence, variability, and timing of
exposure. Int J Obes (Lond). Oct 4–2024.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Christakoudi S, Asimakopoulos AG, Riboli E
and Tsilidis KK: Links between the genetic determinants of morning
plasma cortisol and body shape: A two-sample Mendelian
randomisation study. Sci Rep. 14:32302024. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Kühnel A, Hagenberg J, Knauer-Arloth J,
Ködel M, Czisch M, Sämann PG; BeCOME working group; Binder EB and
Kroemer NB: Stress-induced brain responses are associated with BMI
in women. Commun Biol. 6:10312023. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Lin YC, Lin CY, Saffari M, Tsai MC, Chang
YH, Strong C, Chen JK, Hsieh YP, Yang YN and Latner JD: Weight
stigma is associated with body mass index among college students in
Taiwan: The mediated role of internalized weight stigma. BMC
Psychol. 11:3652023. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Fardet L and Fève B: Systemic
glucocorticoid therapy: A review of its metabolic and
cardiovascular adverse events. Drugs. 74:1731–1745. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Santosa A, Rosengren A, Ramasundarahettige
C, Rangarajan S, Gulec S, Chifamba J, Lear SA, Poirier P, Yeates
KE, Yusuf R, et al: Psychosocial risk factors and cardiovascular
disease and death in a population-based cohort from 21 low-,
middle-, and high-income countries. JAMA Netw Open. 4:e21389202021.
View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Ponti F, Santoro A, Mercatelli D,
Gasperini C, Conte M, Martucci M, Sangiorgi L, Franceschi C and
Bazzocchi A: Aging and imaging assessment of body composition: From
fat to facts. Front Endocrinol (Lausanne). 10:8612020. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Van Der Valk ES, Savas M and Van Rossum
EFC: Stress and obesity: Are there more susceptible individuals?
Curr Obes Rep. 7:193–203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Chen X, Xiao Z, Cai Y, Huang L and Chen C:
Hypothalamic mechanisms of obesity-associated disturbance of
hypothalamic-pituitary-ovarian axis. Trends Endocrinol Metab.
33:206–217. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Morimoto A, Rose RD, Smith KM, Dinh DT,
Umehara T, Winstanley YE, Shibahara H, Russell DL and Robker RL:
Granulosa cell metabolism at ovulation correlates with oocyte
competence and is disrupted by obesity and aging. Hum Reprod.
39:2053–2066. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Schon SB, Cabre HE and Redman LM: The
impact of obesity on reproductive health and metabolism in
reproductive-age females. Fertil Steril. 122:194–203. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Martinuzzi K, Ryan S, Luna M and Copperman
AB: Elevated body mass index (BMI) does not adversely affect in
vitro fertilization outcome in young women. J Assist Reprod Genet.
25:169–175. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Bernardi LA, Carnethon MR, de Chavez PJ,
Ikhena DE, Neff LM, Baird DD and Marsh EE: Relationship between
obesity and anti-Müllerian hormone in reproductive-aged African
American women. Obesity (Silver Spring). 25:229–235. 2017.
View Article : Google Scholar
|
|
226
|
Moy V, Jindal S, Lieman H and Buyuk E:
Obesity adversely affects serum anti-müllerian hormone (AMH) levels
in Caucasian women. J Assist Reprod Genet. 32:1305–1311. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Freeman EW, Gracia CR, Sammel MD, Lin H,
Lim LCL and Strauss JF III: Association of anti-mullerian hormone
levels with obesity in late reproductive-age women. Fertil Steril.
87:101–106. 2007. View Article : Google Scholar
|
|
228
|
Wang Y, Wu L, Yang Z, Xu R, Duan Y, Lin J,
Cui X, Fan C, Zhou Y, Bao W, et al: Association of body mass index
with serum anti-Müllerian hormone and inhibin B levels among 8323
women attending a reproductive medical center: A cross-sectional
study. Endocrine. 75:284–292. 2022. View Article : Google Scholar
|
|
229
|
Douchi T, Kuwahata R, Yamamoto S, Oki T,
Yamasaki H and Nagata Y: Relationship of upper body obesity to
menstrual disorders. Acta Obstet Gynecol Scand. 81:147–150. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Rittmaster RS, Deshwal N and Lehman L: The
role of adrenal hyperandrogenism, insulin resistance, and obesity
in the pathogenesis of polycystic ovarian syndrome. J Clin
Endocrinol Metab. 76:1295–1300. 1993.PubMed/NCBI
|
|
231
|
Di Berardino C, Peserico A, Capacchietti
G, Zappacosta A, Bernabò N, Russo V, Mauro A, El Khatib M, Gonnella
F, Konstantinidou F, et al: High-fat diet and female fertility
across lifespan: A comparative lesson from mammal models.
Nutrients. 14:43412022. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Wang N, Luo LL, Xu JJ, Xu MY, Zhang XM,
Zhou XL, Liu WJ and Fu YC: Obesity accelerates ovarian follicle
development and follicle loss in rats. Metabolism. 63:94–103. 2014.
View Article : Google Scholar
|
|
233
|
Jungheim ES, Schoeller EL, Marquard KL,
Louden ED, Schaffer JE and Moley KH: Diet-induced obesity model:
Abnormal oocytes and persistent growth abnormalities in the
offspring. Endocrinology. 151:4039–4046. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
234
|
Luzzo KM, Wang Q, Purcell SH, Chi M,
Jimenez PT, Grindler N, Schedl T and Moley KH: High fat diet
induced developmental defects in the mouse: Oocyte meiotic
aneuploidy and fetal growth retardation/brain defects. PLoS One.
7:e492172012. View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Reynolds KA, Boudoures AL, Chi MMY, Wang Q
and Moley KH: Adverse effects of obesity and/or high-fat diet on
oocyte quality and metabolism are not reversible with resumption of
regular diet in mice. Reprod Fertil Dev. 27:716–724. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Machtinger R, Combelles CMH, Missmer SA,
Correia KF, Fox JH and Racowsky C: The association between severe
obesity and characteristics of failed fertilized oocytes. Hum
Reprod. 27:3198–3207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Kloos J, Perez J and Weinerman R:
Increased body mass index is negatively associated with ovarian
reserve as measured by anti-Müllerian hormone in patients with
polycystic ovarian syndrome. Clin Obes. 29:e126382024. View Article : Google Scholar
|
|
238
|
Beroukhim G, Esencan E and Seifer DB:
Impact of sleep patterns upon female neuroendocrinology and
reproductive outcomes: A comprehensive review. Reprod Biol
Endocrinol. 20:162022. View Article : Google Scholar
|
|
239
|
Yan J, Zhang X, Zhu K, Yu M, Liu Q, De
Felici M, Zhang T, Wang J and Shen W: Sleep deprivation causes gut
dysbiosis impacting on systemic metabolomics leading to premature
ovarian insufficiency in adolescent mice. Theranostics.
14:3760–3776. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Weng L, Hong H, Zhang Q, Xiao C, Zhang Q,
Wang Q, Huang J and Lai D: Sleep deprivation triggers the excessive
activation of ovarian primordial follicles via β2 adrenergic
receptor signaling. Adv Sci (Weinh). 11:e24023932024. View Article : Google Scholar
|
|
241
|
Andersen ML, Alvarenga TAF, Guindalini C,
Perry JC, Silva A, Zager A and Tufik S: Paradoxical sleep
deprivation influences sexual behavior in female rats. J Sex Med.
6:2162–2172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Koehl M, Battle S and Meerlo P: Sex
differences in sleep: The response to sleep deprivation and
restraint stress in mice. Sleep. 29:1224–1231. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Yang CK, Tsai HD, Wu CH and Cho CL: The
role of glucocorticoids in ovarian development of sleep deprived
rats. Taiwan J Obstet Gynecol. 58:122–127. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
244
|
Yang CK, Wu RSC, Wu CH, Lin TRY and Tsai
HD: Sleep deprivation enhances peripheral serotonin secretion to
regulate the large follicle steroidogenesis of rats. Taiwan J
Obstet Gynecol. 54:260–265. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Stock D, Knight JA, Raboud J, Cotterchio
M, Strohmaier S, Willett W, Eliassen AH, Rosner B, Hankinson SE and
Schernhammer E: Rotating night shift work and menopausal age. Hum
Reprod. 34:539–548. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Reiter RJ, Mayo JC, Tan DX, Sainz RM,
Alatorre-Jimenez M and Qin L: Melatonin as an antioxidant: Under
promises but over delivers. J Pineal Res. 61:253–278. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Tamura H, Takasaki A, Taketani T, Tanabe
M, Kizuka F, Lee L, Tamura I, Maekawa R, Asada H, Yamagata Y and
Sugino N: Melatonin as a free radical scavenger in the ovarian
follicle. Endocr J. 60:1–13. 2013. View Article : Google Scholar
|
|
248
|
Lahiri DK, Ge Y, Sharman EH and Bondy SC:
Age-related changes in serum melatonin in mice: Higher levels of
combined melatonin and 6-hydroxymelatonin sulfate in the cerebral
cortex than serum, heart, liver and kidney tissues. J Pineal Res.
36:217–223. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
249
|
Tanabe M, Tamura H, Taketani T, Okada M,
Lee L, Tamura I, Maekawa R, Asada H, Yamagata Y and Sugino N:
Melatonin protects the integrity of granulosa cells by reducing
oxidative stress in nuclei, mitochondria, and plasma membranes in
mice. J Reprod Dev. 61:35–41. 2015. View Article : Google Scholar :
|
|
250
|
Shen M, Cao Y, Jiang Y, Wei Y and Liu H:
Melatonin protects mouse granulosa cells against oxidative damage
by inhibiting FOXO1-mediated autophagy: Implication of an
antioxidation-independent mechanism. Redox Biol. 18:138–157. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Zhai B, Li X, Zhao Z, Cao Y, Liu X, Liu Z,
Ma H and Lu W: Melatonin protects the apoptosis of sheep granulosa
cells by suppressing oxidative stress via MAP3K8 and FOS pathway.
Genes (Basel). 14:10672023. View Article : Google Scholar : PubMed/NCBI
|
|
252
|
Xu G, Dong Y, Wang Z, Ding H, Wang J, Zhao
J, Liu H and Lv W: Melatonin attenuates oxidative stress-induced
apoptosis of bovine ovarian granulosa cells by promoting mitophagy
via SIRT1/FoxO1 signaling pathway. Int J Mol Sci. 24:128542023.
View Article : Google Scholar : PubMed/NCBI
|
|
253
|
Tamura H, Takasaki A, Miwa I, Taniguchi K,
Maekawa R, Asada H, Taketani T, Matsuoka A, Yamagata Y, Shimamura
K, et al: Oxidative stress impairs oocyte quality and melatonin
protects oocytes from free radical damage and improves
fertilization rate. J Pineal Res. 44:280–287. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
254
|
Zhang H, Li C, Wen D, Li R, Lu S, Xu R,
Tang Y, Sun Y, Zhao X, Pan M and Ma B: Melatonin improves the
quality of maternally aged oocytes by maintaining intercellular
communication and antioxidant metabolite supply. Redox Biol.
49:1022152022. View Article : Google Scholar
|
|
255
|
Lord T, Nixon B, Jones KT and Aitken RJ:
Melatonin prevents postovulatory oocyte aging in the mouse and
extends the window for optimal fertilization in vitro. Biol Reprod.
88:672013. View Article : Google Scholar : PubMed/NCBI
|
|
256
|
Zhang M, Lu Y, Chen Y, Zhang Y and Xiong
B: Insufficiency of melatonin in follicular fluid is a reversible
cause for advanced maternal age-related aneuploidy in oocytes.
Redox Biol. 28:1013272020. View Article : Google Scholar
|
|
257
|
Yang D, Mu Y, Wang J, Zou W, Zou H, Yang
H, Zhang C, Fan Y, Zhang H, Zhang H, et al: Melatonin enhances the
developmental potential of immature oocytes from older
reproductive-aged women by improving mitochondrial function.
Heliyon. 9:e193662023. View Article : Google Scholar : PubMed/NCBI
|
|
258
|
Li C, He X, Huang Z, Han L, Wu X, Li L,
Xin Y, Ge J, Sha J, Yin Z and Wang Q: Melatonin ameliorates the
advanced maternal age-associated meiotic defects in oocytes through
the SIRT2-dependent H4K16 deacetylation pathway. Aging (Albany NY).
12:1610–1623. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
259
|
Takasaki A, Nakamura Y, Tamura H,
Shimamura K and Morioka H: Melatonin as a new drug for improving
oocyte quality. Reprod Med Biol. 2:139–144. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
260
|
Baden MY, Hu FB, Vetter C, Schernhammer E,
Redline S and Huang T: Sleep duration patterns in early to middle
adulthood and subsequent risk of type 2 diabetes in women. Diabetes
Care. 43:1219–1226. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
261
|
Rooney KL and Domar AD: The impact of
stress on fertility treatment. Curr Opin Obstet Gynecol.
28:198–201. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Ramezanzadeh F, Noorbala AA, Abedinia N,
Rahimi Forooshani A and Naghizadeh MM: Psychiatric intervention
improved pregnancy rates in infertile couples. Malays J Med Sci.
18:16–24. 2011.PubMed/NCBI
|
|
263
|
Rock J, Tietze C and McLaughlin HB: Effect
of adoption on infertility. Fertil Steril. 16:305–312. 1965.
View Article : Google Scholar : PubMed/NCBI
|
|
264
|
Weir WC and Weir DR: Adoption and
subsequent conceptions. Fertil Steril. 17:283–288. 1966. View Article : Google Scholar : PubMed/NCBI
|
|
265
|
Nakao M, Shirotsuki K and Sugaya N:
Cognitive-behavioral therapy for management of mental health and
stress-related disorders: Recent advances in techniques and
technologies. Biopsychosoc Med. 15:162021. View Article : Google Scholar : PubMed/NCBI
|
|
266
|
Hoffman SG and Smits JAJ:
Cognitive-behavioral therapy for adult anxiety disorders: A
meta-analysis of randomized placebo-controlled trials. J Clin
Psychiatry. 69:621–632. 2008. View Article : Google Scholar
|
|
267
|
Powers MB, Halpern JM, Ferenschak MP,
Gillihan SJ and Foa EB: A meta-analytic review of prolonged
exposure for posttraumatic stress disorder. Clin Psychol Rev.
30:635–641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Olatunji BO, Davis ML, Powers MB and Smits
JAJ: Cognitive-behavioral therapy for obsessive-compulsive
disorder: A meta-analysis of treatment outcome and moderators. J
Psychiatr Res. 47:33–41. 2013. View Article : Google Scholar
|
|
269
|
Cuijpers P, Berking M, Andersson G,
Quigley L, Kleiboer A and Dobson KS: A meta-analysis of
cognitive-behavioural therapy for adult depression, alone and in
comparison with other treatments. Can J Psychiatry. 58:376–385.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
270
|
Hofmann SG, Asnaani A, Vonk IJJ, Sawyer AT
and Fang A: The efficacy of cognitive behavioral therapy: A review
of meta-analyses. Cogn Ther Res. 36:427–440. 2012. View Article : Google Scholar
|
|
271
|
Golshani F, Hasanpour S, Mirghafourvand M
and Esmaeilpour K: Effect of cognitive behavioral therapy-based
counseling on perceived stress in pregnant women with history of
primary infertility: A controlled randomized clinical trial. BMC
Psychiatry. 21:2782021. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Domar AD, Rooney KL, Wiegand B, Orav EJ,
Alper MM, Berger BM and Nikolovski J: Impact of a group mind/body
intervention on pregnancy rates in IVF patients. Fertil Steril.
95:2269–2273. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Faramarzi M, Pasha H, Esmailzadeh S,
Kheirkhah F, Heidary S and Afshar Z: The effect of the cognitive
behavioral therapy and pharmacotherapy on infertility stress: A
randomized controlled trial. Int J Fertil Steril. 7:199–206.
2013.
|
|
274
|
Sexton MB, Byrd MR, O'Donohue WT and
Jacobs NN: Web-based treatment for infertility-related
psychological distress. Arch Womens Ment Health. 13:347–358. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
275
|
John JB, Chellaiyan DVG, Gupta S and
Nithyanandham R: How effective the mindfulness-based cognitive
behavioral therapy on quality of life in women with menopause. J
Midlife Health. 13:169–174. 2022.PubMed/NCBI
|
|
276
|
Green SM and Furtado M: Cognitive
behavioral therapy for sexual concerns during perimenopause: A four
session study protocol. Front Glob Womens Health. 2:7447482021.
View Article : Google Scholar : PubMed/NCBI
|
|
277
|
Green SM, Donegan E, Frey BN, Fedorkow DM,
Key BL, Streiner DL and McCabe RE: Cognitive behavior therapy for
menopausal symptoms (CBT-Meno): A randomized controlled trial.
Menopause. 26:972–980. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
278
|
Green SM, Haber E, McCabe RE and Soares
CN: Cognitive-behavioral group treatment for menopausal symptoms: A
pilot study. Arch Womens Ment Health. 16:325–332. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
279
|
Reddy NV and Omkarappa DB:
Cognitive-behavioral therapy for depression among menopausal woman:
A randomized controlled trial. J Fam Med Prim Care. 8:1002–1006.
2019. View Article : Google Scholar
|
|
280
|
Donegan E, Frey BN, McCabe RE, Streiner
DL, Fedorkow DM, Furtado M and Green SM: Impact of the CBT-Meno
protocol on menopause-specific beliefs, dysfunctional attitudes,
and coping behaviors. Menopause. 29:963–972. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
281
|
Shabani F, Farvareshi M, Hamdi K,
Sadeghzadeh Oskouei B, Montazeri M and Mirghafourvand M: The effect
of cognitive-behavioral therapy on stress and anxiety of women with
premature ovarian insufficiency: A randomized controlled trial.
Post Reprod Health. 28:211–221. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Berga SL, Marcus MD, Loucks TL, Hlastala
S, Ringham R and Krohn MA: Recovery of ovarian activity in women
with functional hypothalamic amenorrhea who were treated with
cognitive behavior therapy. Fertil Steril. 80:976–981. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
283
|
Michopoulos V, Mancini F, Loucks TL and
Berga SL: Neuroendocrine recovery initiated by cognitive behavioral
therapy in women with functional hypothalamic amenorrhea: A
randomized, controlled trial. Fertil Steril. 99:2084–2091.e1. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
284
|
Sockalingam S, Leung SE, Ma C, Tomlinson
G, Hawa R, Wnuk S, Jackson T, Urbach D, Okrainec A, Brown J, et al:
Efficacy of telephone-based cognitive behavioral therapy for weight
loss, disordered eating, and psychological distress after bariatric
surgery: A randomized clinical trial. JAMA Netw Open.
6:e23270992023. View Article : Google Scholar : PubMed/NCBI
|
|
285
|
Rofey DL, Szigethy EM, Noll RB, Dahl RE,
Lobst E and Arslanian SA: Cognitive-behavioral therapy for physical
and emotional disturbances in adolescents with polycystic ovary
syndrome: A pilot study. J Pediatr Psychol. 34:156–163. 2009.
View Article : Google Scholar
|
|
286
|
Cooney LG, Milman LW, Hantsoo L, Kornfield
S, Sammel MD, Allison KC, Epperson CN and Dokras A:
Cognitive-behavioral therapy improves weight loss and quality of
life in women with polycystic ovary syndrome: A pilot randomized
clinical trial. Fertil Steril. 110:161–171.e1. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
287
|
Ludwig DS and Kabat-Zinn J: Mindfulness in
medicine. JAMA. 300:1350–1352. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
288
|
Grossman P, Niemann L, Schmidt S and
Walach H: Mindfulness-based stress reduction and health benefits. A
meta-analysis. J Psychosom Res. 57:35–43. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
289
|
Rosenzweig S, Greeson JM, Reibel DK, Green
JS, Jasser SA and Beasley D: Mindfulness-based stress reduction for
chronic pain conditions: Variation in treatment outcomes and role
of home meditation practice. J Psychosom Res. 68:29–36. 2010.
View Article : Google Scholar
|
|
290
|
Wong C, Yip BHK, Gao T, Lam KYY, Woo DMS,
Yip ALK, Chin CY, Tang WPY, Choy MMT, Tsang KWK, et al:
Mindfulness-based stress reduction (MBSR) or psychoeducation for
the reduction of menopausal symptoms: A randomized, controlled
clinical trial. Sci Rep. 8:66092018. View Article : Google Scholar : PubMed/NCBI
|
|
291
|
Pyri F, Abedi P, Maraghi E and Jefreh M:
The effect of mindfulness on quality of life among women with
premature ovarian insufficiency: A randomized clinical trial. J
Midlife Health. 12:116–121. 2021.PubMed/NCBI
|
|
292
|
Majeed MH, Ali AA and Sudak DM:
Mindfulness-based interventions for chronic pain: Evidence and
applications. Asian J Psychiatry. 32:79–83. 2018. View Article : Google Scholar
|
|
293
|
Kalhori F, Masoumi SZ, Shamsaei F,
Mohammadi Y and Yavangi M: Effect of mindfulness-based group
counseling on depression in infertile women: Randomized clinical
trial study. Int J Fertil Steril. 14:10–6. 2020.PubMed/NCBI
|
|
294
|
Galhardo A, Cunha M and Pinto-Gouveia J: A
7-year follow-up study of the mindfulness-based program for
infertility: Are there long-term effects? Clin Psychol Psychother.
26:409–417. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
295
|
Aalbers S, Fusar-Poli L, Freeman RE,
Spreen M, Ket JC, Vink AC, Maratos A, Crawford M, Chen XJ and Gold
C: Music therapy for depression. Cochrane Database Syst Rev.
11:CD0045172017.PubMed/NCBI
|
|
296
|
Koçak DY and Varişoğlu Y: The effect of
music therapy on menopausal symptoms and depression: A
randomized-controlled study. Menopause. 29:545–552. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
297
|
Kim S, Kim SM, Hwang H, Kim MK, Kim HJ,
Park S and Han DH: The effects of music therapy on the
psychological status of women with perimenopause syndrome.
Menopause. 30:1045–1052. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
298
|
Mahmoud MY, Labib K, Sileem SA, Mustafa
FA, Hamed WM, Abd Elhamid A, Saleh DM, Alanwar A, Riad AAM,
Abdelhakim AM, et al: The impact of music therapy on anxiety and
pregnancy rate among infertile women undergoing assisted
reproductive technologies: A systematic review and meta-analysis. J
Psychosom Obstet Gynecol. 43:205–213. 2022. View Article : Google Scholar
|
|
299
|
Aba YA, Avci D, Guzel Y, Ozcelik SK and
Gurtekin B: Effect of music therapy on the anxiety levels and
pregnancy rate of women undergoing in vitro fertilization-embryo
transfer: A randomized controlled trial. Appl Nurs Res. 36:19–24.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
300
|
Domar AD, Gross J, Rooney K and Boivin J:
Exploratory randomized trial on the effect of a brief psychological
intervention on emotions, quality of life, discontinuation, and
pregnancy rates in in vitro fertilization patients. Fertil Steril.
104:440–451.e7. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
301
|
Clifton J, Parent J, Seehuus M, Worrall G,
Forehand R and Domar A: An internet-based mind/body intervention to
mitigate distress in women experiencing infertility: A randomized
pilot trial. PLoS One. 15:e02293792020. View Article : Google Scholar : PubMed/NCBI
|
|
302
|
Brinsley J, Schuch F, Lederman O, Girard
D, Smout M, Immink MA, Stubbs B, Firth J, Davison K and Rosenbaum
S: Effects of yoga on depressive symptoms in people with mental
disorders: A systematic review and meta-analysis. Br J Sports Med.
55:992–1000. 2021. View Article : Google Scholar
|
|
303
|
Darbandi S, Darbandi M, Khorram Khorshid
HR and Sadeghi MR: Yoga can improve assisted reproduction
technology outcomes in couples with infertility. Altern Ther Health
Med. 24:50–55. 2018.
|
|
304
|
Oron G, Allnutt E, Lackman T, Sokal-Arnon
T, Holzer H and Takefman J: A prospective study using hatha yoga
for stress reduction among women waiting for IVF treatment. Reprod
Biomed Online. 30:542–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
305
|
Valoriani V, Lotti F, Vanni C, Noci MC,
Fontanarosa N, Ferrari G, Cozzi C and Noci I: Hatha-yoga as a
psychological adjuvant for women undergoing IVF: A pilot study. Eur
J Obstet Gynecol Reprod Biol. 176:158–162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
306
|
Lu X, Liu L and Yuan R: Effect of the
information support method combined with yoga exercise on the
depression, anxiety, and sleep quality of menopausal women.
Psychiatr Danub. 32:380–388. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
307
|
Jorge MP, Santaella DF, Pontes IMO,
Shiramizu VKM, Nascimento EB, Cabral A, Lemos TM, Silva RH and
Ribeiro AM: Hatha yoga practice decreases menopause symptoms and
improves quality of life: A randomized controlled trial. Complement
Ther Med. 26:128–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
308
|
Frederiksen Y, O'Toole MS, Mehlsen MY,
Hauge B, Elbaek HO, Zachariae R and Ingerslev HJ: The effect of
expressive writing intervention for infertile couples: a randomized
controlled trial. Hum Reprod. 32:391–402. 2017. View Article : Google Scholar
|
|
309
|
Hart RJ: Physiological aspects of female
fertility: Role of the environment, modern lifestyle, and genetics.
Physiol Rev. 96:873–909. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
310
|
Waldman M, Cohen K, Yadin D, Nudelman V,
Gorfil D, Laniado-Schwartzman M, Kornwoski R, Aravot D, Abraham NG,
Arad M and Hochhauser E: Regulation of diabetic cardiomyopathy by
caloric restriction is mediated by intracellular signaling pathways
involving 'SIRT1 and PGC-1α'. Cardiovasc Diabetol. 17:1112018.
View Article : Google Scholar
|
|
311
|
Bordone L and Guarente L: Calorie
restriction, SIRT1 and metabolism: Understanding longevity. Nat Rev
Mol Cell Biol. 6:298–305. 2005. View Article : Google Scholar
|
|
312
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
313
|
Selesniemi K, Lee HJ, Muhlhauser A and
Tilly JL: Prevention of maternal aging-associated oocyte aneuploidy
and meiotic spindle defects in mice by dietary and genetic
strategies. Proc Natl Acad Sci USA. 108:12319–12324. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
314
|
Smits A, Marei WFA, Moorkens K, Bols PEJ,
De Neubourg D and Leroy JLMR: Obese outbred mice only partially
benefit from diet normalization or calorie restriction as
preconception care interventions to improve metabolic health and
oocyte quality. Hum Reprod. 37:2867–2884. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
315
|
Mishina T, Tabata N, Hayashi T, Yoshimura
M, Umeda M, Mori M, Ikawa Y, Hamada H, Nikaido I and Kitajima TS:
Single-oocyte transcriptome analysis reveals aging-associated
effects influenced by life stage and calorie restriction. Aging
Cell. 20:e134282021. View Article : Google Scholar : PubMed/NCBI
|
|
316
|
Słuczanowska-Głąbowska S, Laszczyńska M,
Piotrowska K, Grabowska M, Grymuła K and Ratajczak MZ: Caloric
restriction increases ratio of estrogen to androgen receptors
expression in murine ovaries-potential therapeutic implications. J
Ovarian Res. 8:572015. View Article : Google Scholar
|
|
317
|
Sharov AA, Falco G, Piao Y, Poosala S,
Becker KG, Zonderman AB, Longo DL, Schlessinger D and Ko MSh:
Effects of aging and calorie restriction on the global gene
expression profiles of mouse testis and ovary. BMC Biol. 6:242008.
View Article : Google Scholar : PubMed/NCBI
|
|
318
|
Garcia DN, Saccon TD, Pradiee J, Rincón
JAA, Andrade KRS, Rovani MT, Mondadori RG, Cruz LAX, Barros CC,
Masternak MM, et al: Effect of caloric restriction and rapamycin on
ovarian aging in mice. Geroscience. 41:395–408. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
319
|
Isola JVV, Zanini BM, Hense JD,
Alvarado-Rincón JA, Garcia DN, Pereira GC, Vieira AD, Oliveira TL,
Collares T, Gasperin BG, et al: Mild calorie restriction, but not
17α-estradiol, extends ovarian reserve and fertility in female
mice. Exp Gerontol. 159:1116692022. View Article : Google Scholar
|
|
320
|
Liu WJ, Zhang XM, Wang N, Zhou XL, Fu YC
and Luo LL: Calorie restriction inhibits ovarian follicle
development and follicle loss through activating SIRT1 signaling in
mice. Eur J Med Res. 20:222015. View Article : Google Scholar : PubMed/NCBI
|
|
321
|
Zhou XL, Xu JJ, Ni YH, Chen XC, Zhang HX,
Zhang XM, Liu WJ, Luo LL and Fu YC: SIRT1 activator (SRT1720)
improves the follicle reserve and prolongs the ovarian lifespan of
diet-induced obesity in female mice via activating SIRT1 and
suppressing mTOR signaling. J Ovarian Res. 7:972014. View Article : Google Scholar : PubMed/NCBI
|
|
322
|
Li L, Fu YC, Xu JJ, Lin XH, Chen XC, Zhang
XM and Luo LL: Caloric restriction promotes the reserve of follicle
pool in adult female rats by inhibiting the activation of mammalian
target of rapamycin signaling. Reprod Sci. 22:60–67. 2015.
View Article : Google Scholar :
|
|
323
|
Smits A, Marei WFA, De Neubourg D and
Leroy JLMR: Diet normalization or caloric restriction as a
preconception care strategy to improve metabolic health and oocyte
quality in obese outbred mice. Reprod Biol Endocrinol. 19:1662021.
View Article : Google Scholar : PubMed/NCBI
|
|
324
|
Chen G, Xie W, Nah J, Sauvat A, Liu P,
Pietrocola F, Sica V, Carmona-Gutierrez D, Zimmermann A, Pendl T,
et al: 3,4-Dimethoxychalcone induces autophagy through activation
of the transcription factors TFE3 and TFEB. EMBO Mol Med.
11:e104692019. View Article : Google Scholar : PubMed/NCBI
|
|
325
|
Kepp O, Chen G, Carmona-Gutierrez D, Madeo
F and Kroemer G: A discovery platform for the identification of
caloric restriction mimetics with broad health-improving effects.
Autophagy. 16:188–189. 2020. View Article : Google Scholar :
|
|
326
|
Zhang H, Ni W, Yu G, Geng Y, Lou J, Qi J,
Chen Y, Li F, Ye H, Ma H, et al: 3,4-Dimethoxychalcone, a caloric
restriction mimetic, enhances TFEB-mediated autophagy and
alleviates pyroptosis and necroptosis after spinal cord injury.
Theranostics. 13:810–832. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
327
|
Jafari M, Macho-González A, Diaz A,
Lindenau K, Santiago-Fernández O, Zeng M, Massey AC, de Cabo R,
Kaushik S and Cuervo AM: Calorie restriction and
calorie-restriction mimetics activate chaperone-mediated autophagy.
Proc Natl Acad Sci USA. 121:e23179451212024. View Article : Google Scholar : PubMed/NCBI
|
|
328
|
Sciarretta S, Forte M, Castoldi F, Frati
G, Versaci F, Sadoshima J, Kroemer G and Maiuri MC: Caloric
restriction mimetics for the treatment of cardiovascular diseases.
Cardiovasc Res. 117:1434–1449. 2021. View Article : Google Scholar
|
|
329
|
Pallauf K, Günther I, Kühn G, Chin D, de
Pascual-Teresa S and Rimbach G: The potential of resveratrol to act
as a caloric restriction mimetic appears to be limited: Insights
from studies in mice. Adv Nutr. 12:995–1005. 2021. View Article : Google Scholar :
|
|
330
|
Pernicova I and Korbonits M:
Metformin-mode of action and clinical implications for diabetes and
cancer. Nat Rev Endocrinol. 10:143–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
331
|
Viollet B, Guigas B, Garcia NS, Leclerc J,
Foretz M and Andreelli F: Cellular and molecular mechanisms of
metformin: An overview. Clin Sci (Lond). 122:253–270. 2012.
View Article : Google Scholar
|
|
332
|
Masoudi FA, Wang Y, Inzucchi SE, Setaro
JF, Havranek EP, Foody JM and Krumholz HM: Metformin and
thiazolidinedione use in medicare patients with heart failure.
JAMA. 290:81–85. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
333
|
Valencia WM, Palacio A, Tamariz L and
Florez H: Metformin and ageing: Improving ageing outcomes beyond
glycaemic control. Diabetologia. 60:1630–1638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
334
|
Morales DR and Morris AD: Metformin in
cancer treatment and prevention. Annu Rev Med. 66:17–29. 2015.
View Article : Google Scholar
|
|
335
|
Qin X, Du D, Chen Q, Wu M, Wu T, Wen J,
Jin Y, Zhang J and Wang S: Metformin prevents murine ovarian aging.
Aging (Albany NY). 11:3785–3794. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
336
|
Huang CC, Chou CH, Yang YS, Ho HN, Shun
CT, Wen WF, Chen SU and Chen MJ: Metformin: A novel promising
option for fertility preservation during cyclophosphamide-based
chemotherapy. Mol Hum Reprod. 27:gaaa0842021. View Article : Google Scholar : PubMed/NCBI
|
|
337
|
Cao Y, Wang Z, Zhang C, Bian Y, Zhang X,
Liu X, Chen W and Zhao Y: Metformin promotes in vitro maturation of
oocytes from aged mice by attenuating mitochondrial oxidative
stress via SIRT3-dependent SOD2ac. Front Cell Dev Biol.
10:10285102022. View Article : Google Scholar : PubMed/NCBI
|
|
338
|
Ayhan S, Hancerliogullari N, Guney G,
Gozukucuk M, Caydere M, Guney SS, Tokmak A and Ustun Y: Does the
addition of metformin to carboplatin treatment decreases ovarian
reserve damage associated with carboplatin usage? J Ovarian Res.
16:1842023. View Article : Google Scholar : PubMed/NCBI
|
|
339
|
Landry DA, Yakubovich E, Cook DP, Fasih S,
Upham J and Vanderhyden BC: Metformin prevents age-associated
ovarian fibrosis by modulating the immune landscape in female mice.
Sci Adv. 8:eabq14752022. View Article : Google Scholar : PubMed/NCBI
|
|
340
|
Gambini J, Inglés M, Olaso G, Lopez-Grueso
R, Bonet-Costa V, Gimeno-Mallench L, Mas-Bargues C, Abdelaziz KM,
Gomez-Cabrera MC, Vina J and Borras C: Properties of resveratrol:
In vitro and in vivo studies about metabolism, bioavailability, and
biological effects in animal models and humans. Oxid Med Cell
Longev. 2015:8370422015. View Article : Google Scholar : PubMed/NCBI
|
|
341
|
Rajman L, Chwalek K and Sinclair DA:
Therapeutic potential of NAD-boosting molecules: The in vivo
evidence. Cell Metabv. 27:529–547. 2018. View Article : Google Scholar
|
|
342
|
Côté CD, Rasmussen BA, Duca FA,
Zadeh-Tahmasebi M, Baur JA, Daljeet M, Breen DM, Filippi BM and Lam
TK: Resveratrol activates duodenal Sirt1 to reverse insulin
resistance in rats through a neuronal network. Nat Med. 21:498–505.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
343
|
Timmers S, Konings E, Bilet L, Houtkooper
RH, van de Weijer T, Goossens GH, Hoeks J, van der Krieken S, Ryu
D, Kersten S, et al: Calorie restriction-like effects of 30 days of
resveratrol supplementation on energy metabolism and metabolic
profile in obese humans. Cell Metab. 14:612–622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
344
|
Battaglia R, Caponnetto A, Caringella AM,
Cortone A, Ferrara C, Smirni S, Iannitti R, Purrello M, D'Amato G,
Fioretti B and Di Pietro C: Resveratrol treatment induces
mito-miRNome modification in follicular fluid from aged women with
a poor prognosis for in vitro fertilization cycles. Antioxidants
(Basel). 11:10192022. View Article : Google Scholar : PubMed/NCBI
|
|
345
|
Li R, Li E, Kamili G, Ou S and Yang D:
Effect of resveratrol on superovulation in mice. Biomed
Pharmacother. 146:1125652022. View Article : Google Scholar
|
|
346
|
Gou M, Li J, Yi L, Li H, Ye X, Wang H, Liu
L, Sun B, Zhang S, Zhu Z, et al: Reprogramming of ovarian aging
epigenome by resveratrol. PNAS Nexus. 2:pgac3102022. View Article : Google Scholar
|
|
347
|
Wu H, Xue J, Di H, Lv C, Hao Y and Nie Z:
Resveratrol improves ovarian function in aged rat by inhibiting
oxidative stress and activating the Sirt1. Gen Physiol Biophys.
41:53–61. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
348
|
Zhou J, Xue Z, He HN, Liu X, Yin SY, Wu
DY, Zhang X, Schatten H and Miao YL: Resveratrol delays
postovulatory aging of mouse oocytes through activating mitophagy.
Aging (Albany NY). 11:11504–11519. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
349
|
Liang QX, Lin YH, Zhang CH, Sun HM, Zhou
L, Schatten H, Sun QY and Qian WP: Resveratrol increases resistance
of mouse oocytes to postovulatory aging in vivo. Aging (Albany NY).
10:1586–1596. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
350
|
Liu MJ, Sun AG, Zhao SG, Liu H, Ma SY, Li
M, Huai YX, Zhao H and Liu HB: Resveratrol improves in vitro
maturation of oocytes in aged mice and humans. Fertil Steril.
109:900–907. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
351
|
Okamoto N, Sato Y, Kawagoe Y, Shimizu T
and Kawamura K: Short-term resveratrol treatment restored the
quality of oocytes in aging mice. Aging (Albany NY). 14:5628–5640.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
352
|
Sun YL, Tang SB, Shen W, Yin S and Sun QY:
Roles of resveratrol in improving the quality of postovulatory
aging oocytes in vitro. Cells. 8:11322019. View Article : Google Scholar : PubMed/NCBI
|
|
353
|
Cai M, Sun H, Huang Y, Yao H, Zhao C, Wang
J and Zhu H: Resveratrol protects rat ovarian luteinized granulosa
cells from H2O2 -induced dysfunction by
activating autophagy. Int J Mol Sci. 24:109142023. View Article : Google Scholar
|
|
354
|
Moreira-Pinto B, Costa L, Felgueira E,
Fonseca BM and Rebelo I: Low doses of resveratrol protect human
granulosa cells from induced-oxidative stress. Antioxidants
(Basel). 10:5612021. View Article : Google Scholar : PubMed/NCBI
|
|
355
|
Pruessner M, Hellhammer DH, Pruessner JC
and Lupien SJ: Self-reported depressive symptoms and stress levels
in healthy young men: Associations with the cortisol response to
awakening. Psychosom Med. 65:92–99. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
356
|
Campagne DM: Should fertilization
treatment start with reducing stress? Hum Reprod. 21:1651–1658.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
357
|
Obayashi K: Salivary mental stress
proteins. Clin Chim Acta. 425:196–201. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
358
|
Shah K, Kumari R and Jain M: Unveiling
stress markers: A systematic review investigating psychological
stress biomarkers. Dev Psychobiol. 66:e224902024. View Article : Google Scholar : PubMed/NCBI
|
|
359
|
Lavalle S, Masiello E, Iannella G,
Magliulo G, Pace A, Lechien JR, Calvo-Henriquez C, Cocuzza S,
Parisi FM, Favier V, et al: Unraveling the complexities of
oxidative stress and inflammation biomarkers in obstructive sleep
apnea syndrome: A comprehensive review. Life (Basel).
14:4252024.PubMed/NCBI
|
|
360
|
Ok J, Park S, Jung YH and Kim T: Wearable
and implantable cortisol-sensing electronics for stress monitoring.
Adv Mater. 36:22115952024. View Article : Google Scholar
|
|
361
|
Damone AL, Joham AE, Loxton D, Earnest A,
Teede HJ and Moran LJ: Depression, anxiety and perceived stress in
women with and without PCOS: A community-based study. Psychol Med.
49:1510–1520. 2019. View Article : Google Scholar
|
|
362
|
Tay CT, Teede HJ, Hill B, Loxton D and
Joham AE: Increased prevalence of eating disorders, low
self-esteem, and psychological distress in women with polycystic
ovary syndrome: A community-based cohort study. Fertil Steril.
112:353–361. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
363
|
Rowlands IJ, Teede H, Lucke J, Dobson AJ
and Mishra GD: Young women's psychological distress after a
diagnosis of polycystic ovary syndrome or endometriosis. Hum
Reprod. 31:2072–2081. 2016. View Article : Google Scholar : PubMed/NCBI
|