Introduction
Mammalian cells evolved and constantly live in a
highly reactive oxidative environment. In the mammalian organism
H2O2 has a central position within the
reactive oxygen species (ROS) family. Its formation by several
reactions and its controlled inactivation is the basis of redox
homeostasis (1,2), since free radicals are highly
cytotoxic. Oxidative stress is the result of interactions with
macromolecules among highly reactive hydrogen peroxide
(H2O2) and singlet oxygen
(1O2), the radicals superoxide anion
(O2−) and the hydroxyl radical (HO.). Besides
the major interest in identification and advancement of compounds
which are either radical scavengers or antioxidants, ROS can
alternate between a positive and negative cellular outcomes
(3). Insufficient ROS defense
mechanisms are mutagenic and promote cell death, apoptosis and
autophagy (4). Recently autophagy
and apoptosis stimulus by the presence of an
H2O2-induced pathway in human primary and
tumor cell lines and in primary cells (5), was demonstrated. Indeed, ROS may
cause unfavorable cellular outcome or a positive one to eliminate
DNA damaged cells and thus preventing carcinogenesis. From a
mechanistic point of view, a clear and elegant example of the ROS
pivotal effect on cellular outcome has been described by tumor
necrosis factor (TNF-α) binding to tumor necrosis factor receptor 1
(TNFR1). This binding can induce cell proliferation and cell death,
both mediated, by Jun N-terminal kinase (JNK) activation which is
enhanced by ROS production (6).
The natural polyamines (PA), putrescine (PUT), spermidine (SPD) and
spermine (SPM) are ubiquitous low-molecular weight aliphatic
cations essential for eukaryotic cells (7,8) and
tumor cell PA depletion was associated with the downregulation of
Bcl-2 protein and an increase of reactive oxygen species (9,10).
To adequately preserve concentrations, PA metabolism is strictly
regulated by the interconversion stepwise degradation, which is
responsible for the oxidant by-products (8,11).
Increasing interest has been posed on the SPM amino oxidase (SMOX)
and BSAO enzyme activities, since SPM catabolic degradation has
been found closely related to DNA oxidation and apoptosis, mainly
via hydrogen peroxide (H2O2) production
(12–15). High SMOX activity provokes low
level of SPM and the inhibition of the interactions with DNA, thus
causing sensitivity to ionizing radiation exposure and cell death
(15,16). It was also demonstrated that
H2O2 formation caused by bovine serum
spermine oxidase (BSAO) (enzyme able to deaminate endogenous
polyamines generating H2O2 and aldehyde)
delivers deleterious effects in several human cancer cellular
models, noticeable greater in multidrug resistance (MDR) tumor
cells than in their wild-type counterpart (17–19).
Since a variety of tumor cells, including MDR cells (P-glycoprotein
expressing phenotype) produce very high amounts of ROS (13,20)
and in vivo many tumors appear resistant to oxidative stress
and apoptosis, we dissected the influence of chronic sub-lethal DNA
damage and DNA repair by mSMOX ectopically overexpressed in
proficient Chinese hamster AA8 cell line and both deficient
base-excision-repair (BER) EM9 (21) and transcription-coupled
nucleotide-excision-repair (NER) UV61 (22) cell lines to represent cellular
models of priming damage dose of ROS. Low doses of X-irradiation
delivers challenging dose of damage evaluated at 6 and 24 h after
exposure. Summarizing, the priming dose of ROS over-exposure by
mSMOX provokes an adaptive response in N18TG2, AA8 and EM9 cell
lines at 24 h. In the UV61 cell line ROS, mSMOX did not deliver an
adaptive response to radiation. According to Pelicano et
al(23) the escalated ROS
generation in cancer cells serves as an endogenous source of
DNA-damaging agents, which promote genetic instability and
development of drug resistance. Therefore, on the basis of an
increased ROS production, from SPM enzymatic catabolism, in cancer
cells therapeutic strategies have been suggested, which rely on the
assumption that cancer cells, mainly MDR cells, are more sensitive
to additional exposure to radicals than WT cells (12,23).
Once again, the pivotal roles played by SMOX and PA catabolism seem
to evoke the biological processes of stress response, wherein
balance is mandatory to live or to die (3). Thus, these alterations could
represent a multitasking anticancer strategy, addressed not only to
PA metabolism, but involving also radiation biology.
Materials and methods
Cell culture, radiation exposure and
reagents
Reagents were from Sigma-Aldrich (Sigma-Aldrich, St.
Louis, MO, USA), unless otherwise specified. Plastic-wares were
from Nunc (Nunc A/S, Roskilde, Denmark). A human colon
adenocarcinoma cell line (LoVo WT), isolated from a metastatic
nodule and its doxorubicin (DOX)-resistant variant (LoVo DX) were
used in this study. Both cell lines were grown in monolayer in
Ham’s F12 medium (Gibco BRL, Life Technologies, Paisley, UK)
supplemented with 10% fetal bovine serum (FBS) (Hyclone,
Cramlington NE23, UK), 1% L-glutamine (Gibco BRL, Life
Technologies), 1% penicillin (50 U/ml)-streptomycin (50 μg/ml)
(Gibco BRL, Life Technologies), 1% vitamins (Gibco BRL, Life
Technologies) in a humidified atmosphere of 5% CO2 in a
water-jacketed incubator at 37°C. The pleiotropic drug-resistant
cell line LoVo DX was obtained from its drug-sensitive parental
LoVo cell line, by exposure to increasing concentrations of DOX
(Adriblastina, Pharmacia and Upjohn, Milan, Italy) (24). Transfection and growth conditions
of pcDNA3 transfected (NP), pcDNA3-SMOX transfected (NS) and mouse
NB cell lines have been described (15,16).
AA8, EM9 and UV61 cell lines (21,22)
were a generous gift of Professor F. Palitti (Universita’ della
Tuscia, Italy). Stable transfection with pcDNA3 and pcDNA3-SMOX
were performed with Effectene (Qiagen) as described (16). Geneticin (G418) (300 mM) was the
selection agent to isolate stable transfected pool of cells.
X-irradiation of 1 and 10 cGy, were delivered by a Gilardoni CHF
320 G Unit (Gilardoni S.p.A., Mandello L., Italy) tested and
complying with EU standards. Dose/rate was 0.1 Gy/min at 250 KeV,
1,5 A, with 0.5 mm Cu filter. Cells were irradiated on ice and
fresh medium was replaced after exposure. Control cells were
treated similarly, without irradiation. All experimental points
were taken 6 and 24 h after irradiation.
NP, NS, AA8, EM0 and UV61 proliferation
assay (MTT)
To analyse cell proliferation we followed the
instructions of the manufacturer of the Cell Proliferation Kit I
(Roche). We performed triplicate experiments with increasing number
of cell/plate (103, 2×103, 3×103
cell/plate) normalised to 103. Cells were seeded the day
before the experiments and treated with the MTT solution for 2 h at
growth conditions. Isopropanol (0.1 ml) with 0.04 N HCl was added
to each well to quench the red phenol colour and the absorbance was
measured on an ELISA plate reader with a test wavelength of 570 nm
and a reference wavelength of 630 nm.
Reactive oxidative species and flow
cytometry
Level of intracellular hydrogen peroxide was
determined by flow cytometry (FCM) analysis of fluorescence
intensity of 2′,7′-dichlorodihydroflurescein (H2DCFDA)
(Invitrogen). Briefly, cells were treated with H2DCFDA
for 30 min. At least 2×104 cells were analyzed by a
FACSCalibur flow cytometer (Becton-Dickinson, San Josè, CA, USA),
with laser excitation set at 495 nm and a 525-nm emission filter to
detect green fluorescence. The negative control was obtained
omitting fluorescence probe-mix from the reaction and
auto-fluorescence was estimated. To study DNA content, cells were
treated with propidium iodide (PI). At least 105 cells
were analyzed by FACSCalibur previously calibrated by CaliBRITE 3
beads (Becton-Dickinson), with laser excitation set at 488 nm and a
630-nm emission filter to detect red fluorescence. Level of
3′-terminal deoxy-transferase (TdT) by TUNEL method was used to
detect apoptosis, following the manufacturer instructions (In
Situ Cell Death Fluorescent kit, Roche Diagnostic S.p.A.,
Monza, Italy). Laser excitation was set at 488 nm and emission was
at 550-nm for FITC. As an auto-fluorescence control, a sample
treated with label solution but without TdT was carried out for
each set of analyses. FCM histograms were analysed by the Windows
Multiple Document Interface (WinMDI ver. 2.8, The Scripps Research
Institute, La Jolla, CA, USA) dedicated software.
BSAO purification
BSAO was purified from bovine blood as previously
described (25). The purified
enzyme moved as a single band on SDS/PAGE and all samples employed
had a minimum specific benzylamine oxidase activity of 0.35 IU/mg,
with IU defined as μmoles of substrate oxidized per min, assayed
spectrophotometrically at 25°C by monitoring the formation of
benzaldehyde at 250 nm absorbance (ɛ = 12,500
M−1/cm−1). The protein concentration was
measured spectroscopically and from the 280 nm absorbance, assuming
an absorption coefficient of 1.74 l
g−1/cm−1.
LoVo WT and LoVo DX survival
experiments
Cell survival experiments were carried out using
confluent cells that had been incubated for 24 h at 37°C with fresh
culture medium. Cells were harvested with EDTA in phosphate buffer
saline (PBS) and then by addition of trypsin solution in PBS,
washed by centrifugation and resuspended in PBS supplemented with
1% bovine serum albumin (BSA) (Sigma). Freshly harvested LoVo WT
and LoVo DX cells (105/ml) were incubated at 37°C for
varying time intervals in the presence of the following reagents,
used alone or in combination: BSAO (1.03×10−4 μmoles/ml
corresponding to 6.98×10−3 U/ml), spermine (0–6 μM),
catalase (240 U/ml) from bovine liver (Sigma), ALDH (EC 1.2.1.5)
from yeast (0.4 U/ml) and nicotine adenin dinucleotide
(NAD+) (1.8 μg/ml; Boehringer-Mannheim, Mannheim,
Germany). Spermine (Fluka, Buchs, Switzerland) was freshly prepared
before each experiment and, if used, added last. Cells were then
centrifuged, washed in PBS-BSA and finally resuspended in 1 ml
PBS-BSA. The cells were then plated in tissue culture-coated Petri
dishes (60×15 mm) and incubated at 37°C. Cytotoxicity was evaluated
using a colony survival assay, thus determining the ability of
cells to reproduce and form macroscopic colonies (>50 cells).
After three weeks, colonies were fixed with 96% ethanol, stained
with methylene blue and counted manually. Percentage cell survival
was determined as the ratio between the mean number of colonies in
treated and control samples.
cDNA synthesis and real-time PCR
Total RNA was extracted from the hippocampus and
frontal cortex with TRizol reagent (Invitrogen) and subjected to
DNaseI treatment (Promega) according to the manufacturer’s
instructions. Two micrograms of total RNA were then used for cDNA
synthesis, using SuperScript II (BRL Life Technologies) and random
hexamer primers according to the manufacturer’s instructions. One
microliter of cDNA was used for amplification using the following
primers: mGlu2 receptor: Bcl-2 forward, 5′-CTA
CAGTGATGTCTCCATCC-3′, reverse, 5′-AAAGCCTCAATG CCTGTCTC-3′; mGlu3
receptor: forward, 5′-CAAGTGAC TACAGAGTGCAG-3′, reverse,
5′-CTGTCACCAATGCTCAG CTC-3′; β-actin: BAX forward,
5′-TGAACCCTAAGGCCAA CCGTG-3′ reverse, 5′-GCTCATAGCTCTTCTCCAGGG-3′.
Real-time quantitative PCR was performed using a 2X Supermix
mixture (Bio-Rad) containing the double-stranded DNA Binding
fluorescent probe SYBR Green and all necessary components except
primers. Quantitative PCR conditions included an initial
denaturation step of 94°C/10 min, followed by 40 cycles of 94°C/15
sec and 55°C/15 sec. Standards, samples and negative controls (no
template) were analyzed in triplicate. Concentrations of mRNA were
calculated from serially diluted standard curves simultaneously
amplified with the unknown samples and corrected for β-actin mRNA
levels.
Statistical analysis
Significant difference at p<0.05 was evaluated by
the one-way ANOVA, followed by the multiple comparison Tukey
post-hoc test (SPSS-11 statistical dedicated software - SPSS Inc.,
Chicago, IL, USA). Whisker box-plot graphs were obtained by open
source software Gnumeric (Linux environment). All experiments were
repeated three times unless otherwise indicated.
Results
mSMOX radiation and ROS in a
neuroblastoma cell line
In previous studies, mSMOX activity was described to
induce chronic sub-lethal DNA damage, with a 3-fold increase in
oxo8dG residues, but no increase in cell mortality. Upon
2- and 4-Gy doses of X-irradiation, SMOX transfected cells were
sensitized and more prone to die than mock transfected cells.
Treatments with increasing doses of MDL abolished such
radiosensitive predisposition (15,16).
However, the level of X-ray delivered has to be considered as high
doses, thus belonging to the LNT (linear no-threshold) theory of a
linear dose-response. In the present study, we delivered a low dose
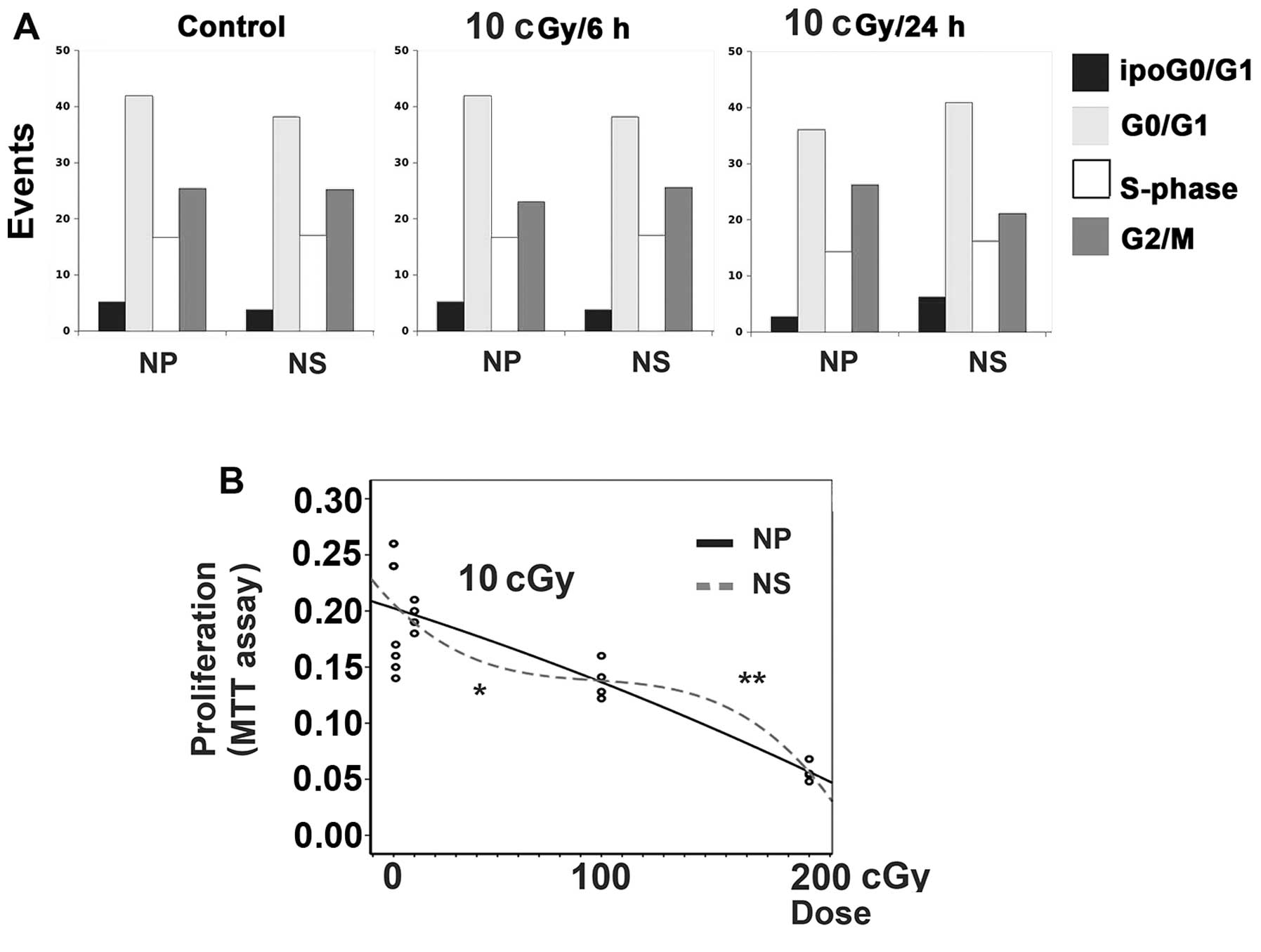
of X-rays (10 cGy) (Fig. 1A,
bar-graphs) with the relative cell cycle compartment composition,
as determined by FCM. To confirm previous data, mSMOX alone does
not apparently alter cell cycle and/or cell mortality, even in the
presence of low dose irradiation at 6 and 24 h after exposure.
Interestingly, when proliferation rates were detected by the more
sensitive MTT assays at 6 h, mSMOX associated with a hypersensitive
dose-effect curve at low dose and with an adaptive response at
higher dose (Fig. 1B).
mSMOX and radiation in AA8 parental, BER
deficient EM9 and NER deficient UV61 cell lines
To dissect the influence of chronic sub-lethal DNA
damage and DNA repair mechanisms, mSMOX was ectopically
overexpressed in proficient Chinese hamster AA8 cell line and both
deficient base-excision-repair EM9 cell line and deficient
transcription-coupled nucleotide-excision-repair UV61 cell line to
represent cellular models of priming damage dose of ROS. In
Fig. 2A, proliferation MTT assays
are represented as dose-effect curves related to low dose
irradiation at 1 and 10 cGy, both determined at 6 and 24 h after
exposure. In the AA8 and BER deficient EM9 cell lines, a
hypersensitive reaction at 1-cGy exposure is detected at 6 h,
independently from SMOX overexpression. At 24 h, the mock
transfected AA8 cells followed a linear dose-effect curve.
Contrarily, mSMOX transfected cells were resistant to both 1 and 10
cGy doses, showing a clear resistance to low dose radiation. In
this case, mSMOX elicits an adaptive response rendering cells more
reactive against DNA damage. In the EM9 cells, mSMOX does not alter
the dose-effect curves of cellular response to irradiation at 24 h,
being both more resistant than AA8 cells. However, the mSMOX
overexpression provokes an additive damage with the BER deficiency
upon low dose irradiation. In the UV61 cells, the 6 h
hypersensitivity is registered only for mock transfected cells. At
24 h, cells keep a consistent proliferation rate independently of
mSMOX and irradiation doses. The overexpression of mSMOX seems to
deliver an earlier adaptive response at 6 h, although the
proliferation-rates of UV61 cell line is much lower than AA8 and
EM9 cell lines. According to the proliferation rates, we determined
the course of apoptosis by means of TUNEL reaction. In Fig. 2B, representative FCM histograms are
shown for the unirradiated and 24 h after 10-cGy exposure. In the
AA8 cell line, the adaptive response delivered by mSMOX is clearly
evidenced by the different reduced amounts of apoptosis at 24 h. In
the EM9 cells, SMOX provokes more damage and, consistently, TUNEL
reaction is more evident for mSMOX transfected cells. In the UV61
cells, TUNEL was barely detectable with no influence by mSMOX,
probably also due to the low proliferation rates.
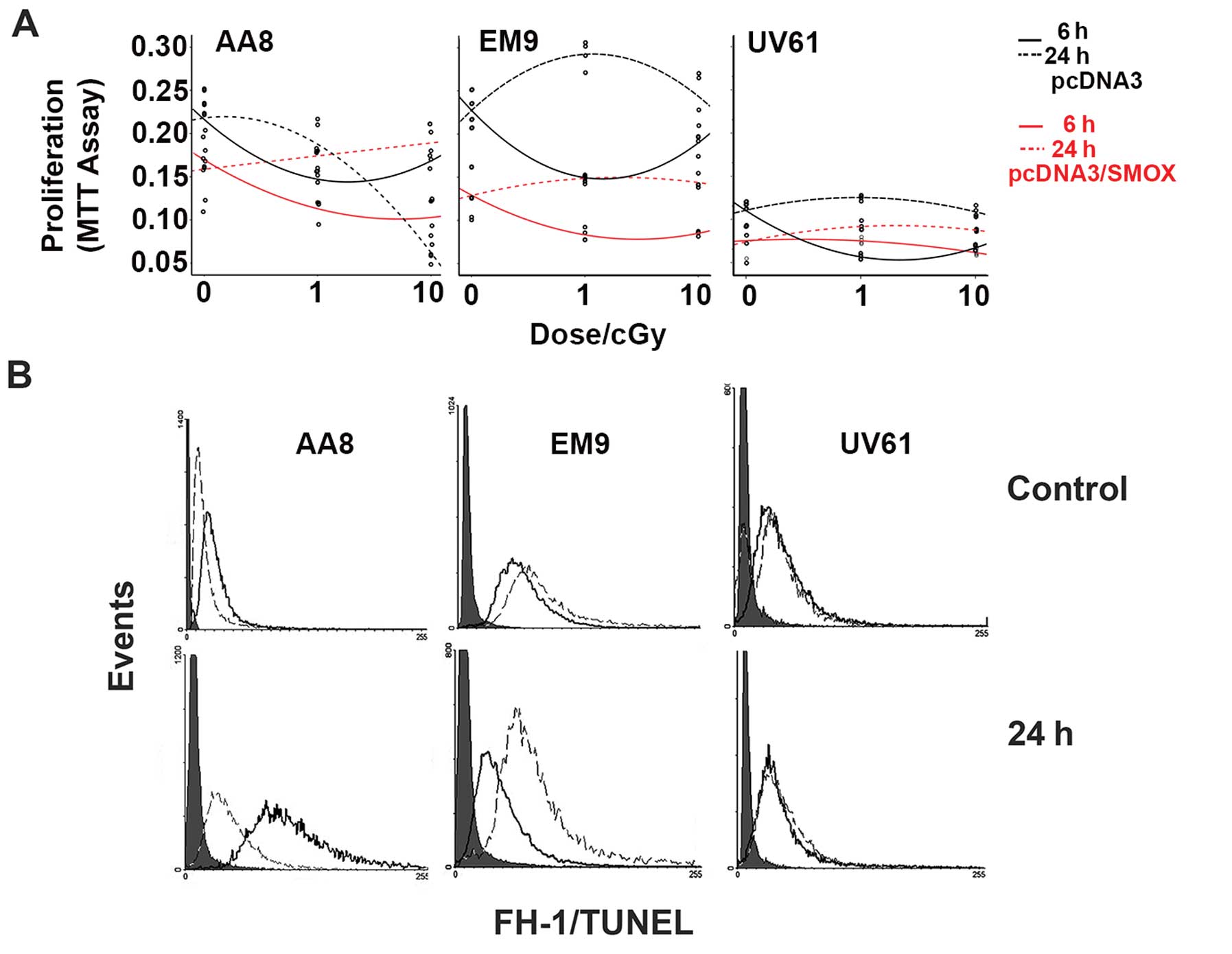 | Figure 2SMOX overexpression in AA8, EM9, UV61
cell lines and low dose IR exposure. (A) MTT proliferation assay
performed at 6 and 24 h after 1- and 10-cGy exposure (three
replicas) in AA8, EM9, UV61 cell lines, as indicated. Black line,
cell line transfected with pcDNA3 plasmid at 6 h; black dotted
line, cell line transfected with pcDNA3 plasmid at 24 h; gray line,
corresponding cell line transfected with pcDNA3/mSMOX plasmid at 6
h; dotted gray line, corresponding cell line transfected with
pcDNA3 plasmid at 24 h. (B) Representative FCM histograms of NP
(black) and NS (dashed) cell lines to analyse TUNEL reaction, at 0
and 10 cGy exposure at 24 h. Filled gray histograms represent
autofluoresence. |
mSMOX and ROS in AA8 parental, BER
deficient EM9 and NER deficient UV61 cell lines
The interconversion metabolism of SPM by SMOX
produces hydrogen-peroxide and substantial evidence has addressed
this fundamental aspect of SMOX activity to cause intracellular ROS
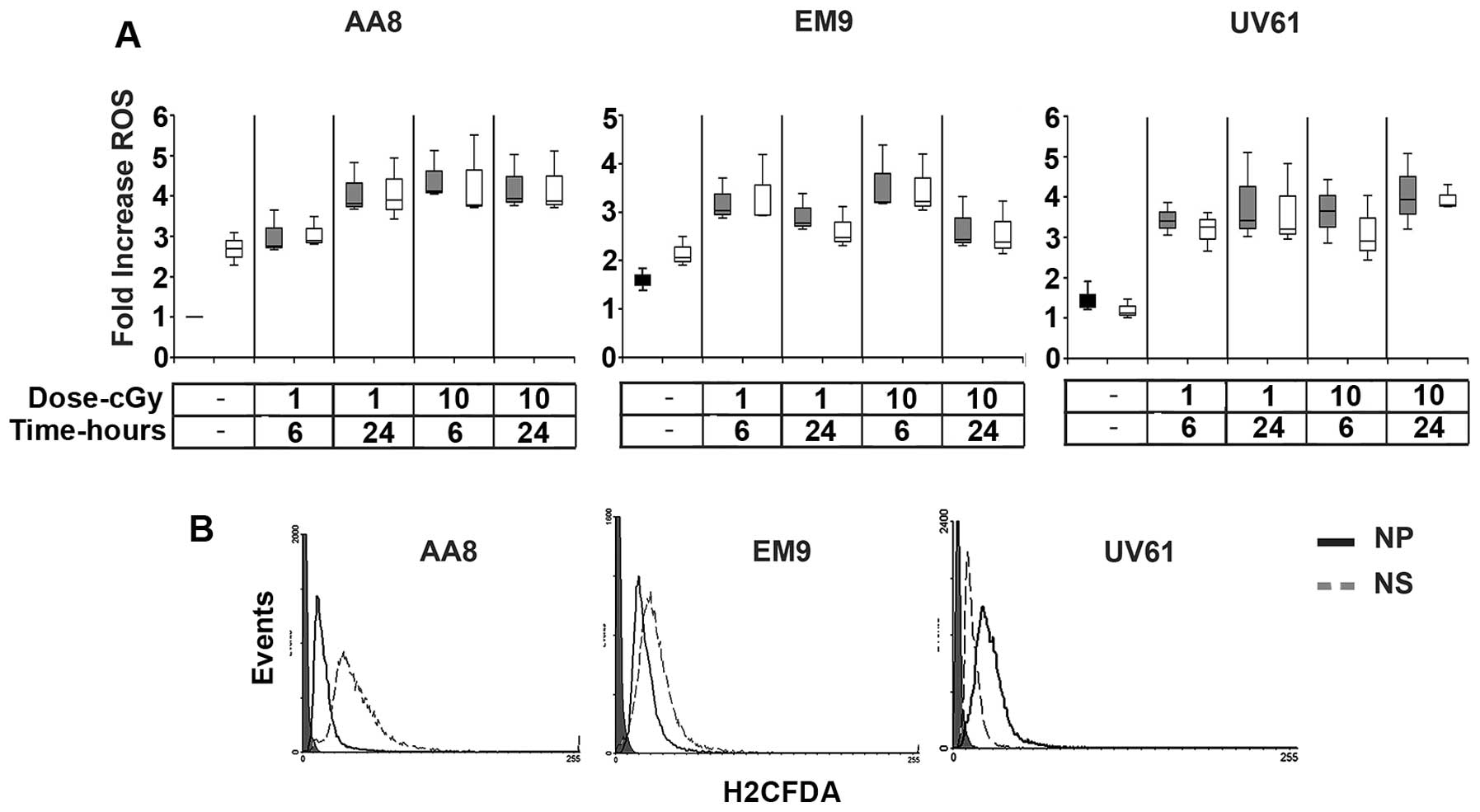
(14–16). In Fig.
3A the fold increase of ROS induced by the mSMOX overexpression
is represented as box-plot graph for all cell lines (representative
histograms are shown in Fig. 3B).
The augmented levels of ROS were significantly higher in AA8 and
EM9 cell lines when transfected with mSMOX. The ROS level in the
UV61 cell line is not influenced by mSMOX. More likely, UV61 are
very sensitive to ROS overproduction, being deficient in NER repair
system and any subtle ROS variation is not compatible with cell
life. Radiation provokes an almost three times ROS increase in all
cell lines tested, reaching a hypothetical threshold, which may
represent a sort of life-death barrier.
LoVo WT and LoVo DX cell viability
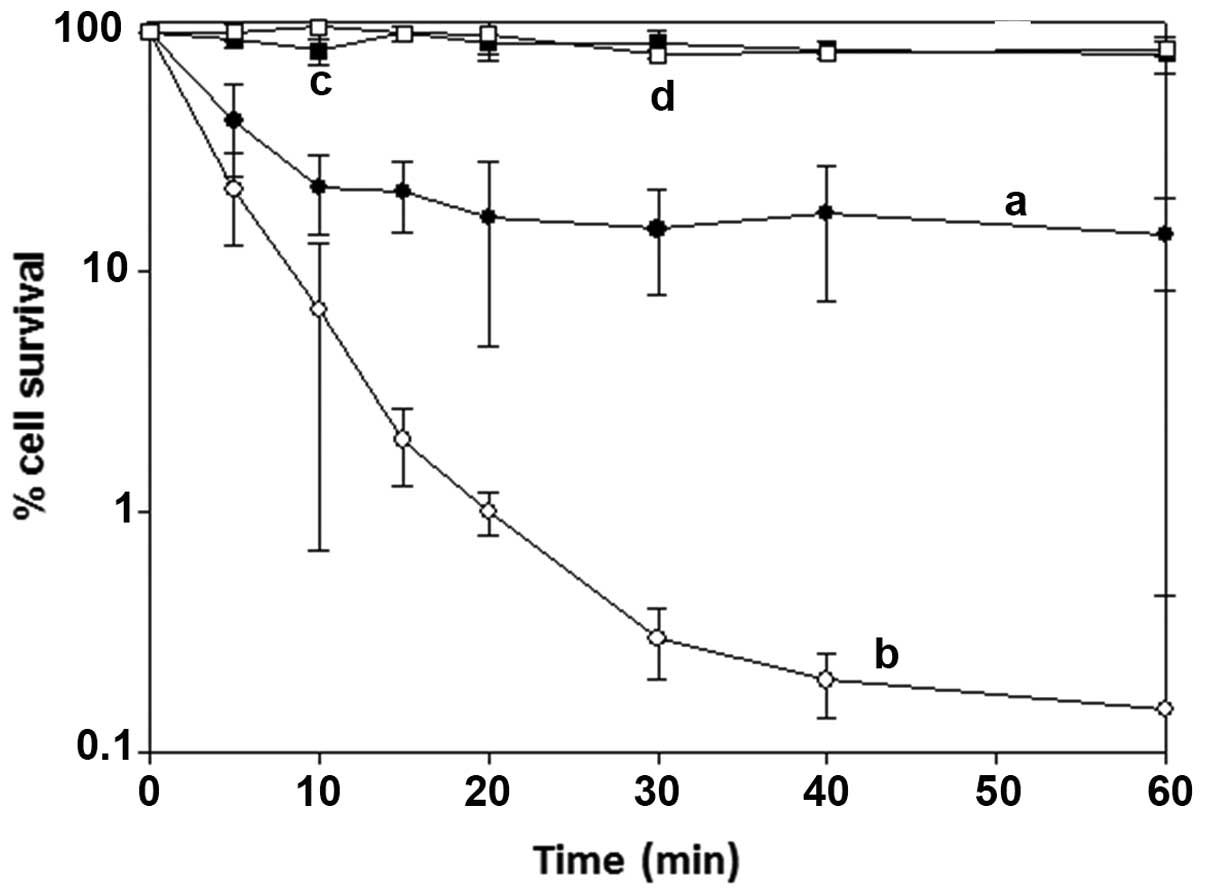
Fig. 4 shows the
percentage of cell survival versus the time of exposure to purified
BSAO (6.98×10−3 U/ml) in the presence of exogenous
spermine (12 μM), with and without catalase, at 37°C. In the
presence of BSAO and spermine alone higher cytotoxicity was
observed in LoVo DX than in LoVo WT cells. The percent cell
survival decreased in both cell lines with increasing exposure
time, resulting in ~18% in LoVo WT (Fig. 4, curve a) and ~0.3% in LoVo DX
cells (Fig. 4, curve b), after 60
min of incubation. In order to evaluate the contribution of each
enzymatic oxidation product in the inhibition of cell growth, the
experiments were performed in the presence of exogenous catalase,
an enzyme which decomposes H2O2, or catalase
and aldehyde dehydrogenase (ALDH) added simultaneously to the
incubation mixture (data not shown). Catalase (240 U/ml) afforded a
marked reduction of the cytotoxic effect, corresponding to ~80%
cell survival, on LoVo WT and LoVo DX cells (Fig. 4, curves c and d, respectively),
probably due to the clearance of hydrogen peroxide, formed in the
catalytic reaction by the enzyme. The result showed that
H2O2 was not the sole toxic factor and that
other products of the enzymatic oxidative deamination were
involved, such as aldehyde(s), including acrolein spontaneously
formed from the aminoaldehydes (27), an aspect still debated. The
addition of exogenous NAD-dependent ALDH (0.4 U/ml) metabolized the
aldehyde form to the corresponding carboxilic acid and prevented
the toxic effects of the aldehyde(s) or acrolein. In fact, after
addition of both exogenous enzymes, catalase and NAD-dependent
ALDH, cytotoxicity was completely inhibited throughout the 60 min
of incubation (data not shown).
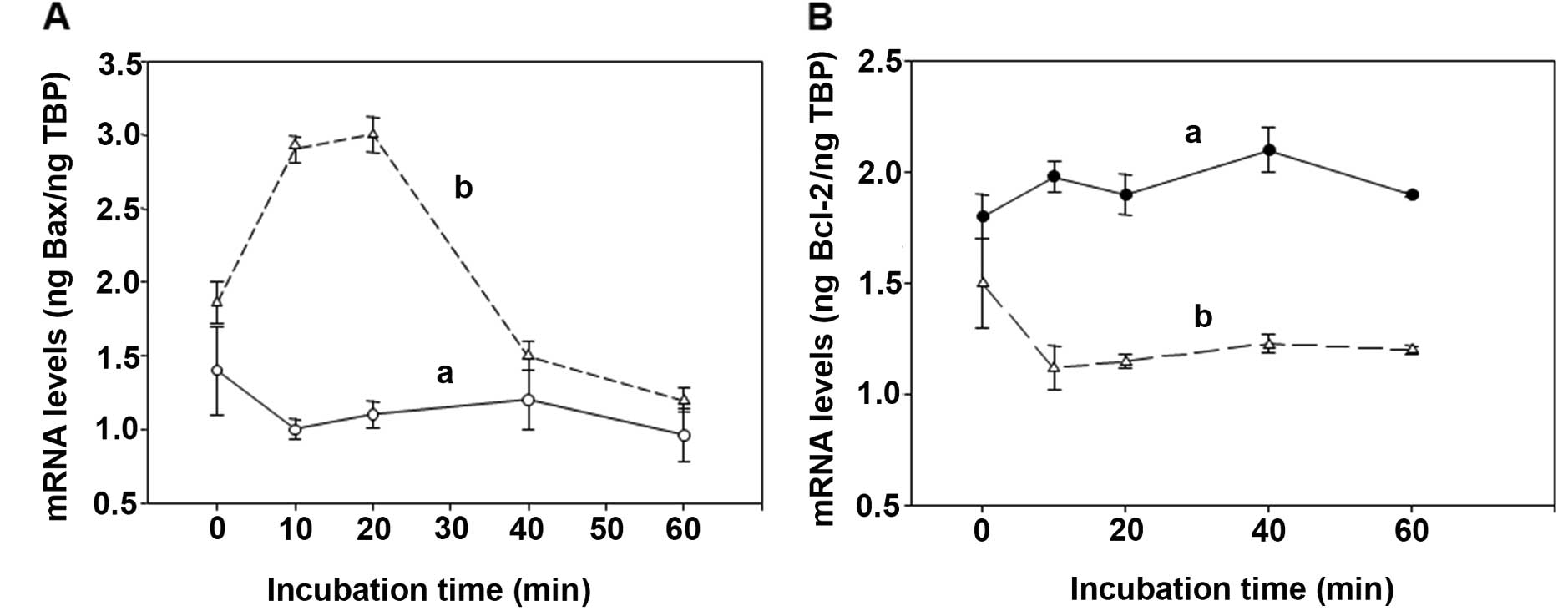
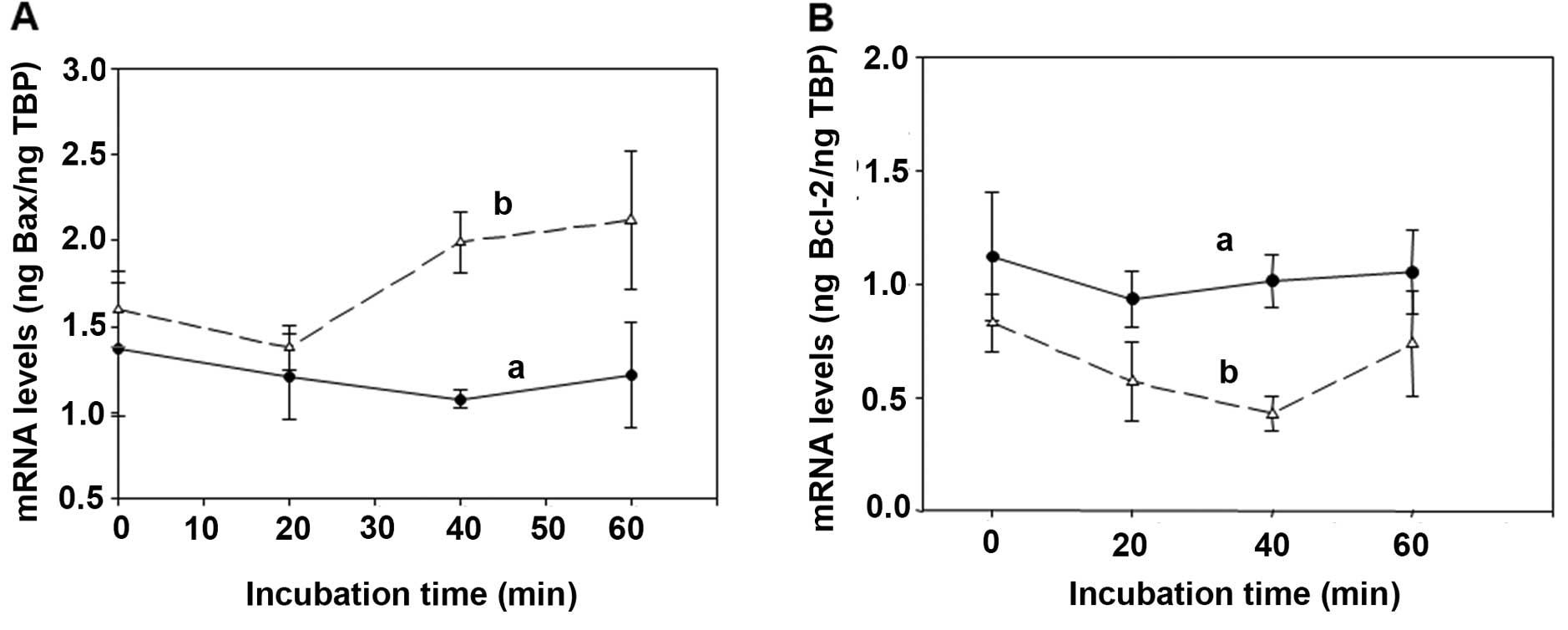
Determination of the overexpression of
the pro-apoptotic BAX and of the pro-survival Bcl-2 genes induced
by the spermine enzymatic oxidation products
Cytotoxicity induced on human cancer cells by bovine
serum amine oxidase (BSAO) and spermine is mainly attributed to
H2O2 generated by the enzymatic reaction.
Results obtained on colon adenocarcinoma LoVo WT and LoVo DX cells
by real-time PCR experiments, showed an increase in mRNA levels for
the BAX pro-apoptotic gene in MDR cells after treatment of both
LoVo cell lines with 12 μM concentration of exogenous
H2O2 (Fig.
5A). Whereas, LoVo cells treated with 12 μM concentration of
exogenous acrolein, showed only a slight overexpression of BAX at
the last stage of the reaction in MDR cells (Fig. 6A). In these experimental
conditions, the pro-survival Bcl-2 gene did not reveal any
variation after LoVo WT and MDR cell treatment with either
exogenous H2O2 (Fig. 5B) or acrolein (Fig. 6B). Therefore, the enhancement in
mRNA levels for the BAX pro-apoptotic gene due to
H2O2 and acrolein was more marked in LoVo MDR
cells than in LoVo WT ones, in agreement with the clonogenic assay
obtained by the treatment of LoVo cells with BSAO in presence of 12
μM spermine concentration, as previously demonstrated by the
authors.
Discussion
In this study, we used the chronic sub-lethal damage
driven by ectopical overexpression of mSMOX as priming dose of
cellular stress, followed by two challenging low doses of
X-irradiation, to evaluate the mSMOX capability to elicit an
adaptive or additive response in cellular models. Moreover, we
studied the influence of the in situ formation of cytotoxic
ROS formed by BSAO and spermine to differentiate the MDR response
in contrast with wild-type human tumor cell lines. Ionising
radiation induces clustered DNA damage, defined as two or more
lesions within one or two helical turns of the DNA double-helix by
a single radiation track (28,29).
The primary cellular repair mechanism is the base excision repair
(BER) pathway to repair base lesions, AP sites and SSBs which are
induced in cells either endogenously or by IR (30,31).
In our models, the overexpression of mSMOX as priming dose of
cellular stress is mainly ROS and DNA suffers oxidative damage from
free radicals produced in living cells. This damage, if not
correctly repaired, can lead to genomic instability and increased
risk of developing cancer. To dissect if chronic sub-lethal DNA
damage driven by SMOX overexpression could deliver adaptive or
additive response, we force SMOX expression in different DNA repair
deficient cellular models, such as EM9 (BER-deficient) and UV61
(NER-deficient) cell lines. The challenging doses of X-irradiation
were in the low dose range to evidence the influence of SMOX
activity, mainly related to proliferation and survival. In fact,
doses <10 cGy do not deliver any mutation and genomic
instability (32), thus are out of
the scope of this study. In NB cell line, SMOX delivered an effect
not in agreement with the LNT theory, causing hypersensitivity at
lower dose and adaptive response at the higher dose of 10 cGy. In
the AA8 and BER deficient EM9 cell lines, mSMOX elicits an adaptive
response rendering cells less sensitive to DNA damage. Moreover, at
the lower dose of 1 cGy, SMOX is additive to BER deficiency, in
agreement with that expected, since BER is the principal mechanism
of DNA repair both for IR and ROS mediated DNA damage. In the UV61
cells, the overexpression of mSMOX seems to deliver an earlier
adaptive response at 6 h. NER deficient cell line seems take
advantage to the priming damage of SMOX to rescue IR damage, as NER
is not primarily involved in this kind of DNA repair.
Controversially, tumor cells acquire resistance to grow in an
oxidative environment, developing also a MDR mechanism to overwhelm
chemotherapy. However, in recent years, we observed that
cytotoxicity induced by ROS, downstream the spermine metabolites by
BSAO, was greater in human tumor MDR LoVo cells than the wild-type
counterpart, Agostinelli et al(18,33).
In addition, our findings also showed that ROS formed by the
combination BSAO/spermine are not only able to prevent tumor cell
growth, but also prevents mass tumor growth. MDR mitochondrial
damage observed by mitochondrial membrane depolarization and
transmission electron microscopy (TEM) was attributed to the
cytotoxic effects induced by ROS, generated during the treatment,
aldehyde(s) also contributed to cytotoxicity, but at a later stage
of the reaction and to a lesser extent (~20%), as demonstrated in
the presence of aldehyde dehydrogenase (12,13,34).
In the present study, we further confirmed these results by
real-time PCR experiments. Increasing mRNA levels for the BAX
pro-apoptotic gene mainly was observed due to
H2O2 and was again more evident in LoVo MDR
cells than in their WT counterparts. In conclusion, SMOX could
deliver a therapeutic gain when forced in NB, parental and cancer
cells with impaired BER repair mechanism at low, fractionated dose
of IR. Contrarily, in cells with deficiency in NER repair
mechanisms, SMOX could play an adaptive role to overwhelm DNA
damage by IR and be deleterious for therapy. In treatment with
chemotherapy alone, cytotoxic PA metabolites might be important as
a new approach in anti-neoplastic therapy in combating cancer,
particularly against MDR cancer cells and this represents an aspect
of particular importance with regard to the potential therapeutic
applications of ROS, since conventional cancer therapy suffers from
the development of drug resistance (35).
Acknowledgements
This study was funded in part by the Italian MIUR
(Ministero dell’Istruzione, dell’Università e della Ricerca), by
Istituto Superiore di Sanità ‘Project Italy-USA’, by Istituto
Pasteur Fondazione Cenci Bolognetti. The authors are indebted with
Professor F. Palitti (Univ. Tuscia, Viterbo, Italy) for supplying
NER and BER deficient cell lines.
Abbreviations:
|
ALDH
|
aldehyde dehydrogenase
|
|
BER
|
base-excision-repair
|
|
BSA
|
bovine serum albumin
|
|
BSAO
|
bovine serum amine oxidase
|
|
DOX
|
doxorubicin
|
|
DX
|
doxorubicin-resistant
|
|
FBS
|
fetal bovine serum
|
|
FCM
|
flow cytometry
|
|
HEPES-BSS
|
HEPES-buffered balanced salt
solution
|
|
H2O2
|
hydrogen peroxide
|
|
H2DCFDA
|
2′,7′-dichlorodihydroflurescein
|
|
IU
|
international units
|
|
LNT
|
linear no-threshold
|
|
MDR
|
multidrug-resistant
|
|
mSMOX
|
mouse spermine oxidase
|
|
NAD+
|
nicotine adenine dinucleotide
|
|
NER
|
nucleotide-excision-repair
|
|
PA
|
polyamine
|
|
PBS
|
phosphate-buffered saline
|
|
P-gp
|
P-glycoprotein
|
|
ROS
|
reactive oxygen species
|
|
SD
|
standard deviation
|
|
SDS/PAGE
|
sodium dodecyl sulphate/polyacrylamide
gel electrophoresis
|
|
SPM
|
spermine
|
|
TEM
|
transmission electron microscopy
|
|
TUNEL
|
3′-terminal deoxy-transferase
|
|
WT
|
wild-type
|
References
|
1
|
Ames BN, Shigenaga MK and Hagen TM:
Oxidants, antioxidants and the degenerative diseases of aging. Proc
Natl Acad Sci USA. 90:7915–7922. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chance B, Sies H and Boveris A:
Hydroperoxide metabolism in mammalian organs. Physiol Rev.
59:527–605. 1979.PubMed/NCBI
|
|
3
|
Schumacker PT: Reactive oxygen species in
cancer cells: live by the sword, die by the sword. Cancer Cell.
10:175–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ghavami S, Eshragi M, Ande SR, et al:
S100A8/A9 induces autophagy and apoptosis via ROS-mediated
cross-talk between mitochondria and lysosomes that involves BNIP3.
Cell Res. 20:314–331. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wong CH, Iskandar KB, Yadav SK, Hirpara
JL, Loh T and Pervaiz S: Simultaneous induction of non-canonical
autophagy and apoptosis in cancer cells by ROS-dependent ERK and
JNK activation. PLoS One. 5:e99962010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kamata H, Honda S, Maeda S, et al:
Reactive oxygen species promote TNFalpha-induced death and
sustained JNK activation by inhibiting MAP kinase phosphatases.
Cell. 120:649–661. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wallace HM, Fraser AV and Hughes A: A
perspective of polyamine metabolism. Biochem J. 376:1–14. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Casero RA Jr and Marton LJ: Targeting
polyamine metabolism and function in cancer and other
hyperproliferative diseases. Nat Rev Drug Discov. 6:373–390. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ploszaj T, Motyl T, Zimowska W, Skierski J
and Zwierzchowski L: Inhibition of ornithine decarboxylase by
α-difluoromethylornithine induces apoptosis in HC11 mouse mammary
epithelial cells. Amino Acids. 19:483–496. 2000.
|
|
10
|
Seiler N and Raul F: Polyamines and
apoptosis. J Cell Mol Med. 9:623–642. 2005. View Article : Google Scholar
|
|
11
|
Amendola R, Cervelli M, Fratini E,
Polticelli F, Sallustio DE and Mariottini P: Spermine metabolism
and anticancer therapy. Cur Cancer Drug Targets. 9:118–130. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Calcabrini A, Arancia G, Marra M, et al:
Enzymatic oxidation products of spermine induce greater cytotoxic
effects on human multidrug-resistant colon carcinoma cells (LoVo)
than on their wild-type counterparts. Int J Cancer. 99:43–52. 2002.
View Article : Google Scholar
|
|
13
|
Arancia G, Calcabrini A, Marra M, et al:
Mitochondrial alterations induced by serum amine oxidase and
spermine on human multidrug resistant tumor cells. Amino Acids.
26:273–282. 2004. View Article : Google Scholar
|
|
14
|
Pledgie A, Huang Y, Hacker A, et al:
Spermine oxidase SMO(PAOh1), Not N1-acetylpolyamine oxidase PAO, is
the primary source of cytotoxic H2O2 in
polyamine analog-treated human breast cancer cell lines. J Biol
Chem. 280:39843–39851. 2005.PubMed/NCBI
|
|
15
|
Bianchi M, Bellini A, Cervelli M, et al:
Chronic sub-lethal oxidative stress by spermine oxidase over
activity induces continuous DNA repair and hypersensitivity to
radiation exposure. Biochim Biophys Acta. 1773:774–783. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Amendola R, Bellini A, Cervelli M, et al:
Direct oxidative DNA damage, apoptosis and radio sensitivity by
spermine oxidase activities in mouse neuroblastoma cells. Biochim
Biophys Acta Rev Cancer. 1775:15–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Agostinelli E, Arancia G, Dalla Vedova L,
et al: The biological functions of polyamine oxidation products by
amine oxidises: perspectives of clinical applications. Amino Acids.
27:347–358. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Agostinelli E, Dalla Vedova L, Belli F,
Condello M, Arancia G and Seiler N: Sensitization of human colon
adenocarcinoma cells (LoVo) to reactive oxygen species by
lysosomotropic compounds. Int J Oncol. 29:947–955. 2006.PubMed/NCBI
|
|
19
|
Agostinelli E, Condello M, Molinari A,
Tempera G, Viceconte N and Arancia G: Cytotoxicity of spermine
oxidation products to multidrug resistant melanoma cells (M14
ADR2): sensitisation by MDL 72527, a lysosomotropic compound. Int J
Oncol. 35:485–498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Soares FA, Shaugnessy SG, MacLarkey WR and
Orr FW: Quantification and morphologic demonstration of reactive
oxygen species produced by Walker 256 tumor cells in vitro and
during metastasis in vivo. Lab Invest. 71:480–489. 1994.PubMed/NCBI
|
|
21
|
Thompson LH, Brookman KW, Dillehay LE, et
al: A CHO-cell strain having hypersensitivity to mutagens, a defect
in DNA strand-break repair and an extraordinary baseline frequency
of sister-chromatid exchange. Mutat Res. 95:427–440. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thompson LH, Salazar EP, Brookman KW, et
al: Recent progress with the DNA repair mutants of Chinese hamster
ovary cells. J Cell Sci. (Suppl 6): 97–110. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pelicano H, Carney D and Huang P: ROS
stress in cancer cells and therapeutic implication. Drug Resist
Updat. 7:97–110. 2004. View Article : Google Scholar
|
|
24
|
Grandi M, Geroni C and Giuliani FC:
Isolation and characterization of a human colon adenocarcinoma cell
line resistant to doxorubicin. Br J Cancer. 54:515–518. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Turini P, Sabatini S, Befani O, et al:
Purification of serum amine oxidase. Anal Biochem. 125:294–298.
1982. View Article : Google Scholar
|
|
26
|
Bahn S, Mimmack M, Ryan M, et al: Neuronal
target genes of the neuron-restrictive silencer factor in
neurospheres derived from fetuses with Down’s syndrome: a gene
expression study. Lancet. 359:310–315. 2002.PubMed/NCBI
|
|
27
|
Sharmin S, Sakata K, Kashiwagi K, et al:
Polyamine cytotoxicity in the presence of bovine serum amine
oxidase. Biochem Biophys Res Commun. 282:228–235. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Regulus P, Duroux B, Bayle PA, et al:
Oxidation of the sugar moiety of DNA by ionizing radiation or
bleomycin could induce the formation of a cluster DNA lesion. Proc
Natl Acad Sci USA. 104:14032–14037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eccles LJ, Lomax ME and O’Neill P:
Hierarchy of lesion processing governs the repair, double-strand
break formation and mutability of three-lesion clustered DNA
damage. Nucleic Acids Res. 38:1123–1134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hitomi K, Iwai S and Tainer JA: The
intricate structural chemistry of base excision repair machinery:
implications for DNA damage recognition, removal and repair. DNA
Repair. 6:410–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zharkov DO: Base excision DNA repair. Cell
Mol Life Sci. 65:1544–1565. 2008. View Article : Google Scholar
|
|
32
|
Maxwell CA, Fleisch MC, Sylvain V, et al:
Targeted and nontargeted effects of ionizing radiation that impact
genomic instability. Cancer Res. 68:8304–8831. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Agostinelli E, Belli F, Molinari A, et al:
Toxicity of enzymatic oxidation products of spermine to human
melanoma cells (M14): sensitization by heat and MDL 72527. Biochim
Biophys Acta. 1763:1040–1050. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Agostinelli E and Seiler N: Lysosomotropic
compounds and spermine enzymatic oxidation products in cancer
therapy (Review). Int J Oncol. 31:473–484. 2007.PubMed/NCBI
|
|
35
|
Agostinelli E and Seiler N:
Non-irradiation-derived reactive oxygen species (ROS) and cancer.
Therapeutic implications. Amino Acids. 31:341–355. 2006. View Article : Google Scholar : PubMed/NCBI
|