Introduction
Malignant melanoma and treatment failure are,
unluckily, two intertwined clinical problems with a profound social
impact. The lifetime risk of melanoma continues to increase
steadily. Unfortunately, melanoma lesions can remain unnoticeable
or asymptomatic for extended periods of time. Melanoma can generate
metastasis at stage IV, without a clearly identifiable primary
lesion.
The success of systemic therapy for melanoma has
been minimal, because melanoma is an extremely aggressive disease
with high metastatic potential and a high resistance to cytotoxic
agents. Indeed, it is notoriously high resistant to standard drugs,
such as nitrosoureas, vinca alkaloids, platinum-associated drugs
and anthracyclines antibiotic, like doxorubicin. A body of
literature data has described several mechanisms and factors
implicated in the resistance of malignant melanoma. In particular,
the multidrug resistant (MDR) phenotype can be costitutively
expressed by melanoma cells. It is mediated by the overexpression
of ATP-binding cassette (ABC) transporters such as ABCA9,
ABCB1/P-gp, ABCB5, ABCB8, ABCC1, ABCC2 and ABCD1, that efflux
anticancer drugs from cells and reduce their concentration and
cytotoxic effect (1,2). Moreover, constitutively activated
Ras/Raf/MEK/ERK (MAPK) and the PI3K/AKT (AKT) signaling pathways
strongly contribute to confer resistance to the apoptotic death
(3).
A broad spectrum of targeted therapeutic strategies,
including targeting of the anti-apoptotic proteins, signal
transduction pathways and tumour angiogenesis factors, are
currently being studied in clinical trials (4). Unfortunately, in conventional cancer
chemotherapy, all the therapies contribute scarsely to overall
patient survival. Numerous difficulties prevent successful
treatment, such as the poor selectivity of the cytotoxic drugs for
tumours and the development of MDR of cancer cells, which represent
the most difficult problems to solve. Thus, alternative therapeutic
strategies are required. In the development of new treatments
against tumours the differences between normal and cancer cells
must be considered.
One such difference is the polyamine content and
metabolism (5,6). The content of polyamines, such as
putrescine, spermidine and spermine (Spm), is increased in tumour
cells in comparison with their normal counterparts. Increased
polyamine levels are associated with increased cell proliferation,
decreased apoptosis and down-expression of genes affecting tumour
invasion and metastasis (7).
Moreover, the natural polyamines are also source of cytotoxic
metabolites, because they are substrates of a large class of
enzymes, named amine oxidases. Our previous studies demonstrated
that these enzymes catalyze the oxidative deamination of polyamines
to generate cytotoxic products, HO and aldehydes, that induce
apoptotic and non-apoptotic cell death. Such products also proved
to induce a higher cell death on MDR melanoma cells than on the
corresponding wild-type cells (8,9). It
was demonstrated that the combination of BSAO/Spm with either
docetaxel or interferon α had a synergistic effect on the
inhibition of cell growth through apoptosis in both human
epidermoid KB and breast cancer MCF-7 cell lines (10,11).
Noteworthy, our previous studies demonstrated that
the induction of cell death, in colon adenocarcinoma (LoVo) and
melanoma (M14) cancer cells, was potentiated by the combined
treatments of BSAO/Spm with MDL 72527
[N1,N4-bis(2,3-butadienyl)-1,4-butanediamine
dihydrochloride], an inactivator of flavin-adenin-dinucleotide
(FAD)-dependent amine oxidase (AO) with lysosomotropic properties
(8,9,12,13).
Thus, the aim of this study was to verify if also CQ, a well known
lysosomotropic compound, was able to sensitize tumour cells to the
cytotoxic activity of spermine metabolites (14). Chloroquine, 4-aminoquinoline, is a
drug widely used for the therapy of malaria and as an
anti-inflammatory agent for the treatment of several diseases such
as rheumatoid arthritis, lupus erythematosus and amoebic hepatitis,
by targeting the polyamine pathway.
A systematic exploration of the potential of this
category of compounds in cancer therapy seems of interest. In fact,
CQ has been studied for its potential as an enhancing agent in
cancer therapies. Therefore, in the present study, the effect of
pre-treatment with CQ on the cytotoxicity of the spermine
metabolites was evaluated on wild-type human melanoma cells (M14
WT) and the corresponding doxorubicin-induced MDR cells (M14 ADR2).
The type of the induced cell death, the mechanism of action and the
main intracellular targets were investigated. Moreover, the
possible cytotoxicity of spermine metabolites on melanocytes were
also studied, in order to exclude undesired side effects in view of
future clinical applications.
Materials and methods
Chemicals
If not stated otherwise chemicals were from Sigma
Chemical Co. (St. Louis, USA). Spermine tetrahydrochloride was from
FLUKA (Buchs, Switzerland), doxorubicin (DOX) was from Pharmacia
and Upjohn (Milan, Italy). Chloroquine (CQ) diphosphate salt was
from Sigma product of United Kingdom.
Purification of BSAO
BSAO was purified to homogeneity essentially as
described by Turini et al (15) with additional ion exchange
chromatographic steps as described previously by Janes et al
(16). The purified enzyme moved
as a single band on sodium dodecyl sulphate (SDS)/PAGE and all
samples employed had a minimum specific benzylamine oxidase
activity of 0.38 IU/mg, with international unit (IU) defined as
micromoles of substrate oxidized per min, assayed
spectrophotometrically at 25°C by monitoring the formation of
benzaldehyde at 250 nm absorbance (ɛ=12,500/M/cm). The protein
concentration was determined spectrophotometrically at 280 nm,
assuming an absorption coefficient of 1.74 l/g/cm.
Melanoma cells and culture
conditions
The parental human M14 cell line, isolated from an
epidermal melanoma, and the corresponding MDR variant M14 ADR2 were
used in the present investigation. The resistant cell line M14 ADR2
was obtained by us culturing the M14 ADR cell line, previously
selected by Molinari et al (17), in medium containing 10 μM
doxorubicin (DOX) constantly, in each passage. Both resistant lines
display multidrug resistant phenotype being, in addition to DOX,
also resistant to other chemotherapeutic agents. Cells were grown
in monolayer in Falcon flasks (initial cell number
8×105) in RPMI-1640 (with glutamine), 10% foetal bovine
serum (FBS) (Hyclone, Europe Ltd., Cramlington, UK), penicillin (50
U/ml), streptomycin (50 μg/ml) and non-essential amino acids in the
same concentration as streptomycin, in a humidified atmosphere of
5% CO in a waterjacketed incubator at 37°C.
Melanocyte culture
Normal human epidermal melanocytes (NHEM) were
cultured in PromoCell Melanocyte Growth Medium M2 (is a serum-free)
and Supplement Mix C-39420 (PromoCell, Heidelberg, Germany).
Briefly, cryopreserved primary melanocytes were transferred into
tissue culture Falcon flasks at a density of 5×105
cells/25 cm2 and were grown in melanocyte growth medium
in a humidified atmosphere of 5% CO, 95% air in a waterjacketed
incubator at 37°C. The culture medium was changed every other day.
After cells became semiconfluent the medium was removed and
melanocytes were subcultured. The Detach kit consists of three
components: i) HEPES-BSS (HEPES-buffered balanced salt solution);
ii) 0.04% Trypsin/0.03% EDTA; 3) 0.05% Trypsin inhibitor, 0.1% BSA.
All solutions were stored at −20°C in the dark immediately after
arrival.
Cell viability: clonogenic assay
Cell viability assays were carried out using
subconfluent cells. Cells were detached by addition of 10 mM EDTA
in phosphate-buffered saline (PBS), washed with PBS supplemented
with 1% bovine serum albumin (BSA) and pelleted by centrifugation
(2 min, 1500 × g). The cells were re-suspended in PBS/BSA. Aliquots
of freshly harvested M14 cells (105/ml) were incubated
at 37°C for different time intervals in the presence of the
following reagents, used alone or in association: BSAO (17.20 μg/ml
corresponding to 1.01×10−4 μmoles/ml or
6.5×10−3 IU/ml), spermine (up to 6 μM). The spermine
solution was freshly prepared before each experiment and, if
present, added last. Other cell survival experiments were performed
using subconfluent cells that had been incubated for 24 h in
RPMI-1640 medium containing CQ at 50 μM, in the presence of serum,
at 37°C. Incubation was carried out as described above, in the
presence or absence of BSAO/spermine. After incubation the cells
were centrifuged, washed twice in PBS/BSA and re-suspended in 1 ml
PBS/BSA. Cytotoxicity was evaluated by a plating efficiency assay,
which determines the ability of the cells to form macroscopic
colonies (>50 cells). Aliquots of cell suspensions containing
105 cells/ml were plated in tissue culture dishes (50 mm
Ø) containing 5 ml complete culture medium and were incubated at
37°C. After 14 days, the colonies were fixed with 96% ethanol,
stained with methylene blue and counted. Control plating
efficiencies were >85 and 80%, for M14 WT and M14 ADR2 cells,
corresponding to 9.1×104±1.0×104 and
8.5×104±1.0×104 number of cells,
respectively. The percentage of colony forming cells was determined
as the ratio between the mean number of colonies in treated and
control samples.
Cell viability: MTT assay
M14 cells and melanocytes were seeded at
1×104 density in 96-well plates and pre-treated with
various concentrations of CQ (0–50 μM) for 24 h and then with
BSAO/spermine for 1 h, at 37°C. Cells were exposed to 10 μl of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide dye (5
mg/ml) and were incubated at 37°C for 4 h for the conversion of MTT
to MTT-formazan crystals by mitochondrial enzymes. In order to
solubilize the formazan crystals, 100 μl DMSO was added. Absorbance
was determined at 570 nm with the spectrophotometer mod.
Perkin-Elmer λ 11 (Bio-Rad, Hercules, CA, USA).
Scanning electron microscopy (SEM)
For SEM studies, control and treated cells were
grown on coverslips and fixed with 2.5% glutaraldehyde in 0.1 M
cacodylate buffer (pH 7.3), added with 2% sucrose, at room
temperature for 20 min. After post-fixation with 1% OsO in 0.1 M
cacodylate buffer (pH 7.3) at room temperature for 30 min, cells
were dehydrated through graded ethanol concentrations, critical
point-dried in CO (CPD 030 Balzers device, Bal-Tec, Balzers) and
gold coated by sputtering (SCD040 Balzers device, Bal-Tec). The
samples were then examined with a Cambridge Stereoscan 360 scanning
electron microscope (Cambridge Instruments, Cambridge, UK).
Transmission electron microscopy
(TEM)
Transmission electron microscopy observations were
performed using confluent cells that had been incubated for 24 h in
RPMI-1640 medium containing CQ at 50 μM, in the presence of serum,
at 37°C. Then, cells were harvested as described above, washed with
PBS/1% BSA, centrifuged and re-suspended in 2 ml of RPMI-1640
medium (without FBS). After incubation for 60 min at 37°C in the
presence or absence of BSAO (6.5×10−3 IU/ml) and
spermine (6 μM), cells were washed with RPMI-1640 medium and then
processed for TEM, as described previously by Molinari et al
(18). Ultrathin sections were
examined with a Philips EM 208S electron microscope (FEI Co.,
Eindhoven, The Netherlands).
Morphological analysis of apoptosis by
Hoechst 33258 labeling
To analyze the apoptotic morphology by fluorescence
microscopy, both control and BSAO/Spm-treated melanocytes and
melanoma cells, grown on coverslips, were fixed with 3.7%
paraformaldehyde in PBS for 10 min at room temperature. Then,
samples were labeled with Hoechst 33258 at the concentration of 0.1
μg/ml for 15 min to counterstain DNA and observed with a CCD camera
equipped light microscope Olympus BX51 (Olympus, Tokyo, Japan).
Flow cytometry analyses
All flow cytometric analyses were carried out on
cell suspensions (~106 cells/ml) obtained by incubating
monolayer cell cultures with EDTA. The fluorescent signals were
analyzed by a BDLSRII flow cytometer (Becton-Dickinson, Mountain
View, CA) equipped with a 5 mW, 488 nm, air-cooled argon ion laser
and a Kimmon HeCd 325 nm laser. The fluorescence emissions were
collected through a 530-nm band pass filter for fluorescein
isothiocyanate conjugated antibody (FITC), a 575-nm band pass
filter for propidium iodide (PI). At least 10,000 events/sample
were acquired in linear for cell cycle studies or log mode for
Annexin V-FITC labeling. Percentages of cells in subG1, G1, S and
G2/M phases of cell cycle and percentages of early and late
apoptotic cells were calculated using the FACS Diva Software
(Becton-Dickinson).
Determination of apoptotic cell death by
Annexin V-FITC staining
To detect phosphatidylserine translocated from the
inner face to the outer surface of plasma membrane in the initial
step of apoptosis, an Annexin V-FITC apoptosis detection kit (MBL,
Medical and Biological Laboratories Co., Ltd., Japan) was used as
described by Van Engeland et al (19). About 50 phosphatidylserine monomers
are estimated to be bound per each Annexin V molecule. M14 WT and
M14 ADR2 cells (1×106/ml) were treated with 50 μM CQ for
24 h or with BSAO (6.5×10−3 IU/ml) and spermine (6 μM)
for 1 h at 37°C or with the combination CQ and BSAO/spermine. Cell
suspensions were then plated in tissue culture-coated Petri dishes
containing culture medium supplemented with FBS. After incubation
at 37°C for 48 and 72 h, cells were detached, centrifuged and
resuspended in binding buffer (10 mM HEPES/NaOH, pH 7.5, 140 mM
NaCl and 2.5 mM CaCl). Cell suspension (~5×105) was then
incubated with 1 μg/ml of Annexin V-FITC and with 1 μg/ml of PI for
10 min at room temperature in the dark. The populations of Annexin
V-FITC negative/PI negative cells (viable cells), Annexin V-FITC
positive/PI negative cells (early apoptosis) and Annexin V-FITC
positive/PI positive cells (late apoptosis) were evaluated by flow
cytometry (19). The dot plots
reported in Fig. 2 were obtained
from one out of three independent experiments which gave very
comparable results.
Cell cycle analysis
Cell cycle distribution was analyzed by labelling
cells with PI. Assays were carried out as described by Nicoletti
et al (20). M14 WT and
ADR2 cells (1×106/ml), obtained from different
treatments, as described in Annexin V-FITC labeling, were
collected, washed twice with cold PBS and then centrifuged. The
pellet was fixed in 70% ethanol in PBS at 4°C for 1 h, washed twice
and then re-suspended in PBS containing 100 μg/ml Ribonuclease A
(RNAse) and 40 μg/ml PI. After incubation at 37°C for 30 min, cells
were analyzed by flow cytometry.
Confocal microscopy observations of
acidic vacuoles
To analyze by laser scanning confocal microscopy
(LSCM) the formation of acidic vacuoles in M14 cells, induced by
treatment with 50 μM CQ, the vital staining with acridine orange
solution was performed. Cells were grown on the coverslips until
confluent state and then treated with the lysosomotropic compound
for 24 h. The cells were incubated with acridine orange solution
(2.5 μg/ml) for 15 min at 37°C; after 20 min in drug-free medium at
37°C, then washed with PBS and immediately observed. The
observations of acidic vacuoles were carried out using a Leica TCS
SP2 spectral confocal microscope (Leica Microsystems, Wetzlar,
Germany) equipped with Argon-Helium-Neon lasers. In these
organelles, the stacked form of acridine orange generates a bright
red fluorescence; on the contrary in the nucleus and cytoplasm,
unstacked acridine orange displays variable green fluorescence.
Statistical analysis
Values reported in the figures are means ± standard
deviation (SD) from 3–6 independent experiments. Statistical
analyses were performed using Student’s t-test, with p<0.05
considered statistically significant.
Results
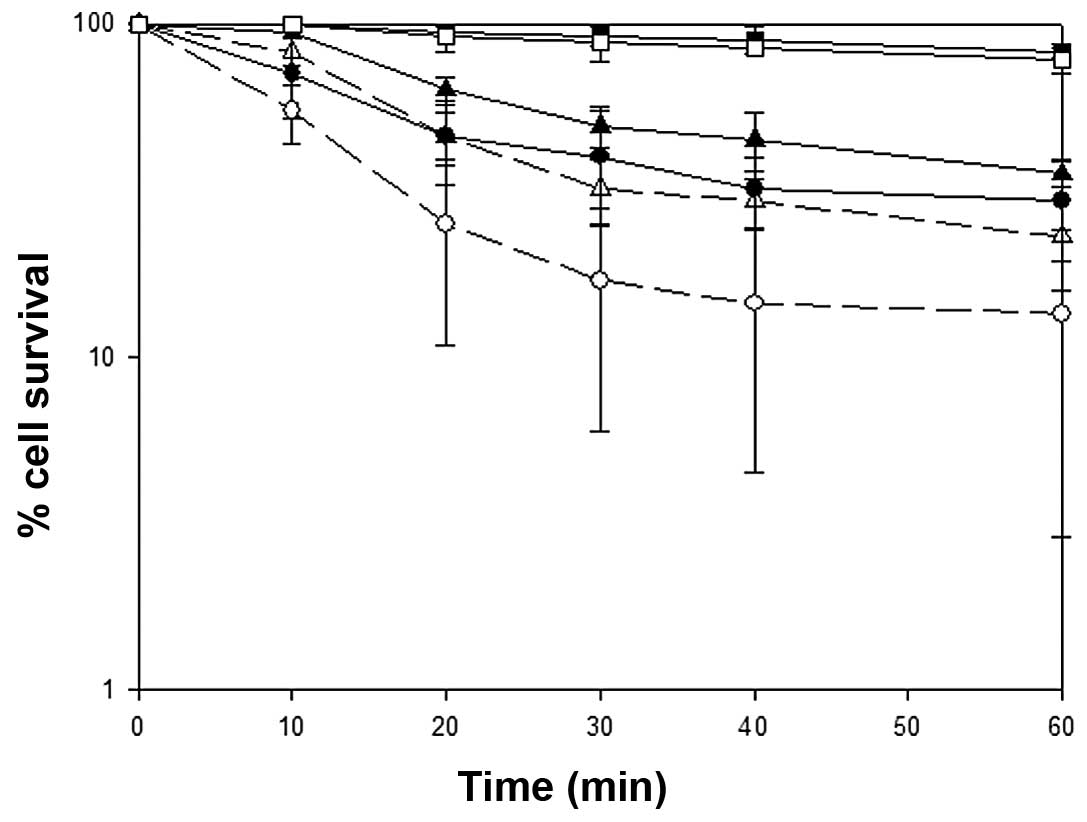
Enhancement of the cytotoxic effect of
polyamine metabolites by pre-incubation with chloroquine
Cloning efficiency test demonstrated that the
lysosomotropic compound chloroquine (CQ) was able to increase the
cytotoxic activity of BSAO/Spm enzymatic system on both drug
sensitive and drug resistant melanoma cells (Fig. 1). Since treatment with 50 μM CQ
alone for 24 h did not impair the survival of both cell lines, this
subcytotoxic concentration was chosen to sensitize cells to
enzymatic products of BSAO/Spm.
When M14 WT cells were pre-incubated with 50 μM CQ
for 24 h and then exposed for up to 60 min to BSAO
(6.5×10−3 IU/ml) and Spm (6 μM) at 37°C, cell survival
percentage significantly decreased, when compared with that of
cultures treated with the enzymatic system BSAO/Spm alone. The
chemosensitizing effect exerted by CQ was much more evident on the
MDR melanoma cell line, whose cell survival decreased to about 10%
after 60 min of BSAO/Spm treatment (Fig. 1).
Induction of apoptotic cell death by
treatment with the combination CQ+BSAO/Spm
The induction of apoptosis was analyzed by flow
cytometry at 48 and 72 h after the end of the treatment (Fig. 2) on control and treated cells
labelled with Annexin V-FITC and PI. According to clonogenic data,
the treatment with CQ alone did not change the fraction of early
and late apoptotic cells in comparison with the untreated cells.
The treatment with BSAO/Spm alone induced a slight increase of
early (8.0 and 11.9%) and late (7.0 and 5.1%) apoptotic cells on
M14 WT and ADR2 cells, respectively. These percentages
significantly increased after pre-incubation with CQ (9.7 and 32.7%
for the early fraction, and 11.2 and 11.4% for the late fraction on
M14 WT and ADR2 cells, respectively). The apoptotic effect was
clearly more evident in drug resistant M14 cells, and after 72 h
from the end of the combined treatment. In fact, at this time,
pre-treatment with CQ significantly increased the percentage of
apoptotic cells (63.9%) in drug resistant melanoma cultures
compared to that determined in the sensitive ones (27.6%).
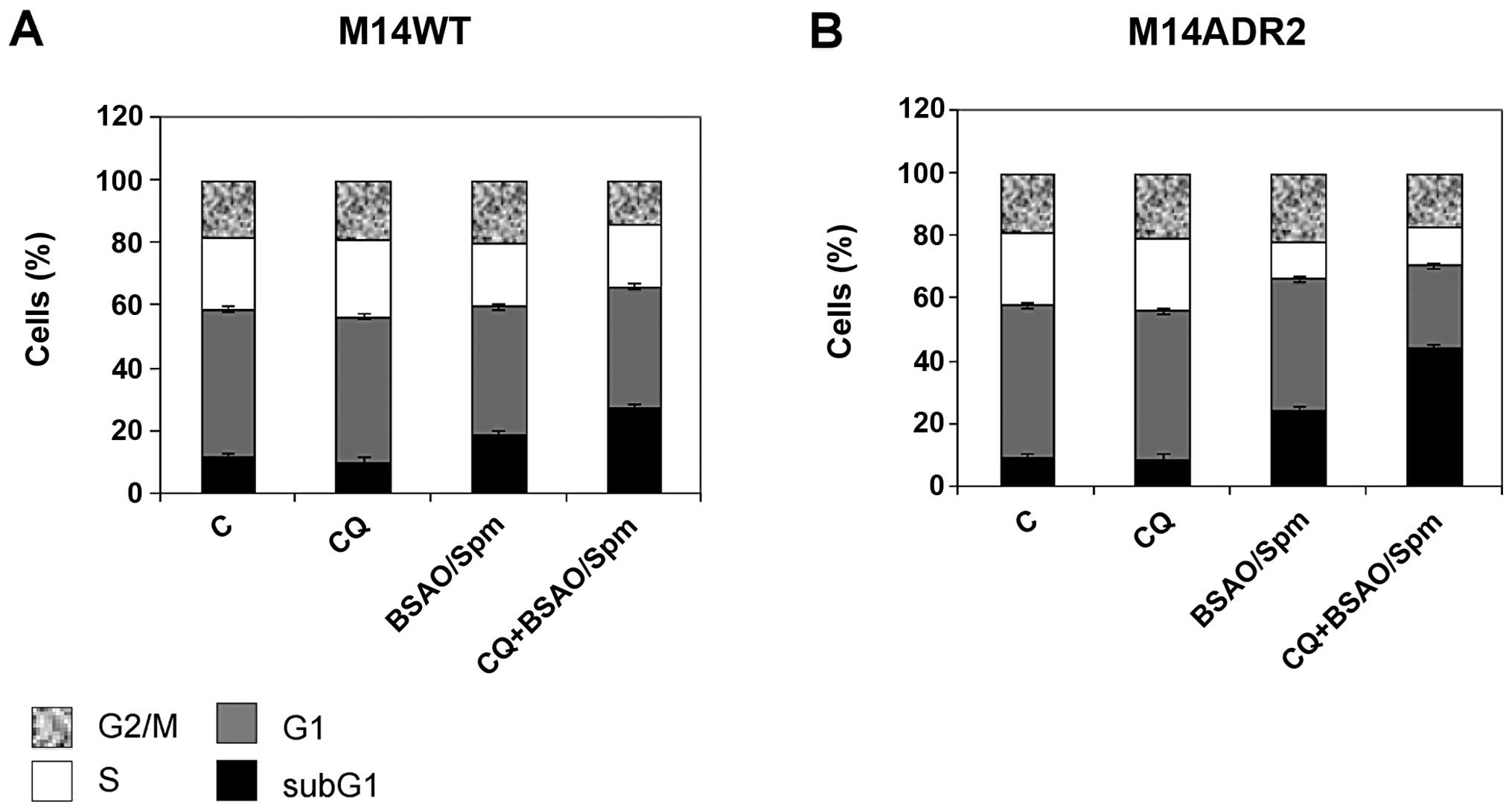
The analysis of the cell cycle (Fig. 3) confirmed the data obtained by
Annexin V-FITC/PI test. The treatment with CQ alone did not change
the percentage of subG1, G1, S and G2/M fractions on either cell
line. The treatment with BSAO/Spm induced a slight increase of
subG1 cell fraction on M14 WT cells (19.0%) and a more significant
increase on M14 ADR2 cells (24.4%). When M14 WT cells were
incubated for 24 h with CQ and then treated with BSAO/Spm a
remarkable increase of the subG1 cell fraction was revealed
(27.3%). This effect was much more evident on M14 ADR2, where the
subG1 fraction reached the value of 44.2%.
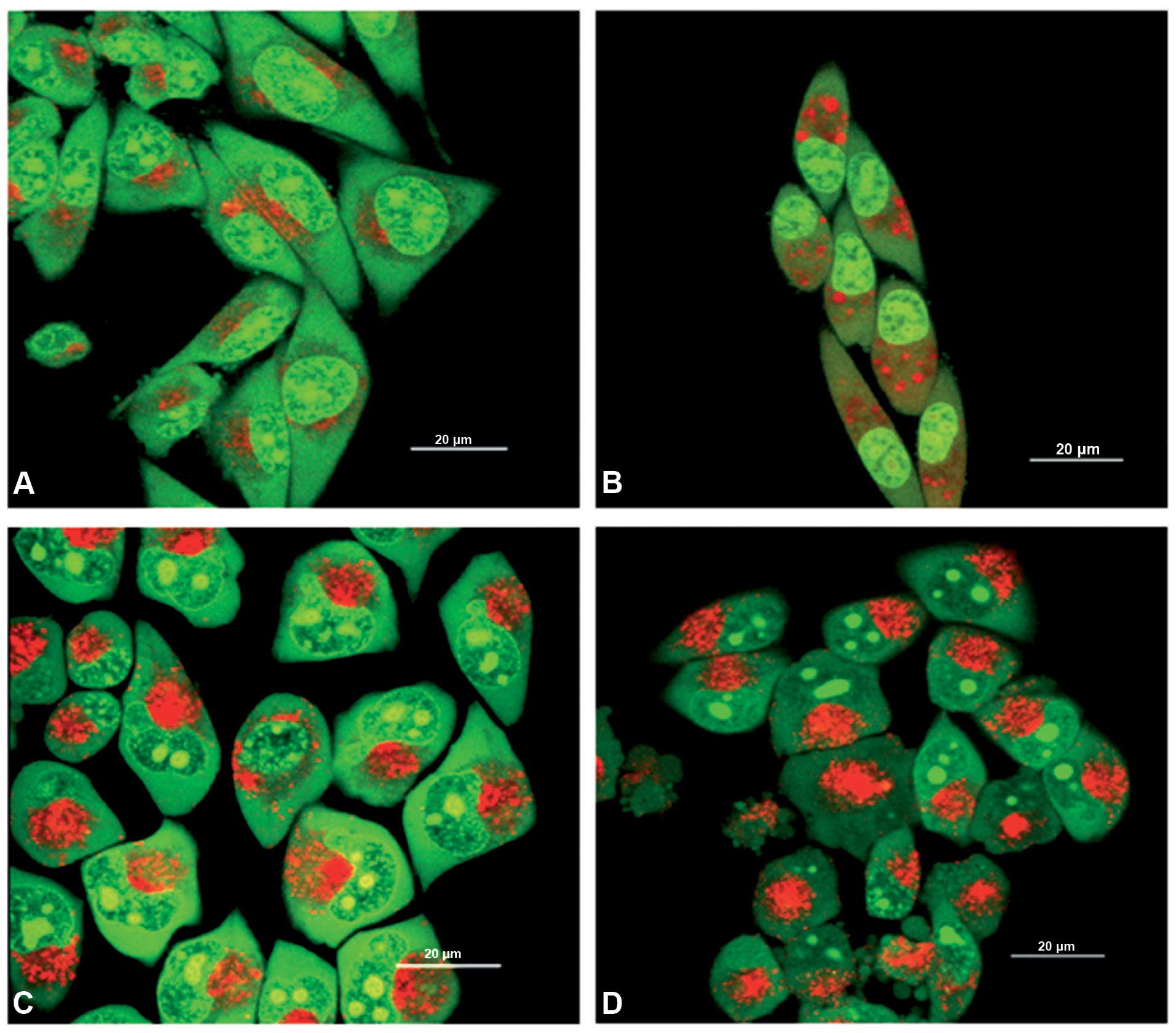
Confocal microscopy observations of the
lysosomotropic effect induced by CQ
The treatment with 50 μM CQ for 24 h induced an
increase of the acidic compartments in drug sensitive and resistant
melanoma cells, as demonstrated by the staining with the acridine
orange solution and observations by LSCM of vital cells. In control
M14 WT (Fig. 4A) and M14 ADR2
cells (Fig. 4B) the red signal
appeared localized in cytoplasmic perinuclear organelles that
noticeably increased in number after the treatment with CQ in both
sensitive and resistant cells (Fig. 4C
and D, respectively), suggesting the induction of a
lysosomotropic effect.
Ultrastructural alterations induced by
the combined treatment with CQ+BSAO/Spm
Transmission electron microscopy observations on
ultrathin sectioned melanoma cells allowed the detection of the
subcellular changes related to the potency of the cytotoxic effect
of BSAO/Spm exerted by CQ pre-treatment (Fig. 5). According to the above reported
observations, the treatment with CQ alone did not induce any
evident alteration of the subcellular structures, either on
sensitive or resistant cells (Fig. 5B
and F, respectively), which appeared to be well preserved as in
the corresponding control untreated cells (Fig. 5A and E, respectively). However,
after CQ treatment the appearance of a large number of lysosomes
and cytoplasmic vacuoles were observed. After incubation with
BSAO/Spm enzymatic system, M14 WT cells did not show significant
ultrastructural modifications (Fig.
5C), according to previous reported observations by Agostinelli
et al (9). On the contrary,
the mitochondria of M14 ADR2 cells showed rarefied matrix and
swollen cristae (Fig. 5G).
Finally, in both melanoma cell lines treated for 24 h with CQ and
then with BSAO/Spm, numerous lysosomes and cytoplasmic vacuoles
were still observable inside most cells (Fig. 5D and H, respectively). Moreover,
the mitochondria of cells treated with the combination CQ+BSAO/Spm
showed alterations much more evident than those present in cells
treated with BSAO/Spm alone. Again, M14 ADR2 cells were more
susceptible than M14 WT cells to the enzymatic oxidation products
of BSAO/Spm system.
Effects of BSAO/Spm enzymatic system,
acrolein and hydrogen peroxide on the cell survival of melanoma
cells and melanocytes
On the basis of the above reported results
demonstrating that the enzymatic oxidation products of
BSAO/Spermine system induce cytotoxic effect on cultured melanoma
cells and in order to exclude undesirable side-effects in view of
clinical applications, we then analyzed by MTT test the possible
cytotoxic effect of the enzymatic system on human melanocytes. The
results of the cytotoxicity test, besides confirming that melanoma
resistant cells (M14 ADR2) are more sensitive than their wild-type
counterparts (M14 WT) to the BSAO/Spm treatment, and showed that
melanocytes display a greater resistance than melanoma cells
against the treatment with the enzymatic system, up to 60 μM
spermine concentration for 1 h (Fig.
6 insert). In particular, after treatment with 9 μM spermine
>90% of melanocytes were viable.
Fig. 6 also shows
the comparison between melanocytes and melanoma cells regarding
their survival after treatment with BSAO/Spm alone or with the
combined treatment CQ+BSAO/Spm. The time course for 60 min clearly
shows that melanocytes, differently from melanoma cells, are
virtually unaffected by the enzymatic system, even when preceded by
CQ treatment.
Moreover, the cytotoxic effects induced by the
enzymatic system were compared to those exerted singularly by the
two oxidation products, HO and aldehyde (acrolein) at the same
molar concentration of spermine (12 μM), on melanoma cells and
melanocytes (Fig. 7). Since
acrolein is considered as the potential cytotoxic product of
spermine (21), and since it is
not known which of the aldehydes is formed preferentially from
spermine under the experimental conditions (8,12),
acrolein was used as a model compound. Exogenous acrolein
administered alone for 60 min induced low cytotoxicity on melanoma
cells and, noteworthy, it was completely ineffective against
melanocytes. Conversely, at the same concentration and incubation
time, exogenous HO caused a light effect on melanocytes and
noticeable cytotoxicity on melanoma cells.
Morphological and ultrastructural
modifications induced by enzymatic system on melanoma cells and
melanocytes
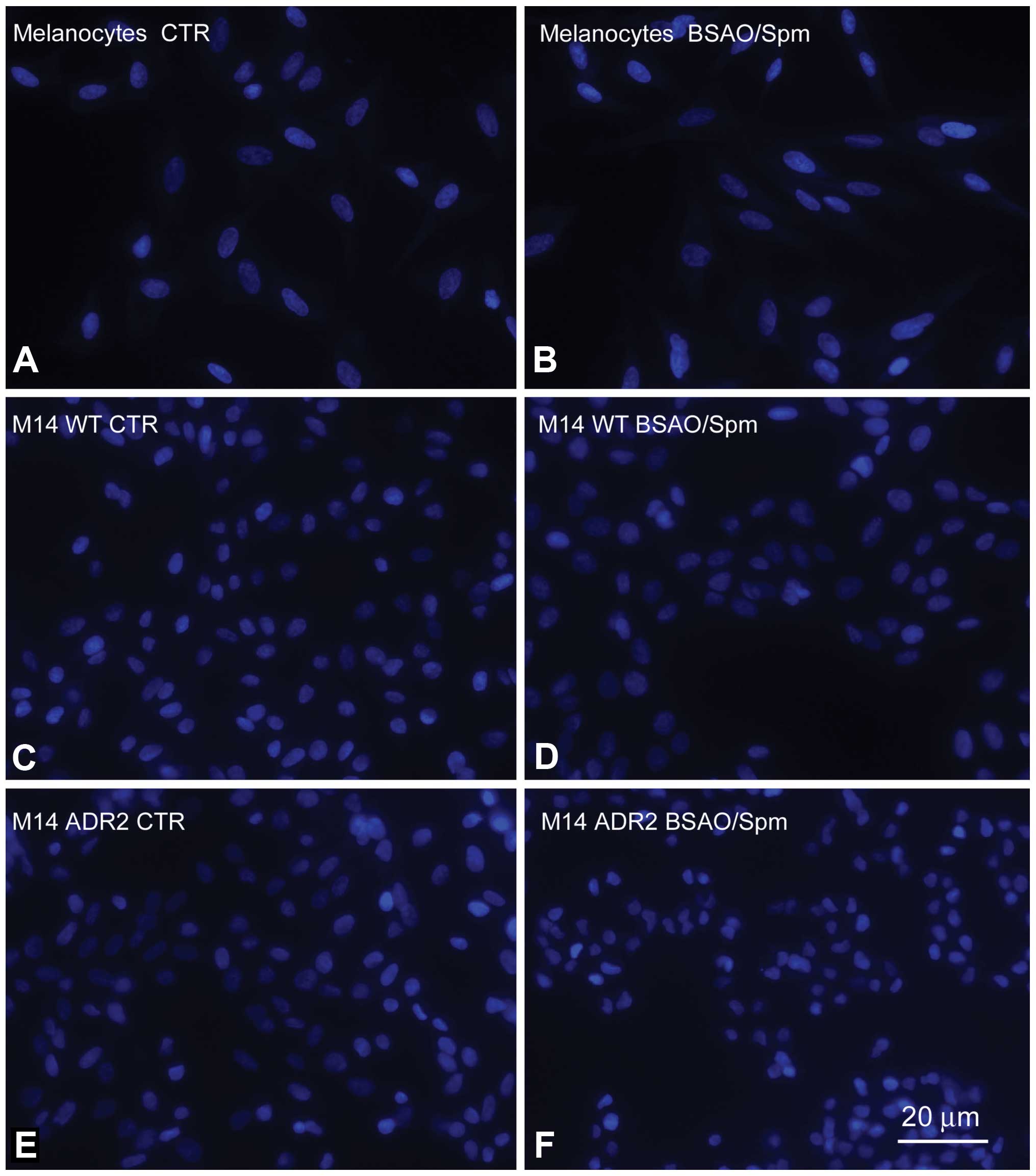
The evaluation of the apoptosis induction carried
out by fluorescence microscopy after Hoechst 33258 staining
confirmed the results obtained by the cytotoxicity test. After
treatment with BSAO/Spm the nuclei of normal melanocytes (Fig. 8B) appeared to be unaltered when
compared to those of control ones (Fig. 8A). Conversely, after the same
treatment numerous apoptotic nuclei were observed in both M14 WT
and M14 ADR2 cell cultures (Fig. 8D
and F, respectively), while they were not detectable in the
corresponding control cultures (Fig.
8C and E).
The effects of BSAO/Spm treatment on the morphology
of melanoma cells and melanocytes were also analyzed by scanning
electron microscopy (Fig. 9).
Control melanocytes (Fig. 9A), M14
WT (Fig. 9D) and M14 ADR2
(Fig. 9G) cells displayed very
similar morphology characterized by bipolar elongated shape and the
cell surface covered by short and randomly dispersed microvilli.
Accordingly to cytotoxicity data, melanocytes showed unaltered
morphological appearance either after 24 or 48 h of BSAO/Spm
treatment (Fig. 9B and C,
respectively). On the contrary, after treatment for 24 h both
sensitive (Fig. 9E) and resistant
(Fig. 9H) melanoma cells appeared
to be rounded in shape and tended to partially detach from the
substrate. Moreover, numerous thin and long cell protrusions
connecting adjacent cells were easily detectable. Such alterations
were even more evident after treatment for 48 h and more dramatic
on M14 ADR2 cells (Fig. 9I) than
in the sensitive counterparts (Fig.
9F).
Transmission electron microscopy observations of
ultrathin sectioned cells substantially confirmed the scarce
susceptibility of melanocytes to BSAO/Spm treatment (Fig. 10). In fact, melanocytes treated
with spermine (Fig. 10B) at the
concentration (6 μM) that induces cytotoxicity in melanoma cells,
displayed an unaltered ultrastructural appearance when compared to
untreated cells (Fig. 10A). In
particular, mitochondria, the main subcellular target of the
oxidation products of the enzymatic system (22), maintained the orthodox feature,
characterized by regular shape and parallel cristae. To induce
light intracristal swelling and matrix rarefaction in melanocyte
mitochondria, 10 times higher concentration of spermine (60 μM) had
to be administered (Fig.
10C).
As expected, many treated melanoma cells observed by
transmission electron microscopy showed the typical apoptotic
changes consisting in evident alterations of the cytoplasmic
organelles and chromatin condensation and marginalization. Fig. 10D shows an apoptotic melanoma cell
treated with BSAO/Spm (6 μM).
Discussion
In conventional cancer chemotherapy, numerous
obstacles exist preventing successful treatment, such as the poor
selectivity of the cytotoxic drugs for tumours and the development
of MDR of cancer cells, which represent the most difficult problems
to solve. Therefore, alternative therapeutic strategies are
required. Among these, although still at a beginning, enzymatically
formed ROS have certain potential in cancer therapy. The in
situ formation of toxic compounds or radicals by
enzyme-catalyzed reactions is a promising start (5,23,24).
In previous papers we reported on investigations on the potential
use, as antitumour agent, of an enzyme capable of forming cytotoxic
products (hydrogen peroxide and aldehydes) from polyamines that are
present at elevated levels in cancer cells. In vitro studies
were carried out on Chinese hamster ovary (CHO) cells (25), and human colon adenocarcinoma cells
(12) as well as on human melanoma
(M14) (8) and mouse melanoma
tumour cells (B16-F0) (26). It
was observed that hydrogen peroxide and aldehydes, enzymatic
oxidation products of polyamines, exerted greater cytotoxic effects
on colchicine- and doxorubicin-resistant cells (CHO-CHC5 and LoVo
DX, respectively) than on their parental sensitive counterparts
(CHO-AuxB1 and LoVo WT) (22,27).
The growth of the mouse melanoma was inhibited by exposure to BSAO
and spermine (26,28) and for the slow release of toxic
enzymatic oxidation products of spermine into the tumour, the use
of BSAO conjugated to biocompatible polymers was considered
(28,29). However, in the case of BSAO, the
administration of the substrate spermine, might be unnecessary in a
new therapeutic approach, since cancer cells have high polyamine
concentrations (5,6,30),
and the polyamines are released into the cellular environment, if
cells are damaged (23).
Therefore, the aim of the present study was to further improve our
knowledge of polyamine-derived cytotoxic products in causing cell
death.
From a therapeutic point of view, the improvement of
the efficacy of in situ formation of the cytotoxic oxidation
products of spermine is fundamental. This may be pursued by the
combined treatment with lysosomotropic compounds that might improve
cell damage caused by spermine metabolites. In this study, emphasis
was put on the ability of CQ to sensitize melanoma cells to the
cytotoxic effect induced by HO and aldehyde, formed from
BSAO/spermine enzymatic system (22).
Cell survival experiments were carried out on WT and
MDR melanoma cells (M14). As previously observed by using the
lysosomotropic compound MDL 72527 (8), shorter survival was observed in both
cell lines after pre-incubation with CQ (50 μM for 24 h) before the
subsequent exposure to spermine metabolites. The sensitization
began at low concentration (10 μM) but was more evident at 50 μM
CQ. Cytotoxicity was significantly greater on cells treated with
the combined treatment than on those treated with BSAO/spermine
alone, even though CQ alone did not reduce the number of viable
cells under the experimental conditions. However, an impairment of
cell metabolism by CQ was indicated by an increase of the number of
lysosomes. The treatment with 50 μM CQ for 24 h induced an
enhancement of the acidic compartments in both drug sensitive and
resistant melanoma cells, as demonstrated by the staining with the
acridine orange solution and observations by LSCM of vital cells,
hypothesizing the induction of a lysosomotropic effect.
Interestingly, the test of cytotoxicity showed that
melanocytes display a higher resistance than melanoma cells against
the treatment with the enzymatic system. In fact, more than 90% of
melanocytes were viable after treatment with 9 μM spermine.
Moreover, an interesting result was obtained when melanocytes were
treated with CQ alone or in combination with BSAO/spermine.
Melanocytes, differently from melanoma cells, were unaffected by
the enzymatic system, even when preceded by CQ treatment (Fig. 6). This phenomenon could be due to
the higher reactivity of the cellular defense systems in
melanocytes than in melanoma cells. It was reported that CQ induced
cell death dependent on GSH depletion in human glioblastoma cells
(31) and cell growth inhibition,
cell cycle arrest and apoptosis on breast cancer cells (32). In addition, it was suggested that
autophagy, a lysosomal degradation pathway, is necessary for
anti-immune responses. In response to chemotherapy, autophagy in
dying cancer cells enables the release of ATP, which attracts
immune cells and triggers an anti-immune response (33,34).
In addition to its anticancer effects (31,32),
CQ is a well known lysosomotropic compound (35,36),
because it accumulates in lysosomes, increasing their volume,
causing permeability and release of lysosomial enzymes. Its
lysosomotropic properties are probably responsible for several of
the biological effects of this drug. Among these, CQ can sensitize
cancer cells to the cytotoxic effects of chemotherapeutic agents,
mainly due to modulating effect on ABC transporters by inhibiting
the drug efflux mediated by ABC proteins (37). Therefore, its application as an
effective sensitizer in cancer therapies could be a promising new
therapeutic strategy (38). The
cancer-specific chemosensitizer effect of CQ may be partly
dependent on its ability to inhibit autophagy (35,36,38).
However, whereas other autophagy inhibitors, such as
3-methyladenine, inhibit autophagy in its early phase, consequently
resulting in inhibition of the formation of acidic vesicular
organelles, as autophagosomes and autolysosomes, CQ inhibits
autophagy in its late phase, when autophagosomes have been formed.
Moreover, in the cytoplasm of CQ-treated cells, accumulation of
acidic vesicular organelles has been reported by Tasdemir et
al (36). So, mechanistically,
CQ seems to exert its effects through the weak-base lysosomotropic
feature (39,40). This drug prevents lysosome
acidification, which is not specific to autophagy, but may have
other potentially beneficial antitumour effects in addition to
autophagy inhibition. For this reason, inhibition of autophagy
could be a promising new strategy to improve cancer treatment.
The ultrastructural alterations observed by
transmission electron microscopy in the present study (Fig. 6) are evidence in favour of the
lysosomotropic properties of CQ. The modifications observed after
exposure of M14 cells, both sensitive and resistant, to CQ alone or
in combination with BSAO and spermine, are similar to those
previously observed in LoVo and M14 cells after treatment with MDL
72527 (8,9,12,41).
The finding support the view that CQ acts as a lysosomotropic
compound and that the sensitization of melanoma cells to the
treatment with BSAO and spermine is mainly due to the effects
induced by CQ on the endosomal-lysosomal system. Among the
ultrastructural changes produced by CQ, the enhancement of
lysosomes and the rapid formation of cytoplasmic vacuoles were the
most striking. A major role of lysosomes in cell death is presently
not doubted (42), and it is known
that the release of lysosomal enzymes causes oxidative stress
(43,44). The amplification of oxidative
stress by the subsequent exposure of cells to hydrogen peroxide,
and the toxic effects of the highly reactive spermine-derived
aldehydes, explains the increase of cytotoxicity observed in cell
survival experiments, in both M14 WT and M14 ADR2 cells. According
to Zdolsek et al (45),
even the cytotoxicity of hydrogen peroxide is linked with the
damage of lysosomal membranes. This observation may be considered
as an additional argument in favour of a major role of lysosomes in
the cytotoxicity of spermine-derived metabolites.
Analysis by scanning electron microscopy revealed
characteristic morphological changes in M14 melanoma cells treated
with BSAO and spermine. In general, the morphological alterations
of M14 WT and M14 ADR2 cells reflected the results of the cell
survival experiments. We presume that the phenotypic changes
(transformation of elongated cells into polygonal and rounded
shapes) were the result of the impairment of the cytoskeleton due
to reaction with spermine metabolites. Such alterations were more
marked after treatment for 48 h on M14 ADR2 cells than in the WT
counterparts. On the contrary, according to cytotoxicity data,
melanocytes showed unaltered morphological appearance, even after
48 h of BSAO/Spm treatment. Severe changes of the mitochondrial
structure, such as dilatation of the cristae and disruption of
membranes were observed by transmission electron microscopy in M14
cells, mainly in multidrug-resistant phenotype. These observations
substantially confirmed the scarce susceptibility of melanocytes to
BSAO/Spm exposure. In fact, melanocytes treated with 6 μM spermine,
that induces cytotoxicity in melanoma cells, displayed an unaltered
ultrastructural aspect when compared to untreated cells. To induce
light intracristal swelling and matrix rarefaction in melanocyte
mitochondria, a concentration of at least 60 μM spermine was
necessary.
There is an ongoing debate on the question, whether
hydrogen peroxide or the spermine-derived aldehydes are the major
effectors in causing cytotoxicity. We previously demonstrated that
catalase added to the culture medium prevented the loss of about
80% of cell viability, suggesting that hydrogen peroxide is the
most important cytotoxic metabolite of spermine in our experimental
setting (8,9). If formed by the glucose oxidase
reaction, or added as such to the cell suspension, hydrogen
peroxide was cytotoxic at lower concentrations than acrolein
(26). So, comparing the cytotoxic
effect of 12 μM spermine in the presence of BSAO, with that of the
same concentrations of exogenous hydrogen peroxide and acrolein,
the enzymatically formed spermine metabolites were considerably
more toxic, on M14 WT and M14 ADR2 cells, than the individual
agents. Interestingly, while the enzymatic system induced low
cytotoxicity on melanocytes (approximately 20%) during 60 min of
incubation, exogenous 12 μM acrolein administered alone for 60 min
was completely ineffective against melanocytes. Conversely, at the
same concentration and incubation time, exogenous HO caused a light
cytotoxic effect on the same cells. These findings suggest that a
contribution of the aldehydes on the cytotoxicity induced on
melanoma cells is clear. According to Sharmin et al
(21) acrolein is the actual toxic
metabolite of spermine. However, acrolein is formed by spontaneous
β-elimination from the aldehydes generated by BSAO-catalyzed
oxidative deamination (8,12,26).
The rate of β-elimination is dependent on environmental conditions,
and it is unknown under most experimental settings. In view of our
lack of knowledge of the composition of the aldehyde mixture formed
from spermine under our experimental conditions, it seemed
justified, as reported in the Results section, to use acrolein as
model compound. However, particularly interesting and promising is
the finding, as demonstrated in this experimental work, that M14
ADR2 cells were much more sensitive to the toxic metabolites of
spermine than their wild-type counterpart.
The exposure of phosphatidylserine, a negative
charged phospholipid, on the outer surface of the cytoplasmic
membrane clearly showed the onset of the apoptotic process. Flow
cytometric analysis was performed by Annexin V-FITC and propidium
iodide (PI) assay (Fig. 2). The
measurement of Annexin V binding, executed simultaneously with the
exclusion dye PI, provided a suitable assay to detect apoptotic
cells and to discriminate between apoptosis (both early and late)
and necrosis. The cytotoxic products hydrogen peroxide and
aldehyde(s) induced a significant increase of early apoptotic
fraction on both cell lines 72 h after the end of the treatment
(11.1% in M14 WT and 23.7%) in M14 ADR2 cells when compared to 1.5
and 7.3% in the respective control cells. Pre-treatment with CQ,
followed by the treatment with spermine metabolites, enhanced the
percentage of apoptotic cells on both lines: 17.5% on M14 WT but
particularly on MDR cells, where it approximately doubled
(48.1%).
A further investigation on the cell cycle analysis
by flow cytometry (Fig. 3),
revealed that subG1 hypodiploid cells, a subpopulation of apoptotic
cells with fragmented DNA, accumulated in response to the treatment
of M14 WT and M14 ADR2 cells with BSAO and 6 μM spermine or when
both cells lines were pre-treated with CQ and then treated with
BSAO/spermine enzymatic system. The increase of the subG1 fraction,
induced by the oxidation products of spermine, was much higher in
M14 ADR2 cells (approximately 2 times) than in M14 WT ones after
the combined treatment. This finding is in agreement with the
results obtained either by Annexin V-FITC/PI assay or TEM
observations. Severe changes of the mitochondrial structure, such
as dilatation of the cristae, disruption of membranes and chromatin
condensation produced the characteristic apoptotic phenotype, were
mainly observed in multidrug-resistant cells.
Taking into consideration the findings described
above, the difference between MDR and WT melanoma cells in
sensitivity to treatment with BSAO and spermine, particularly after
sensitization with 50 μM CQ, is evident, and explained as follows.
Several human MDR tumour cells express P-glycoprotein, a
membrane-bound transporter protein, which functions as
energy-dependent pump capable of extruding drugs out of cells
(46). However,
P-glycoprotein-mediated excretion of the toxic products of spermine
oxidation appears to play no, or only a minor role, because M14
ADR2 cells were not less, but significantly more sensitive to
exposure to BSAO and spermine than M14 WT cells. The higher
sensitivity to cytotoxic spermine derivatives of colon
adenocarcinoma LoVo DX cells, as compared with WT cells, has
previously been attributed to an earlier and higher mitochondrial
membrane depolarisation, and a higher basal production of ROS that
was not related to the glutathione content, because MDR and WT LoVo
cells have the same glutathione pool (22,47).
At present no data exist to allow to draw the same conclusion on
M14 melanoma cells (our studies are in progress), although the same
physiological factors are possibly responsible for the enhanced
sensitivity in both MDR cell lines.
The evaluation of the apoptosis induction carried
out by fluorescence microscopy after Hoechst 33258 staining
supported the findings described above. Moreover, Hoechst assay
confirmed the results obtained by the cytotoxicity MTT test. As
reported in the Results, after treatment with spermine metabolites
the nuclei of normal melanocytes (Fig.
8B) appeared to be unaltered when compared to those of control
ones (Fig. 8A). Instead, after the
same treatment numerous apoptotic nuclei were observed in both M14
WT and M14 ADR2 cell cultures, not detectable in the corresponding
control ones.
In conclusion, from the enhanced sensitivity of
immortalised cells to lysosomal death pathways (48), new possibilities in cancer therapy
are expected (49). The
sensitisation of tumour cells to anticancer drugs by lysosomotropic
compounds, and particularly the sensitisation of MDR cells,
recommend scrutinizing the potential of lysosomotropic drugs in
cancer therapy. In clinical applications, the enhancement of the
efficacy of in situ formation of cytotoxic polyamine
metabolites, hydrogen peroxide and aldehyde enzymatically
generated, deserves to be taken into consideration. Sensitising
cells by a lysosomotropic compound, as for example chloroquine or
MDL 72527 (8,12), improved cell damage by hydrogen
peroxide and other spermine metabolites, activating the stress
signal transduction pathway and leading to apoptotic cell death.
Therefore, an extension of these experiments with the aim to
further improve the apoptotic effect of spermine metabolites
appears important. In our opinion, the systematic exploration of
chloroquine in combination with conventional anticancer drugs
promises new and efficient anticancer therapies within a short
period. It has been demonstrated that chloroquine has antiviral
effects (50) and, more important,
it improves the therapy of glioblastoma multiforme (51). Thus, the studies carried out in our
laboratories on human colon adenocarcinoma and melanoma cells,
suggest that the anti-malarial drug chloroquine, in association
with BSAO/spermine, could potentiate the effects of the enzymatic
oxidation products of spermine and might be important as a new
approach in anti-neoplastic therapy, particularly against MDR
cancer cells.
Acknowledgements
This study was partially supported by the Italian
MIUR (Ministero dell’Istruzione, dell’Università e della Ricerca),
by Istituto Superiore di Sanità ‘Project Italy-USA’, by funds
MIUR-PRIN (Cofin) and by Istituto Pasteur-Fondazione Cenci
Bolognetti. Thanks are also due to Fondazione ‘Enrico ed Enrica
Sovena’ for the scholarships to Giampiero Tempera for supporting
his post-doc (E.A.).
Abbreviations:
|
ADR
|
adriamycin
|
|
AO
|
amine oxidase
|
|
BSA
|
bovine serum albumin
|
|
BSAO
|
bovine serum amine oxidase
|
|
CQ
|
chloroquine
|
|
DMSO
|
dimethyl sulfoxide
|
|
DOX
|
doxorubicin
|
|
EDTA
|
ethylenediaminetetraacetic acid
|
|
FAD
|
flavin-adenin-dinucleotide
|
|
FBS
|
foetal bovine serum
|
|
FITC
|
fluoresceine isothiocyanate
conjugated
|
|
HEPES-BSS
|
HEPES-buffered balanced salt
solution
|
|
IU
|
international units
|
|
LSCM
|
laser scanning confocal microscopy
|
|
MFC
|
means fluorescence channel
|
|
MDL 72527
|
[N1,N4-bis(2,3-butadienyl)-1,4-butanediamine
dihydrochloride]
|
|
MDR
|
multidrug resistance
|
|
PBS
|
phosphate-buffered saline
|
|
P-gp
|
P-glycoprotein
|
|
PI
|
propidium iodide
|
|
RNAse A
|
ribonuclease A
|
|
ROS
|
reactive oxygen species
|
|
SEM
|
scanning electron microscopy
|
|
SD
|
standard deviation
|
|
SDS/PAGE
|
sodium dodecyl
sulphate/polyacrylamide gel electrophoresis
|
|
Spm
|
spermine
|
|
TEM
|
transmission electron microscopy
|
|
WT
|
wild-type
|
References
|
1
|
La Porta CA: Mechanism of drug sensitivity
and resistance in melanoma. Curr Cancer Drug Targets. 3:391–397.
2009.PubMed/NCBI
|
|
2
|
Setia N, Abbas O, Sousa Y, Garb JL and
Mahalingam M: Profiling of ABC transporters ABCB5, ABCF2 and
nestin-positive stem cells in nevi, in situ and invasive melanoma.
Mod Pathol. 25:1169–1175. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Davies MA: The role of the PI3K-AKT
pathway in melanoma. Cancer J. 18:142–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nikolaous VA, Stratigos AJ, Flaherty KT,
et al: Melanoma: new insights and new therapies. J Invest Dermatol.
132:854–863. 2012. View Article : Google Scholar
|
|
5
|
Goodwin AC, Jadallah S, Toubaji A,
Lecksell K, Hicks JL, Kowaski J, Bova GS, De Marzo AM, Netto GJ and
Casero RA Jr: Increased spermine oxidase expression in human
prostate cancer and prostatic intraepithelial neoplasia tissues.
Prostate. 68:766–772. 2008. View Article : Google Scholar
|
|
6
|
Simoneau AR, Gerner EW, Nagle R, Ziogas A,
Fujikawa-Brooks S, Yerushalmi H, Ahlering TE, Lieberman R, Mclaren
CE, Anton-Culver H and Meyskens FL Jr: The effect of
difluoromethylornithine on decreasing prostate size and polyamines
in men: results of a year-long phase IIb randomized
placebo-controlled chemoprevention trial. Cancer Epidemiol
Biomarkers Prev. 17:292–299. 2008.
|
|
7
|
Gerner EW and Meyskens FL: Polyamines and
cancer: old molecules, new understanding. Nat Rev Cancer.
4:782–792. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Agostinelli E, Belli F, Molinari A,
Condello M, Palmigiani P, Dalla Vedova L, Marra M, Seiler N and
Arancia G: Toxicity of enzymatic oxidation products of spermine to
human melanoma cells (M14): sensitization by heat and MDL 72527.
Biochim Biophis Acta. 1763:1040–1050. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Agostinelli E, Condello M, Molinari A,
Tempera G, Viceconte N and Arancia G: Cytotoxicity of spermine
oxidation products to multidrug resistant melanoma cells (M14
ADR2): sensitisation by MDL 72527, a lysosomotropic compound. Int J
Oncol. 35:485–498. 2009. View Article : Google Scholar
|
|
10
|
Marra M, Agostinelli E, Tempera G,
Lombardi A, Meo G, Budillon A, Abbruzzese A, Giuberti G and
Caraglia M: Anticancer drugs and hyperthermia enhance cytotoxicity
induced by polyamine enzymatic oxidation products. Amino Acids.
33:273–281. 2007. View Article : Google Scholar
|
|
11
|
Marra M, Lombardi A, Agostinelli E,
Giuberti G, Zappavigna S, Tempera G, Vitale G, Bifulco M,
Abbruzzese A and Caraglia M: Bovine serum amine oxidase and spm
potentiate docetaxel and interferon-alpha effects in inducing
apoptosis on human cancer cells through the generation of oxidative
stress. BBA-Mol Cell Res. 1783:2269–2278. 2008.
|
|
12
|
Agostinelli E, Dalla Vedova L, Belli F,
Condello M, Arancia G and Seiler N: Sensitization of human colon
adenocarcinoma cells (LoVo) to reactive oxygen species by
lysosomotropic compounds. Int J Oncol. 29:947–955. 2006.PubMed/NCBI
|
|
13
|
Dai H, Kramer DL, Yang C, Murti KG, Porter
CW and Cleveland JL: The polyamine oxidase inactivator MDL-72527
selectively induces apoptosis in transformed hematopoietic cells
through lysosomotropic effects. Cancer Res. 59:4944–4954. 1999.
|
|
14
|
Iyamu E, Perdew H and Woods G: Growth
inhibitory and differentiation effect of chloroquine and its
analogue on human leukemic cells potentiate fetal hemoglobin
production by targeting the polyamine pathway. Biochem Pharmacol.
77:1021–1029. 2009. View Article : Google Scholar
|
|
15
|
Turini P, Sabatini S, Befani O, Chimenti
F, Casanova C, Riccio P and Mondovi B: Purification of serum amine
oxidase. Anal Biochem. 125:294–298. 1982. View Article : Google Scholar
|
|
16
|
Janes SM, Mu D, Wemmer D, Smith AJ, Kaur
S, Maltby D, Burlingam AL and Klinman JP: A new redox cofactor in
eukaryotic enzymes: 6-Hydroxydopa at the active site of bovine
serum amine oxidase. Science. 248:981–987. 1990. View Article : Google Scholar
|
|
17
|
Molinari A, Toccaceli L, Calcabrini A,
Diociaiuti M, Cianfriglia M and Arancia G: Induction of
P-glycoprotein expression on the plasma membrane of human melanoma
cells. Anticancer Res. 20:2691–2696. 2000.PubMed/NCBI
|
|
18
|
Molinari A, Calcabrini A, Crateri P and
Arancia G: Interaction of anthracyclin with cytoskeletal components
of cultured carcinoma cells (CG5). Exp Mol Pathol. 53:11–33. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van Engeland M, Nieland LJ, Ramaekers FC,
Schutte B and Reutelingsperger CP: Annexin V-affinity assay: a
review on an apoptosis detection system based on phosphatidylserine
exposure. Cytometry. 31:1–9. 1998.PubMed/NCBI
|
|
20
|
Nicoletti I, Migliorati G, Pagliacci MC,
Grignani F and Riccardi C: A rapid and simple method for measuring
thymocyte apoptosis by propidium iodide staining and flow
cytometry. J Immunol Methods. 139:271–279. 1991. View Article : Google Scholar
|
|
21
|
Sharmin S, Sakata K, Kashiwagi K, Ueda S,
Iwasaki S, Shirahata A and Igarashi K: Polyamine cytotoxicity in
the presence of bovine serum amine oxidase. Biochem Biophys Res
Commun. 282:228–235. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Calcabrini A, Arancia G, Marra M, Crateri
P, Befani O, Martone A and Agostinelli E: Enzymatic oxidation
products of spermine induce greater cytotoxic effects on human
multidrug-resistant colon carcinoma cells (LoVo) than on their
wild-type counterparts. Int J Cancer. 99:43–52. 2002. View Article : Google Scholar
|
|
23
|
Agostinelli E and Seiler N:
Non-irradiation-derived reactive oxygen species (ROS) and cancer.
Therapeutic implications. Amino Acids. 31:341–355. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Agostinelli E and Seiler N: Lysosomotropic
compounds and spermine enzymatic oxidation products in cancer
therapy (review). Int J Oncol. 31:473–484. 2007.PubMed/NCBI
|
|
25
|
Agostinelli E, Przybytkowskj E, Mondovì B
and Averill-Bates DA: Heat enhancement of cytotoxicity induced by
oxidation products of spermine in Chinese hamster ovary cells.
Biochem Pharmacol. 72:36–42. 1994.PubMed/NCBI
|
|
26
|
Averill-Bates DA, Ke Q, Tanel A, Roy J,
Fortier G and Agostinelli E: Mechanism of cell death induced by
spermine and amine oxidase in mouse melanoma cells. Int J Oncol.
32:79–88. 2008.PubMed/NCBI
|
|
27
|
Lord-Fontaine S, Agostinelli E,
Przybytkowskj E and Averill-Bates DA: Amine oxidase, spermine, and
hyperthermia induce cytotoxicity in P-glycoprotein overexpressing
multidrug resistant Chinese hamster ovary cells. Biochem Cell Biol.
79:165–175. 2001. View Article : Google Scholar
|
|
28
|
Averill-Bates DA, Cherif A, Agostinelli E,
Tanel A and Fortier G: Anti-tumoral effect of native and
immobilized bovine serum amine oxidase in a mouse melanoma model.
Biochem Pharmacol. 69:1693–1704. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Demers N, Agostinelli E, Averill-Bates DA
and Fortier G: Immobilization of native and poly(ethylene
glycol)-treated (‘PEGylated’) bovine serum amine oxidase into a
biocompatible hydrogel. Biotechnol Appl Biochem. 33:201–207.
2001.PubMed/NCBI
|
|
30
|
Gerner EW and Meyskens Fl Jr: Combination
chemoprevention for colon cancer targeting polyamine synthesis and
inflammation. Clin Cancer Res. 15:758–761. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Park BC, Park SH, Paek SH, et al:
Chloroquine-induced nitric oxide increase and cell death is
dependent on cellular GSH depletion in A172 human glioblastoma
cells. Toxicol Lett. 178:52–60. 2008. View Article : Google Scholar
|
|
32
|
Jiang P, Zhao Y, Shi W, et al: Cell growth
inhibition, G2/M cell cycle arrest, and apoptosis induced by
Chloroquine in human breast cancer cell line Bcap-37. Cell Physiol
Biochem. 22:431–440. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kimura T, Yoshitsugu T, Atsushi T, et al:
Chloroquine in cancer therapy: a double-edged sword of autophagy.
Cancer Res. 73:3–7. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Michaud M, Martins I, Sukkurwala AQ,
Adjemian S, Ma Y, Pellegatti P, et al: Autophagy-dependent
anticancer immuneresponses induced by chemotherapeutic agents in
mice. Science. 334:1573–1577. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Amaravadi RK and Thompson CB: The roles of
therapy-induced autophagy and necrosis in cancer treatment. Clin
Cancer Res. 13:7271–7279. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tasdemir E, Galluzzi L, Maiuri MC, Criollo
A, Vitale I, Hangen E, Modjtahedi N and Kroemer G: Methods for
assessing autophagy and autophagic cell death. Methods Mol Biol.
445:29–76. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vezmar M and Georges E: Reversal of
MRP-mediated doxorubicin resistance with quinoline-based drugs.
Biochem Pharmacol. 59:1245–1252. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Solomon VR and Lee H: Chloroquine and its
analogs: a new promise of an old drug for effective and safe cancer
therapies. Eur J Pharmacol. 625:220–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Homewood CA, Warhurst DC, Peters W and
Baggaley VC: Lysosomes, pH and the anti-malarial action of
chloroquine. Nature. 235:50–52. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Slater AF: Chloroquine: mechanism of drug
action and resistance in Plasmodium falciparum. Pharmacol Ther.
57:203–235. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Agostinelli E, Tempera G, Dalla Vedova L,
Condello M and Arancia G: MDL 72527 and spermine oxidation products
induce a lysosomotropic effect and mitochondrial alterations on
tumor cells. Biochem Soc Trans. 35:343–348. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guicciardi ME, Leist M and Gores GJ:
Lysosomes in cell death. Oncogene. 23:2881–2890. 2004. View Article : Google Scholar
|
|
43
|
Zhao M, Antunes F, Eaton JW and Brunk UT:
Lysosomal enzymes promote mitochondrial oxidant production,
cytochrome c release and apoptosis. Eur J Biochem. 270:13778–13786.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu Z, Li W and Brunk UT: 3-Aminopropanal
is a lysosomotropic aldehyde that causes oxidative stress and
apoptosis by rupturing lysosomes. APMIS. 111:643–652. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zdolsek J, Zhang H, Roberg K and Brunk U:
HO-mediated damage to lysosomal membranes of J-774 cells. Free
Radic Res Commun. 18:71–85. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gottesman MM and Pastan I: Biochemistry of
multidrug resistance mediated by the multidrug transporter. Ann Rev
Biochem. 62:385–342. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Arancia G, Calcabrini A, Marra M, Crateri
P, Artico M, Martone A, Martelli F and Agostinelli E: Mitochondrial
alterations induced by serum amine oxidase and spermine on human
multidrug resistant tumor cells. Amino Acids. 26:273–282. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fehrenbacher N, Gyrd-Hansen M, Poulsen B,
Felbor U, Kallunki T, Boes M, Weber E, Leist M and Jäättelä M:
Sensitization to the lysosomal cell death pathway upon
immortalization and transformation. Cancer Res. 64:5301–5310. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fehrenbacher N and Jäättelä M: Lysosomes
as targets for cancer therapy. Cancer Res. 65:2993–2995.
2005.PubMed/NCBI
|
|
50
|
Savarino A, Lucia MB, Giordano F and Cauda
R: Risks and benefits of chloroquine use in anticancer strategies.
Lancet Oncol. 7:792–793. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Briceño E, Reyes S and Sotelo J: Therapy
of glioblastoma multiforme improved by the antimutagenic
chloroquine. Neurosurg Focus. 14:e32003.PubMed/NCBI
|