Introduction
Gastric cancer is one of the most fatal cancers,
especially in eastern Asia. In recent years, despite the progress
made in diagnosis and treatment of gastric cancer, the mortality
rate of gastric cancer remains high, mainly due to complex
biological characteristics of gastric cancer and its high rate of
malignancy (1). Most patients are
diagnosed at an advanced stage and therefore it remains one of the
most life-threatening pathological events involving the spread of
tumor cells from a primary tumor to other organs. Currently, some
studies have described the metastasis of gastric cancer, but its
exact mechanism remains unknown. Similar to tumorigenesis, tumor
metastasis also includes multiple factors and genes. Metastasis
consists of five distinct steps: i) detachment of tumor cells from
the primary tumor; ii) invasion into surrounding tissue; iii)
intravasation into blood or lymphatic vessels; iv) dissemination in
the blood stream or the lymphatic system; and, v) extravasation and
outgrowth at secondary sites. To detach from the primary tumor and
to invade into the surrounding tissue, tumor cells need to
dismantle cell-cell junction, remodel cell-matrix contact sites,
and follow a chemo-attractive path through the extracellular
matrix, mined by secreted proteinases. These processes are commonly
observed in various normal conditions, such as in developmental
processes i.e. gastrulation or neural crest cell migration, where
differentiated, epithelial cells dedifferentiate, move to a distant
site, and then re-differentiate to form a new structure. This
temporary and reversible phenomenon is known as the
epithelial-mesenchymal transition (EMT), a process that is
currently in the limelight of investigating the onset of cancer
cell migration, invasion and metastatic dissemination (2,3).
Recent studies have shown that abnormal EMT is closely related to
tumorigenesis and progression of epithelial cancers, such as lung
cancer, colon cancer and breast cancer (4,5).
The association of thrombin and cancer is documented
(6). But, the role of thrombin in
propagation of the pro-malignant phenotype has not been fully
elucidated. This pro-malignant role of thrombin has been attributed
to its stimulation of tumor adhesion (7–10),
growth (11), metastasis (12–14),
and angiogenesis (15,16). Thrombin, a multifunctional serine
protease, activates platelets and regulates the behavior of other
cells through G-protein coupled protease-activated receptors (PARs)
(17,18). Protease-activated receptor-1
(PAR1), the prototype of this family, is activated when thrombin
cleaves the amino-terminal extracellular domain at a specific site
(19). PAR1 is overexpressed in
invasive and metastatic tumors and the expression levels directly
correlate with the degree of invasiveness of the cancer. PAR1 has
been found to be instrumental in cell growth and invasion of
tumor-derived cells (20,21).
We previously reported that the expression of PAR1,
along with a metalloproteinase known to activate PAR1 (MMP1) were
associated with poorer prognoses, compared with expression-negative
tumors, and activated PAR1 promotes gastric cancer cell invasion
in vivo as well as cell morphological changes (22–24).
These results underline the relevance of EMT and PAR1 activity to
gastric cancer, however there are few reports on the role of PAR1
in EMT. Therefore, this study investigated the role of PAR1
activity in EMT by observing the expression levels of epithelial
and mesenchymal markers, such as E-cadherin and vimentin. This
study on the relationship between PAR1 activity and EMT in gastric
cancer further elucidates metastasis which in turn could serve to
impact gastric cancer prognoses and potentially add knowledge for
targeted therapy.
Materials and methods
Reagents
An antibody against PAR1 (clone WEDE15) was
purchased from Beckman Coulter (Fuller ton, CA, USA).
Anti-E-cadherin, fibronectin, vimentin, β-catenin, Twist, E12/E47
and Snail were purchased from Abcam (Cambridge, UK). Anti-GAPDH was
from Imgenex (San Diego, CA, USA). Human α-thrombin was purchased
from Sigma-Aldrich (catalog no. T1063) (St. Louis, MO, USA). The
selective PAR1 antagonist SCH79797 (catalog no. 1592) (IC50=70 nM)
was purchased from Tocris Bioscience (Avonmouth, UK) (25).
Cell culture
The human gastric cancer cell lines, MKN45 and MKN74
cells, were obtained from the Riken Cell Bank (Tsukuba, Japan).
Cells were cultured at 37°C in 5% CO2 in RPMI-1640
medium containing 10% fetal bovine serum (FBS). Cells were
propagated by mechanical re-suspension using a scraper, without the
use of trypsin. PAR1 is expressed in MKN74, not in MKN45 (23).
Western blot assay
Whole, cytoplasmic and nuclear cell protein was
extracted using RIPA-buffer (Wako, Osaka, Japan) containing 1%
protease inhibitor cocktail (Sigma-Aldrich) and NE-PER (Thermo
Scientific, Rockford, IL, USA). Protein concentration was measured
with a protein assay reagent (Bio-Rad, Hercules, CA, USA). Proteins
were resolved by SDS-PAGE using a 5–20% SuperSep gel (Wako) and
analyzed by western blotting using polyvinylidene difluoride
membranes (Millipore, Bedford, MA, USA) according to the
manufacturer’s instructions. Membranes were blocked with 3% non-fat
dry milk in TBS plus 0.1% Tween-20 (TBST). The membranes were
probed with anti-E-cadherin antibody at 1:1,000 dilution,
anti-fibronectin and anti-vimentin antibody at 1:750 dilution,
anti-β-catenin antibody at 1:2,000 in TBST. Protein bands were
incubated with primary antibody overnight at 4°C. An enhanced
chemiluminescence detection system (ImmunoStar; Wako) was used for
visualized of immunoreactive bands after the reaction with the
HRP-labeled secondary antibody against mouse or rabbit IgG. Equal
loading of samples was confirmed by probing the membranes with
GAPDH antibody.
Measurement of Snail activation by
electrophoretic mobility shift assay
MKN45/mock, MKN45/PAR1 and MKN74 were treated for 24
h with 15 nM α-thrombin or 15 nM α-thrombin and 70 nM SCH79797.
Nuclear fractions were extracted from the cultured cells using
NE-PER (Thermo Scientific).
In the electrophoretic mobility shift assay (EMSA),
the double-stranded oligonucleotide probes were labeled with
biotin. Non-labeled oligonucleotides were used as competitors for
biotin-labeled oligonucleotide binding. The sequences flanking the
E-boxes are for E-box 1, 5′-CTGAGATTAC ATGCGTGAGTCACTA-3′;
for E-box 2, 5′-TGTTGTGTG CATGTGTTATGGAACC-3′; and for E-box
3, 5′-ATTACA ACACGAGCCCACCATGCCT-3′. Briefly, 20 μl of
binding reaction containing 2 μg of nuclear extract was prepared
according to the kit instruction manual and incubated at room
temperature for 20 min. Complexes were resolved on 5–20% SuperSep
gel (Wako) in 0.5X TBE buffer at 4°C. When electrophoresis was
completed, binding reactions were electrophoretically transferred
to N+ nylon membrane. The transferred DNA then was cross-linked to
the N+ nylon membrane, and the biotin-labeled DNA-protein complex
was detected by streptavidin-HRP using a LightShift
Chemiluminescent EMSA kit (Thermo Scientific).
Immunofluorescence
Cultured cells were fixed with 4% paraformaldehyde
at room temperature, permeabilized with 0.1% Triton X-100 in PBS
and blocked with 3% FBS in PBS. Following overnight incubation at
4°C with primary antibodies, and incubation in the dark with Alexa
405 and 488 Fluor dye-labeled secondary antibodies,
immunofluorescence was detected using a Leica DMLB confocal laser
fluorescence microscope (Leica Microsystems, Mannheim,
Germany).
Results
PAR1 activation via α-thrombin induces
epithelial marker downregulation and mesenchymal marker
upregulation
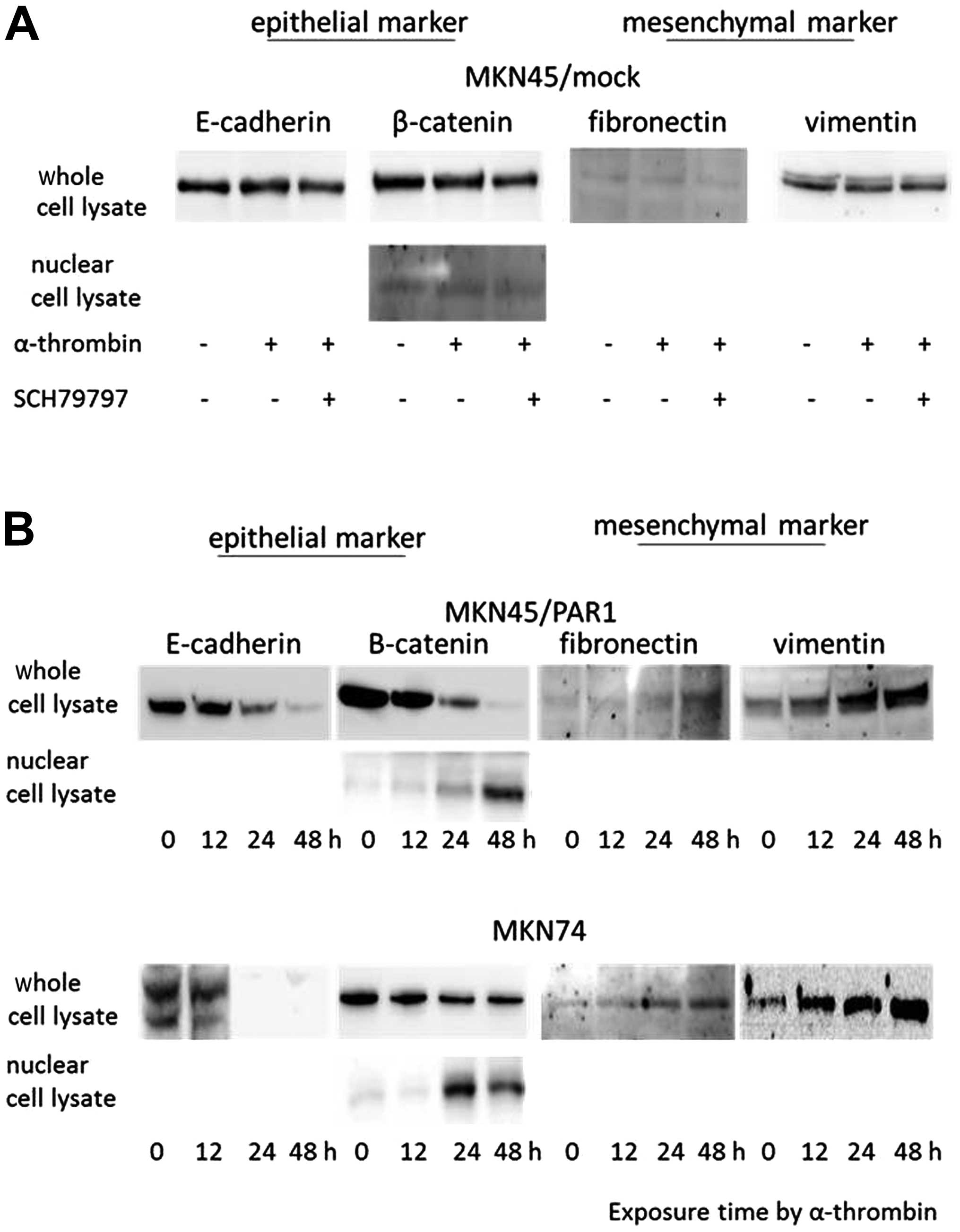
Given the shape change caused by α-thrombin
activation of PAR1 (24), we
hypothesized that EMT would have been induced in gastric cancer
cells. To test this hypothesis, we collected cell lysate from
MKN45/mock, MKN45/PAR1 and MKN74 treated with 15 nM α-thrombin or
15 nM α-thrombin and 70 nM SCH79797. When MKN45/mock was treated
with α-thrombin or α-thrombin plus SCH7979 for 24 h, we showed that
the epithelial and mesenchymal markers were stably expressed at a
level that did not change over time (Fig. 1A). MKN45/PAR1 and MKN74 treated
with α-thrombin presented reduced expression levels of the
epithelial markers E-cadherin and β-catenin in cytoplasmic lysate
and an icreased expression level of β-catenin in nuclear lysate. On
the other hand, the expression levels of fibronectin and vimentin,
which are mesenchymal markers, were increased in cytoplasmic lysate
(Fig. 1B). MKN45/PAR1 and MKN74
treated with α-thrombin along with SCH79797 presented expression
levels of fibronectin and vimentin similar to that of MKN45/mock
(Fig. 1C).
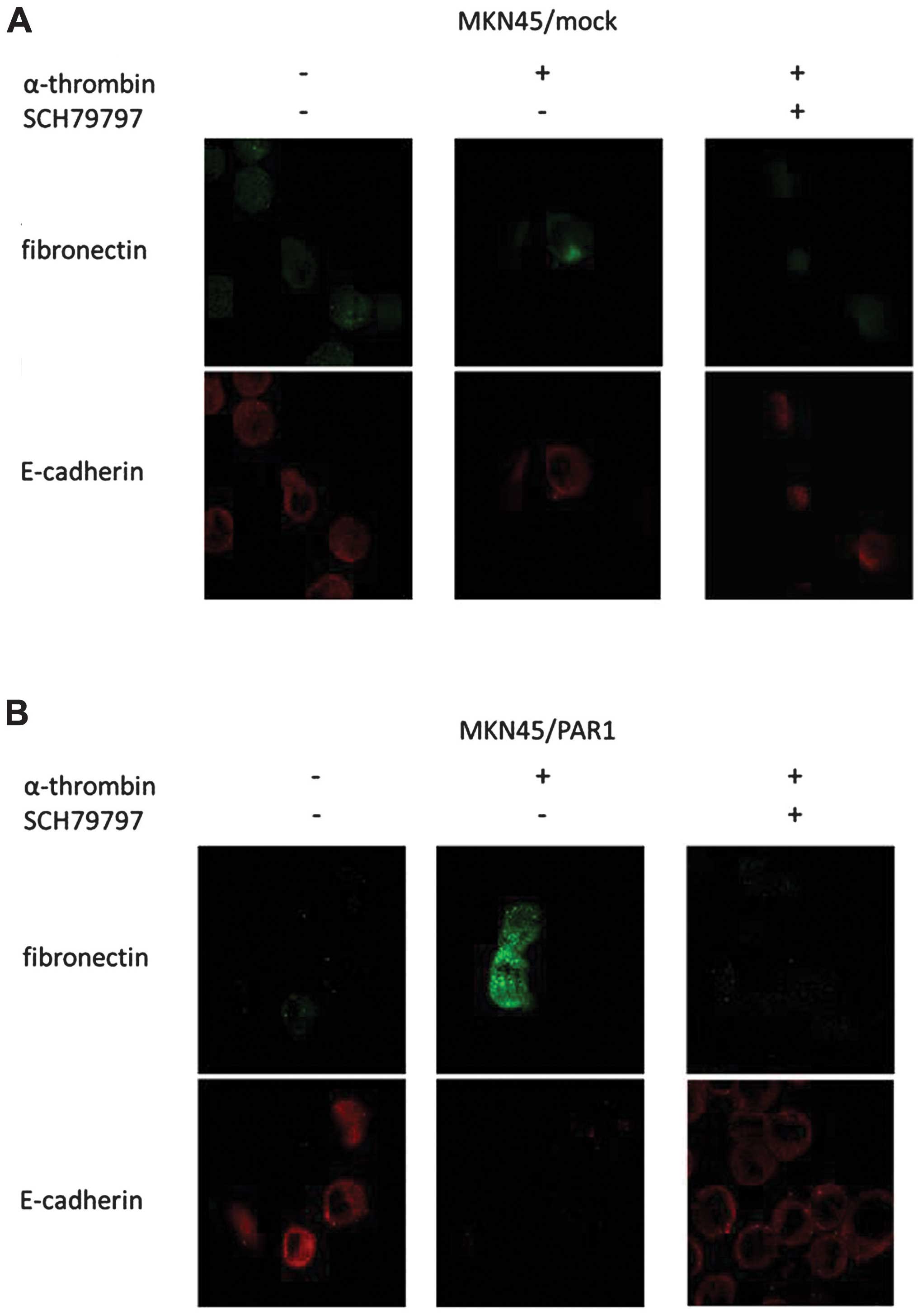
Immunofluorescence of E-cadherin and
fibronectin expression
Our investigation of the role of thrombin in the
morphology of MKN45/PAR1 and MKN74 cells employed
immunofluorescence staining to visualize the expression levels of
E-cadherin and fibronectin in untreated MKN45/mock, MKN45/PAR1 and
MKN74 cells and these cells were treated with 15 nM α-thrombin in
the absence and presence of 70 nM SCH79797.
MKN45/PAR1 and MKN74 cells underwent a morphological
change from round- or epithelial-like to a spindle-like shape
phenotype upon treatment with α-thrombin and this morphological
change was not observed in response to treatment with a combination
of α-thrombin and SCH79797 (Fig.
2). The expression of E-cadherin decreased and the expression
of fibronectin increased in response to treatment of MKN45/PAR1 and
MKN74 cells with α-thrombin while no such change in expression
levels were observed in these cells that were either untreated or
underwent α-thrombin treatment in the presence of SCH79797
(Fig. 2).
Snail, transcriptional factor, travels
into the nucleus
Next, we investigated the impact of α-thrombin
induced PAR1 activation upon E-cadherin expression. Several
E-cadherin transcriptional repressors have been characterized
(Snail, E12/E47, Twist, and SIP-1) and shown to interact with
proximal E-boxes of the E-cadherin promoter.
To test whether or not PAR1 agonist-mediated
elevation is associated with an increase in binding to the
E-cadherin promoter, we examined the binding of these
transcriptional repressors to E-boxes by means of EMSA and western
blotting. Assessment was carried out of E-cadherin promoter E-box
binding by nuclear proteins from MKN45/mock and MKN74 as control,
MKN45/PAR1- and MKN74-treated α-thrombin, and MKN45/PAR1 and MKN74
treated with α-thrombin and SCH79797. We designed oligonucleotide
probes comprising E-box binding sequences as described previously
(26). No shifted band was evident
when the labeled oligonucleotide was exposed to nuclear lysates
derived from MKN45/mock and MKN74 cells (Fig. 3A). In contrast, labeled
oligonucleotide incubated with nuclear lysate from MKN45/PAR1 and
MKN74 cells with α-thrombin clearly shows a retarded
electrophoretic mobility band (Fig.
3B). Slightly leaky expression of a shifted band was observed
when the labelled oligonucleotide was incubated with nuclear lysate
from MKN45/PAR1 and MKN74 cells treated with a combination of
α-thrombin and SCH79797 (Fig.
3C).
Subsequently, we employed western blotting to
examine nuclear expression of Twist, Snail and E12/E47. Twist and
E12/E47 were not detected in nuclear lysates of MKN45/PAR1 and
MKN74 that underwent α-thrombin induced PAR1 activation (Fig. 4). Snail was present in the nuclear
lysates derived from MKN45/PAR1 and MKN74 that underwent α-thrombin
induced PAR1 activation while Snail was not detected in nuclear
lysates of these cells in which α-thrombin-induced PAR1 activation
was suppressed by SCH79797 (Fig.
4).
Discussion
In the current study, we demonstrate that PAR1
activity induces pro-migratory effects in gastric cancer cell lines
by inducing EMT. Malignant tumors including gastric cancer are
multi-step processes associated with changes in gene expression
involved in cell proliferation, invasion, and metastasis. The most
troubling metastasis in gastric cancer is peritoneal dissemination,
for which anticancer therapies have been developed. However,
despite the proven benefits of these treatments (27,28),
patients with peritoneal dissemination of gastric cancer still have
poor prognosis and outcomes of gastric cancer require further
improvement.
Many metastasis-related proteins have been reported
in various cancers including gastric cancer. PAR1, G-protein
coupled receptor, was identified as one of the most upregulated
proteins in advanced gastric cancer by immunohistochemistory
(23). PAR1, also known as the
coagulation factor II (thrombin) receptor, is a protein that in
humans is encoding the F2R gene (29). Thrombin activates PAR1 in alveolar
epithelial cells and this pathway is associated with epithelial to
myofibroblats transition and collagen secretion, and
thrombin-induced EMT is mediated through PAR1 (30). Additionally, PAR1-mediated
extracellular signal-regulated kinase activation and promoted the
transition of tubular epithelial cells into cells of mesenchymal
phenotype.
Recent studies have revealed that PAR1 activity
encouraged cancer invasion in various cancers (23,31,32).
EMT, characterized by a gain of mesenchymal cell markers
(fibronectin, vimentin, smooth muscle actin, and N-cadherin) and a
loss of epithelial markers (E-cadherin and β-catenins) (33,34),
is a process whereby cells undergo molecular alterations that
facilitate cell motility and invasion (35). In this study we shown that thrombin
via PAR1 induced EMT in gastric cancer MKN45/PAR1 and MKN74 cell
lines and this was associated with time-dependent downregulation of
E-cadherin accompanied by an upregulation of vimentin and
β-catenin. Thrombin also induced a morphological alteration in
MKN45/PAR1 and MKN74 cells to a spindle-shaped phenotype.
Additionally, we show that expression of E-cadherin, vimentin and
β-catenin remained unchanged in MKN45/PAR1 cells exposed to
SCH79797, which is a specific PAR1 antagonist. The most common
biochemical change associated with EMT is the loss of E-cadherin
expression. These data imply that PAR1 activation by thrombin
induces EMT in gastric cancer and may be conducive to invasion and
metastasis.
E-cadherin transcriptional repressors such as Snail,
Slug, E12/E47, and Twist have traditionally been implicated in
promoting EMT in various systems of embryonic development and tumor
progression (36,37). The transcription factor Twist is a
component of embryonic morphogenesis (38). Snail, a Zinc-finger transcription
factor, has been isolated in Drosophila embryos (39). During the embryonic development,
Snail has been implicated in the triggering of EMT in the precursor
cells of the mesoderm and neural crest (40). Snail is known to directly repress
E-cadherin gene transcription by binding to the E-box on the
E-cadherin promoter (41). Several
reports have also implicated that Snail not only in E-cadherin
repression but also in the acceleration of cancer invasion in
various carcinomas (41,42). Snail expression correlated with
histological grade and lymph node status in breast carcinomas
(43), and Snail mRNA expression
levels independently correlated with capsular invasion in
hepatocellular carcinoma (44). We
showed by means of western blotting and EMSA that the transcription
factor Snail moved from cell cytoplasm to the nucleus. These data
indicated that PAR1 activation via thrombin induced Snail-mediated
E-cadherin transcriptional repressor and ultimately EMT in gastric
cancer cells.
We conclude that at least one of the facets of PAR1
activity by thrombin induced upregulation of malignant phenotype is
via the upregulation of Snail in gastric cancer cells. SCH79797,
which is a PAR1 antagonist, blocked these effects of thrombin.
Thrombin, MMP1 and urokinase-type plasminogen activator, which are
PAR1 agonist, contribute to tumor invasion and tumorigenesis
(45,46). These results suggest that
inhibition of PAR1 signaling pathway may be a new strategy to
prevent tumor invasion and metastasis.
References
|
1
|
Memon MA, Subramanya MS, Khan S, Hossain
MB, Osland E and Memon B: Meta-analysis of D1 versus D2 gastrectomy
for gastric adenocarcinoma. Ann Surg. 253:900–911. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grünert S, Jechlinger M and Beug H:
Diverse cellular and molecular mechanisms contribute to epithelial
plasticity and metastasis. Nat Rev Mol Cell Biol. 4:657–665.
2003.PubMed/NCBI
|
|
4
|
Jain VK and Cunningham D: Targeting
angiogenesis in advanced gastric cancer: Is this end of the road?
Transl Gastrointest Cancer. 1:119–121. 2012.
|
|
5
|
Rose CS and Malcolm S: A TWIST in
development. Trends Genet. 13:384–387. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nierodzik ML and Karpatkin S: Thrombin
induces tumor growth, metastasis, and angiogenesis: evidence for a
thrombin-regulated dormant tumor phenotype. Cancer Cell.
10:355–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nierodzik ML, Plotkin A, Kajumo F and
Karpatkin S: Thrombin stimulates tumor-platelet adhesion in vitro
and metastasis in vivo. J Clin Invest. 87:229–236. 1991. View Article : Google Scholar
|
|
8
|
Nierodzik ML, Kajumo F and Karpatkin S:
Effect of thrombin treatment of tumor cells on adhesion of tumor
cells to platelets in vitro and tumor metastasis in vivo. Cancer
Res. 52:3267–3272. 1992.PubMed/NCBI
|
|
9
|
Nierodzik ML, Bain RM, Liu LX, Shivji M,
Takeshita K and Karpatkin S: Presence of the seven transmembrane
thrombin receptor on human tumour cells: effect of activation on
tumour adhesion to platelets and tumor tyrosine phosphorylation. Br
J Haematol. 92:452–457. 1996. View Article : Google Scholar
|
|
10
|
Klepfish A, Greco MA and Karpatkin S:
Thrombin stimulates melanoma tumor-cell binding to endothelial
cells and subendothelial matrix. Int J Cancer. 53:978–982. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zain J, Huang YQ, Feng X, Nierodzik ML, Li
JJ and Karpatkin S: Concentration dependent dual effect of thrombin
on impaired growth/apoptosis or mitogenesis in tumor cells. Blood.
95:3133–3138. 2000.PubMed/NCBI
|
|
12
|
Nierodzik ML, Chen K, Takeshita K, et al:
Protease-activated receptor 1 (PAR-1) is required and rate-limiting
for thrombin-enhanced experimental pulmonary metastasis. Blood.
92:3694–3700. 1998.PubMed/NCBI
|
|
13
|
Hu L, Lee M, Campbell W, Perez-Soler R and
Karpatkin S: Role of endogenous thrombin in tumor implantation,
seeding and spontaneous metastasis. Blood. 104:2746–2751. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wojtukiewicz MZ, Tang DG, Ciarelli JJ, et
al: Thrombin increases the metastatic potential of tumor cells. Int
J Cancer. 54:793–806. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang YQ, Li JJ, Hu L, Lee M and Karpatkin
S: Thrombin induces increased expression and secretion of VEGF from
human FS4 fibroblasts, DU145 prostate cells and CHRF
megakaryocytes. Thromb Haemost. 86:1094–1098. 2001.PubMed/NCBI
|
|
16
|
Caunt M, Huang YQ, Brooks PC and Karpatkin
S: Thrombin induces neoangiogenesis in the chick chorioallantoic
membrane. J Thromb Haemost. 1:2097–2102. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Coughlin SR: How the protease thrombin
talks to cells. Proc Natl Acad Sci USA. 96:11023–11027. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vu TK, Hung DT, Wheaton VI and Coughlin
SR: Molecular cloning of a functional thrombin receptor reveals a
novel proteolytic mechanism of receptor activation. Cell.
64:1057–1068. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rasmussen UB, Vouret-Craviari V, Jallat S,
et al: cDNA cloning and expression of a hamster alpha-thrombin
receptor coupled to Ca2+ mobilization. FEBS Lett.
288:123–128. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Even-Ram S, Uziely B, Cohen P, et al:
Thrombin receptor overexpression in malignant and physiological
invasion processes. Nat Med. 4:909–914. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Even-Ram SC, Maoz M, Pokroy E, et al:
Tumor cell invasion is promoted by activation of protease activated
receptor-1 in cooperation with the alpha vbeta 5 integrin. J Biol
Chem. 276:10952–10962. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujimoto D, Hirono Y, Goi T, Katayama K
and Yamaguchi A: Prognostic value of protease-activated receptor-1
(PAR-1) and matrix metalloproteinase-1 (MMP-1) in gastric cancer.
Anticancer Res. 28:847–854. 2008.PubMed/NCBI
|
|
23
|
Fujimoto D, Hirono Y, Goi T, Katayama K,
Matsukawa S and Yamaguchi A: The activation of proteinase-activated
receptor-1 (PAR1) mediates gastric cancer cell proliferation and
invasion. BMC Cancer. 10:4432010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fujimoto D, Hirono Y, Goi T, Katayama K,
Matsukawa S and Yamaguchi A: The activation of proteinase-activated
receptor-1 (PAR1) promotes gastric cancer cell alteration of
cellular morphology related to cell motility and invasion. Int J
Oncol. 42:565–573. 2013.PubMed/NCBI
|
|
25
|
Ahn HS, Foster C, Boykow G, Stamford A,
Manna M and Graziano M: Inhibition of cellular action of thrombin
by N3-cyclopropyl-7-[[4-(1-methylethyl)phenyl]methyl]-7H-pyrrolo[3,
2-f ]quinazoline-1,3-diamine (SCH 79797), a nonpeptide thrombin
receptor antagonist. Biochem Pharmacol. 60:1425–1434.
2000.PubMed/NCBI
|
|
26
|
Manohar CF, Bray JA, Salwen HR, et al:
MYCN-mediated regulation of the MRP1 promoter in human
neuroblastoma. Oncogene. 23:753–762. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fushida S, Kinoshita J, Yagi Y, et al:
Dual anti-cancer effects of weekly intraperitoneal docetaxel in
treatment of advanced gastric cancer patients with peritoneal
carcinomatosis: A feasibility and pharmacokinetic study. Oncol Rep.
19:1305–1310. 2008.
|
|
28
|
Yonemura Y, Elnemr A, Endou Y, et al:
Multidisciplinary therapy for treatment of patients with peritoneal
carcinomatosis from gastric cancer. World J Gastrointest Oncol.
2:85–97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Coughlin SR, Vu TK, Hung DT and Wheaton
VI: Characterization of a functional thrombin receptor. Issues and
opportunities. J Clin Invest. 89:351–355. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song JS, Kang CM, Park CK and Yoon HK:
Thrombin induces epithelial-mesenchymal transition via PAR-1, PKC,
and ERK1/2 pathways in A549 cells. Exp Lung Res. 39:336–348. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Juncker-Jensen A, Deryugina EI, Rimann I,
et al: Tumor MMP-1 activates endothelial PAR1 to facilitate
vascular intravasation and metastatic dissemination. Cancer Res.
73:4196–4211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ohshiro K, Bui-Nguyen TM, Divijendra Natha
RS, Schwartz AM, Levine P and Kumar R: Thrombin stimulation of
inflammatory breast cancer cells leads to aggressiveness via the
EGFR-PAR1-Pak1 pathway. Int J Biol Markers. 27:e305–e313. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Radisky DC: Epithelial-mesenchymal
transition. J Cell Sci. 118:4325–4326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thiery JP: Epithelial-mesenchymal
transitions in development and pathologies. Curr Opin Cell Biol.
15:740–746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang J, Mani SA and Weinberg RA: Exploring
a new twist on tumor metastasis. Cancer Res. 66:4549–4552. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Peinado H, Portillo F and Cano A:
Transcriptional regulation of cadherins during development and
carcinogenesis. Int J Dev Biol. 48:365–375. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang J, Mani SA, Donaher JL, et al: Twist,
a master regulator of morphogenesis, plays an essential role in
tumor metastasis. Cell. 117:927–939. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Thisse B, el Messal M and Perrin-Schmitt
F: The twist gene: isolation of a Drosophila zygotic gene
necessary for the establishment of dorsoventral pattern. Nucleic
Acids Res. 15:3439–3453. 1987.PubMed/NCBI
|
|
39
|
Grau Y, Carteret C and Simpson P:
Mutations and chromosomal rearrangements affecting the expression
of snail, a gene involved in embryonic patterning in Drosophila
melanogaster. Genetics. 108:347–360. 1984.PubMed/NCBI
|
|
40
|
Cano A, Pérez-Moreno MA, Rodrigo I, et al:
The transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Batlle E, Sancho E, Franci C, et al: The
transcription factor snail is a repressor of E-cadherin gene
expression in epithelial tumour cells. Nat Cell Biol. 2:84–89.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yokoyama K, Kamata N, Hayashi E, et al:
Reverse correlation of E-cadherin and snail expression in oral
squamous cell carcinoma cells in vitro. Oral Oncol. 37:65–71. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Blanco MJ, Moreno-Bueno G, Sarrio D, et
al: Correlation of Snail expression with histological grade and
lymph node status in breast carcinomas. Oncogene. 21:3241–3246.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sugimachi K, Tanaka S, Kameyama T, et al:
Transcription factor snail and progression of human hepatocellular
carcinoma. Clin Cancer Res. 9:2657–2664. 2003.PubMed/NCBI
|
|
45
|
Boire A, Covic L, Agarwal A, Jacques S,
Sherifi S and Kuliopulos A: PAR1 is a matrix metalloprotease-1
receptor that promotes invasion and tumorigenesis of breast cancer
cells. Cell. 120:303–313. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rømer J, Bugge TH, Pyke C, et al:
Plasminogen and wound healing. Nat Med. 2:7251996.
|